
Rise of the BQ.1.1.37 SARS-CoV-2 Sublineage, Italy
 , , ,
, , , 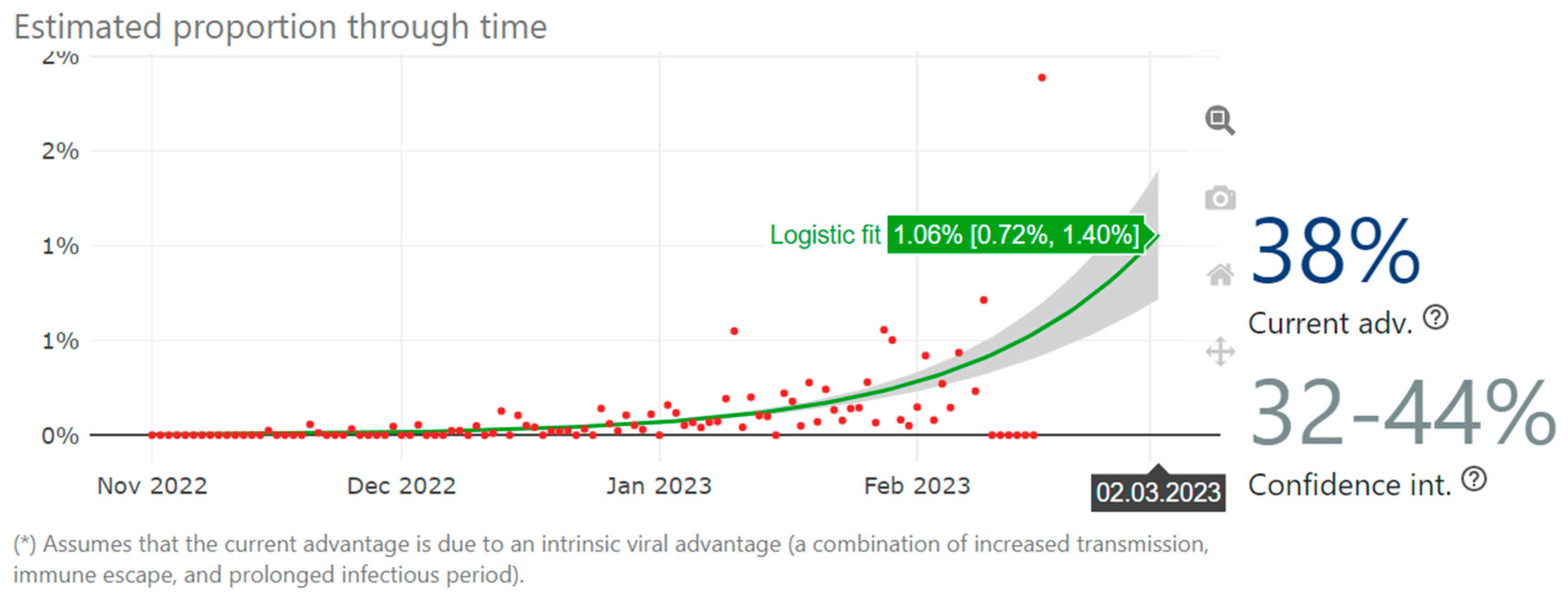{kind=link}
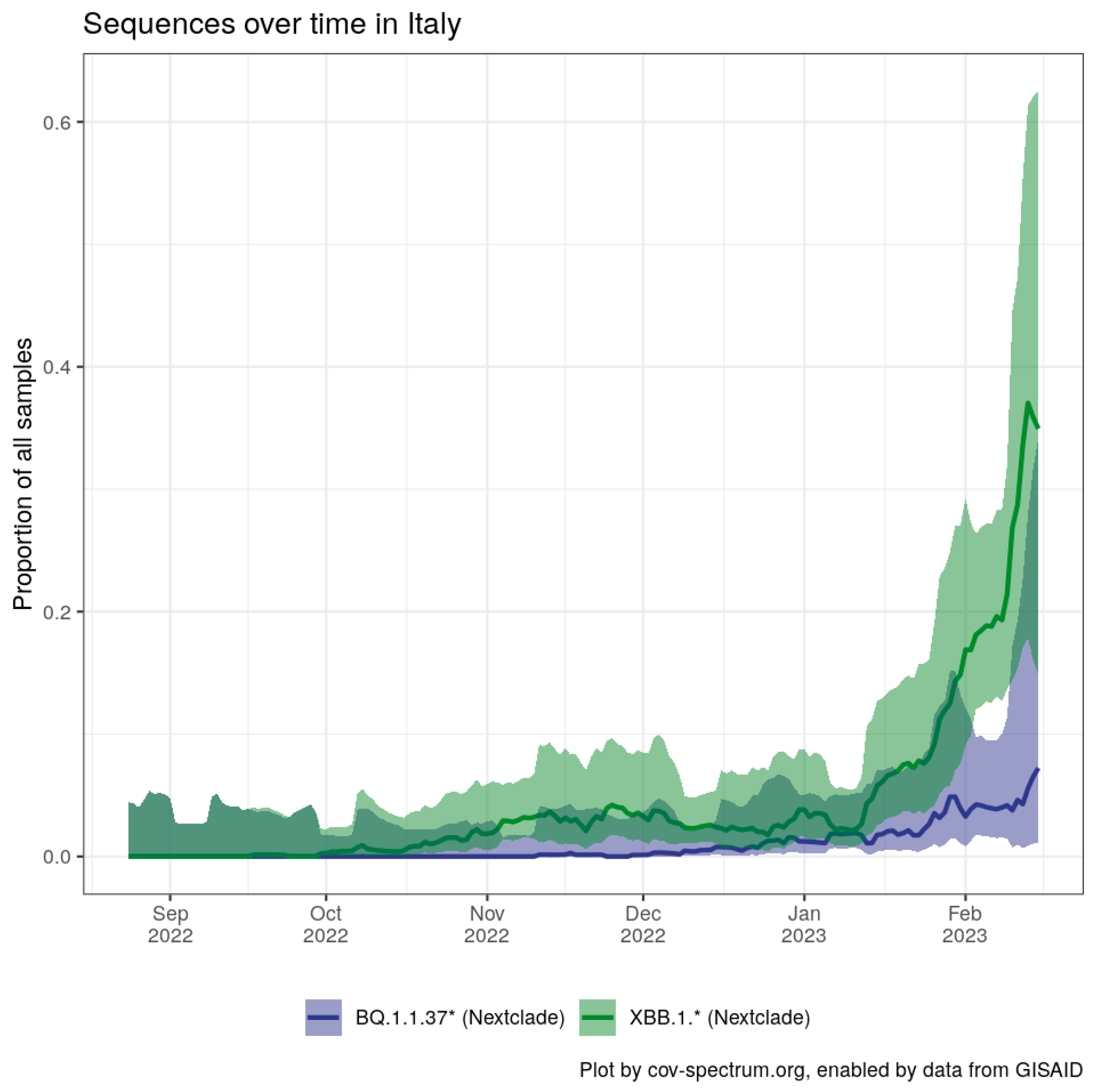{kind=link}
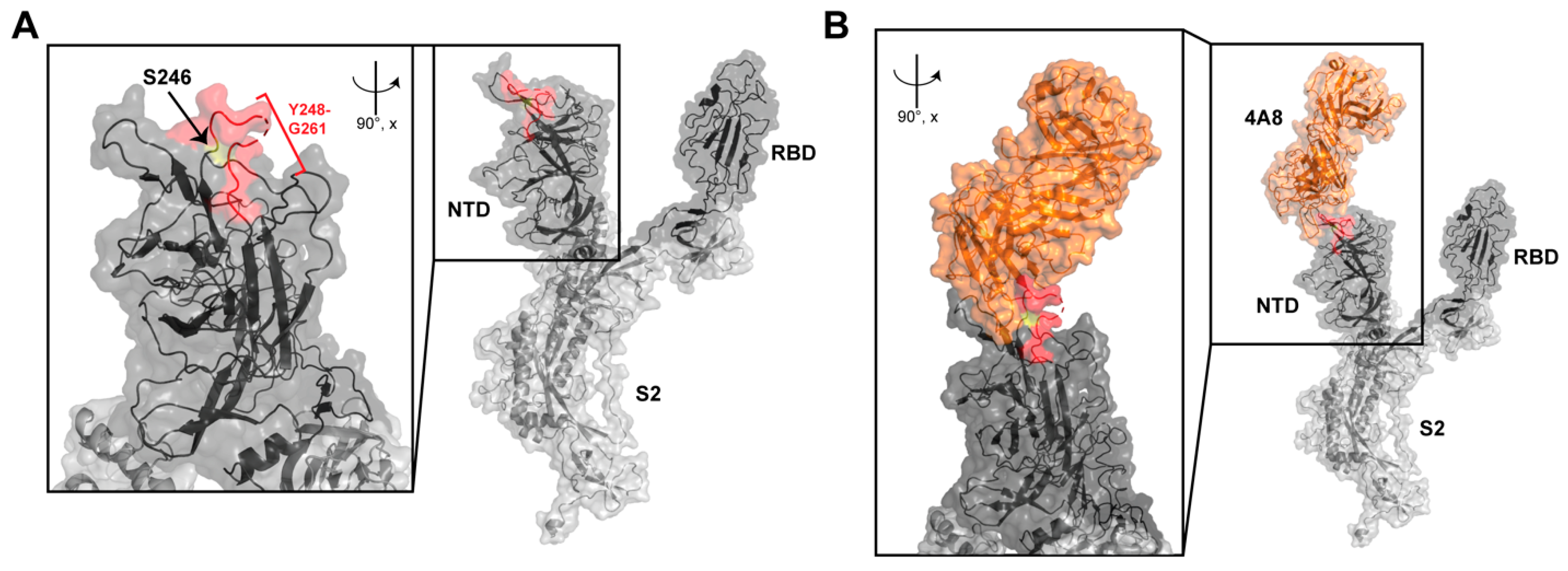{kind=link}
Abstract
:


Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- PANGO Lineage Notes. Available online: https://github.com/cov-lineages/pango-designation/blob/master/lineage_notes.txt (accessed on 25 January 2023).
- BQ.1.1 + T1453C Sublineage with Insertion at S:247/248. Available online: https://github.com/cov-lineages/pango-designation/issues/1571 (accessed on 24 January 2023).
- Elbe, S.; Buckland-Merrett, G. Data, disease and diplomacy: GISAID’s innovative contribution to global health. Glob. Chall. 2017, 1, 33–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.J.; Lapedes, A.S.; de Jong, J.C.; Bestebroer, T.M.; Rimmelzwaan, G.F.; Osterhaus, A.D.; Fouchier, R.A. Mapping the antigenic and genetic evolution of influenza virus. Science 2004, 305, 371–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Nadeau, S.; Yared, M.; Voinov, P.; Xie, N.; Roemer, C.; Stadler, T. CoV-Spectrum: Analysis of globally shared SARS-CoV-2 data to identify and characterize new variants. Bioinformatics 2021, 38, 1735–1737. [Google Scholar] [CrossRef] [PubMed]
- Collection #24 Tracking/Designated Lineages by Federico Gueli, Nick Frohnberg, and Andrew Urquhart. Available online: https://cov-spectrum.org/collections/24 (accessed on 26 January 2023).
- Finkelstein, M.T.; Mermelstein, A.G.; Parker Miller, E.; Seth, P.C.; Stancofski, E.D.; Fera, D. Structural Analysis of Neutralizing Epitopes of the SARS-CoV-2 Spike to Guide Therapy and Vaccine Design Strategies. Viruses 2021, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Andreano, E.; Piccini, G.; Licastro, D.; Casalino, L.; Johnson, N.V.; Paciello, I.; Dal Monego, S.; Pantano, E.; Manganaro, N.; Manenti, A.; et al. SARS-CoV-2 escape in vitro from a highly neutralizing COVID-19 convalescent plasma. Proc. Natl. Acad. Sci. USA 2020, 118, e2103154118. [Google Scholar] [CrossRef] [PubMed]
- Garry, R.; Gallaher, W.R. Naturally Occurring Indels in Multiple Coronavirus Spikes. 2020. Available online: https://virological.org/t/naturally-occurring-indels-in-multiple-coronavirus-spikes/560 (accessed on 26 January 2023).
- Garry, R.; Andersen, K.; Gallaher, W.; Tsan-Yuk Lam, T.; Gangaparapu, K.; Latif, A.; Beddingfield, B.; Rambaut, A.; Holmes, E. Spike Protein Mutations in Novel SARS-CoV-2 ‘Variants of Concern’ Commonly Occur in or Near Indels. Available online: https://virological.org/t/spike-protein-mutations-in-novel-sars-cov-2-variants-of-concern-commonly-occur-in-or-near-indels/605 (accessed on 25 February 2023).
- Gerdol, M.; Dishnica, K.; Giorgetti, A. Emergence of a recurrent insertion in the N-terminal domain of the SARS-CoV-2 spike glycoprotein. Virus Res. 2022, 310, 198674. [Google Scholar] [CrossRef]
- Uriu, K.; Cárdenas, P.; Muñoz, E.; Barragan, V.; Kosugi, Y.; Shirakawa, K.; Takaori-Kondo, A.; Ecuador-COVID19 Consortium, T.G.t.P.J.C.; Sato, K. Characterization of the Immune Resistance of Severe Acute Respiratory Syndrome Coronavirus 2 Mu Variant and the Robust Immunity Induced by Mu Infection. J. Infect. Dis. 2022, 226, 1200–1203. [Google Scholar] [CrossRef] [PubMed]
- Greco, S.; Gerdol, M. Independent acquisition of short insertions at the RIR1 site in the spike N-terminal domain of the SARS-CoV-2 BA.2 lineage. Transbounrday Emerg. Dis. 2022, 69, e3408–e3415. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Tang, W.; Wang, J.; Xiong, Y.; Li, B.; Yang, L.; Li, T.; Yang, L.; Su, X.; Li, Q.; et al. Outbreak Reports: An Outbreak of SARS-CoV-2 Omicron Subvariant BA.2.76 in an Outdoor Park—Chongqing Municipality, China, August 2022. China CDC Wkly. 2022, 4, 1039–1042. [Google Scholar]
- Peacock, T.; Bauer, D.; Barclay, W. Putative Host Origins of RNA Insertions in SARS-CoV-2 Genomes SARS-CoV-2 Coronavirus. Available online: https://virological.org/t/putative-host-origins-of-rna-insertions-in-sars-cov-2-genomes/761 (accessed on 9 December 2021).
- Venkatakrishnan, A.J.; Anand, P.; Lenehan, P.J.; Suratekar, R.; Raghunathan, B.; Niesen, M.J.M.; Soundararajan, V. On the origin of Omicron’s unique Spike gene insertion. Vaccines 2022, 10, 1509. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Nadeau, S.A.; Topolsky, I.; Manceau, M.; Huisman, J.S.; Jablonski, K.P.; Fuhrmann, L.; Dreifuss, D.; Jahn, K.; Beckmann, C.; et al. Quantification of the spread of SARS-CoV-2 variant B.1.1.7 in Switzerland. Epidemics 2021, 37, 100480. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Focosi, D.; Spezia, P.G.; Capria, A.-L.; Gueli, F.; McConnell, S.; Novazzi, F.; Pistello, M. Rise of the BQ.1.1.37 SARS-CoV-2 Sublineage, Italy. Diagnostics 2023, 13, 1000. https://doi.org/10.3390/diagnostics13051000
Focosi D, Spezia PG, Capria A-L, Gueli F, McConnell S, Novazzi F, Pistello M. Rise of the BQ.1.1.37 SARS-CoV-2 Sublineage, Italy. Diagnostics. 2023; 13(5):1000. https://doi.org/10.3390/diagnostics13051000
Chicago/Turabian StyleFocosi, Daniele, Pietro Giorgio Spezia, Anna-Lisa Capria, Federico Gueli, Scott McConnell, Federica Novazzi, and Mauro Pistello. 2023. "Rise of the BQ.1.1.37 SARS-CoV-2 Sublineage, Italy" Diagnostics 13, no. 5: 1000. https://doi.org/10.3390/diagnostics13051000