

Comparison of Two Commercially Available Interferon-γ Release Assays for T-Cell-Mediated Immunity and Evaluation of Humoral Immunity against SARS-CoV-2 in Healthcare Workers
 , ,
, , 
Abstract
:1. Introduction
2. Methods
2.1. Subjects and Sample Collection
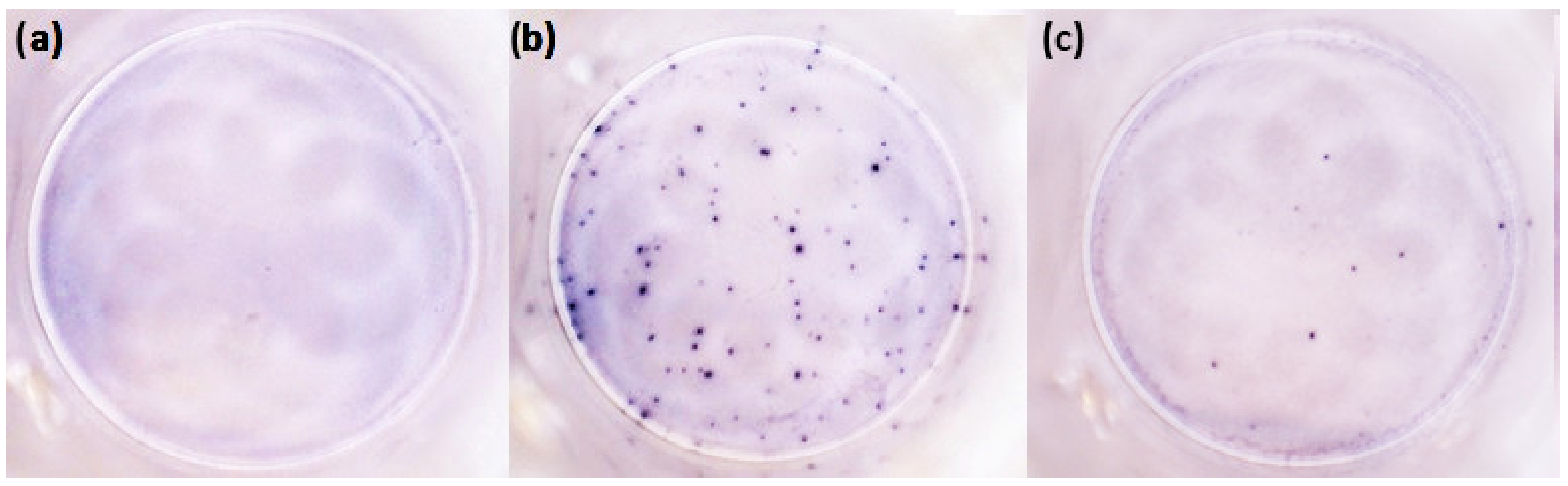
2.2. T-Cell-Mediated Immunity
2.3. Humoral Immunity
2.4. Statistical Methods
3. Results
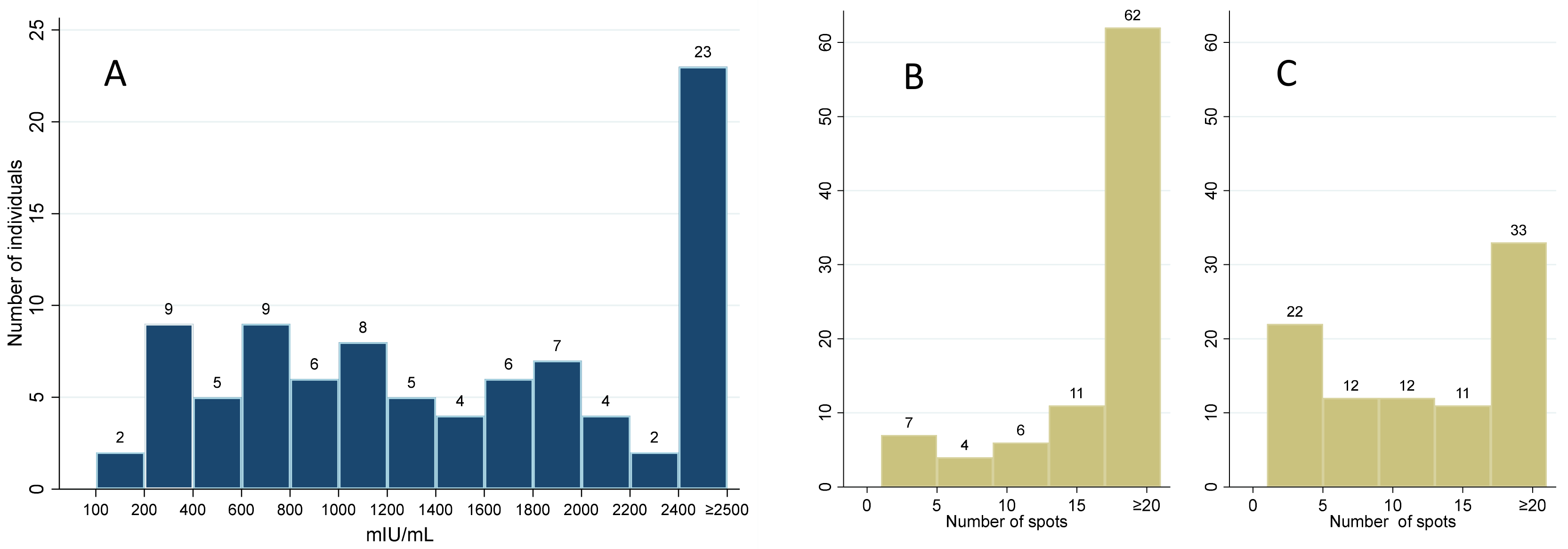
3.1. Study Group
3.2. Comparison of the Quan-T and T-Spot Tests
3.3. Correlations between Tests
3.4. Comparison of Humoral and Cellular Immunity Considering the Variant of the Past Infection
4. Discussion
4.1. T-Cell-Mediated Immunity and Comparison of the Quan-T and T-Spot Tests
4.2. Humoral Immunity
4.3. Associations between Humoral and Cellular Immunity
4.4. Study Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rashidzadeh, H.; Danafar, H.; Rahimi, H.; Mozafari, F.; Salehiabar, M.; Rahmati, M.A.; Rahamooz-Haghighi, S.; Mousazadeh, N.; Mohammadi, A.; Ertas, Y.N.; et al. Nanotechnology against the novel coronavirus (severe acute respiratory syndrome coronavirus 2): Diagnosis, treatment, therapy and future perspectives. Nanomedicine 2021, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Grunewald, M.; Perlman, S. Coronaviruses: An Updated Overview of Their Replication and Pathogenesis. In Methods in Molecular Biology; Humana: New York, NY, USA, 2020; Volume 2203, pp. 1–29. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.H.; Li, W.; Choe, H.; Farzan, M. What’s new in the renin-angiotensin system? CMLS Cell. Mol. Life Sci. 2004, 61, 2738–2743. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, A.; Hashemi, V.; Shomali, N.; Asghari, F.; Gharibi, T.; Akbari, M.; Gholizadeh, S.; Jafari, A. Innate and adaptive immune responses against coronavirus. Biomed Pharm. 2020, 132, 110859. [Google Scholar] [CrossRef]
- Zmievskaya, E.; Valiullina, A.; Ganeeva, I.; Petukhov, A.; Rizvanov, A.; Bulatov, E. Application of CAR-T Cell Therapy beyond Oncology: Autoimmune Diseases and Viral Infections. Biomedicines 2021, 9, 59. [Google Scholar] [CrossRef]
- Terreri, S.; Mortari, E.P.; Vinci, M.R.; Russo, C.; Alteri, C.; Albano, C.; Colavita, F.; Gramigna, G.; Agrati, C.; Linardos, G.; et al. Persistent B cell memory after SARS-CoV-2 vaccination is functional during breakthrough infections. Cell Host Microbe 2022, 30, 400–408.e4. [Google Scholar] [CrossRef]
- Primorac, D.; Brlek, P.; Matišić, V.; Molnar, V.; Vrdoljak, K.; Zadro, R.; Parčina, M. Cellular Immunity—The Key to Long-Term Protection in Individuals Recovered from SARS-CoV-2 and after Vaccination. Vaccines 2022, 10, 442. [Google Scholar] [CrossRef]
- Noh, J.Y.; Jeong, H.W.; Shin, E.-C. SARS-CoV-2 mutations, vaccines, and immunity: Implication of variants of concern. Signal Transduct. Target. Ther. 2021, 6, 203. [Google Scholar] [CrossRef]
- Tang, F.; Quan, Y.; Xin, Z.-T.; Wrammert, J.; Ma, M.-J.; Lv, H.; Wang, T.-B.; Yang, H.; Richardus, J.H.; Liu, W.; et al. Lack of peripheral memory B cell responses in recovered patients with severe acute respiratory syndrome: A six-year follow-up study. J. Immunol. 2011, 186, 7264–7268. [Google Scholar] [CrossRef]
- Swadling, L.; Maini, M.K. T cells in COVID-19—United in diversity. Nat. Immunol. 2020, 21, 1307–1308. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Dhawan, M.; Rabaan, A.A.; Fawarah, M.M.A.; Almuthree, S.A.; Alsubki, R.A.; Alfaraj, A.H.; Mashraqi, M.M.; Alshamrani, S.A.; Abduljabbar, W.A.; Alwashmi, A.S.S.; et al. Updated Insights into the T Cell-Mediated Immune Response against SARS-CoV-2: A Step towards Efficient and Reliable Vaccines. Vaccines 2023, 11, 101. [Google Scholar] [CrossRef]
- Oh, H.-L.J.; Chia, A.; Chang, C.X.L.; Leong, H.N.; Ling, K.L.; Grotenbreg, G.M.; Gehring, A.J.; Tan, Y.J.; Bertoletti, A. Engineering T Cells Specific for a Dominant Severe Acute Respiratory Syndrome Coronavirus CD8 T Cell Epitope. J. Virol. 2011, 85, 10464–10471. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, K.; Jagannathan, P.; Pham, T.D.; Pandey, S.; Bonilla, H.F.; Jacobson, K.; Parsonnet, J.; Andrews, J.R.; Weiskopf, D.; Sette, A.; et al. Interferon-γ Release Assay for Accurate Detection of Severe Acute Respiratory Syndrome Coronavirus 2 T-Cell Response. Clin. Infect. Dis. 2021, 73, e3130–e3132. [Google Scholar] [CrossRef]
- Mouton, W.; Compagnon, C.; Saker, K.; Daniel, S.; Djebali, S.; Lacoux, X.; Pozzetto, B.; Oriol, G.; Laubreton, D.; Prieux, M.; et al. Specific detection of memory T-cells in COVID-19 patients using standardized whole-blood Interferon gammarelease assay. Eur. J. Immunol. 2021, 51, 3239–3242. [Google Scholar] [CrossRef]
- Masiá, M.; Fernández-González, M.; García, J.A.; Padilla, S.; García-Abellán, J.; Botella, Á.; Mascarell, P.; Agulló, V.; Gutiérrez, F. Robust long-term immunity to SARS-CoV-2 in patients recovered from severe COVID-19 after interleukin-6 blockade. EBioMedicine 2022, 82, 104153. [Google Scholar] [CrossRef]
- Barreiro, P.; Sanz, J.C.; San Román, J.; Pérez-Abeledo, M.; Carretero, M.; Megías, G.; Viñuela-Prieto, J.M.; Ramos, B.; Canora, J.; Martínez-Peromingo, F.J.; et al. A Pilot Study for the Evaluation of an Interferon Gamma Release Assay (IGRA) To Measure T-Cell Immune Responses after SARS-CoV-2 Infection or Vaccination in a Unique Cloistered Cohort. J. Clin. Microbiol. 2022, 60, e0219921. [Google Scholar] [CrossRef]
- Seraceni, S.; Zocca, E.; Cervone, T.E.; Tomassetti, F.; Polidori, I.; Valisi, M.; Broccolo, F.; Calugi, G.; Bernardini, S.; Pieri, M. T-Cell Assay after COVID-19 Vaccination Could Be a Useful Tool? A Pilot Study on Interferon-Gamma Release Assay in Healthcare Workers. Diseases 2022, 10, 49. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, L.; Schank, M.; Dang, X.; Lu, Z.; Cao, D.; Khanal, S.; Nguyen, L.N.; Nguyen, L.N.T.; Zhang, J.; et al. SARS-CoV-2 specific memory T cell epitopes identified in COVID-19-recovered subjects. Virus Res. 2021, 304, 198508. [Google Scholar] [CrossRef]
- Guo, L.; Wang, G.; Wang, Y.; Zhang, Q.; Ren, L.; Gu, X.; Huang, T.; Zhong, J.; Wang, Y.; Wang, X.; et al. SARS-CoV-2-specific antibody and T-cell responses 1 year after infection in people recovered from COVID-19: A longitudinal cohort study. Lancet Microbe 2022, 3, e348–e356. [Google Scholar] [CrossRef] [PubMed]
- Kaaijk, P.; Pimentel, V.O.; Emmelot, M.E.; Poelen, M.C.M.; Cevirgel, A.; Schepp, R.M.; den Hartog, G.; Reukers, D.F.M.; Beckers, L.; van Beek, J.; et al. Children and Adults with Mild COVID-19: Dynamics of the Memory T Cell Response up to 10 Months. Front. Immunol. 2022, 13, 817876. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Dowell, A.C.; Pearce, H.; Verma, K.; Long, H.M.; Begum, J.; Aiano, F.; Amin-Chowdhury, Z.; Hoschler, K.; Brooks, T.; et al. Robust SARS-CoV-2-specific T cell immunity is maintained at 6 months following primary infection. Nat. Immunol. 2021, 22, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Šimánek, V.; Pecen, L.; Krátká, Z.; Fürst, T.; Řezáčková, H.; Topolčan, O.; Fajfrlík, K.; Sedláček, D.; Šín, R.; Pazdiora, P.; et al. Five Commercial Immunoassays for SARS-CoV-2 Antibody Determination and Their Comparison and Correlation with the Virus Neutralization Test. Diagnostics 2021, 11, 593. [Google Scholar] [CrossRef]
- Homza, M.; Zelena, H.; Janosek, J.; Tomaskova, H.; Jezo, E.; Kloudova, A.; Mrazek, J.; Svagera, Z.; Prymula, R. Covid-19 antigen testing: Better than we know? A test accuracy study. Infect. Dis. 2021, 53, 661–668. [Google Scholar] [CrossRef]
- Janošek, J.; Komárek, A. Post-infection immunity provides excellent protection from COVID-19 ICU hospitalization during Delta and Omicron waves. Infect. Dis. 2022, 55, 74–77. [Google Scholar] [CrossRef]
- Lindemann, M.; Klisanin, V.; Thümmler, L.; Fisenkci, N.; Tsachakis-Mück, N.; Ditschkowski, M.; Schwarzkopf, S.; Klump, H.; Reinhardt, H.C.; Horn, P.A.; et al. Humoral and Cellular Vaccination Responses against SARS-CoV-2 in Hematopoietic Stem Cell Transplant Recipients. Vaccines 2021, 9, 1075. [Google Scholar] [CrossRef]
- Tassignon, J.; Burny, W.; Dahmani, S.; Zhou, L.; Stordeur, P.; Byl, B.; De Groote, D. Monitoring of cellular responses after vaccination against tetanus toxoid: Comparison of the measurement of IFN-gamma production by ELISA, ELISPOT, flow cytometry and real-time PCR. J. Immunol. Methods 2005, 305, 188–198. [Google Scholar] [CrossRef]
- Phillips, E.; Adele, S.; Malone, T.; Deeks, A.; Stafford, L.; Dobson, S.L.; Amini, A.; Skelly, D.; Eyre, D.; Jeffery, K.; et al. Comparison of two T-cell assays to evaluate T-cell responses to SARS-CoV-2 following vaccination in naïve and convalescent healthcare workers. Clin. Exp. Immunol. 2022, 209, 90–98. [Google Scholar] [CrossRef]
- Prendecki, M.; Clarke, C.; Edwards, H.; McIntyre, S.; Mortimer, P.; Gleeson, S.; Martin, P.; Thomson, T.; Randell, P.; Shah, A.; et al. Humoral and T-cell responses to SARS-CoV-2 vaccination in patients receiving immunosuppression. Ann. Rheum. Dis. 2021, 80, 1322–1329. [Google Scholar] [CrossRef]
- Angyal, A.; Longet, S.; Moore, S.C.; Payne, R.P.; Harding, A.; Tipton, T.; Rongkard, P.; Ali, M.; Hering, L.M.; Meardon, N.; et al. T-cell and antibody responses to first BNT162b2 vaccine dose in previously infected and SARS-CoV-2-naive UK health-care workers: A multicentre prospective cohort study. Lancet Microbe 2022, 3, e21–e31. [Google Scholar] [CrossRef]
- Schiavoni, I.; Olivetta, E.; Natalucci, F.; Olivieri, G.; Lo Presti, A.; Fedele, G.; Stefanelli, P.; Ceccarelli, F.; Conti, F. Evidence of immune response to BNT162b2 COVID-19 vaccine in systemic lupus erythematosus patients treated with Belimumab. Lupus 2023. [Google Scholar] [CrossRef]
- Jordan, S.C. Innate and adaptive immune responses to SARS-CoV-2 in humans: Relevance to acquired immunity and vaccine responses. Clin. Exp. Immunol. 2021, 204, 310–320. [Google Scholar] [CrossRef]
- Cox, M.; Peacock, T.P.; Harvey, W.T.; Hughes, J.; Wright, D.W.; Willett, B.J.; Thomson, E.; Gupta, R.K.; Peacock, S.J.; Robertson, D.L.; et al. SARS-CoV-2 variant evasion of monoclonal antibodies based on in vitro studies. Nat. Rev. Microbiol. 2022, 21, 112–124. [Google Scholar] [CrossRef]
- Gallais, F.; Velay, A.; Nazon, C.; Wendling, M.-J.; Partisani, M.; Sibilia, J.; Candon, S.; Fafi-Kremer, S. Intrafamilial Exposure to SARS-CoV-2 Associated with Cellular Immune Response without Seroconversion, France. Emerg. Infect. Dis. 2021, 27, 113–121. [Google Scholar] [CrossRef]
- Goldberg, Y.; Mandel, M.; Bar-On, Y.M.; Bodenheimer, O.; Freedman, L.S.; Ash, N.; Alroy-Preis, S.; Huppert, A.; Milo, R. Protection and Waning of Natural and Hybrid Immunity to SARS-CoV-2. N. Engl. J. Med. 2022, 386, 2201–2212. [Google Scholar] [CrossRef]
- Altarawneh, H.N.; Chemaitelly, H.; Ayoub, H.H.; Tang, P.; Hasan, M.R.; Yassine, H.M.; Al-Khatib, H.A.; Smatti, M.K.; Coyle, P.; Al-Kanaani, Z.; et al. Effects of Previous Infection and Vaccination on Symptomatic Omicron Infections. N. Engl. J. Med. 2022, 387, 21–34. [Google Scholar] [CrossRef]
- Kim, T.-S.; Shin, E.-C. The activation of bystander CD8+ T cells and their roles in viral infection. Exp. Mol. Med. 2019, 51, 1–9. [Google Scholar] [CrossRef]
- Piler, P.; Thon, V.; Andrýsková, L.; Doležel, K.; Kostka, D.; Pavlík, T.; Dušek, L.; Pikhart, H.; Bobák, M.; Matic, S.; et al. Nationwide increases in anti-SARS-CoV-2 IgG antibodies between October 2020 and March 2021 in the unvaccinated Czech population. Commun. Med. 2022, 2, 19. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| N = 90 Individuals (100%) | T-Spot (Combined S and N) | |||
|---|---|---|---|---|
| Positive | Borderline Positive | Negative | ||
| Quan-T | positive | 81 (90.0%) | 2 (2.2%) | 5 (5.6%) |
| borderline pos. | 2 (2.2%) | 0 | 0 | |
| negative | 0 | 0 | 0 | |
| p-value (Test of symmetry) | 0.082 | |||
| Overall agreement * | 94.4% (95% CI: 87.5–98.2%) | |||
| Subgroup | ||||||
|---|---|---|---|---|---|---|
| All | All Vaccinated | All Convalescents | Unvaccinated Convalescents | Vaccinated Convalescents | Vaccinated Uninfected | |
| (Sub)group size (N) | 90 | 72 | 68 | 18 | 50 | 22 |
| T-Spot S vs. T-Spot N | 0.373 | 0.541 | 0.271 | 0.228 | 0.472 | 0.542 |
| T-Spot S vs. Quan-T | 0.376 | 0.343 | 0.346 | −0.082 | 0.208 | 0.545 |
| T-Spot S vs. IgG | 0.316 | 0.146 | 0.445 | 0.126 | 0.183 | 0.173 |
| T-Spot N vs. IgG | −0.117 | −0.140 | −0.017 | 0.321 | −0.144 | 0.086 |
| Quan-T vs. IgG | 0.454 | 0.304 | 0.497 | 0.005 | 0.321 | 0.305 |
| T-Spot S vs. VNT | 0.092 | −0.161 | 0.216 | −0.033 | −0.190 | −0.026 |
| T-Spot N vs. VNT | −0.172 | −0.164 | −0.058 | 0.147 | −0.108 | −0.081 |
| Quan-T vs. VNT | 0.304 | 0.090 | 0.385 | −0.194 | 0.182 | −0.056 |
| IgG vs. VNT | 0.668 | 0.421 | 0.725 | 0.885 | 0.435 | 0.334 |
| Group | N | Days from Last Immunization Impulse—Median (IQR) | Humoral Immunity | T-Cell-Mediated Immunity | ||||
|---|---|---|---|---|---|---|---|---|
| IgG ELISA | VNT | Quan-T | T-Spot (Total) | T-Spot (S) | T-Spot (N) | |||
| Omicron unvaccinated | 6 | 209 | 33.3% | 100% | 100% | 100% | 83.3% | 83.3% |
| (138–221) | (4/2/0) | (0/2/4) | (0/1/5) | (0/0/6) | (1/1/4) | (1/1/4) | ||
| Wuhan/Alpha/Delta unvaccinated | 6 | 315 (310–616) | 83.3% (1/0/5) | 83.3% (1/0/5) | 100% (0/0/6) | 100% (0/0/6) | 83.3% (1/0/5) | 100% (0/2/4) |
| Reinfected unvaccinated | 6 | 263 | 100% | 100% | 100% | 66.6% | 66.6% | 50.0% |
| (201–316) | (0/0/6) | (0/0/6) | (0/0/6) | (2/0/4) | (2/0/4) | (3/0/3) | ||
| Vaccinated convalescents | 50 | 238 | 100% | 100% | 100% | 100% | 100% | 84.0% |
| (204–307) | (0/0/50) | (0/0/50) | (0/1/49) | (0/1/49) | (0/1/49) | (8/4/38) | ||
| Vaccinated uninfected | 22 | 286 | 100% | 100% | 100% | 86.4% | 86.4% | 54.5% |
| (284–304) | (0/0/22) | (0/0/22) | (0/0/22) | (3/1/18) | (3/2/17) | (10/2/10) | ||
| Total | 90 | 284 | 94.4% | 98.9% | 100% | 94.4% | 92.2% | 75.6% |
| (209–307) | (5/2/83) | (1/2/87) | (0/2/88) | (5/2/83) | (7/4/79) | (22/9/59) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lochmanová, A.; Martinek, J.; Tomášková, H.; Zelená, H.; Dieckmann, K.; Grage-Griebenow, E.; Ježo, E.; Janošek, J. Comparison of Two Commercially Available Interferon-γ Release Assays for T-Cell-Mediated Immunity and Evaluation of Humoral Immunity against SARS-CoV-2 in Healthcare Workers. Diagnostics 2023, 13, 637. https://doi.org/10.3390/diagnostics13040637
Lochmanová A, Martinek J, Tomášková H, Zelená H, Dieckmann K, Grage-Griebenow E, Ježo E, Janošek J. Comparison of Two Commercially Available Interferon-γ Release Assays for T-Cell-Mediated Immunity and Evaluation of Humoral Immunity against SARS-CoV-2 in Healthcare Workers. Diagnostics. 2023; 13(4):637. https://doi.org/10.3390/diagnostics13040637
Chicago/Turabian StyleLochmanová, Alexandra, Jan Martinek, Hana Tomášková, Hana Zelená, Kersten Dieckmann, Evelin Grage-Griebenow, Eduard Ježo, and Jaroslav Janošek. 2023. "Comparison of Two Commercially Available Interferon-γ Release Assays for T-Cell-Mediated Immunity and Evaluation of Humoral Immunity against SARS-CoV-2 in Healthcare Workers" Diagnostics 13, no. 4: 637. https://doi.org/10.3390/diagnostics13040637