

Computational Design and Experimental Evaluation of MERS-CoV siRNAs in Selected Cell Lines
 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Analysis and siRNAs Selection
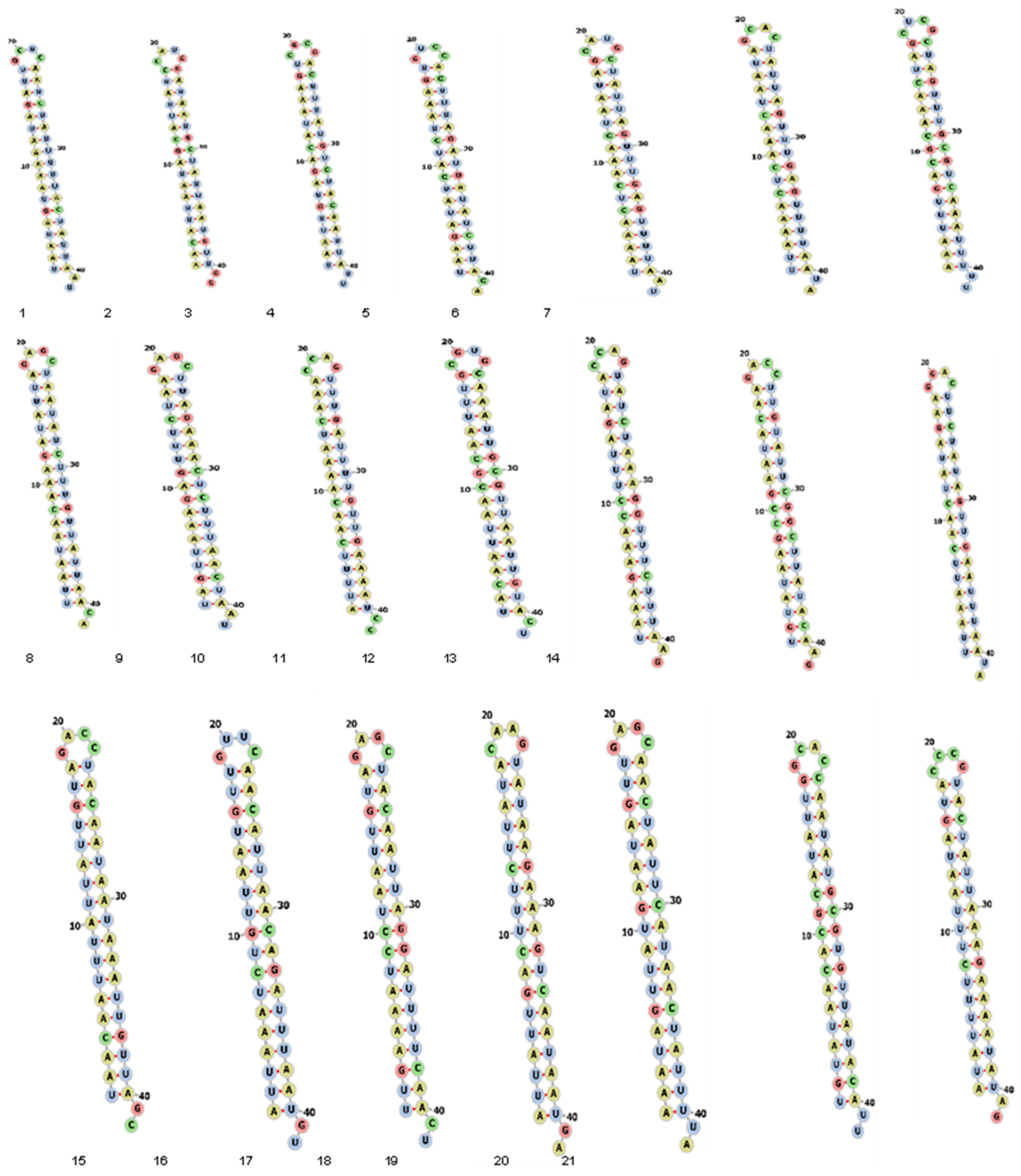
2.2. Secondary Structure Prediction of siRNAs
2.3. Cell Culture, Virus Propagation, and Virus Titering
2.4. siRNA Transfection and Cytotoxicity Assay
2.5. Evaluation of Virus Replication Inhibition by qRT-PCR
3. Results
3.1. Sequence Analysis and Prediction of siRNAs
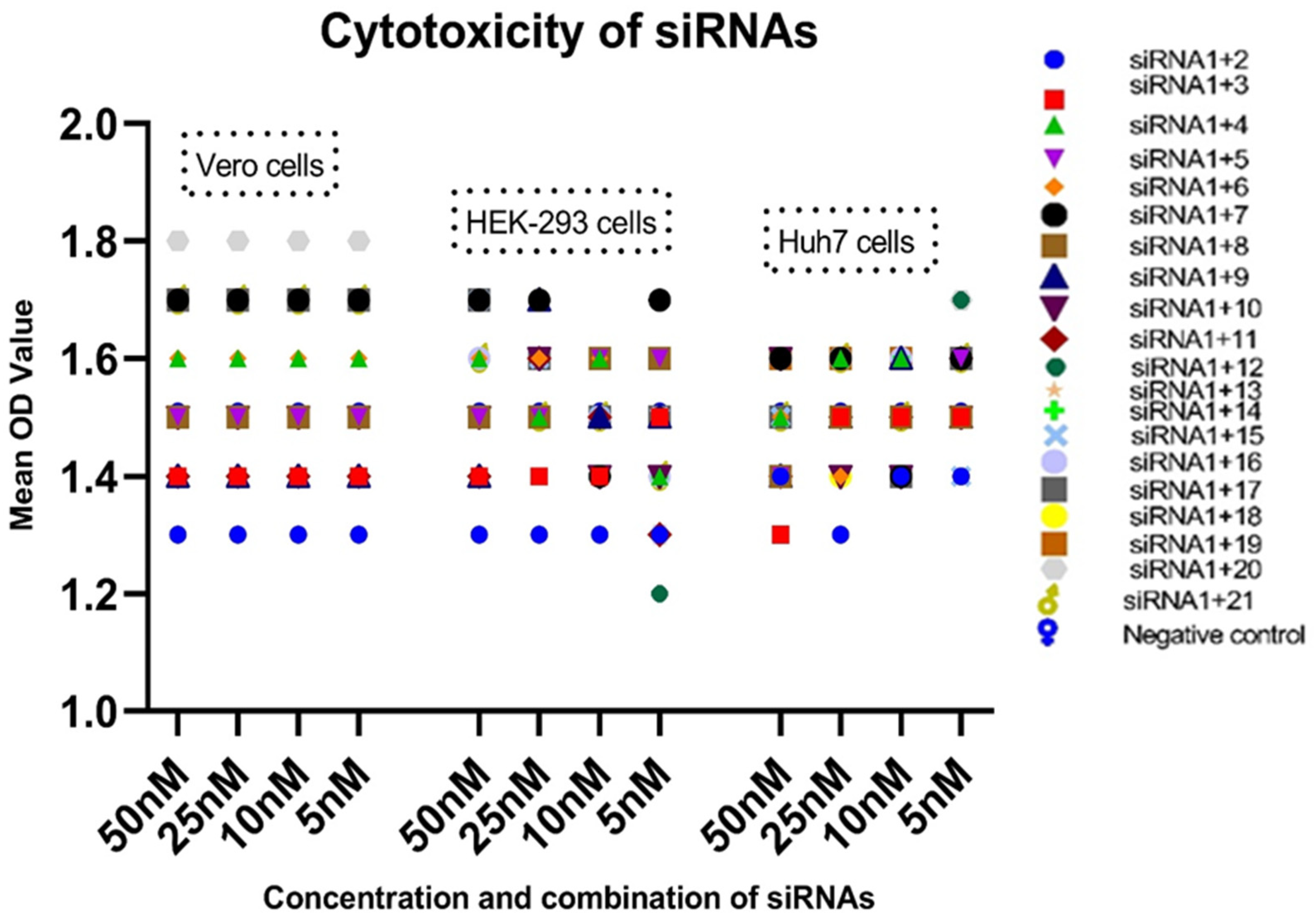
3.2. Cytotoxicity Assay
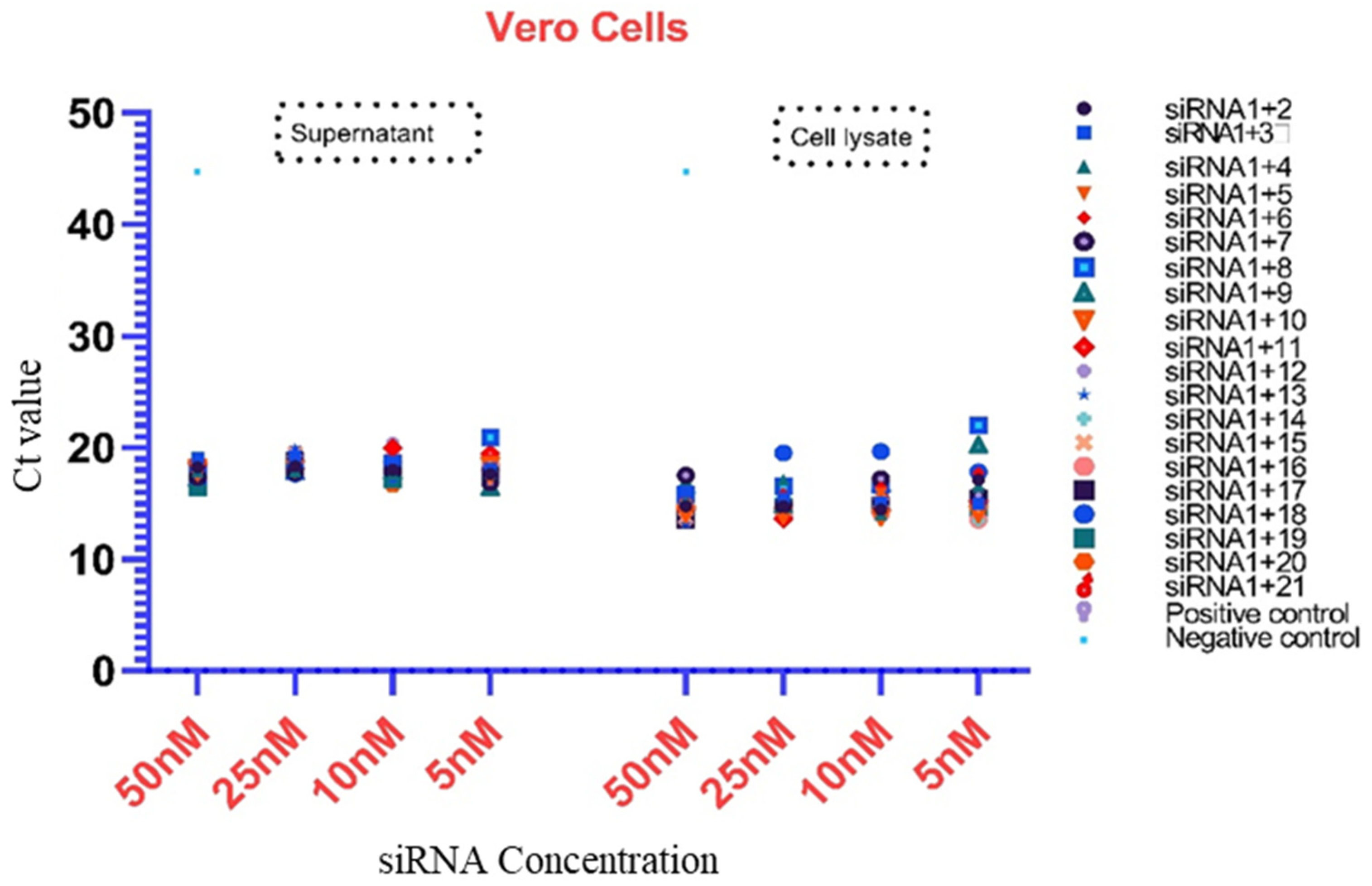
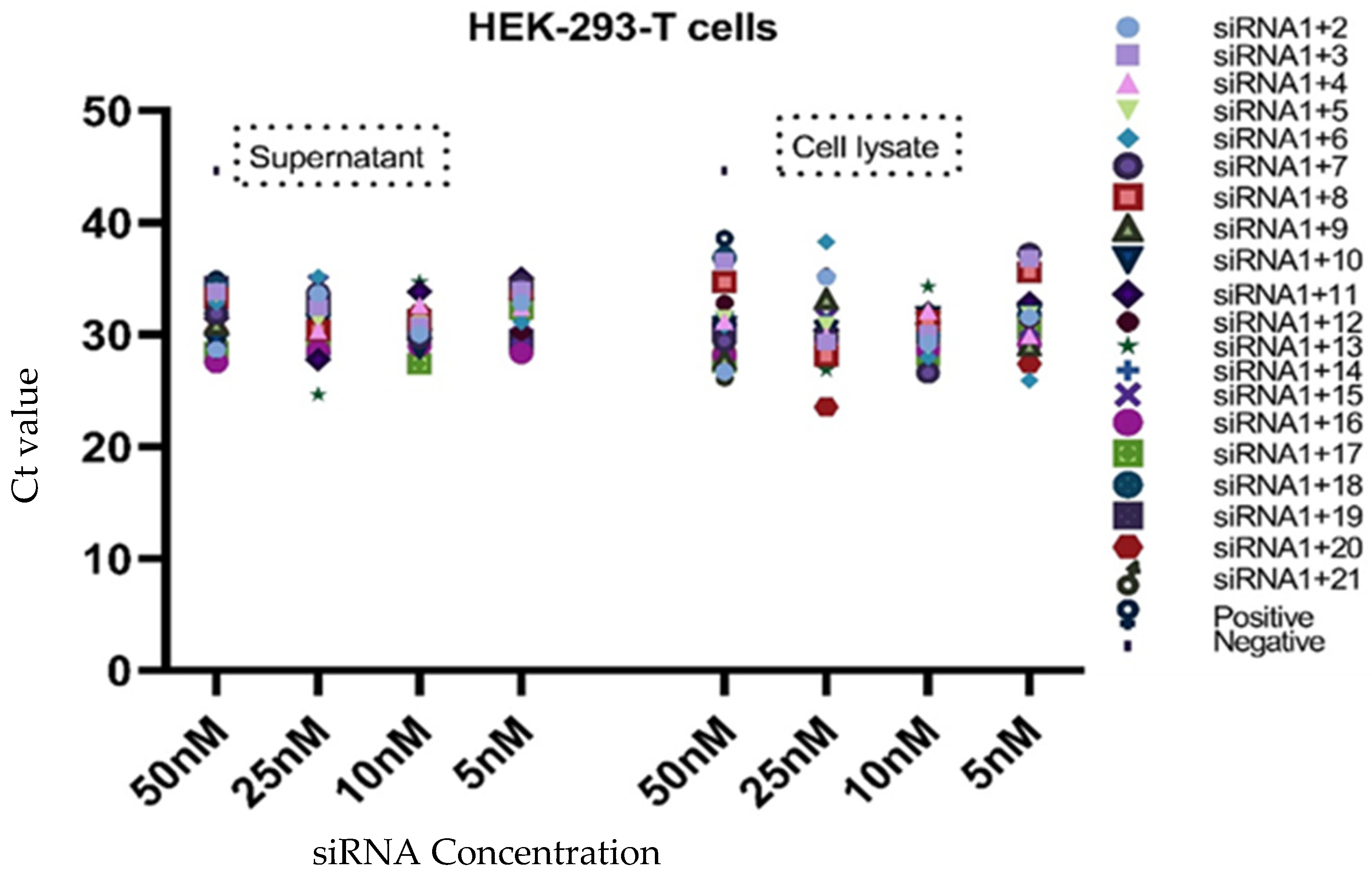
3.3. Analysis of Virus Inhibition
3.4. Prediction of Secondary Structure of siRNA
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Azhar, E.I.; El-Kafrawy, S.A.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Hashem, A.M.; Madani, T.A. Evidence for camel-to-human transmission of MERS coronavirus. N. Engl. J. Med. 2014, 370, 2499–2505. [Google Scholar] [CrossRef] [PubMed]
- Memish, Z.A.; Assiri, A.; Almasri, M.; Alhakeem, R.F.; Turkestani, A.; Al Rabeeah, A.A.; Al-Tawfiq, J.A.; Alzahrani, A.; Azhar, E.; Makhdoom, H.Q.; et al. Prevalence of MERS-CoV nasal carriage and compliance with the Saudi health recommendations among pilgrims attending the 2013 Hajj. J. Infect. Dis. 2014, 210, 1067–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oboho, I.K.; Tomczyk, S.M.; Al-Asmari, A.M.; Banjar, A.A.; Al-Mugti, H.; Aloraini, M.S.; Alkhaldi, K.Z.; Almohammadi, E.L.; Alraddadi, B.M.; Gerber, S.I.; et al. 2014 MERS-CoV outbreak in Jeddah--a link to health care facilities. N. Engl. J. Med. 2015, 372, 846–854. [Google Scholar] [CrossRef]
- Lee, S.S.; Wong, N.S. Probable transmission chains of Middle East respiratory syndrome coronavirus and the multiple generations of secondary infection in South Korea. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2015, 38, 65–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assiri, A.; Al-Tawfiq, J.A.; Al-Rabeeah, A.A.; Al-Rabiah, F.A.; Al-Hajjar, S.; Al-Barrak, A.; Flemban, H.; Al-Nassir, W.N.; Balkhy, H.H.; Al-Hakeem, R.F. Epidemiological, demographic, and clinical characteristics of 47 cases of Middle East respiratory syndrome coronavirus disease from Saudi Arabia: A descriptive study. Lancet Infect. Dis. 2013, 13, 752–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Wunderink, R.G. MERS, SARS and other coronaviruses as causes of pneumonia. Respirology 2018, 23, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Omari, A.; Rabaan, A.A.; Salih, S.; Al-Tawfiq, J.A.; Memish, Z.A. MERS coronavirus outbreak: Implications for emerging viral infections. Diagn. Microbiol. Infect. Dis. 2019, 93, 265–285. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Ithete, N.L.; Richards, L.R.; Schoeman, M.C.; Preiser, W.; Drosten, C.; Drexler, J.F. Rooting the phylogenetic tree of middle East respiratory syndrome coronavirus by characterization of a conspecific virus from an African bat. J. Virol. 2014, 88, 11297–11303. [Google Scholar] [CrossRef] [Green Version]
- Mahmoodi Chalbatani, G.; Dana, H.; Gharagouzloo, E.; Grijalvo, S.; Eritja, R.; Logsdon, C.D.; Memari, F.; Miri, S.R.; Rad, M.R.; Marmari, V. Small interfering RNAs (siRNAs) in cancer therapy: A nano-based approach. Int. J. Nanomed. 2019, 14, 3111–3128. [Google Scholar] [CrossRef] [Green Version]
- Hattab, D.; Gazzali, A.M.; Bakhtiar, A. Clinical Advances of siRNA-Based Nanotherapeutics for Cancer Treatment. Pharmaceutics 2021, 13, 1009. [Google Scholar] [CrossRef]
- Menanteau-Ledouble, S.; Schachner, O.; Lawrence, M.L.; El-Matbouli, M. Effects of siRNA silencing on the susceptibility of the fish cell line CHSE-214 to Yersinia ruckeri. Vet. Res. 2020, 51, 45. [Google Scholar] [CrossRef] [Green Version]
- Bruch, A.; Kelani, A.A.; Blango, M.G. RNA-based therapeutics to treat human fungal infections. Trends Microbiol. 2022, 30, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, H.; Chen, N.; Yang, J.; Zhou, H. LncRNA: A Potential Target for Host-Directed Therapy of Candida Infection. Pharmaceutics 2022, 14, 621. [Google Scholar] [CrossRef] [PubMed]
- Somarathne, M.; Gunawardene, Y.; Chandrasekharan, N.V.; Dassanayake, R.S. Development of siRNA mediated RNA interference and functional analysis of novel parasitic nematode-specific protein of Setaria digitata. Exp. Parasitol. 2018, 186, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Portet, A.; Galinier, R.; Lassalle, D.; Faille, A.; Gourbal, B.; Duval, D. Hemocyte siRNA uptake is increased by 5′ cholesterol-TEG addition in Biomphalaria glabrata, snail vector of schistosome. PeerJ 2021, 9, e10895. [Google Scholar] [CrossRef] [PubMed]
- Levanova, A.; Poranen, M.M. RNA interference as a prospective tool for the control of human viral infections. Front. Microbiol. 2018, 9, 2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setten, R.L.; Rossi, J.J.; Han, S.-P. The current state and future directions of RNAi-based therapeutics. Nat. Rev. Drug Discov. 2019, 18, 421–446. [Google Scholar] [CrossRef] [PubMed]
- Escalona-Noguero, C.; López-Valls, M.; Sot, B. CRISPR/Cas technology as a promising weapon to combat viral infections. BioEssays News Rev. Mol. Cell. Dev. Biol. 2021, 43, e2000315. [Google Scholar] [CrossRef]
- Baddeley, H.J.E.; Isalan, M. The Application of CRISPR/Cas Systems for Antiviral Therapy. Front. Genome Ed. 2021, 3. [Google Scholar] [CrossRef]
- Kong, H.; Ju, E.; Yi, K.; Xu, W.; Lao, Y.H.; Cheng, D.; Zhang, Q.; Tao, Y.; Li, M.; Ding, J. Advanced Nanotheranostics of CRISPR/Cas for Viral Hepatitis and Hepatocellular Carcinoma. Adv. Sci. 2021, 8, e2102051. [Google Scholar] [CrossRef]
- Lin, H.; Li, G.; Peng, X.; Deng, A.; Ye, L.; Shi, L.; Wang, T.; He, J. The Use of CRISPR/Cas9 as a Tool to Study Human Infectious Viruses. Front. Cell. Infect. Microbiol. 2021, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Najafi, S.; Tan, S.C.; Aghamiri, S.; Raee, P.; Ebrahimi, Z.; Jahromi, Z.K.; Rahmati, Y.; Sadri Nahand, J.; Piroozmand, A.; Jajarmi, V.; et al. Therapeutic potentials of CRISPR-Cas genome editing technology in human viral infections. Biomed. Pharmacother. 2022, 148, 112743. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.M.; Akter, R.; Ullah, M.S.; Abedin, M.J.; Ullah, G.M.; Hossain, M.Z. A Computational Approach for Predicting Role of Human MicroRNAs in MERS-CoV Genome. Adv. Bioinform. 2014, 2014, 967946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nur, S.M.; Hasan, M.A.; Amin, M.A.; Hossain, M.; Sharmin, T. Design of Potential RNAi (miRNA and siRNA) Molecules for Middle East Respiratory Syndrome Coronavirus (MERS-CoV) Gene Silencing by Computational Method. Interdiscip. Sci. Comput. Life Sci. 2015, 7, 257–265. [Google Scholar] [CrossRef]
- Fakhr, E.; Zare, F.; Teimoori-Toolabi, L. Precise and efficient siRNA design: A key point in competent gene silencing. Cancer Gene Ther. 2016, 23, 73–82. [Google Scholar] [CrossRef]
- Sohrab, S.S.; El-Kafrawy, S.A.; Mirza, Z.; Kamal, M.A.; Azhar, E.I. Design and Delivery of Therapeutic siRNAs: Application to MERS-Coronavirus. Curr. Pharm. Des. 2018, 24, 62–77. [Google Scholar] [CrossRef]
- Zhang, D.; Lu, J. In Silico Design of siRNAs Targeting Existing and Future Respiratory Viruses with VirusSi. bioRxiv 2020. [Google Scholar] [CrossRef]
- Sohrab, S.S.; Abbas, A.; Bajrai, L.; Azhar, E. In silico Prediction and Designing of Potential siRNAs to be Used as Antivirals Against SARS-CoV-2. Curr. Pharm. Des. 2021, 27, 3490–3500. [Google Scholar] [CrossRef]
- Sohrab, S.S.; Aly El-Kafrawy, S.; Mirza, Z.; Hassan, A.M.; Alsaqaf, F.; Azhar, E.I. In silico prediction and experimental validation of siRNAs targeting ORF1ab of MERS-CoV in Vero cell line. Saudi J. Biol. Sci. 2020, 23, 1348–1355. [Google Scholar] [CrossRef]
- Liu, C.; Liang, Z.; Kong, X. Efficacy Analysis of Combinatorial siRNAs against HIV Derived from One Double Hairpin RNA Precursor. Front. Microbiol. 2017, 8, 1651. [Google Scholar] [CrossRef]
- Taning, C.N.T.; Christiaens, O.; Li, X.; Swevers, L.; Casteels, H.; Maes, M.; Smagghe, G. Engineered Flock House Virus for Targeted Gene Suppression Through RNAi in Fruit Flies (Drosophila melanogaster) in Vitro and in Vivo. Front. Physiol. 2018, 9, 805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idrees, S.; Ashfaq, U.A. RNAi: Antiviral therapy against dengue virus. Asian Pac. J. Trop. Biomed. 2013, 3, 232–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahid, I.; AlMalki, W.H.; AlRabia, M.W.; Mukhtar, M.H.; Almalki, S.S.R.; Alkahtani, S.A.; Ashgar, S.S.; Faidah, H.S.; Hafeez, M.H. In vitro inhibitory analysis of consensus siRNAs against NS3 gene of hepatitis C virus 1a genotype. Asian Pac. J. Trop. Med. 2017, 10, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.T.; Lu, C.Y.; Shao, P.L.; Chang, L.Y.; Wang, J.Y.; Chang, Y.H.; Lai, M.J.; Chi, Y.H.; Huang, L.M. In vivo inhibition of influenza A virus replication by RNA interference targeting the PB2 subunit via intratracheal delivery. PLoS ONE 2017, 12, e0174523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, K.; Courtney, D.G.; Kennedy, E.M.; Cullen, B.R. Influenza A virus-derived siRNAs increase in the absence of NS1 yet fail to inhibit virus replication. RNA 2018, 24, 1172–1182. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.J.; Yang, L.; Zuo, J.P. Recent developments in antivirals against hepatitis B virus. Virus Res. 2016, 213, 205–213. [Google Scholar] [CrossRef]
- Zeng, J.; Yang, S.; Wang, X.; Gao, Y.; Zhang, M. Effects of siRNA-mediated suppression of HPV-11 L1 expression on the proliferation and apoptosis of vaginal epithelial cells. Exp. Ther. Med. 2017, 13, 1561–1565. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Jung, Y.S.; Liang, P.H. Anti-SARS coronavirus agents: A patent review (2008–present). Expert Opin. Ther. Pat. 2013, 23, 1337–1348. [Google Scholar] [CrossRef] [Green Version]
- Naito, Y.; Ui-Tei, K. siRNA Design Software for a Target Gene-Specific RNA Interference. Front. Genet. 2012, 3, 102. [Google Scholar] [CrossRef] [Green Version]
- ElHefnawi, M.; Kim, T.; Kamar, M.A.; Min, S.; Hassan, N.M.; El-Ahwany, E.; Kim, H.; Zada, S.; Amer, M.; Windisch, M.P. In Silico Design and Experimental Validation of siRNAs Targeting Conserved Regions of Multiple Hepatitis C Virus Genotypes. PLoS ONE 2016, 11, e0159211. [Google Scholar] [CrossRef]
- Coleman, C.M.; Frieman, M.B. Growth and Quantification of MERS-CoV Infection. Curr. Protoc. Microbiol. 2015, 37, 15E.2.1–15E.2.9. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.-W.; Chan, K.-H.; Choi, G.K.-Y.; To, K.K.-W.; Tse, H.; Cai, J.-P.; Yeung, M.L.; Cheng, V.C.-C.; Chen, H.; Che, X.-Y.; et al. Differential cell line susceptibility to the emerging novel human betacoronavirus 2c EMC/2012: Implications for disease pathogenesis and clinical manifestation. J. Infect. Dis. 2013, 207, 1743–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Wilde, A.H.; Raj, V.S.; Oudshoorn, D.; Bestebroer, T.M.; van Nieuwkoop, S.; Limpens, R.; Posthuma, C.C.; van der Meer, Y.; Bárcena, M.; Haagmans, B.L.; et al. MERS-coronavirus replication induces severe in vitro cytopathology and is strongly inhibited by cyclosporin A or interferon-α treatment. J. Gen. Virol. 2013, 94, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- Eckerle, I.; Corman, V.M.; Müller, M.A.; Lenk, M.; Ulrich, R.G.; Drosten, C. Replicative Capacity of MERS Coronavirus in Livestock Cell Lines. Emerg. Infect. Dis. 2014, 20, 276–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Kafrawy, S.A.; Sohrab, S.S.; Mirza, Z.; Hassan, A.M.; Alsaqaf, F.; Azhar, E.I. In Vitro Inhibitory Analysis of Rationally Designed siRNAs against MERS-CoV Replication in Huh7 Cells. Molecules 2021, 26, 2610. [Google Scholar] [CrossRef]
- Hashem, A.M.; Algaissi, A.; Agrawal, A.S.; Al-Amri, S.S.; Alhabbab, R.Y.; Sohrab, S.S.; Almasoud, A.; Alharbi, N.K.; Peng, B.-H.; Russell, M. A highly immunogenic, protective, and safe adenovirus-based vaccine expressing Middle East respiratory syndrome coronavirus S1-CD40L fusion protein in a transgenic human dipeptidyl peptidase 4 mouse model. J. Infect. Dis. 2019, 220, 1558–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mubarak, A.; Alturaiki, W.; Hemida, M.G. Middle East respiratory syndrome coronavirus (MERS-CoV): Infection, immunological response, and vaccine development. J. Immunol. Res. 2019, 6491738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Yang, Y.; Huang, J.; Jiang, S.; Du, L. Advances in MERS-CoV vaccines and therapeutics based on the receptor-binding domain. Viruses 2019, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Jia, W.; Wang, P.; Zhang, S.; Shi, X.; Wang, X.; Zhang, L. Antibodies and vaccines against Middle East respiratory syndrome coronavirus. Emerg. Microbes Infect. 2019, 8, 841–856. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-D.; Chi, W.-Y.; Su, J.-H.; Ferrall, L.; Hung, C.-F.; Wu, T.-C. Coronavirus vaccine development: From SARS and MERS to COVID-19. J. Biomed. Sci. 2020, 27, 104. [Google Scholar] [CrossRef]
- Folegatti, P.M.; Bittaye, M.; Flaxman, A.; Lopez, F.R.; Bellamy, D.; Kupke, A.; Mair, C.; Makinson, R.; Sheridan, J.; Rohde, C. Safety and immunogenicity of a candidate Middle East respiratory syndrome coronavirus viral-vectored vaccine: A dose-escalation, open-label, non-randomised, uncontrolled, phase 1 trial. Lancet Infect. Dis. 2020, 20, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, B.; Braga, A.C.; Batista, M.N.; Harris, M.; Rahal, P. Evaluation of canonical siRNA and Dicer substrate RNA for inhibition of hepatitis C virus genome replication—A comparative study. PLoS ONE 2015, 10, e0117742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.S.; Lee, S.H.; Kim, E.J.; Cho, H.; Lee, W.; Kim, G.W.; Park, H.J.; Cho, S.W.; Lee, C.; Oh, J.W. Inhibition of Hepatitis C Virus in Mice by a Small Interfering RNA Targeting a Highly Conserved Sequence in Viral IRES Pseudoknot. PLoS ONE 2016, 11, e0146710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Doss, C.G.P.; Lee, S.S. Therapeutic miRNA and siRNA: Moving from Bench to Clinic as Next Generation Medicine. Mol. Ther. Nucleic Acids 2017, 8, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Fukushige, K.; Tagami, T.; Naito, M.; Goto, E.; Hirai, S.; Hatayama, N.; Yokota, H.; Yasui, T.; Baba, Y.; Ozeki, T. Developing spray-freeze-dried particles containing a hyaluronic acid-coated liposome-protamine-DNA complex for pulmonary inhalation. Int. J. Pharm. 2020, 583, 119338. [Google Scholar] [CrossRef]
- Molina-Mora, J.A. Insights into the mutation T1117I in the spike and the lineage B.1.1.389 of SARS-CoV-2 circulating in Costa Rica. Gene Rep. 2022, 27, 101554. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Position of siRNA in the Genome (Start–End) | Target Sequence | Predicted RNA Oligo Sequences (5′→3′) | Minimum Free Energy (MFE (kcal/mol) and Frequency of Thermodynamic Ensemble (%) |
|---|---|---|---|---|
| 1 | 791–813 | agcaatctatttttactattaat | UAAUAGUAAAAAUAGAUUGCU CAAUCUAUUUUUACUAUUAAU | −17.96, 65.21 |
| 2 | 1615–1637 | atggataatgctattaatgttgg | AACAUUAAUAGCAUUAUCCAU GGAUAAUGCUAUUAAUGUUGG | −21.80, 77.37 |
| 3 | 1910–1932 | gcgactttatgtctacaattatt | UAAUUGUAGACAUAAAGUCGC GACUUUAUGUCUACAAUUAUU | −22.02, 69.74 |
| 4 | 4018–4040 | gacactttagatgatatcttaca | UAAGAUAUCAUCUAAAGUGUC CACUUUAGAUGAUAUCUUACA | −22.62, 69.54 |
| 5 | 5597–5619 | atgctattagtttgagttttaat | UAAAACUCAAACUAAUAGCAU GCUAUUAGUUUGAGUUUUAAU | −21.51, 83.64 |
| 6 | 5598–5620 | tgctattagtttgagttttaata | UUAAAACUCAAACUAAUAGCA CUAUUAGUUUGAGUUUUAAUA | −19.74, 57.91 |
| 7 | 5819–5841 | gagctagtttgcgtcaaattttt | AAAUUUGACGCAAACUAGCUC GCUAGUUUGCGUCAAAUUUUU | −24.28, 53.63 |
| 8 | 9495–9517 | ctctaatatctttgttattaaca | UUAAUAACAAAGAUAUUAGAG CUAAUAUCUUUGUUAUUAACA | −17.97, 54.45 |
| 9 | 9533–9555 | ctcttagaaactctttaactaat | UAGUUAAAGAGUUUCUAAGAG CUUAGAAACUCUUUAACUAAU | −22.37, 64.54 |
| 10 | 13,605–13,627 | tggtttgattttgttgaaaatcc | AUUUUCAACAAAAUCAAACCA GUUUGAUUUUGUUGAAAAUCC | −18.34, 35.22 |
| 11 | 14,005–14,027 | acgcaaattgcgttaattgtact | UACAAUUAACGCAAUUUGCGU GCAAAUUGCGUUAAUUGUACU | −22.34, 79.46 |
| 12 | 14,389–14,411 | tggtatctaaaggtttctttaag | UAAAGAAACCUUUAGAUACCA GUAUCUAAAGGUUUCUUUAAG | −22.04, 67.95 |
| 13 | 16,177–16,199 | gtcttgtattcggcttatacaag | UGUAUAAGCCGAAUACAAGAC CUUGUAUUCGGCUUAUACAAG | −26.53, 58.68 |
| 14 | 16,217–16,239 | tccttctatagttgaatttaata | UUAAAUUCAACUAUAGAAGGA CUUCUAUAGUUGAAUUUAAUA | −20.24, 48.81 |
| 15 | 17,283–17,305 | gtctacaataataaattgttagc | UAACAAUUUAUUAUUGUAGAC CUACAAUAAUAAAUUGUUAGC | −17.87, 75.42 |
| 16 | 17,583–17,605 | aacaacattaacagatttaatgt | AUUAAAUCUGUUAAUGUUGUU CAACAUUAACAGAUUUAAUGU | −19.59, 62.23 |
| 17 | 18,028–18,050 | ctctacaattaggattttcaact | UUGAAAAUCCUAAUUGUAGAG CUACAAUUAGGAUUUUCAACU | −22.08, 53.94 |
| 18 | 19,806–19,828 | ttgtataagaaagtcaataatga | AUUAUUGACUUUCUUAUACAA GUAUAAGAAAGUCAAUAAUGA | −19.97, 64.53 |
| 19 | 20,090–20,112 | ctcaactattcataactatttta | AAAUAGUUAUGAAUAGUUGAG CAACUAUUCAUAACUAUUUUA | −19.63, 42.01 |
| 20 | 20,498–20,520 | tgccaatatgcgtgttatacatt | UGUAUAACACGCAUAUUGGCA CCAAUAUGCGUGUUAUACAUU | −25.98, 74.21 |
| 21 | 20,948–20,970 | gggtactattaaagaaaatatag | AUAUUUUCUUUAAUAGUACCC GUACUAUUAAAGAAAAUAUAG | −17.65, 66.76 |
| siRNAs Combination | siRNA Concentrations (nM)/OD Value in Vero Cells | siRNA Concentrations (nM)/OD Value in HEK-293-T Cells | siRNA Concentrations (nM)/OD Value in Huh7 Cells | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 50 | 25 | 10 | 5.0 | CC50 | 50 | 25 | 10 | 5.0 | CC50 | 50 | 25 | 10 | 5.0 | CC50 | |
| siRNA-1+2 | 1.3 | 1.3 | 1.3 | 1.3 | >100 | 1.3 | 1.3 | 1.3 | 1.3 | >100 | 1.4 | 1.3 | 1.4 | 1.4 | >100 |
| siRNA-1+3 | 1.4 | 1.4 | 1.4 | 1.4 | >100 | 1.4 | 1.4 | 1.4 | 1.5 | >100 | 1.3 | 1.5 | 1.5 | 1.5 | >100 |
| siRNA-1+4 | 1.6 | 1.6 | 1.6 | 1.6 | >100 | 1.6 | 1.5 | 1.6 | 1.4 | >100 | 1.5 | 1.6 | 1.6 | 1.5 | >100 |
| siRNA-1+5 | 1.5 | 1.5 | 1.5 | 1.5 | >100 | 1.5 | 1.5 | 1.6 | 1.6 | >100 | 1.4 | 1.5 | 1.5 | 1.6 | >100 |
| siRNA-1+6 | 1.6 | 1.6 | 1.6 | 1.6 | >100 | 1.6 | 1.6 | 1.6 | 1.5 | >100 | 1.5 | 1.4 | 1.5 | 1.5 | >100 |
| siRNA-1+7 | 1.7 | 1.7 | 1.7 | 1.7 | >100 | 1.7 | 1.7 | 1.4 | 1.7 | >100 | 1.6 | 1.6 | 1.4 | 1.6 | >100 |
| siRNA-1+8 | 1.5 | 1.5 | 1.5 | 1.5 | >100 | 1.5 | 1.5 | 1.6 | 1.6 | >100 | 1.4 | 1.5 | 1.5 | 1.5 | >100 |
| siRNA-1+9 | 1.4 | 1.4 | 1.4 | 1.4 | >100 | 1.4 | 1.7 | 1.5 | 1.5 | >100 | 1.4 | 1.5 | 1.6 | 1.5 | >100 |
| siRNA-1+10 | 1.5 | 1.5 | 1.5 | 1.5 | >100 | 1.5 | 1.6 | 1.4 | 1.4 | >100 | 1.6 | 1.4 | 1.4 | 1.5 | >100 |
| siRNA-1+11 | 1.4 | 1.4 | 1.4 | 1.4 | >100 | 1.4 | 1.6 | 1.5 | 1.3 | >100 | 1.4 | 1.5 | 1.5 | 1.6 | >100 |
| siRNA-1+12 | 1.5 | 1.5 | 1.5 | 1.5 | >100 | 1.5 | 1.4 | 1.3 | 1.2 | >100 | 1.6 | 1.3 | 1.4 | 1.7 | >100 |
| siRNA-1+13 | 1.4 | 1.4 | 1.4 | 1.4 | >100 | 1.6 | 1.4 | 1.5 | 1.5 | >100 | 1.5 | 1.4 | 1.5 | 1.6 | >100 |
| siRNA-1+14 | 1.5 | 1.5 | 1.5 | 1.5 | >100 | 1.5 | 1.6 | 1.5 | 1.4 | >100 | 1.4 | 1.5 | 1.6 | 1.5 | >100 |
| siRNA-1+15 | 1.5 | 1.5 | 1.5 | 1.5 | >100 | 1.7 | 1.6 | 1.5 | 1.5 | >100 | 1.5 | 1.6 | 1.5 | 1.4 | >100 |
| siRNA-1+16 | 1.5 | 1.5 | 1.5 | 1.5 | >100 | 1.6 | 1.5 | 1.5 | 1.4 | >100 | 1.6 | 1.6 | 1.6 | 1.5 | >100 |
| siRNA-1+17 | 1.7 | 1.7 | 1.7 | 1.7 | >100 | 1.7 | 1.6 | 1.5 | 1.5 | >100 | 1.5 | 1.5 | 1.4 | 1.6 | >100 |
| siRNA-1+18 | 1.5 | 1.5 | 1.5 | 1.5 | >100 | 1.5 | 1.6 | 1.5 | 1.4 | >100 | 1.4 | 1.4 | 1.5 | 1.5 | >100 |
| siRNA-1+19 | 1.7 | 1.7 | 1.7 | 1.7 | >100 | 1.7 | 1.6 | 1.5 | 1.5 | >100 | 1.6 | 1.6 | 1.6 | 1.6 | >100 |
| siRNA-1+20 | 1.8 | 1.8 | 1.8 | 1.8 | >100 | 1.5 | 1.5 | 1.4 | 1.3 | >100 | 1.4 | 1.4 | 1.5 | 1.7 | >100 |
| siRNA-1+21 | 1.7 | 1.7 | 1.7 | 1.7 | >100 | 1.6 | 1.5 | 1.5 | 1.4 | >100 | 1.5 | 1.6 | 1.5 | 1.6 | >100 |
| Negative | 1.5 | 1.5 | 1.5 | 1.5 | >100 | 1.5 | 1.5 | 1.5 | 1.5 | >100 | 1.5 | 1.5 | 1.5 | 1.5 | >100 |
| siRNAs Combinations | Vero Cells | |||||||
|---|---|---|---|---|---|---|---|---|
| (Cell Supernatant)—(nM) | (Cell Lysate)—(nM) | |||||||
| 50 | 25 | 10 | 5.0 | 50 | 25 | 10 | 5.0 | |
| siRNA1+2 | 18.25 | 18.31 | 18.01 | 17.64 | 14.75 | 14.74 | 14.43 | 17.13 |
| siRNA1+3 | 19.07 | 19.24 | 18.68 | 18.08 | 16.06 | 15.48 | 15.06 | 14.99 |
| siRNA1+4 | 17.87 | 17.89 | 17.66 | 17.77 | 15.32 | 17.11 | 14.06 | 16.59 |
| siRNA1+5 | 17.41 | 17.76 | 17.97 | 17.13 | 14.80 | 14.44 | 15.87 | 16.53 |
| siRNA1+6 | 17.83 | 18.99 | 19.68 | 17.82 | 15.61 | 15.89 | 16.43 | 17.74 |
| siRNA1+7 | 17.34 | 17.80 | 17.97 | 16.87 | 17.50 | 14.86 | 17.22 | 15.75 |
| siRNA1+8 | 17.48 | 17.85 | 18.58 | 20.90 | 15.82 | 16.56 | 16.63 | 21.96 |
| siRNA1+9 | 17.25 | 17.84 | 17.91 | 16.46 | 16.62 | 14.88 | 16.76 | 20.23 |
| siRNA1+10 | 18.24 | 18.14 | 18.06 | 18.46 | 14.09 | 13.96 | 13.78 | 14.11 |
| siRNA1+11 | 18.59 | 18.10 | 19.97 | 19.46 | 14.46 | 13.60 | 14.33 | 15.21 |
| siRNA1+12 | 18.08 | 17.71 | 20.45 | 18.24 | 14.62 | 14.28 | 14.80 | 14.61 |
| siRNA1+13 | 18.26 | 19.97 | 18.07 | 18.63 | 13.17 | 14.50 | 14.15 | 15.60 |
| siRNA1+14 | 18.32 | 18.63 | 18.02 | 17.42 | 15.19 | 14.31 | 14.38 | 13.48 |
| siRNA1+15 | 18.32 | 19.61 | 19.68 | 19.12 | 13.67 | 14.41 | 15.07 | 14.76 |
| siRNA1+16 | 17.90 | 18.06 | 18.97 | 18.71 | 14.73 | 14.01 | 14.11 | 13.50 |
| siRNA1+17 | 17.64 | 18.91 | 18.09 | 17.97 | 13.52 | 14.41 | 14.88 | 15.48 |
| siRNA1+18 | 17.20 | 17.67 | 17.66 | 17.12 | 15.75 | 19.54 | 19.67 | 17.81 |
| siRNA1+19 | 16.42 | 17.81 | 17.10 | 17.48 | 14.51 | 14.78 | 14.76 | 14.59 |
| siRNA1+20 | 18.48 | 17.68 | 16.67 | 17.97 | 15.27 | 15.17 | 15.14 | 15.35 |
| siRNA1+21 | 17.86 | 18.97 | 18.25 | 18.37 | 14.88 | 14.94 | 15.31 | 14.70 |
| Positive control | 17.85 | 16.04 | ||||||
| Negative control | 45.00 | 45.00 | ||||||
| siRNAs Combinations | HEK-293-T Cells | |||||||
|---|---|---|---|---|---|---|---|---|
| (Cell Supernatant)—(nM) | (Cell Lysate)—(nM) | |||||||
| 50 | 25 | 10 | 5.0 | 50 | 25 | 10 | 5.0 | |
| siRNA1+2 | 28.62 | 33.66 | 30.06 | 32.81 | 26.69 | 35.10 | 29.21 | 31.55 |
| siRNA1+3 | 33.84 | 32.42 | 30.77 | 33.96 | 36.58 | 29.32 | 30.10 | 36.76 |
| siRNA1+4 | 33.88 | 30.34 | 32.63 | 32.44 | 31.20 | 29.89 | 32.14 | 29.89 |
| siRNA1+5 | 33.56 | 31.24 | 30.92 | 32.94 | 31.40 | 30.83 | 29.02 | 31.75 |
| siRNA1+6 | 32.86 | 35.17 | 29.87 | 31.10 | 30.78 | 38.28 | 27.96 | 25.90 |
| siRNA1+7 | 31.87 | 33.71 | 29.73 | 34.48 | 29.42 | 29.78 | 26.60 | 37.23 |
| siRNA1+8 | 33.23 | 30.37 | 31.35 | 34.04 | 34.71 | 28.11 | 31.24 | 35.56 |
| siRNA1+9 | 30.88 | 30.37 | 31.35 | 34.04 | 27.77 | 33.15 | 31.82 | 29.01 |
| siRNA1+10 | 29.35 | 30.92 | 28.85 | 34.17 | 30.72 | 30.32 | 31.60 | 31.73 |
| siRNA1+11 | 31.44 | 27.76 | 33.83 | 35.02 | 30.86 | 30.35 | 31.96 | 32.82 |
| siRNA1+12 | 28.58 | 31.44 | 31.36 | 30.21 | 32.84 | 35.20 | 30.61 | 31.23 |
| siRNA1+13 | 30.60 | 24.63 | 34.69 | 32.46 | 29.46 | 26.86 | 34.27 | 28.80 |
| siRNA1+14 | 32.03 | 31.94 | 31.72 | 33.09 | 31.97 | 31.17 | 30.59 | 31.96 |
| siRNA1+15 | 29.57 | 34.56 | 30.26 | 29.91 | 29.91 | 31.45 | 31.53 | 30.58 |
| siRNA1+16 | 27.51 | 28.53 | 29.00 | 28.33 | 28.15 | 29.48 | 28.59 | 29.14 |
| siRNA1+17 | 28.42 | 29.69 | 27.40 | 32.28 | 27.54 | 29.25 | 28.12 | 30.53 |
| siRNA1+18 | 34.18 | 28.25 | 29.47 | 31.94 | 36.87 | 27.93 | 29.93 | 31.84 |
| siRNA1+19 | 34.23 | 32.65 | 30.54 | 29.49 | 30.31 | 29.67 | 29.66 | 31.06 |
| siRNA1+20 | 30.28 | 28.46 | 31.86 | 29.04 | 28.49 | 23.53 | 31.28 | 27.38 |
| siRNA1+21 | 29.06 | 29.94 | 30.36 | 29.46 | 26.61 | 28.42 | 28.77 | 29.66 |
| Positive control | 34.50 | 38.11 | ||||||
| Negative control | 45.00 | 45.00 | ||||||
| siRNAs Combination | Huh-7 Cells | |||||||
|---|---|---|---|---|---|---|---|---|
| (Cell Supernatant)—(nM) | (Cell Lysate)—(nM) | |||||||
| 50 | 25 | 10 | 5.0 | 50 | 25 | 10 | 5.0 | |
| siRNA1+2 | 20.77 | 21.97 | 21.02 | 21.07 | 15.71 | 17.99 | 16.25 | 16.60 |
| siRNA1+3 | 23.42 | 23.54 | 21.31 | 22.77 | 18.04 | 19.54 | 16.90 | 18.17 |
| siRNA1+4 | 20.77 | 21.94 | 21.22 | 22.20 | 17.68 | 19.07 | 17.89 | 19.03 |
| siRNA1+5 | 22.82 | 22.10 | 21.86 | 21.87 | 17.05 | 19.00 | 16.72 | 17.08 |
| siRNA1+6 | 21.80 | 22.59 | 20.53 | 19.43 | 17.55 | 17.17 | 15.19 | 16.22 |
| siRNA1+7 | 21.15 | 20.91 | 20.66 | 21.00 | 17.57 | 17.27 | 16.32 | 16.18 |
| siRNA1+8 | 21.24 | 20.44 | 20.53 | 20.41 | 17.21 | 16.41 | 19.15 | 16.93 |
| siRNA1+9 | 20.10 | 18.97 | 18.96 | 20.09 | 16.42 | 15.37 | 16.06 | 15.73 |
| siRNA1+10 | 21.17 | 21.51 | 20.68 | 21.12 | 18.11 | 18.50 | 23.11 | 17.25 |
| siRNA1+11 | 19.99 | 20.48 | 21.10 | 19.86 | 23.87 | 17.88 | 18.02 | 18.29 |
| siRNA1+12 | 22.03 | 22.49 | 19.85 | 22.70 | 17.92 | 18.72 | 18.48 | 18.97 |
| siRNA1+13 | 24.33 | 20.02 | 21.31 | 23.48 | 18.85 | 18.60 | 18.03 | 19.05 |
| siRNA1+14 | 23.53 | 22.03 | 20.95 | 22.26 | 18.42 | 19.12 | 18.66 | 17.26 |
| siRNA1+15 | 20.81 | 22.31 | 23.25 | 22.65 | 18.97 | 19.02 | 23.94 | 20.26 |
| siRNA1+16 | 21.06 | 22.05 | 21.95 | 20.61 | 18.90 | 18.74 | 20.82 | 20.40 |
| siRNA1+17 | 19.99 | 19.93 | 19.36 | 19.05 | 19.38 | 18.93 | 17.12 | 16.61 |
| siRNA1+18 | 24.39 | 21.99 | 23.44 | 23.74 | 18.36 | 18.17 | 18.60 | 22.40 |
| siRNA1+19 | 22.36 | 22.17 | 23.18 | 23.26 | 18.10 | 17.89 | 20.95 | 19.88 |
| siRNA1+20 | 23.97 | 23.39 | 23.87 | 22.92 | 18.62 | 19.69 | 20.76 | 21.18 |
| siRNA1+21 | 22.89 | 23.42 | 23.36 | 22.98 | 18.86 | 21.25 | 19.98 | 19.73 |
| Positive control | 21.10 | 19.18 | ||||||
| Negative control | 45.00 | 45.00 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sohrab, S.S.; El-Kafrawy, S.A.; Mirza, Z.; Hassan, A.M.; Alsaqaf, F.; Azhar, E.I. Computational Design and Experimental Evaluation of MERS-CoV siRNAs in Selected Cell Lines. Diagnostics 2023, 13, 151. https://doi.org/10.3390/diagnostics13010151
Sohrab SS, El-Kafrawy SA, Mirza Z, Hassan AM, Alsaqaf F, Azhar EI. Computational Design and Experimental Evaluation of MERS-CoV siRNAs in Selected Cell Lines. Diagnostics. 2023; 13(1):151. https://doi.org/10.3390/diagnostics13010151
Chicago/Turabian StyleSohrab, Sayed S., Sherif A. El-Kafrawy, Zeenat Mirza, Ahmed M. Hassan, Fatima Alsaqaf, and Esam I. Azhar. 2023. "Computational Design and Experimental Evaluation of MERS-CoV siRNAs in Selected Cell Lines" Diagnostics 13, no. 1: 151. https://doi.org/10.3390/diagnostics13010151