
HLA-G 14bp Ins/Del Polymorphism, Plasma Level of Soluble HLA-G, and Association with IL-6/IL-10 Ratio and Survival of Glioma Patients

 ,
,
Abstract
:1. Introduction
2. Subjects and Methods
2.1. Study Groups
2.2. Procedures and Sample Processing
2.3. Statistical Analysis
3. Results
3.1. Comparison of HLA-G 14bp Ins/Del Allele and Genotype Frequencies between Glioma Patients and Control Group of Healthy Subjects
3.2. Comparison of Plasma Levels of sHLA-G in Glioma Patients and Healthy Controls
3.3. The Effect of HLA-G 14bp Ins/Del Polymorphism on sHLA-G Plasma Levels in Glioma Patients
3.4. Analysis of the Association of Plasma Levels of sHLA-G with a O6-Methylguanine-DNA Methyl-Transferase (MGMT) Promoter Methylation Status in Glioma Patients
3.5. Association of Plasma Levels of Immunosuppressive Molecule sHLA-G with the Levels of Pro-Inflammatory Cytokine IL-6, Anti-Inflammatory Cytokine IL-10, and IL-10/IL-6 Ratio in Glioma Patients
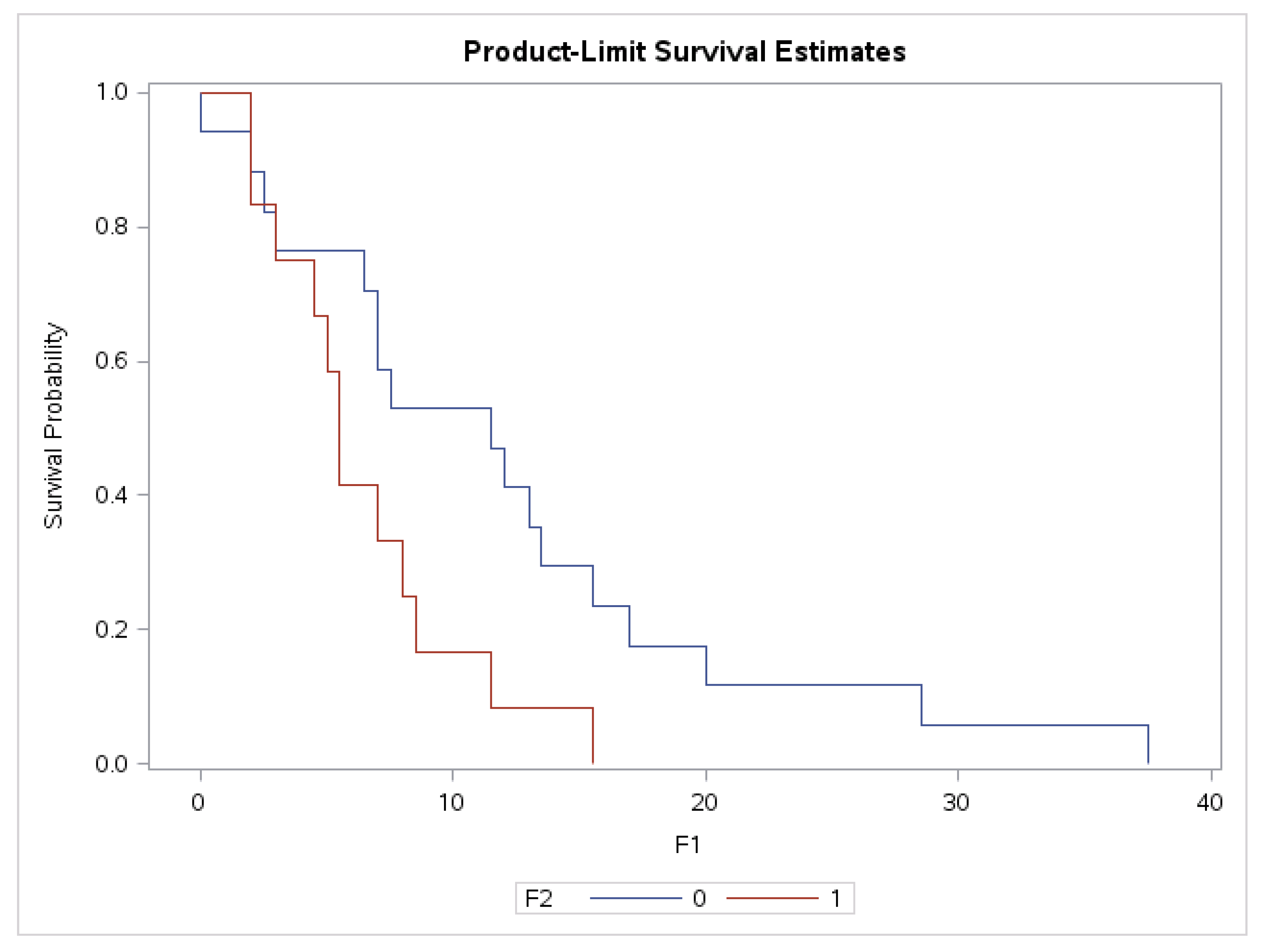
3.6. Association of Plasma Levels of sHLA-G with Survival Time in Grade II and IV Glioma Patients
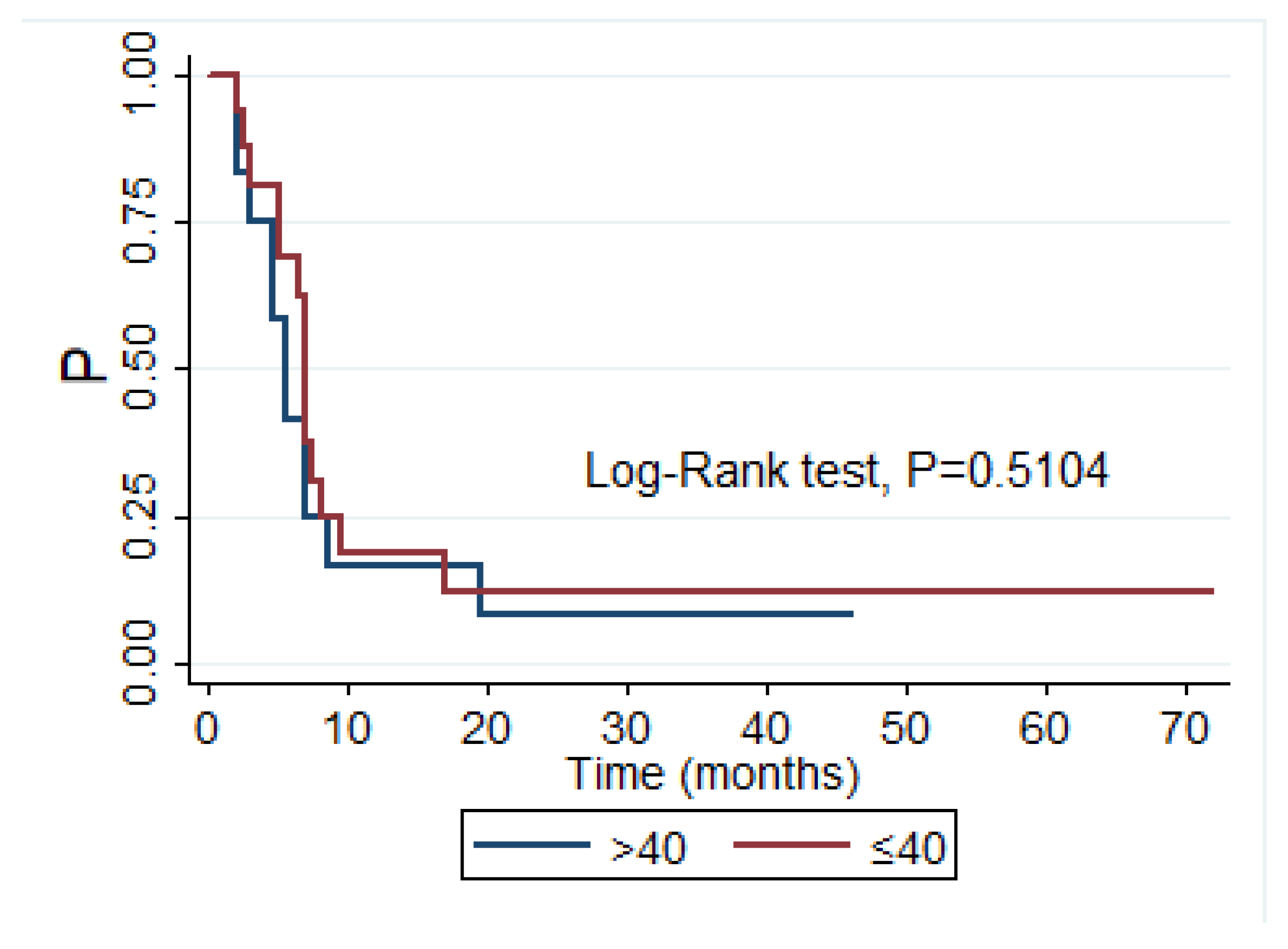
3.7. Correlation between Plasma Level of sHLA with Overall Survival in Whole Group of Glioma Patients and in the Subgroup of Glioblastoma Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carosella, E.D.; Rouas-Freiss, N.; Roux, D.T.; Moreau, P.; LeMaoult, J. HLA-G: An Immune Checkpoint Molecule. Adv. Immunol. 2015, 127, 33–144. [Google Scholar]
- Carosella, E.D.; Favier, B.; Rouas-Freiss, N.; Moreau, P.; Lemaoult, J. Beyond the increasing complexity of the immunomodulatory HLA-G molecule. Blood 2008, 111, 4862–4870. [Google Scholar] [CrossRef]
- Ferreira, L.M.R.; Meissner, T.B.; Tilburgs, T.; Strominger, J.L. HLA-G: At the interface of maternal-fetal tolerance. Trends Immunol. 2017, 38, 272–286. [Google Scholar] [CrossRef]
- Ishitani, A.; Geraghty, D.E. Alternative splicing of HLA-G transcripts yields proteins with primary structures resembling both class I and class II antigens. Proc. Natl. Acad. Sci. USA 1992, 89, 3947–3951. [Google Scholar] [CrossRef] [Green Version]
- Kirszenbaum, M.; Moreau, P.; Gluckman, E.; Dausset, J.; Carosella, E. An alternatively spliced form of HLA-G mRNA in human trophoblasts and evidence for the presence of HLA-G transcript in adult lymphocytes. Proc. Natl. Acad. Sci. USA 1994, 91, 4209–4213. [Google Scholar] [CrossRef] [Green Version]
- Donadi, E.A.; Castelli, E.C.; Arnaiz-Villena, A.; Roger, M.; Rey, D.; Moreau, P. Implications of the polymorphism of HLA-G on its function, regulation, evolution and disease association. Cell. Mol. Life Sci. 2011, 68, 369–395. [Google Scholar] [CrossRef] [Green Version]
- Kovats, S.; Main, E.K.; Librach, C.; Stubblebine, M.; Fisher, S.J.; DeMars, R. A class I antigen, HLA-G, expressed in human trophoblasts. Science 1990, 248, 220–223. [Google Scholar] [CrossRef]
- Loustau, M.; Anna, F.; Dréan, R.; Lecomte, M.; Langlade-Demoyen, P.; Caumartin, J. HLA-G Neo-Expression on Tumors. Front. Immunol. 2020, 11, 1685. [Google Scholar] [CrossRef]
- Contini, P.; Murdaca, G.; Puppo, F.; Negrini, S. HLA-G Expressing Immune Cells in Immune Mediated Diseases. Front. Immunol. 2020, 11, 1613. [Google Scholar] [CrossRef]
- Rizzo, R.; Bortolotti, D.; Bolzani, S.; Fainardi, E. HLA-G molecules in autoimmune diseases and infections. Front. Immunol. 2014, 5, 592. [Google Scholar] [CrossRef] [Green Version]
- Negrini, S.; Contini, P.; Pupo, F.; Greco, M.; Murdaca, G.; Puppo, F. Expression of membrane-bound human leucocyte antigen-G in systemic sclerosis and systemic lupus erythematosus. Hum. Immunol. 2020, 81, 162–167. [Google Scholar] [CrossRef]
- Bu, X.; Zhong, J.; Li, W.; Cai, S.; Gao, Y.; Ping, B. Immunomodulating functions of human leukocyte antigen-G and its role in graft-versus-host disease after allogeneic hematopoietic stem cell transplantation. Ann. Hematol. 2021, 100, 1391–1400. [Google Scholar] [CrossRef]
- Xu, C.X.; Zhang, Y.L.; Huang, X.Y.; Han, F.; Jin, Z.K.; Tian, P.X.; Dou, M. Prediction of acute renal allograft rejection by combined HLA-G 14-bp insertion/deletion genotype analysis and detection of kidney injury molecule-1 and osteopontin in the peripheral blood. Transpl. Immunol. 2021, 65, 101371. [Google Scholar] [CrossRef]
- Le Rond, S.; Le Maoult, J.; Creput, C.; Menier, C.; Deschamps, M.; Le Friec, G.; Amiot, L.; Durrbach, A.; Jean Dausset, J.; Carosella, E.D.; et al. Alloreactive CD4+ and CD8+ T cells express the immunotolerant HLA-G molecule in mixed lymphocyte reactions: In vivo implications in transplanted patients. Eur. J. Immunol. 2004, 34, 649–660. [Google Scholar] [CrossRef]
- Brugiere, O.; Thabut, G.; Krawice-Radanne, I.; Rizzo, R.; Dauriat, G.; Danel, C.; Suberbielle, C.; Mal, H.; Stern, M.; Schilte, C.; et al. Role of HLA-G as a predictive marker of low risk of chronic rejection in lung transplant recipients: A clinical prospective study. Am. J. Transplant. 2015, 15, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Amodio, G.; Sales de Albuquerque, R.; Gregori, S. New insights into HLA-G mediated tolerance. Tissue Antigens 2014, 84, 255–263. [Google Scholar] [CrossRef]
- Jaakola, A.; Roger, M.; Faucher, M.C.; Syrjänen, K.; Grénman, S.; Syrjänen, S.; Louvanto, K. HLA-G polymorphism impacts the outcome of oral HPV infections in women. BMC Infect. Dis. 2021, 21, 419. [Google Scholar] [CrossRef]
- Rashidi, S.; Farhadi, L.; Ghasemi, F.; Sheikhesmaeili, F.; Mohammadi, A. The potential role of HLA-G in the pathogenesis of HBV infection: Immunosuppressive or immunoprotective? Infect. Genet. Evol. 2020, 85, 104580. [Google Scholar] [CrossRef]
- Kaprio, T.; Sariola, H.; Linder, N.; Lundin, J.; Kere, J.; Haglund, C.; Wedenoja, S. HLA-G expression correlates with histological grade but not with prognosis in colorectal carcinoma. HLA 2021, 98, 213–217. [Google Scholar] [CrossRef]
- Rouas-Freiss, N.; Moreau, P.; LeMaoult, J.; Carosella, E.D. The dual role of HLA-G in cancer. J. Immunol. Res. 2014, 2014, 359748. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.H. HLA-G expression in cancers: Potential role in diagnosis, prognosis and therapy. Endocr. Metab. Immune Disord. Drug Targets 2011, 11, 76–89. [Google Scholar] [CrossRef]
- Lin, A.; Zhang, X.; Xu, H.H.; Xu, D.P.; Ruan, Y.Y.; Yan, W.H. HLA-G expression is associated with metastasis and poor survival in the Balb/c nu/nu murine tumor model with ovarian cancer. Int. J. Cancer 2012, 131, 150–157. [Google Scholar] [CrossRef] [Green Version]
- Urosevic, M.; Dummer, R. Human Leukocyte Antigen–G and Cancer Immunoediting. Cancer Res. 2008, 68, 627–630. [Google Scholar] [CrossRef] [Green Version]
- Dhatchinamoorthy, K.; Colbert, J.D.; Rock, K.L. Cancer Immune Evasion Through Loss of MHC Class I Antigen Presentation. Front. Immunol. 2021, 12, 636568. [Google Scholar] [CrossRef]
- Ścieżyńska, A.; Komorowski, M.; Soszyńska, M.; Malejczyk, J. NK Cells as Potential Targets for Immunotherapy in Endometriosis. J. Clin. Med. 2019, 8, 1468. [Google Scholar] [CrossRef] [Green Version]
- Llano, M.; Lee, N.; Navarro, F.; Garcia, P.; Albar, J.P.; Geraghty, D.E.; Lopez-Botet, M. HLA-E-bound peptides influence recognition by inhibitory and triggering CD94/NKG2 receptors: Preferential response to an HLA-G-derived nonamer. Eur. J. Immunol. 1998, 28, 2854–2863. [Google Scholar] [CrossRef]
- LeMaoult, J.; Zafaranloo, K.; Le Danff, C.; Carosella, E.D. HLA-G up-regulates ILT2, ILT3, ILT4, and KIR2DL4 in antigen presenting cells, NK cells, and T cells. FASEB J. 2005, 6, 662–664. [Google Scholar] [CrossRef]
- Chen, B.G.; Xu, D.P.; Lin, A.; Yan, W.H. NK cytolysis is dependent on the proportion of HLA-G expression. Hum. Immunol. 2013, 74, 286–289. [Google Scholar] [CrossRef]
- Riteau, B.; Menier, C.; Khalil-Daher, I.; Martinozzi, S.; Pla, M.; Dausset, J.; Carosella, E.D.; Rouas-Freiss, N. HLA-G1 co-expression boosts the HLA class I-mediated NK lysis inhibition. Int. Immunol. 2001, 13, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Kapasi, K.; Albert, S.E.; Yie, S.; Zavazava, N.; Librach, C.L. HLA-G has a concentration-dependent effect on the generation of anallo-CTL response. Immunology 2000, 101, 191–200. [Google Scholar] [CrossRef]
- Naji, A.; Menier, C.; Morandi, F.; Agaugue, S.; Maki, G.; Ferretti, E.; Bruel, S.; Pistoia, V.; Carosella, E.D.; Rouas-Freiss, N. Binding of HLA-G to ITIM-bearing Ig-like transcript 2 receptor suppresses B cell responses. J. Immunol. 2014, 192, 1536–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naji, A.; Le Rond, S.; Durrbach, A.; Krawice-Radanne, I.; Creput, C.; Daouya, M.; Caumartin, J.; LeMaoult, J.; Carosella, E.D.; Rouas-Freiss, N. CD3+CD4 low and CD3+CD8 low are induced by HLA-G: Novel human peripheral blood suppressor T-cell subsets involved in transplant acceptance. Blood 2007, 110, 3936–3948. [Google Scholar] [CrossRef] [PubMed]
- Dumont, C.; Jacquier, A.; Verine, J.; Noel, F.; Goujon, A.; Wu, C.L.; Hung, T.M.; Desgrandchamps, F.; Culine, S.; Carosella, E.D.; et al. CD8+PD-1-ILT2+ T Cells Are an Intratumoral Cytotoxic Population Selectively Inhibited by the Immune-Checkpoint HLA-G. Cancer. Immunol. Res. 2019, 7, 1619–1632. [Google Scholar] [CrossRef] [PubMed]
- LeMaoult, J.; Krawice-Radanne, I.; Dausset, J.; Carosella, E.D. HLA-G1-expressing antigen-presenting cells induce immunosuppressive CD4+T cells. Proc. Natl. Acad. Sci. USA 2004, 101, 7064–7069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horuzsko, A.; Lenfant, F.; Munn, D.H.; Mellor, A.L. Maturation of antigen-presenting cells is compromised in HLA-G transgenicmice. Int. Immunol. 2001, 13, 385–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.; Wang, L.; Han, Y.; Gao, W.; Wei, X.; Gong, R.; Zhu, M.; Sun, Y.; Yu, S. Immunoglobulin like transcript 4 and human leukocyte antigen G interaction promotes the progression of human colorectal cancer. Int. J. Oncol. 2019, 54, 1943–1954. [Google Scholar] [CrossRef] [Green Version]
- Contini, P.; Ghio, M.; Poggi, A.; Filaci, G.; Indiveri, F.; Ferrone, S.; Puppo, F. Soluble HLA-F-A,-B,-C and -G molecules induce apoptosis in T and NK CD8+ cells and inhibit cytotoxic T cell activity through CD8 ligation. Eur. J. Immunol. 2003, 33, 125–134. [Google Scholar] [CrossRef]
- Lindaman, A.; Dowden, A.; Zavazava, N. Soluble HLA-G molecules induce apoptosis in natural killer cells. Am. J. Reprod. Immunol. 2006, 56, 68–76. [Google Scholar] [CrossRef]
- Ajith, A.; Portik-Dobos, V.; Nguyen-Lefebvre, A.T.; Callaway, C.; Horuzsko, D.D.; Kapoor, R.; Zayas, C.; Maenaka, K.; Mulloy, L.L.; Horuzsko, A. HLA-G dimer targets Granzyme B pathway to prolong human renal allograft survival. FASEB J. 2019, 33, 5220–5236. [Google Scholar] [CrossRef]
- Bainbridge, D.R.; Ellis, S.A.; Sargent, I.L. HLA-G suppresses proliferation of CD4 (+) T-lymphocytes. J. Reprod. Immunol. 2000, 48, 17–26. [Google Scholar] [CrossRef]
- Amodio, G.; Gregori, S. Human tolerogenic DC-10: Perspectives for clinical applications. Transpl. Res. 2012, 1, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregori, S.; Tomasoni, D.; Pacciani, V.; Scirpoli, M.; Battaglia, M.; Magnani, C.F.; Hauben, E.; Roncarolo, M.G. Dierentiation of type 1 T regulatory cells (Tr1) by tolerogenic DC-10 requires the IL-10-dependent ILT4/HLA-G pathway. Blood 2010, 116, 935–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, A.; Yan, W.H. Intercellular transfer of HLA-G: Its potential in cancer immunology. Clin. Transl. Immunol. 2019, 8, e1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiot, L.; Ferrone, S.; Grosse-Wilde, H.; Seliger, B. Biology of HLA-G in cancer: A candidate molecule for therapeutic intervention? Cell. Mol. Life Sci. 2011, 68, 417–431. [Google Scholar] [CrossRef] [Green Version]
- Wastowski, I.J.; Simões, R.T.; Yaghi, L.; Donadi, E.A.; Pancoto, J.T.; Poras, I.; Lechapt-Zalcman, E.; Bernaudin, M.; Valable, S.; Carlotti, C.G., Jr.; et al. Human leukocyte antigen-G is frequently expressed in glioblastoma and may be induced in vitro by combined 5-aza-2′-deoxycytidine and interferon-γ treatments: Results from a multicentric study. Am. J. Pathol. 2013, 182, 540–552. [Google Scholar] [CrossRef]
- Krijgsman, D.; Roelands, J.; Hendrickx, W.; Bedognetti, D.; Kuppen, P.J.K. HLA-G: A New Immune Checkpoint in Cancer? Editorial. Int. J. Mol. Sci. 2020, 21, 4528. [Google Scholar] [CrossRef]
- Hviid, T.V.; Rizzo, R.; Melchiorri, L.; Stignani, M.; Baricordi, O.R. Polymorphism in the 5′ upstream regulatory and 3′ untranslated regions of the HLA-G gene in relation to soluble HLA-G and IL-10 expression. Hum. Immunol. 2006, 67, 53–62. [Google Scholar] [CrossRef]
- Hviid, T.V.; Hylenius, S.; Hoegh, A.M.; Kruse, C.; Christiansen, O.B. HLA-G polymorphisms in couples with recurrent spontaneous abortions. Tissue Antigens 2002, 60, 122–132. [Google Scholar] [CrossRef]
- Rousseau, P.; Le Discorde, M.; Mouillot, G.; Marcou, C.; Carosella, E.D.; Moreau, P. The 14bp deletion insertion polymorphism in the 3′ UT region of the HLA-G gene influences HLA-G mRNA stability. Hum. Immunol. 2003, 64, 1005–1010. [Google Scholar] [CrossRef]
- Chun, S.J.; Park, S.H.; Park, C.K.; Kim, J.W.; Kim, T.M.; Choi, S.H.; Lee, S.T.; Kim, I.H. Survival gain with re-Op/RT for recurred high-grade gliomas depends upon risk groups. Radiother. Oncol. 2018, 128, 254–259. [Google Scholar] [CrossRef]
- Liang, R.; Chen, N.; Li, M.; Wang, X.; Mao, Q.; Liu, Y. Significance of systemic immune-inflammation index in the differential diagnosis of high- and low-grade gliomas. Clin. Neurol. Neurosurg. 2018, 164, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Anast, J.; Bassett, W.; Kawakami, T.; Sakuragi, N.; Dahiya, R. Bisulfite conversion-specific and methylation-specific PCR: A sensitive technique for accurate evaluation of CpG methylation. Biochem. Biophys. Res. Commun. 2003, 309, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.snpstats.net/start.htm (accessed on 20 February 2018).
- Lin, A.; Yan, W.H. Heterogeneity of HLA-G expression in cancers: Facing the Challenges. Front. Immunol. 2018, 9, 2164. [Google Scholar] [CrossRef]
- Wagner, S.N.; Rebmann, V.; Willers, C.P.; Grosse-Wilde, H.; Goos, M. Expression analysis of classic and non-classic HLA molecules before interferon alfa-2b treatment of melanoma. Lancet 2000, 56, 220–221. [Google Scholar] [CrossRef]
- Boujelbene, N.; Ben, Y.H.; Babay, W.; Gadria, S.; Zemni, I.; Azaiez, H.; Dhouioui, S.; Zidi, N.; Mchiri, R.; Mrad, K.; et al. HLA-G, HLA-E, and IDO overexpression predicts a worse survival of Tunisian patients with vulvar squamous cell carcinoma. HLA 2019, 94, 11–24. [Google Scholar] [CrossRef]
- Imani, R.; Seyedmajidi, M.; Ghasemi, N.; Moslemi, D.; Shafaee, S.; Bijani, A. HLA-G Expression is Associated with an Unfavorable Prognosis of Oral Squamous Cell Carcinoma. Asian Pac. J. Cancer Prev. 2018, 19, 2527–2533. [Google Scholar]
- De Kruijf, E.M.; Sajet, A.; van Nes, J.G.; Natanov, R.; Putter, H.; Smit, V.T.; Liefers, G.J.; van den Elsen, P.J.; van de Velde, C.J.; Kuppen, P.J. HLA-E and HLA-G expression in classical HLA class I-negative tumors is of prognostic value for clinical outcome of early breast cancer patients. J. Immunol. 2010, 185, 7452–7459. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.Y.; Lv, Y.G.; Wang, L.; Shi, S.J.; Yang, F.; Zheng, G.X.; Wen, W.H.; Yang, A.G. Predictive value of HLA-G and HLA-E in the prognosis of colorectal cancer patients. Cell. Immunol. 2015, 293, 10–16. [Google Scholar] [CrossRef]
- Durmanova, V.; Kluckova, K.; Filova, B.; Minarik, G.; Kozak, J.; Rychly, B.; Svajdler, M.; Matejcik, V.; Steno, J.; Bucova, M. HLA-G 5’URR regulatory polymorphisms are associated with the risk of developing gliomas. Int. J. Neurosci. 2021, 1–10, online ahead of print. [Google Scholar] [CrossRef]
- de Magalhães, K.C.S.F.; Silva, K.R.; Gomes, N.A.; Sadissou, I.; Carvalho, G.T.; Buzellin, M.A.; Tafuri, L.S.; Nunes, C.B.; Nunes, M.B.; Donadi, E.A.; et al. HLA-G 14bp In/Del and +3142 C/G Genotypes Are Differentially Expressed Between Patients with Grade IV Gliomas and Controls. Int. J. Neurosci. 2021, 131, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Lau, D.T.; Norris, M.D.; Marshall, G.M.; Haber, M.; Ashton, L.J. HLA-G polymorphisms, genetic susceptibility, and clinical outcome in childhood neuroblastoma. Tissue Antigens 2011, 78, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, R.; Ramanakumar, A.V.; Koushik, A.; Coutlée, F.; Franco, E.; Roger, M. Human Leukocyte Antigen G Polymorphism Is Associated with an Increased Risk of Invasive Cancer of the Uterine Cervix. Int. J. Cancer 2012, 131, E312–E319. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.Z.; Ge, Q.; Li, M.H.; Shi, G.M.; Xu, X.; Xu, L.W.; Xu, Z.; Lu, T.Z.; Wu, R.; Zhou, L.H.; et al. Association between human leukocyte antigen-G 14-bp insertion/deletion polymorphism and cancer risk: A meta-analysis and systematic review. Hum. Immunol. 2014, 75, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, S.; Jia, S.; Zhu, Z.; Gao, X.; Dong, D.; Gao, Y. Association of HLA-G 3′ UTR 14-bp insertion/deletion polymorphism with hepatocellular carcinoma susceptibility in a Chinese population. DNA Cell Biol. 2011, 30, 1027–1032. [Google Scholar] [CrossRef]
- Eskandari-Nasab, E.; Hashemi, M.; Hasani, S.S.; Omrani, M.; Taheri, M.; Mashhadi, M.A. Association between HLA-G 3’UTR 14-bp ins/del polymorphism and susceptibility to breast cancer. Cancer Biomark. 2013, 13, 253–259. [Google Scholar] [CrossRef]
- Li, T.; Huang, H.; Liao, D.; Ling, H.; Su, B.; Cai, M. Genetic Polymorphism in HLA-G 3’UTR 14-bp Ins/Del and Risk of Cancer: A Meta-Analysis of Case-Control Study. Mol. Genet. Genom. 2015, 290, 1235–1245. [Google Scholar] [CrossRef]
- Fan, X.; Wang, Y.; Zhang, C.; Liu, X.; Qian, Z.; Jiang, T.J. Human leukocyte antigen-G overexpression predicts poor clinical outcomes in low-grade gliomas. Neuroimmunology 2016, 294, 27–31. [Google Scholar] [CrossRef]
- Li, Y.; Patel, S.P.; Roszik, J.; Qin, Y. Hypoxia-Driven Immunosuppressive Metabolites in the Tumor Microenvironment: New Approaches for Combinational Immunotherapy. Front. Immunol. 2018, 9, 1591. [Google Scholar] [CrossRef] [Green Version]
- Ziliotto, M.; Rodrigues, R.M.; Bogo, A.J.C. Controlled hypobaric hypoxia increases immunological tolerance by modifying HLA-G expression, a potential therapy to inflammatory diseases. Med. Hypotheses 2020, 140, 109664. [Google Scholar] [CrossRef]
- Gupta, S.K.; Kizilbash, S.H.; Carlson, B.L.; Mladek, A.C.; Boakye-Agyeman, F.; Bakken, K.K.; Pokorny, J.L.; Schroeder, M.A.; Decker, P.A.; Cen, L.; et al. Delineation of MGMT Hypermethylation as a Biomarker for Veliparib-Mediated Temozolomide-Sensitizing Therapy of Glioblastoma. J. Natl. Cancer Inst. 2015, 108, djv369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konig, L.; Kasimir-Bauer, S.; Hoffmann, O.; Bittner, A.K.; Wagner, B.; Manvailer, L.F.; Sabine Schramm, S.; Bankfalvi, A.; Giebel, B.; Rainer Kimmig, R.; et al. The prognostic impact of soluble and vesicular HLA-G and its relationship to circulating tumor cells in neoadjuvant treated breast cancer patients. Hum. Immunol. 2016, 77, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Ben, A.A.; Beauchemin, K.; Faucher, M.C.; Hamzaoui, A.; Hamzaoui, K.; Roger, M. Human Leukocyte Antigen G Polymorphism and Expression Are Associated with an Increased Risk of Non-Small-Cell Lung Cancer and Advanced Disease Stage. PLoS ONE 2016, 11, e0161210. [Google Scholar]
- Ben, Y.H.; Babay, W.; Bortolotti, D.; Boujelbene, N.; Laaribi, A.B.; Zidi, N.; Kehila, M.; Chelbi, H.; Boudabous, A.; Mrad, K.; et al. Increased plasmatic soluble HLA-G levels in endometrial cancer. Mol. Immunol. 2018, 99, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Lázaro-Sánchez, A.D.; Salces-Ortiz, P.; Velásquez, L.I.; Orozco-Beltrán, D.; Díaz-Fernández, N.; Juárez-Marroquí, A. HLA-G as a new tumor biomarker: Detection of soluble isoforms of HLA-G in the serum and saliva of patients with colorectal cancer. Clin. Transl. Oncol. 2020, 22, 1166–1171. [Google Scholar] [CrossRef]
- Kirana, C.; Ruszkiewicz, A.; Stubbs, R.S.; Hardingham, J.E.; Hewett, P.J.; Maddern, G.J.; Hauben, E. Soluble HLA-G is a differential prognostic marker in sequential colorectal cancer disease stages. Int. J. Cancer 2017, 140, 2577–2586. [Google Scholar] [CrossRef]
- Hasanah, N.; Mintaroem, K.; Fitri, L.E.; Noorhamdani, N. Interleukin 10 Induces the Expression of Membrane-Bound HLA- G and the Production of Soluble HLA-G on HeLa CCL-2 Cells. Open Access Maced. J. Med. Sci. 2019, 7, 3554–3558. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Parameter (Mean ± SD) | Brain Gliomas N = 59 |
|---|---|
| Age at diagnosis | 53.36 |
| Sex (women/men) | 25/34 |
| Grade of gliomas | |
| Grade II | 19 |
| Grade III | 11 |
| Grade IV | 29 |
| Primary diagnosis of glioma | 49 |
| Relapse | 10 |
| Allele/Genotype | Brain Gliomas (N = 59) | Controls (N = 159) | Univariate Analysis | Multivariate Analysis | ||
|---|---|---|---|---|---|---|
| p | OR (95% CI) | p | OR (95% CI) | |||
| −14 | 58 (49.15%) | 191 (60.06%) | ||||
| +14 | 60 (50.85%) | 127 (39.94%) | 0.05 | 1.56 (1.02–2.38) | - | - |
| −14/−14 | 12 (20.34%) | 55 (34.59%) | 1.00 | 1.00 | ||
| −14/+14 | 34 (57.63%) | 81 (50.94%) | 0.09 | 1.92 (0.92–4.04) | 0.35 | 1.64 (0.72–3.72) |
| +14/+14 | 13 (22.03%) | 23 (14.47%) | 2.59 (1.03–6.52) | 2.00 (0.71–5.65) | ||
| −14/−14 | 12 (20.34%) | 55 (34.59%) | 1.00 | 1.00 | ||
| −14/+14 and +14/+14 | 47 (79.66%) | 104 (65.41%) | 0.03 | 2.07 (1.02–4.23) | 0.17 | 1.72 (0.78–3.78) |
| −14/−14 and −14/+14 | 46 (77.97%) | 136 (85.53%) | 1.00 | 1.00 | ||
| +14/+14 | 13 (22.03%) | 23 (14.47%) | 0.19 | 1.67 (0.78–3.57) | 0.43 | 1.43 (0.60–3.40) |
| −14/−14 and +14/+14 | 25 (42.37%) | 78 (49.06%) | 1.00 | 1.00 | ||
| −14/+14 | 34 (57.63%) | 81 (50.94%) | 0.38 | 1.31 (0.72–2.39) | 0.54 | 1.24 (0.63–2.44) |
| Patients (N = 59) | Healthy Controls (N = 43) | p (Student’s t-Test) | |
|---|---|---|---|
| sHLA-G (U/mL) (Mean ± SD) | 42.17 ± 38.50 | 23.06 ± 9.53 | 0.048 |
| Grade 2 N = 13 | Grade 3 N = 9 | Grade 4 N = 27 | p (2 vs. 3) | p (2 vs. 4) | p (3 vs. 4) | |
|---|---|---|---|---|---|---|
| sHLA-G (U/mL) (Mean ± SD) | 39.19 ± 40.96 | 32.05 ± 13.69 | 43.23 ± 39.42 | 0.45 | 0.22 | 0.70 |
| Allele/Genotype | sHLA-G (U/mL) Mean ± SD | p |
|---|---|---|
| A. −14 (n = 58) | 44.586 ± 38.34 | |
| B. +14 (n = 60) | 37.942 ± 29.81 | 0.499 (A vs. B) |
| C. −14/−14 (n = 12) | 45.76 ± 36.17 | 0.395 (C vs. D) |
| D. −14/+14 (n = 34) | 41.06 ± 35.77 | 0.468 (D vs. E) |
| E. +14/+14 (n = 13) | 48.56 ± 39.06 | 0.957 (C vs. E) |
| MGMT Promoter | Patients (N = 32) | sHLA-G (U/mL) (Mean ± SD) | p |
|---|---|---|---|
| methylated | 17 | 29.51 ± 23.50 | 0.03 |
| unmethylated | 15 | 54.30 ± 43.12 |
| Patients (N = 32) | Correlation of sHLA-G with IL-6 | Correlation of sHLA-G with IL-10 | Correlation of sHLA-G with IL-10/IL-6 Ratio |
|---|---|---|---|
| Spearman r | −0.584 | 0.208 | 0.622 |
| 95% CI | −0.779–(−0.286) | −0.527–0.163 | −0.0034–0.633 |
| p | 0.0004 | 0.26 | 0.046 |
| Parameter | Parameter Estimate | Standard Error | Chi-Square | p | Hazard Ratio | |
|---|---|---|---|---|---|---|
| sHLA-G in G. II (n = 19) | Survival time (months) | 0.02234 | 0.00852 | 6.8696 | 0.0088 | 1.023 |
| sHLA-G in G. IV (n = 29) | Survival time (months) | 0.00435 | 0.00212 | 4.2222 | 0.0399 | 1.004 |
| sHLA-G (U/mL) | Pts. Surviving < 1 Year N = 21 | Pts. Surviving > 1 Year N = 8 |
|---|---|---|
| Median | 46.74 | 21.50 |
| IQR | 74.80 | 12.60 |
| 95% CI | 30.02–119.10 | 14.59–30.17 |
| p | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bucova, M.; Kluckova, K.; Kozak, J.; Rychly, B.; Suchankova, M.; Svajdler, M.; Matejcik, V.; Steno, J.; Zsemlye, E.; Durmanova, V. HLA-G 14bp Ins/Del Polymorphism, Plasma Level of Soluble HLA-G, and Association with IL-6/IL-10 Ratio and Survival of Glioma Patients. Diagnostics 2022, 12, 1099. https://doi.org/10.3390/diagnostics12051099
Bucova M, Kluckova K, Kozak J, Rychly B, Suchankova M, Svajdler M, Matejcik V, Steno J, Zsemlye E, Durmanova V. HLA-G 14bp Ins/Del Polymorphism, Plasma Level of Soluble HLA-G, and Association with IL-6/IL-10 Ratio and Survival of Glioma Patients. Diagnostics. 2022; 12(5):1099. https://doi.org/10.3390/diagnostics12051099
Chicago/Turabian StyleBucova, Maria, Kristina Kluckova, Jan Kozak, Boris Rychly, Magda Suchankova, Marian Svajdler, Viktor Matejcik, Juraj Steno, Eszter Zsemlye, and Vladimira Durmanova. 2022. "HLA-G 14bp Ins/Del Polymorphism, Plasma Level of Soluble HLA-G, and Association with IL-6/IL-10 Ratio and Survival of Glioma Patients" Diagnostics 12, no. 5: 1099. https://doi.org/10.3390/diagnostics12051099