
Alpha-Enolase (ENO1) Correlates with Invasiveness of Cutaneous Melanoma—An In Vitro and a Clinical Study

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Lysis
2.3. Western Blotting
2.4. Immunofluorescence
2.5. Enolase Activity Assay
2.6. Immunohistochemistry
2.7. Patients
2.8. Statistical Analysis
3. Results
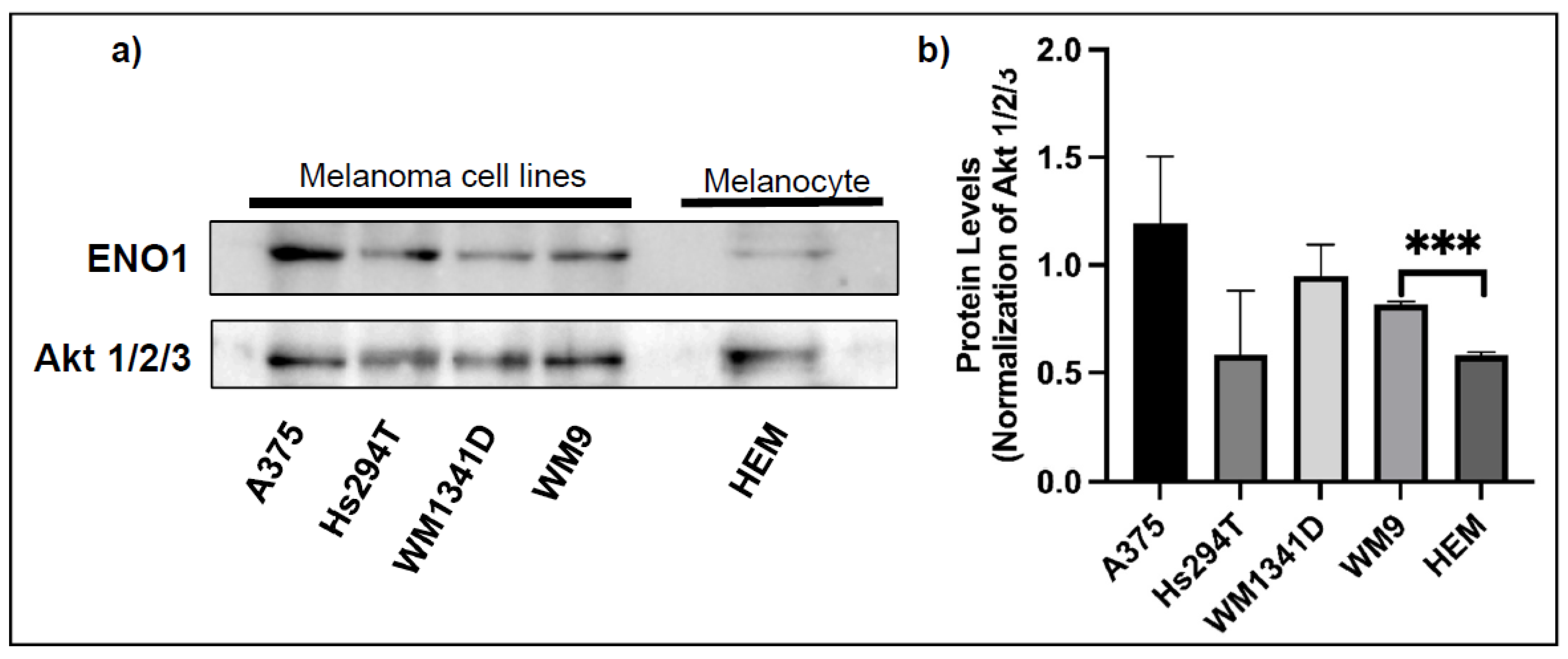
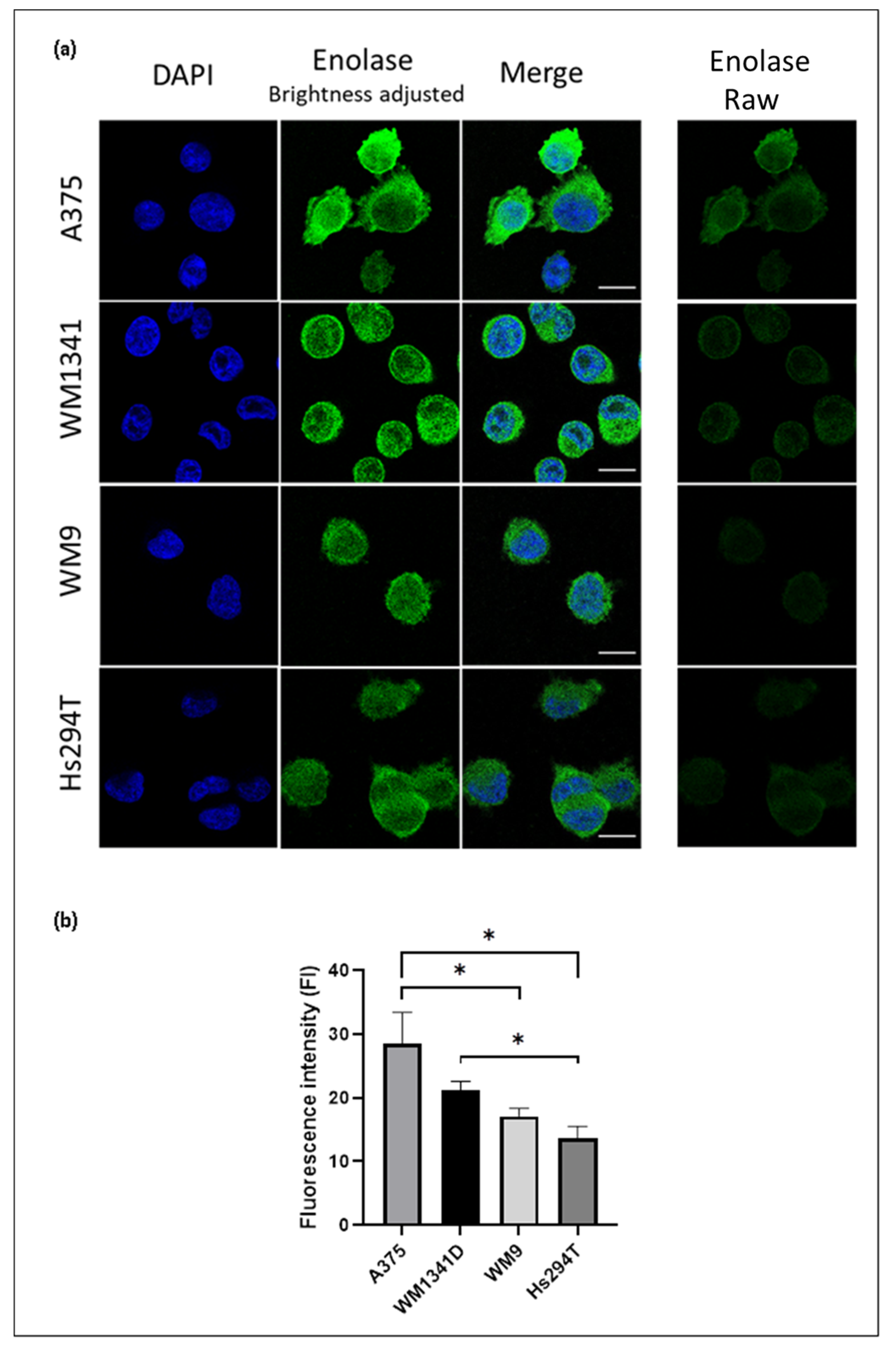
3.1. Expression of ENO1 in Melanoma Cell Lines
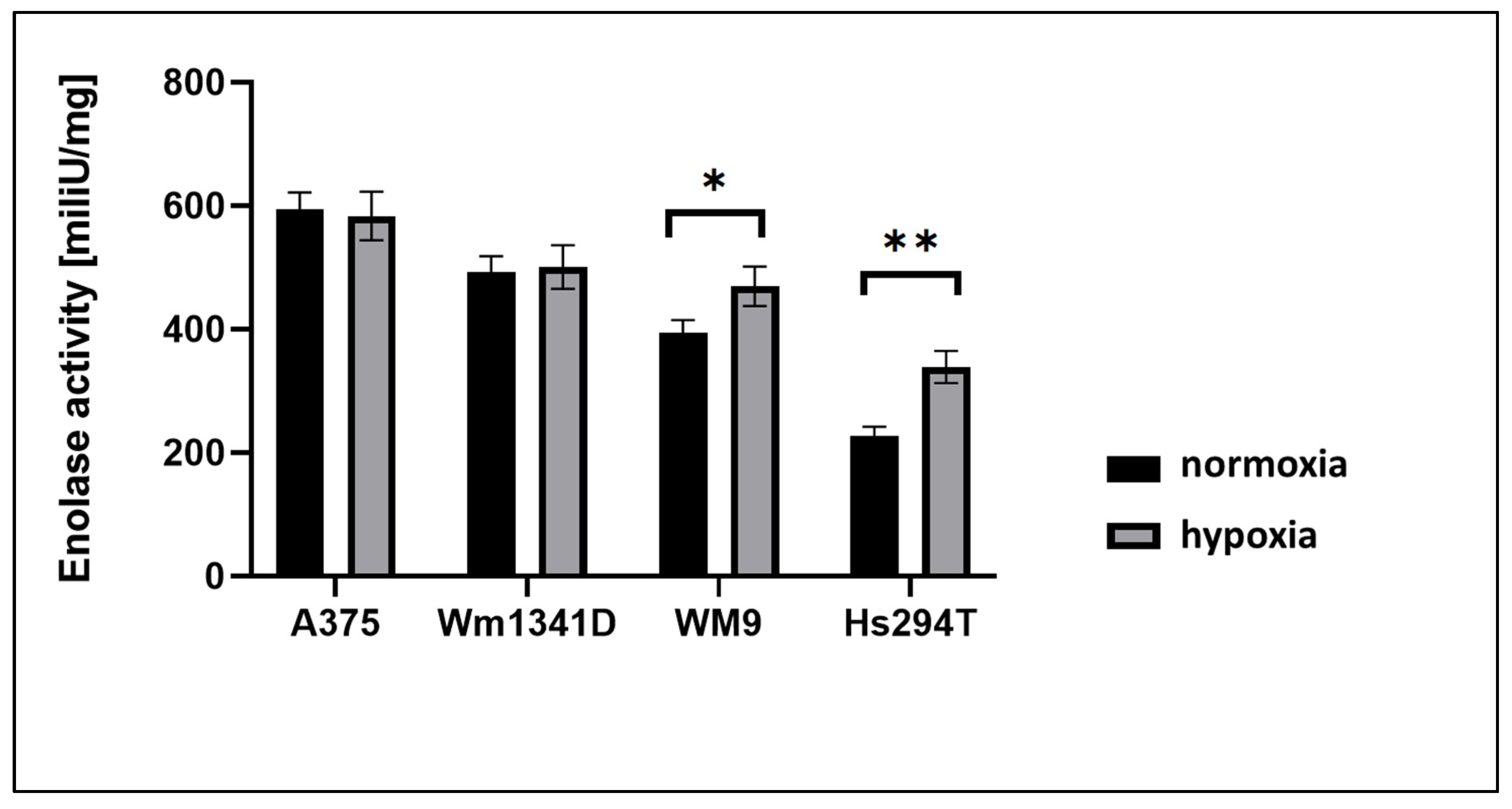
3.2. ENO1 Enzymatic Activity in Melanoma Cell Lines
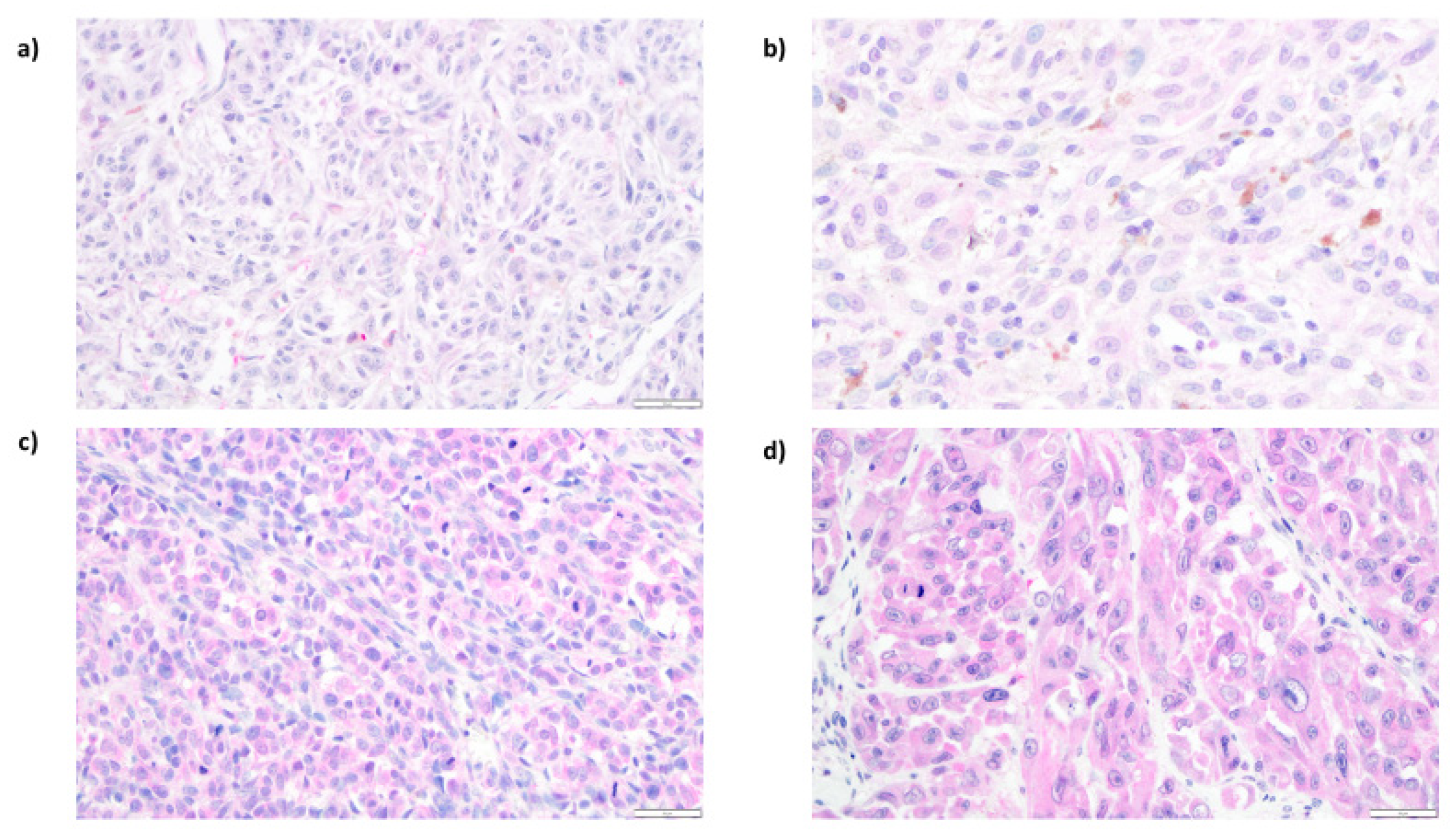
3.3. Expression of ENO1 in Cutaneous Melanoma Patients
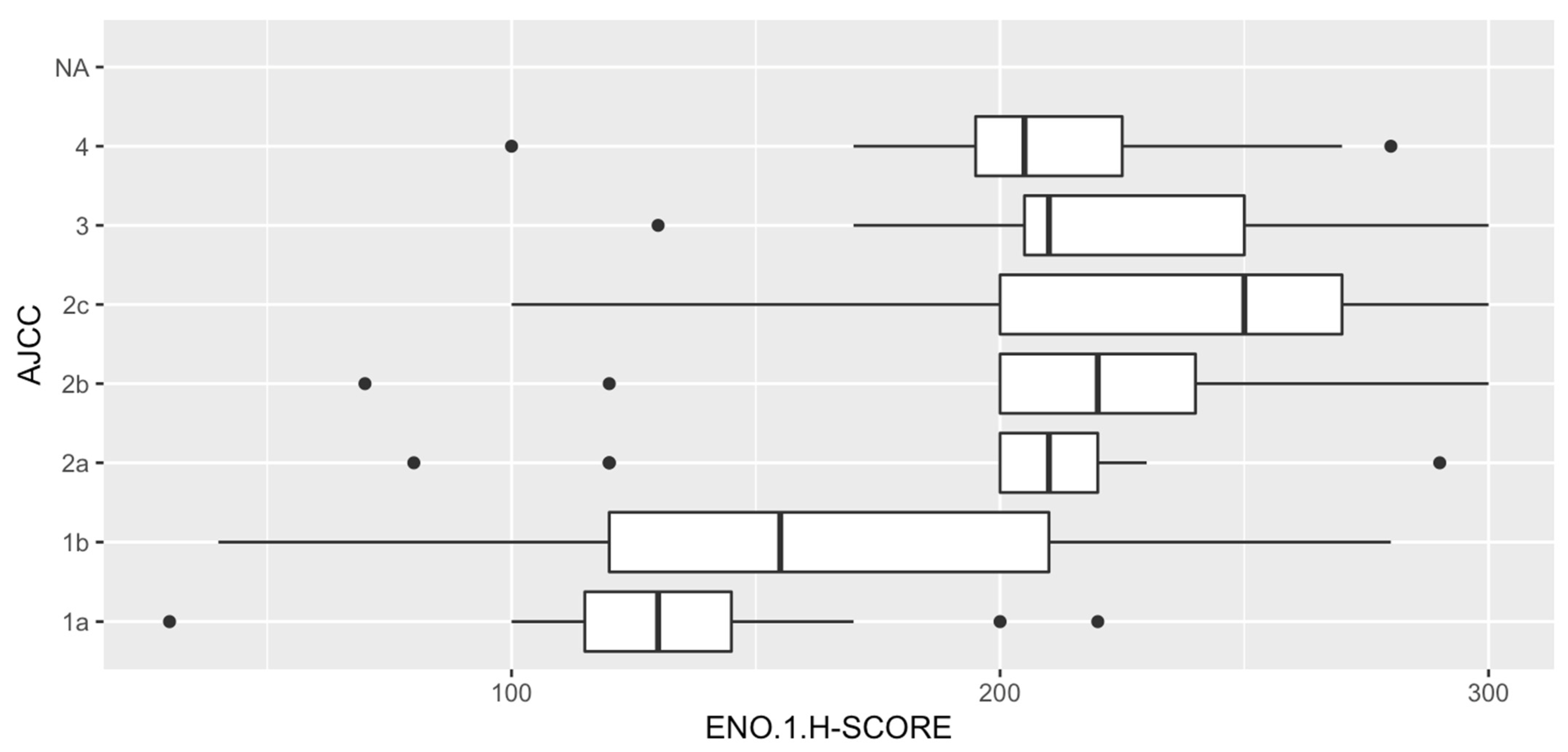
3.4. Analysis of Correlations between ENO1 Expression and Clinical Parameters
3.5. Analysis of Correlations between ENO1 Expression and Histopathologic Parameters of Primary Tumors
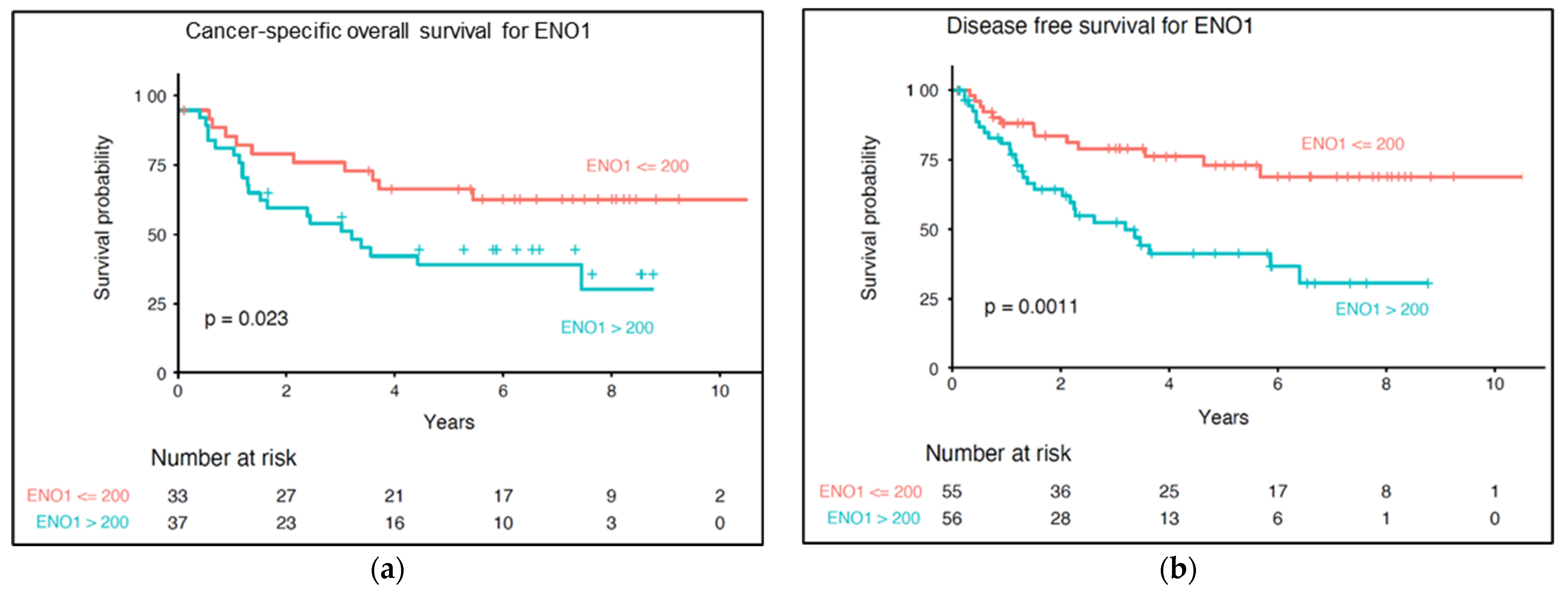
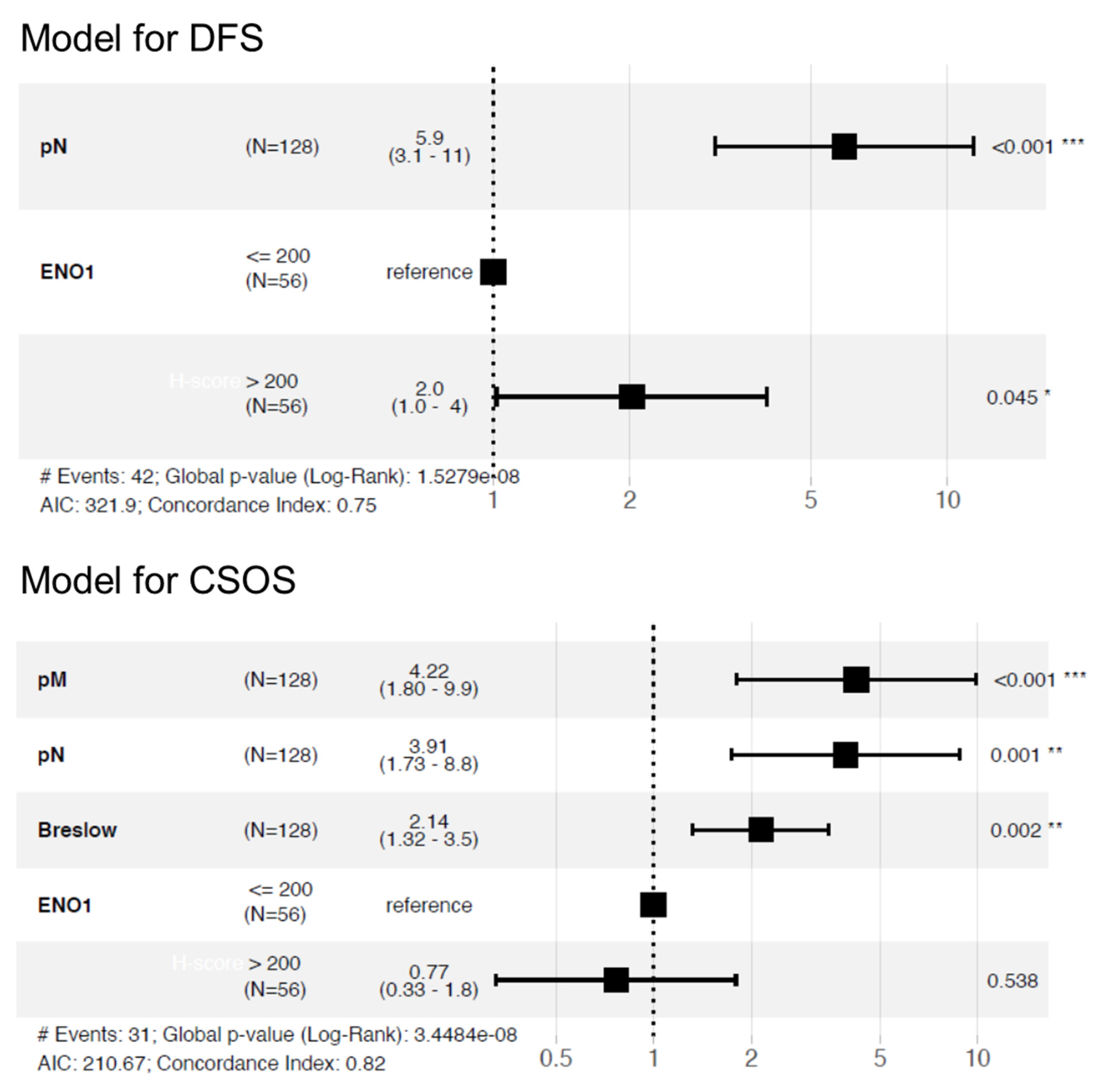
3.6. Impact of ENO1 Expression of Long-Term Prognosis of Cutaneous Melanoma Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rastrelli, M.; Tropea, S.; Rossi, C.R.; Alaibac, M. Melanoma: Epidemiology, Risk Factors, Pathogenesis, Diagnosis and Classification. Vivo 2014, 28, 1005–1011. [Google Scholar]
- Robsahm, T.E.; Bergva, G.; Hestvik, U.E.; Møller, B. Sex Differences in Rising Trends of Cutaneous Malignant Melanoma in Norway, 1954–2008. Melanoma Res. 2013, 23, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Minini, R.; Rohrmann, S.; Braun, R.; Korol, D.; Dehler, S. Incidence Trends and Clinical–Pathological Characteristics of Invasive Cutaneous Melanoma from 1980 to 2010 in the Canton of Zurich, Switzerland. Melanoma Res. 2017, 27, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conforti, C.; Zalaudek, I. Epidemiology and Risk Factors of Melanoma: A Review. Dermatol Pract. Concept. 2021, 11 (Suppl. 1), e2021161S. Available online: https://dpcj.org/index.php/dpc/article/view/dermatol-pract-concept-articleid-dp11S1a161S (accessed on 12 March 2021). [CrossRef] [PubMed]
- Lucas, R.M.; Norval, M.; Neale, R.E.; Young, A.R.; de Gruijl, F.R.; Takizawa, Y.; van der Leun, J.C. The Consequences for Human Health of Stratospheric Ozone Depletion in Association with Other Environmental Factors. Photochem. Photobiol. Sci. 2015, 14, 53–87. [Google Scholar] [CrossRef] [Green Version]
- Long, G.V.; Stroyakovskiy, D.; Gogas, H.; Levchenko, E.; de Braud, F.; Larkin, J.; Garbe, C.; Jouary, T.; Hauschild, A.; Grob, J.J.; et al. Combined BRAF and MEK Inhibition versus BRAF Inhibition Alone in Melanoma. N. Engl. J. Med. 2014, 371, 1877–1888. [Google Scholar] [CrossRef] [Green Version]
- Shoushtari, A.N.; Munhoz, R.R.; Kuk, D.; Ott, P.A.; Johnson, D.B.; Tsai, K.K.; Rapisuwon, S.; Eroglu, Z.; Sullivan, R.J.; Luke, J.J.; et al. The Efficacy of Anti-PD-1 Agents in Acral and Mucosal Melanoma. Cancer 2016, 122, 3354–3362. [Google Scholar] [CrossRef] [Green Version]
- Algazi, A.P.; Tsai, K.K.; Shoushtari, A.N.; Munhoz, R.R.; Eroglu, Z.; Piulats, J.M.; Ott, P.A.; Johnson, D.B.; Hwang, J.; Daud, A.I.; et al. Clinical Outcomes in Metastatic Uveal Melanoma Treated with PD-1 and PD-L1 Antibodies. Cancer 2016, 122, 3344–3353. [Google Scholar] [CrossRef]
- Capello, M.; Ferri-Borgogno, S.; Cappello, P.; Novelli, F. α-Enolase: A Promising Therapeutic and Diagnostic Tumor Target. FEBS J. 2011, 278, 1064–1074. [Google Scholar] [CrossRef] [Green Version]
- Didiasova, M.; Schaefer, L.; Wygrecka, M. When Place Matters: Shuttling of Enolase-1 Across Cellular Compartments. Front. Cell Dev. Biol. 2019, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Perconti, G.; Pratesi, F.; Angelotti, F.; Manca, L.; Puxeddu, I.; Rubino, P.; Maranto, C.; Giallongo, A.; Migliorini, P. Fingerprinting of Anti-Alpha Enolase Antibodies in Systemic Sclerosis. Clin. Exp. Rheumatol. 2020, 38 (Suppl. 1), 115–119. [Google Scholar] [PubMed]
- Li, M.; Li, J.; Wang, J.; Li, Y.; Yang, P. Serum Level of Anti-α-Enolase Antibody in Untreated Systemic Lupus Erythematosus Patients Correlates with 24-Hour Urine Protein and D-Dimer. Lupus 2018, 27, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Um, J.; Lee, J.-H.; Kim, W.-H.; Kang, W.S.; Kim, S.H.; Ha, H.-H.; Kim, Y.-C.; Ahn, Y.-K.; Jung, D.-W.; et al. ENOblock, a Unique Small Molecule Inhibitor of the Non-Glycolytic Functions of Enolase, Alleviates the Symptoms of Type 2 Diabetes. Sci. Rep. 2017, 7, 44186. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Lange, M.L.B. Multifunctional Roles of Enolase in Alzheimer’s Disease Brain: Beyond Altered Glucose Metabolism. J. Neurochem. 2009, 111, 915–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altenberg, B.; Greulich, K.O. Genes of Glycolysis Are Ubiquitously Overexpressed in 24 Cancer Classes. Genomics 2004, 84, 1014–1020. [Google Scholar] [CrossRef]
- Semenza, G.L.; Roth, P.H.; Fang, H.M.; Wang, G.L. Transcriptional Regulation of Genes Encoding Glycolytic Enzymes by Hypoxia-Inducible Factor 1. J. Biol. Chem. 1994, 269, 23757–23763. [Google Scholar] [CrossRef]
- Peng, C.; Hou, S.; Deng, C.; Zhang, Y. Function of DHX33 in Promoting Warburg Effect via Regulation of Glycolytic Genes. J. Cell. Physiol. 2021, 236, 981–996. [Google Scholar] [CrossRef] [PubMed]
- Capello, M.; Ferri-Borgogno, S.; Riganti, C.; Chattaragada, M.S.; Principe, M.; Roux, C.; Zhou, W.; Petricoin, E.F.; Cappello, P.; Novelli, F. Targeting the Warburg Effect in Cancer Cells through ENO1 Knockdown Rescues Oxidative Phosphorylation and Induces Growth Arrest. Oncotarget 2016, 7, 5598–5612. [Google Scholar] [CrossRef] [Green Version]
- Schofield, L.; Lincz, L.F.; Skelding, K.A. Unlikely Role of Glycolytic Enzyme α-Enolase in Cancer Metastasis and Its Potential as a Prognostic Biomarker. J. Cancer Metastasis Treat. 2020, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.S.; Wu, W.; Walsh, G.; Hong, W.K.; Mao, L. Enolase-Alpha Is Frequently down-Regulated in Non-Small Cell Lung Cancer and Predicts Aggressive Biological Behavior. Clin. Cancer Res. 2003, 9 Pt 1, 3641–3644. [Google Scholar] [PubMed]
- Cecconi, D.; Carbonare, L.D.; Mori, A.; Cheri, S.; Deiana, M.; Brandi, J.; Degaetano, V.; Masiero, V.; Innamorati, G.; Mottes, M.; et al. An Integrated Approach Identifies New Oncotargets in Melanoma. Oncotarget 2018, 9, 11489–11502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. Available online: http://www.nature.com/articles/nmeth.2019 (accessed on 12 March 2021). [CrossRef] [PubMed] [Green Version]
- Donizy, P.; Wu, C.-L.; Mull, J.; Fujimoto, M.; Chłopik, A.; Peng, Y.; Shalin, S.C.; Selim, M.A.; Puig, S.; Fernandez-Figueras, M.-T.; et al. Up-Regulation of PARP1 Expression Significantly Correlated with Poor Survival in Mucosal Melanomas. Cells 2020, 9, 1135. [Google Scholar] [CrossRef]
- Kassambara, A.; Kosinski, M.; Biecek, P. Survminer: Drawing Survival Curves Using “Ggplot”. Available online: https://cran.r-project.org/package=survminer (accessed on 12 March 2020).
- Park, H.S.; Kim, J.H.; Sun, B.K.; Song, S.U.; Suh, W.; Sung, J.-H. Hypoxia Induces Glucose Uptake and Metabolism of Adipose-Derived Stem Cells. Mol. Med. Rep. 2016, 14, 4706–4714. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Suo, C.; Zheng, C.; Zhang, H. Hypoxia and Metabolism in Metastasis. Adv. Med. Exp. Biol. 2019, 1136, 87–95. [Google Scholar] [CrossRef]
- Rofstad, E.K.; Rasmussen, H.; Galappathi, K.; Mathiesen, B.; Nilsen, K.; Graff, B.A. Hypoxia Promotes Lymph Node Metastasis in Human Melanoma Xenografts by Up-Regulating the Urokinase-Type Plasminogen Activator Receptor. Cancer Res. 2002, 62, 1847–1853. [Google Scholar]
- Fu, Q.-F.; Liu, Y.; Fan, Y.; Hua, S.-N.; Qu, H.-Y.; Dong, S.-W.; Li, R.-L.; Zhao, M.-Y.; Zhen, Y.; Yu, X.-L.; et al. Alpha-Enolase Promotes Cell Glycolysis, Growth, Migration, and Invasion in Non-Small Cell Lung Cancer through FAK-Mediated PI3K/AKT Pathway. J. Hematol. Oncol. 2015, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Ludvigsen, M.; Bjerregård Pedersen, M.; Lystlund Lauridsen, K.; Svenstrup Poulsen, T.; Hamilton-Dutoit, S.J.; Besenbacher, S.; Bendix, K.; Møller, M.B.; Nørgaard, P.; D’Amore, F.; et al. Proteomic Profiling Identifies Outcome-Predictive Markers in Patients with Peripheral T-Cell Lymphoma, Not Otherwise Specified. Blood Adv. 2018, 2, 2533–2542. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Sun, Y.; Sun, J.; Wang, Z.; Zhou, Y.; Yao, G.; Gu, Y.; Zhang, H.; Zhao, H. Differentially Expressed and Survival-Related Proteins of Lung Adenocarcinoma with Bone Metastasis. Cancer Med. 2018, 7, 1081–1092. [Google Scholar] [CrossRef]
- Cheng, Z.; Shao, X.; Xu, M.; Zhou, C.; Wang, J. ENO1 Acts as a Prognostic Biomarker Candidate and Promotes Tumor Growth and Migration Ability Through the Regulation of Rab1A in Colorectal Cancer. Cancer Manag. Res. 2019, 11, 9969–9978. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, B.; Zhu, S.; Zhang, J.; He, X.; Cao, G.; Chen, B. Hyperglycemia Promotes Snail-Induced Epithelial–Mesenchymal Transition of Gastric Cancer via Activating ENO1 Expression. Cancer Cell Int. 2019, 19, 344. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, Q.; Sun, H.; Wang, X.; Saiyin, H.; Zhang, H. The Circ-AMOTL1/ENO1 Axis Implicated in the Tumorigenesis of OLP-Associated Oral Squamous Cell Carcinoma. Cancer Manag. Res. 2020, 12, 7219–7230. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Wang, L.; Liu, H.-L. ENO1 Overexpression in Pancreatic Cancer Patients and Its Clinical and Diagnostic Significance. Gastroenterol. Res. Pract. 2018, 2018, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Dong, C.; Yin, Z.; Li, R.; Mao, J.; Wang, C.; Zhang, J.; Gao, Z.; Liang, R.; Wang, Q.; et al. Exosome-Derived ENO1 Regulates Integrin A6β4 Expression and Promotes Hepatocellular Carcinoma Growth and Metastasis. Cell Death Dis. 2020, 11, 972. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Liu, H.; Liang, Z.; Wang, F.; Zhou, C.; Zheng, X.; Zhang, Y.; Song, Y.; Hu, J.; He, X.; et al. Tumor-Intrinsic CD47 Signal Regulates Glycolysis and Promotes Colorectal Cancer Cell Growth and Metastasis. Theranostics 2020, 10, 4056–4072. [Google Scholar] [CrossRef]
- Pietraszek-Gremplewicz, K.; Simiczyjew, A.; Dratkiewicz, E.; Podgórska, M.; Styczeń, I.; Matkowski, R.; Ziętek, M.; Nowak, D. Expression Level of EGFR and MET Receptors Regulates Invasiveness of Melanoma Cells. J. Cell. Mol. Med. 2019, 23, 8453–8463. [Google Scholar] [CrossRef]
- Simiczyjew, A.; Pietraszek-Gremplewicz, K.; Dratkiewicz, E.; Podgórska, M.; Matkowski, R.; Ziętek, M.; Nowak, D. Combination of Selected MET and EGFR Inhibitors Decreases Melanoma Cells’ Invasive Abilities. Front. Pharmacol. 2019, 10, 1116. [Google Scholar] [CrossRef]
- Makowiecka, A.; Simiczyjew, A.; Nowak, D.; Mazur, A.J. Varying Effects of EGF, HGF and TGFβ on Formation of Invadopodia and Invasiveness of Melanoma Cell Lines of Different Origin. Eur. J. Histochem. 2016, 60, 2728. [Google Scholar] [CrossRef] [Green Version]
- Dratkiewicz, E.; Pietraszek-Gremplewicz, K.; Simiczyjew, A.; Mazur, A.J.; Nowak, D. Gefitinib or lapatinib with foretinib synergistically induce a cytotoxic effect in melanoma cell lines. Oncotarget 2018, 9, 18254–18268. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Li, N.; Huang, Z.; Chen, R.; Yi, P.; Kang, R.; Tang, D.; Hu, X.; Fan, X. A Novel LncRNA, TCONS_00006195, Represses Hepatocellular Carcinoma Progression by Inhibiting Enzymatic Activity of ENO1. Cell Death Dis. 2018, 9, 1184. [Google Scholar] [CrossRef]
- Wang, L.; Bi, R.; Yin, H.; Liu, H.; Li, L. ENO1 Silencing Impaires Hypoxia-Induced Gemcitabine Chemoresistance Associated with Redox Modulation in Pancreatic Cancer Cells. Am. J. Transl. Res. 2019, 11, 4470–4480. [Google Scholar] [PubMed]
- Zhang, W.; Gao, J.; Cheng, C.; Zhang, M.; Liu, W.; Ma, X.; Lei, W.; Hao, E.; Hou, X.; Hou, Y.; et al. Cinnamaldehyde Enhances Antimelanoma Activity through Covalently Binding ENO1 and Exhibits a Promoting Effect with Dacarbazine. Cancers 2020, 12, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gou, Y.; Li, F.; Huo, X.; Hao, C.; Yang, X.; Pei, Y.; Li, N.; Liu, H.; Zhu, B. ENO1 Monoclonal Antibody Inhibits Invasion, Proliferation and Clone Formation of Cervical Cancer Cells. Am. J. Cancer Res. 2021, 11, 1946–1961. [Google Scholar] [PubMed]
- Liu, Z.; Zhang, A.; Zheng, L.; Johnathan, A.F.; Zhang, J.; Zhang, G. The Biological Significance and Regulatory Mechanism of c-Myc Binding Protein 1 (MBP-1). Int. J. Mol. Sci. 2018, 19, 3868. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Parameters | ENO1 Expression | ||
|---|---|---|---|
| Low (H-Score ≤200) (N = 56) | High (H-Score >200) (N = 56) | p Value | |
| Age (18−86 years) a | 64 (52–73) | 65 (54–74) | 0.40 |
| Gender b | 1.00 | ||
| Female | 29 (52%) | 28 (50%) | |
| Male | 27 (48%) | 28 (50%) | |
| Primary tumor location c | 0.063 | ||
| Head/neck | 2 (4%) | 9 (16%) | |
| Extremities | 22 (39%) | 24 (43%) | |
| Trunk | 31 (55%) | 21 (38%) | |
| Hand/foot | 1 (2%) | 2 (4%) | |
| Primary tumor (pT) a | <0.001 | ||
| pT1 | 20 (36%) | 5 (9%) | |
| pT2 | 13 (23%) | 6 (11%) | |
| pT3 | 11 (20%) | 16 (29%) | |
| pT4 | 12 (21%) | 29 (52%) | |
| Sentinel lymph node biopsy status (SNLB) b | 0.042 | ||
| No metastases (SNLB−) | 19 (76%) | 11 (44%) | |
| Metastases present (SNLB+) | 6 (24%) | 14 (56%) | |
| Regional lymph nodes status (pN) b | 0.007 | ||
| Metastases absent (pN−) | 49 (88%) | 36 (64%) | |
| Metastases present (pN+) | 7 (12%) | 20 (36%) | |
| Distant metastases (pM) b | 0.53 | ||
| No metastases (pM−) | 52 (93%) | 49 (88%) | |
| Metastases present (pM+) | 4 (7%) | 7 (12%) | |
| AJCC (8th edition) stage a | <0.001 | ||
| I | 30 (54%) | 8 (14%) | |
| II | 17 (30%) | 25 (45%) | |
| III | 5 (9%) | 16 (29% | |
| IV | 4 (7%) | 7 (12%) | |
| Recurrence b | 0.018 | ||
| No | 42 (75%) | 29 (52%) | |
| Yes | 14 (25%) | 27 (48%) | |
| Histopathological Parameters | ENO1 Expression | ||
|---|---|---|---|
| Low (H-Score ≤200) (N = 56) | High (H-Score >200) (N = 56) | p Value | |
| Breslow thickness a | <0.001 | ||
| ≤1 mm | 20 (36%) | 5 (9%) | |
| 1.01−2.00 mm | 13 (23%) | 6 (11%) | |
| 2.01−4.00 mm | 11 (20%) | 16 (29%) | |
| >4 mm | 12 (21%) | 29 (52%) | |
| Clark level a | <0.001 | ||
| I | 0 (0%) | 0 (0%) | |
| II | 24 (43%) | 5 (9%) | |
| III | 17 (30%) | 24 (43%) | |
| IV | 11 (20%) | 20 (36%) | |
| V | 4 (7%) | 7 (12%) | |
| Histological type c | <0.001 | ||
| Superficial spreading melanoma | 35 (62%) | 13 (23%) | |
| Nodular melanoma | 20 (36%) | 41 (73%) | |
| Acral lentiginous melanoma | 1 (2%) | 2 (4%) | |
| Mitotic rate a | <0.001 | ||
| 0 | 22 (39%) | 2 (4%) | |
| 1−2 | 11 (20%) | 5 (9%) | |
| >2 | 23 (41%) | 49 (87%) | |
| Ulceration c | 0.013 | ||
| No | 39 (70%) | 25 (45%) | |
| Yes | 17 (30%) | 21 (55%) | |
| Lymphangioinvasion c | 1.0 | ||
| No | 54 (96%) | 52 (93%) | |
| Yes | 2 (4%) | 4 (7%) | |
| Tumor-infiltrating lymphocytes c | 0.039 | ||
| No | 5 (9%) | 2 (4%) | |
| Non-brisk | 28 (50%) | 41 (73%) | |
| Brisk | 23 (41%) | 13 (23%) | |
| Microsatellitosis c | 1.00 | ||
| No | 54 (96%) | 54 (96%) | |
| Yes | 2 (4%) | 2 (4%) | |
| Regression c | 1.00 | ||
| No | 53 (95%) | 54 (96%) | |
| Yes | 3 (5%) | 2 (4%) | |
| N | Cancer-Specific Overall Survival | Disease-Free Survival | |||||
|---|---|---|---|---|---|---|---|
| HR | 95% CI | p-Value | HR | 95% CI | p-Value | ||
| Sex | 128 | 0.4 | 0.2−0.8 | 0.015 * | 0.6 | 0.4−1.1 | 0.118 |
| Age | 128 | 3.3 | 1−10.4 | 0.043 * | 2.0 | 0.8−4.7 | 0.126 |
| pN | 128 | 5.3 | 2.6−11 | <0.001 * | 7.3 | 4−13 | <0.001 * |
| pM | 128 | 3.5 | 1.6−7.4 | <0.001 * | 3.3 | 1.7−6.8 | <0.001 * |
| AJCC (8th edition) stage | 112 | 17.4 | 3.9−77.9 | <0.001 * | 12.5 | 4.7−33 | <0.001 * |
| Breslow thickness | 128 | 9.0 | 2.1−38.7 | 0.003 * | 7.9 | 2.7−23 | <0.001 * |
| Clark level | 128 | 2.0 | 1.3−2.9 | <0.001 * | 1.8 | 1.4−2.5 | <0.001 * |
| Histologic type | 128 | 2.7 | 1.5−5 | 0.001 * | 3.3 | 1.9−5.6 | <0.001 * |
| Ulceration | 128 | 2.9 | 1.5−6 | 0.003 * | 2.3 | 1.3−4.1 | 0.005 * |
| Lymphovascular invasion | 128 | 2.0 | 0.6−6.6 | 0.249 | 1.1 | 0.3−4.6 | 0.877 |
| TILs | 128 | 0.12 | 0.1−0.5 | 0.002 * | 0.2 | 0.1−0.9 | 0.041 * |
| Microsatellitosis | 128 | 3.1 | 1.1−8.9 | 0.035 * | 3.5 | 1.3−9.9 | 0.016 * |
| ENO1 H-score | 112 | 2.4 | 1.1−5 | 0.027 * | 2.6 | 1.3−4.9 | 0.005 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hippner, M.; Majkowski, M.; Biecek, P.; Szkudlarek, T.; Simiczyjew, A.; Pieniazek, M.; Nowak, D.; Miazek, A.; Donizy, P. Alpha-Enolase (ENO1) Correlates with Invasiveness of Cutaneous Melanoma—An In Vitro and a Clinical Study. Diagnostics 2022, 12, 254. https://doi.org/10.3390/diagnostics12020254
Hippner M, Majkowski M, Biecek P, Szkudlarek T, Simiczyjew A, Pieniazek M, Nowak D, Miazek A, Donizy P. Alpha-Enolase (ENO1) Correlates with Invasiveness of Cutaneous Melanoma—An In Vitro and a Clinical Study. Diagnostics. 2022; 12(2):254. https://doi.org/10.3390/diagnostics12020254
Chicago/Turabian StyleHippner, Miriam, Michal Majkowski, Przemyslaw Biecek, Teresa Szkudlarek, Aleksandra Simiczyjew, Malgorzata Pieniazek, Dorota Nowak, Arkadiusz Miazek, and Piotr Donizy. 2022. "Alpha-Enolase (ENO1) Correlates with Invasiveness of Cutaneous Melanoma—An In Vitro and a Clinical Study" Diagnostics 12, no. 2: 254. https://doi.org/10.3390/diagnostics12020254