
The Profile of MicroRNA Expression in Bone Marrow in Non-Hodgkin’s Lymphomas

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Samples
2.2. Isolation of Total RNA
2.3. Selection of miRNAs
2.4. Reverse Transcription
2.5. Real-Time PCR
2.6. Statistical Analysis
2.7. Functional Analysis of miRNAs Using DIANA-miRPath
3. Results
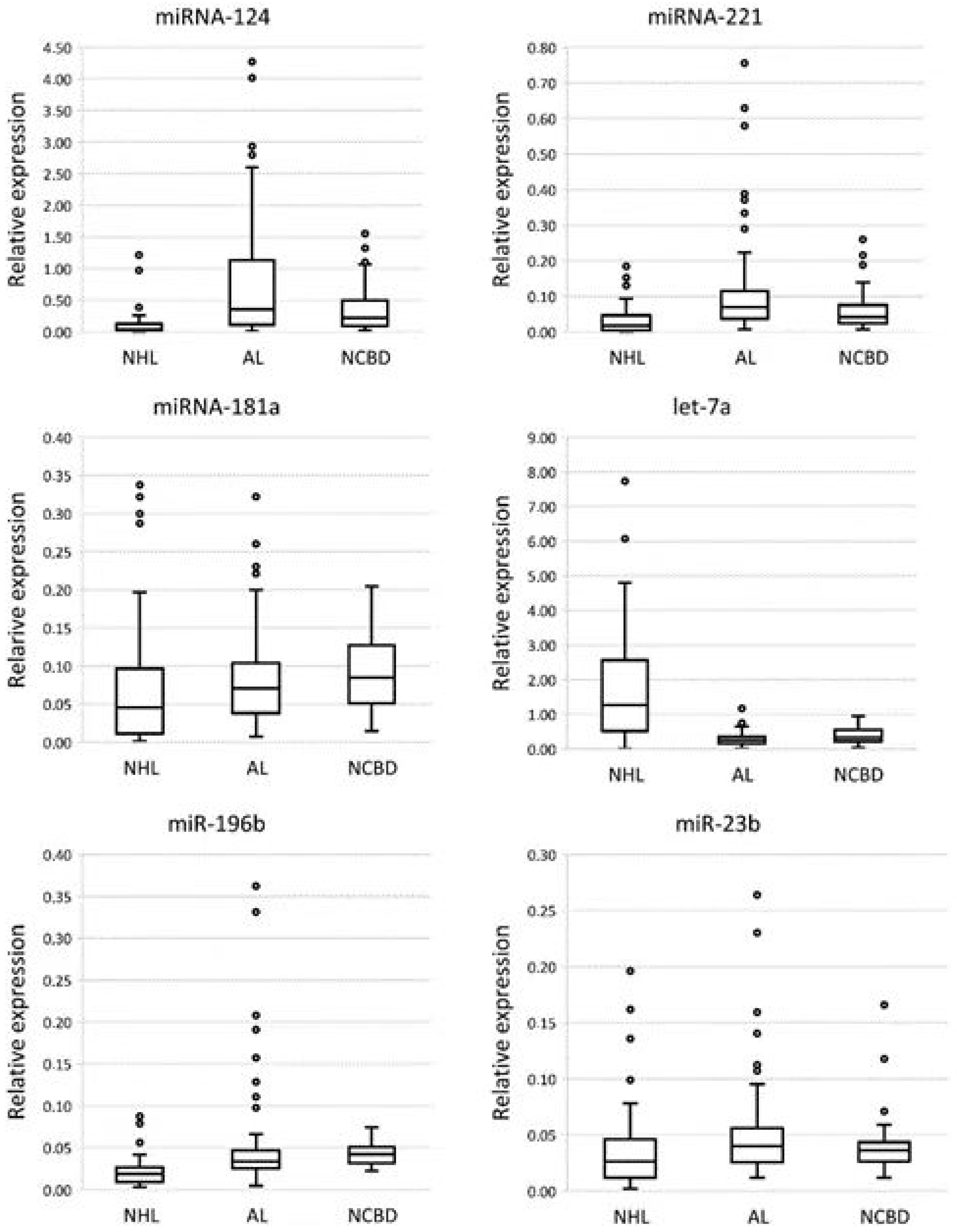
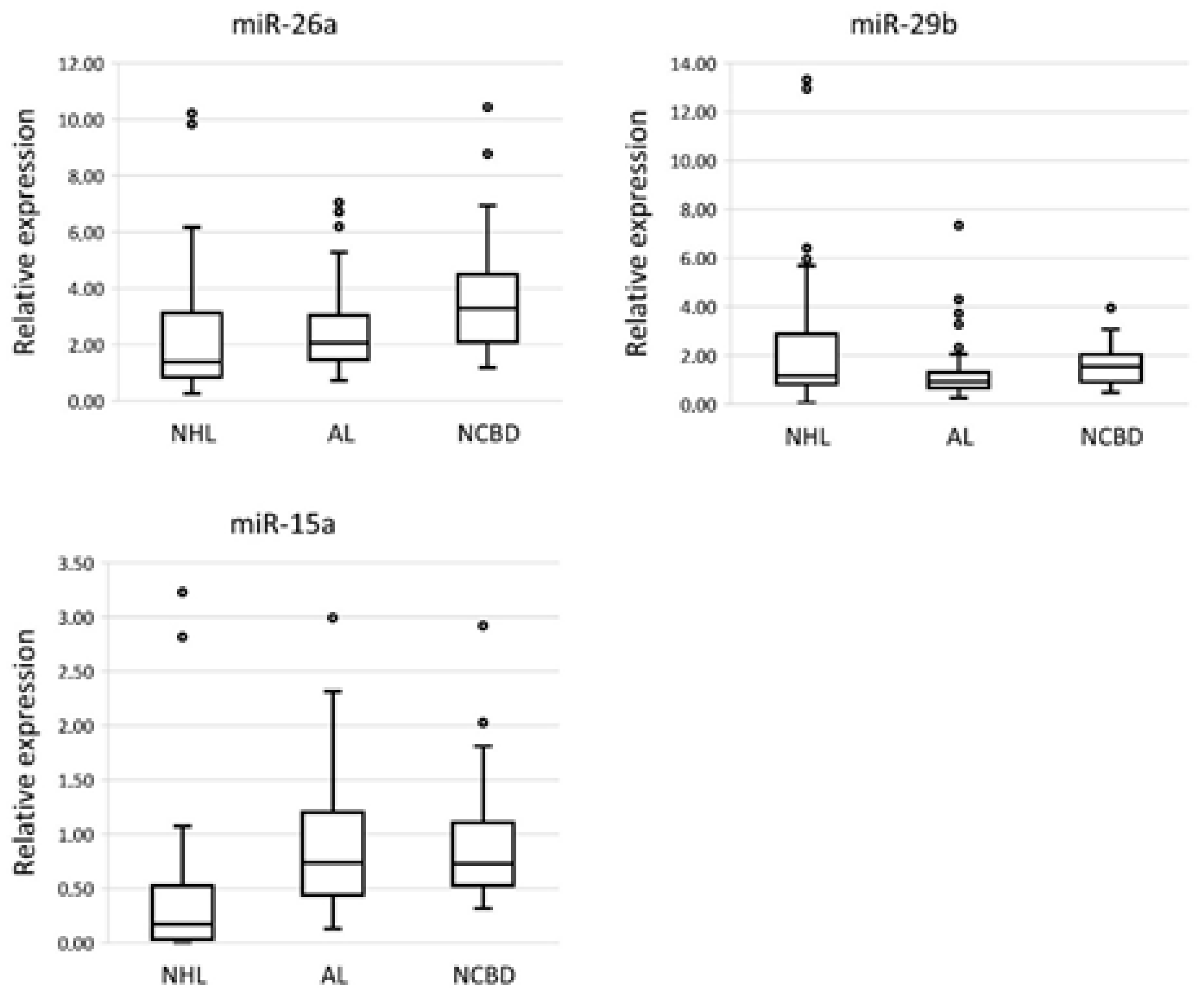
3.1. The Profile of miRNA Expression in BM Tumor Specimens and NCBD
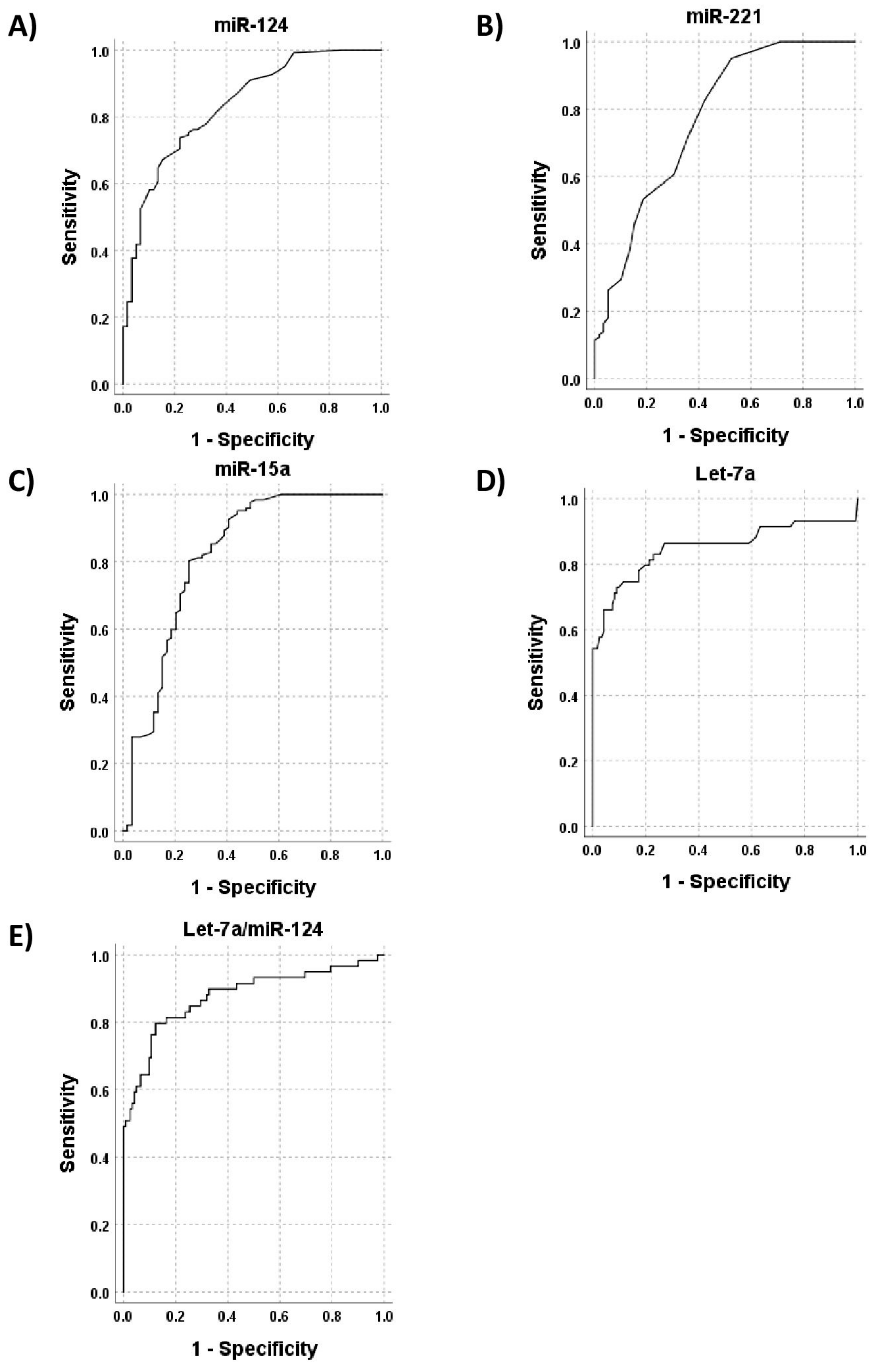
3.2. Diagnostic Significance
3.3. Bioinformatic Analysis of Pathways Involved in Cancer Processes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Singh, R.; Shaik, S.; Negi, B.S.; Rajguru, J.P.; Patil, P.B.; Parihar, A.S.; Sharma, U. Non-Hodgkin’s lymphoma: A review. J. Fam. Med. Prim. Care 2020, 9, 1834–1840. [Google Scholar]
- International Non-Hodgkin’s Lymphoma Prognostic Factors Project. A predictive model for aggressive non-Hodgkin’s lymphoma. N. Engl. J. Med. 1993, 329, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Park, B.B.; Jeong, J.Y.; Kim, W.Y.; Jang, S.; Shin, B.K.; Lee, D.S.; Han, J.H.; Park, C.J.; Suh, C.; et al. Assessment of bone marrow involvement in patients with lymphoma: Report on a consensus meeting of the Korean Society of Hematology Lymphoma Working Party. Korean J. Intern. Med. 2016, 31, 1030–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fend, F.; Kremer, M. Diagnosis and classification of malignant lymphoma and related entities in the bone marrow trephine biopsy. Pathobiology 2007, 74, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Albano, D.; Patti, C.; Lagalla, R.; Midiri, M.; Galia, M. Whole-body MRI, FDG-PET/CT, and bone marrow biopsy, for the assessment of bone marrow involvement in patients with newly diagnosed lymphoma. J. Magn. Reason. Imaging. 2017, 45, 1082–1089. [Google Scholar] [CrossRef]
- Kwoun, W.J.; Ahn, J.Y.; Park, P.W.; Seo, Y.H.; Kim, K.H.; Seo, J.Y.; Lee, H.T.; Yoo, K.H. How useful is bone marrow study as an initial investigative tool without lymph node biopsy in malignant lymphoma? Eleven years of experience at a single institution. J. Clin. Lab. Anal. 2019, 33, e22841. [Google Scholar] [CrossRef]
- Ribrag, V.; Vanel, D.; Leboulleux, S.; Lumbroso, J.; Couanet, D.; Bonniaud, G.; Aupérin, A.; Masson, F.; Bosq, J.; Edeline, V.; et al. Prospective study of bone marrow infiltration in aggressive lymphoma by three independent methods: Whole-body MRI, PET/CT and bone marrow biopsy. Eur. J. Radiol. 2008, 66, 325–331. [Google Scholar] [CrossRef]
- Kumar, S.; Rau, A.R.; Naik, R.; Kini, H.; Mathai, A.M.; Pai, M.R.; Khadilkar, U.N. Bone marrow biopsy in non-Hodgkin lymphoma: A morphological study. Indian J. Pathol. Microbiol. 2009, 52, 332–338. [Google Scholar] [CrossRef]
- Gilotra, M.; Gupta, M.; Singh, S.; Sen, R. Comparison of bone marrow aspiration cytology with bone marrow trephine biopsy histopathology: An observational study. J. Lab. Physicians 2017, 9, 182–189. [Google Scholar] [CrossRef]
- Kim, J.M.; Park, C.J.; Jang, S.S.; Chi, H.S.; Huh, J.R.; Kang, Y.K.; Seo, J.J. Clinico-hematologic findings of leukemic bone marrow involvement in Korean patients with non-hodgkin lymphoma. Lab. Med. Online 2011, 1, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Veryaskina, Y.A.; Titov, S.E.; Kovynev, I.B.; Fedorova, S.S.; Pospelova, T.I.; Zhimulev, I.F. MicroRNAs in the Myelodysplastic Syndrome. Acta Nat. 2021, 13, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Getaneh, Z.; Asrie, F.; Melku, M. MicroRNA profiles in B-cell non-Hodgkin lymphoma. EJIFCC 2019, 30, 195–214. [Google Scholar] [PubMed]
- Zheng, R.L.; Jiang, Y.J.; Wang, X. Role of microRNAs on therapy resistance in Non-Hodgkin’s lymphoma. Int. J. Clin. Exp. Med. 2014, 7, 3818–3832. [Google Scholar] [PubMed]
- Veryaskina, Y.A.; Titov, S.E.; Kovynev, I.B.; Pospelova, T.I.; Zhimulev, I.F. The miRNA Profile in Non-Hodgkin’s Lymphoma Patients with Secondary Myelodysplasia. Cells 2020, 9, 2318. [Google Scholar] [CrossRef] [PubMed]
- Bedewy, A.M.L.; Elmaghraby, S.M.; Shehata, A.A.; Kandil, N.S. Prognostic Value of miRNA-155 Expression in B-Cell Non-Hodgkin Lymphoma. Turk. J. Haematol. 2017, 34, 207–212. [Google Scholar]
- Larrabeiti-Etxebarria, A.; Lopez-Santillan, M.; Santos-Zorrozua, B.; Lopez-Lopez, E.; Garcia-Orad, A. Systematic Review of the Potential of MicroRNAs in Diffuse Large B Cell Lymphoma. Cancers 2019, 11, 144. [Google Scholar] [CrossRef] [Green Version]
- Titov, S.E.; Ivanov, M.K.; Karpinskaya, E.V.; Tsivlikova, E.V.; Shevchenko, S.P.; Veryaskina, Y.A.; Akhmerova, L.G.; Poloz, T.L.; Klimova, O.A.; Gulyaeva, L.F.; et al. miRNA profiling, detection of BRAF V600E mutation and RET-PTC1 translocation in patients from Novosibirsk oblast (Russia) with different types of thyroid tumors. BMC Cancer 2016, 16, 201. [Google Scholar] [CrossRef] [Green Version]
- Kluiver, J.; Kroesen, B.-J.; Poppema, S.; Berg, A. The role of microRNAs in normal hematopoiesis and hematopoietic malignancies. Leukemia 2006, 20, 1931–1936. [Google Scholar] [CrossRef]
- Roden, C.; Lu, J. MicroRNAs in Control of Stem Cells in Normal and Malignant Hematopoiesis. Curr. Stem. Cell Rep. 2016, 2, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Petriv, O.I.; Kuchenbauer, F.; Delaney, A.D.; Lecault, V.; White, A.; Kent, D.; Marmolejo, L.; Heuser, M.; Berg, T.; Copley, M.; et al. Comprehensive microRNA expression profiling of the hematopoietic hierarchy. Proc. Natl. Acad. Sci. USA 2010, 107, 15443–15448. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Sankaran, V.G.; Lodish, H.F. MicroRNAs in erythroid and megakaryocytic differentiation and megakaryocyte-erythroid progenitor lineage commitment. Leukemia 2012, 26, 2310–2316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Huang, X.; Timani, K.A.; Broxmeyer, H.E.; He, J.J. MicroRNA-124 Targets Tip110 Expression and Regulates Hematopoiesis. Stem. Cells Dev. 2015, 24, 2009–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovynev, I.B.; Titov, S.E.; Ruzankin, P.S.; Agakishiev, M.M.; Veryaskina, Y.A.; Nedel’ko, V.M.; Pospelova, T.I.; Zhimulev, I.F. Profiling 25 Bone Marrow microRNAs in Acute Leukemias and Secondary Nonleukemic Hematopoietic Conditions. Biomedicines 2020, 8, 607. [Google Scholar] [CrossRef]
- Chen, C.; Ridzon, D.A.; Broomer, A.J.; Zhou, Z.; Lee, D.H.; Nguyen, J.T.; Barbisin, M.; Xu, N.L.; Mahuvakar, V.R.; Andersen, M.R.; et al. Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res. 2005, 33, e179. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, X.; Wang, X.; Guo, X.; Ji, J.; Lou, G.; Zhao, J.; Zhou, W.; Guo, M.; Zhang, M.; Li, C.; et al. MicroRNA-124: An emerging therapeutic target in cancer. Cancer Med. 2019, 8, 5638–5656. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Jeong, D.; Nam, J.; Aung, T.N.; Gim, J.A.; Park, K.U.; Kim, S.W. MicroRNA-124 regulates glucocorticoid sensitivity by targeting phosphodiesterase 4B in diffuse large B cell lymphoma. Gene 2015, 558, 173–180. [Google Scholar] [CrossRef]
- Shim, H.; Nam, J.; Kim, S.W. NF-κB p65 represses microRNA-124 transcription in diffuse large B-cell lymphoma. Genes Genom. 2020, 42, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.; Kim, J.; Nam, J.; Sun, H.; Lee, Y.-H.; Lee, T.-J.; Aquiar, R.C.T.; Kim, S.-W. MicroRNA-124 links p53 to the NF-κB pathway in B-cell lymphomas. Leukemia 2015, 29, 1868–1874. [Google Scholar] [CrossRef]
- Fassina, A.; Marino, F.; Siri, M.; Zambello, R.; Ventura, L.; Fassan, M.; Simonato, F.; Cappellesso, R. The miR-17-92 microRNA cluster: A novel diagnostic tool in large B-cell malignancies. Lab. Investig. 2012, 92, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Di Lisio, L.; Sánchez-Beato, M.; Gómez-López, G.; Rodríguez, M.E.; Montes-Moreno, S.; Mollejo, M.; Menárguez, J.; Martínez, M.A.; Alves, F.J.; Pisano, D.G.; et al. MicroRNA signatures in B-cell lymphomas. Blood Cancer J. 2012, 2, e57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, J.; Shen, Y.; Huang, X.; Liu, Y.; Wake, L.; Liu, C.; Deffenbacher, K.; Lachel, C.M.; Wang, C.; Rohr, J.; et al. Global microRNA expression profiling uncovers molecular markers for classification and prognosis in aggressive B-cell lymphoma. Blood 2015, 125, 1137–1145. [Google Scholar] [CrossRef]
- Hezaveh, K.; Kloetgen, A.; Bernhart, S.H.; Mahapatra, K.D.; Lenze, D.; Richter, J.; Haake, A.; Bergmann, A.K.; Brors, B.; Burkhardt, B.; et al. ICGC MMML-Seq Project. Alterations of microRNA and microRNA-regulated messenger RNA expression in germinal center B-cell lymphomas determined by integrative sequencing analysis. Haematologica 2016, 101, 1380–1389. [Google Scholar] [CrossRef] [Green Version]
- Hershkovitz-Rokah, O.; Geva, P.; Salmon-Divon, M.; Shpilberg, O.; Liberman-Aronov, S. Network analysis of microRNAs, genes and their regulation in diffuse and follicular B-cell lymphomas. Oncotarget 2018, 9, 7928–7941. [Google Scholar] [CrossRef] [PubMed]
- Lenze, D.; Leoncini, L.; Hummel, M.; Volinia, S.; Liu, C.G.; Amato, T.; De Falco, G.; Githanga, J.; Horn, H.; Nyagol, J.; et al. The different epidemiologic subtypes of Burkitt lymphoma share a homogenous micro RNA profile distinct from diffuse large B-cell lymphoma. Leukemia 2011, 25, 1869–1876. [Google Scholar] [CrossRef] [Green Version]
- Zajdel, M.; Rymkiewicz, G.; Chechlinska, M.; Blachnio, K.; Pienkowska-Grela, B.; Grygalewicz, B.; Goryca, K.; Cieslikowska, M.; Bystydzienski, Z.; Swoboda, P.; et al. miR expression in MYC-negative DLBCL/BL with partial trisomy 11 is similar to classical Burkitt lymphoma and different from diffuse large B-cell lymphoma. Tumour. Biol. 2015, 36, 5377–5388. [Google Scholar] [CrossRef]
- Lawrie, C.H.; Chi, J.; Taylor, S.; Tramonti, D.; Ballabio, E.; Palazzo, S.; Saunders, N.J.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Expression of microRNAs in diffuse large B cell lymphoma is associated with immunophenotype, survival and transformation from follicular lymphoma. J. Cell Mol. Med. 2009, 13, 1248–1260. [Google Scholar] [CrossRef]
- Tan, L.P.; Wang, M.; Robertus, J.L.; Schakel, R.N.; Gibcus, J.H.; Diepstra, A.; Harms, G.; Peh, S.C.; Reijmers, R.M.; Pals, S.T.; et al. Mirna profiling of B-cell subsets: Specific miRNA profile for germinal center B cells with variation between centroblasts and centrocytes. Lab. Investig. 2009, 89, 708–716. [Google Scholar] [CrossRef] [Green Version]
- Sampson, V.B.; Rong, N.H.; Han, J.; Yang, Q.; Aris, V.; Soteropoulos, P.; Petrelli, N.J.; Dunn, S.P.; Krueger, L.J. MicroRNA let-7a down-regulates MYC and reverts MYC-induced growth in Burkitt lymphoma cells. Cancer Res. 2007, 67, 9762–9770. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Chen, X.; Lin, J.; Lwin, T.; Wright, G.; Moscinski, L.C.; Dalton, W.S.; Seto, E.; Wright, K.; Sotomayor, E.; et al. Myc represses miR-15a/miR-16-1 expression through recruitment of HDAC3 in mantle cell and other non-Hodgkin B-cell lymphomas. Oncogene 2012, 31, 3002–3008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgantas, R.W.; Hildreth, R.; Morisot, S.; Alder, J.; Liu, C.G.; Heimfeld, S.; Calin, G.A.; Croce, C.M.; Civin, C.I. CD34+ hematopoietic stem-progenitor cell microRNA expression and function: A circuit diagram of differentiation control. Proc. Natl. Acad. Sci. USA 2007, 104, 2750–2755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solé, C.; Larrea, E.; Di Pinto, G.; Tellaetxe, M.; Lawrie, C.H. miRNAs in B-cell lymphoma: Molecular mechanisms and biomarker potential. Cancer Lett. 2017, 405, 79–89. [Google Scholar] [CrossRef]
- Kim, B.; Lee, S.T.; Kim, H.J.; Kim, S.H. Bone marrow flow cytometry in staging of patients with B-cell non-Hodgkin lymphoma. Ann. Lab. Med. 2015, 35, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Bethge, N.; Honne, H.; Hilden, V.; Troen, G.; Eknaes, M.; Liestol, K.; Holte, H.; Delabie, J.; Smeland, E.B.; Lind, G.E. Identification of highly methylated genes across various types of B-cell non-hodgkin lymphoma. PLoS ONE 2013, 8, e79602. [Google Scholar] [CrossRef] [Green Version]
- Brueckner, B.; Stresemann, C.; Kuner, R.; Mund, C.; Musch, T.; Meister, M.; Sültmann, H.; Lyko, F. The human let-7a-3 locus contains an epigenetically regulated microRNA gene with oncogenic function. Cancer Res. 2007, 67, 1419–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazarlou, F.; Kadkhoda, S.; Ghafouri-Fard, S. Emerging role of let-7 family in the pathogenesis of hematological malignancies. Biomed. Pharm. 2021, 144, 112334. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| NHL vs. NCBD | Corrected p-Value | AL’s vs. NCBD | Corrected p-Value | NHL vs. ALs | Corrected p-Value | |
|---|---|---|---|---|---|---|
| miR-16 | 1 | NS | −1.23 | NS | 1.23 | NS |
| miR-155 | 1.02 | NS | 1.04 | NS | −1.03 | NS |
| miR-124 | −5.29 | 5 × 10−8 | 1.68 | NS | −8.87 | 5 × 10−10 |
| miR-221 | −2.34 | 0.004 | 1.7 | NS | −3.98 | 1 × 10−10 |
| miR-181a | −1.63 | 0.007 | −1.09 | NS | −1.49 | 0.002 |
| miR-30a | −1.17 | NS | −1.22 | NS | 1.05 | NS |
| miR-182 | −1.42 | NS | −1.04 | NS | −1.37 | NS |
| miR-7 | −1.63 | NS | −1.03 | NS | −1.59 | NS |
| let-7a | 3.88 | 4 × 10−7 | −1.3 | NS | 5.04 | 1 × 10−13 |
| miR-196b | −2.22 | 1 × 10−7 | −1.23 | NS | −1.81 | 1 × 10−7 |
| miR-20a | 1 | NS | −1.2 | NS | 1.21 | NS |
| miR-23b | −1.37 | NS | 1.1 | NS | −1.51 | 0.022 |
| miR-26a | −2.36 | NS | −1.59 | 0.006 | −1.49 | NS |
| miR-29b | −1.32 | NS | −1.66 | 0.003 | 1.25 | NS |
| miR-145 | 1.11 | NS | −1.24 | NS | 1.37 | NS |
| miR-15a | −4.25 | 3 × 10−8 | 1.01 | NS | −4.3 | 1 × 10−8 |
| miR-10b | −1.82 | NS | 1.08 | NS | −1.98 | NS |
| SLL vs. NCBD | Corrected p-Value | MCL vs. NCBD | Corrected p-Value | FL vs. NCBD | Corrected p-Value | MZL vs. NCBD | Corrected p-Value | DLBCL vs. NCBD | Corrected p-Value | |
|---|---|---|---|---|---|---|---|---|---|---|
| miR-16 | 1.07 | NS | 1.39 | NS | 1.27 | NS | −1.51 | NS | 1.22 | NS |
| miR-155 | 6.04 | NS | 3.91 | NS | −1.90 | NS | −1.03 | NS | −1.30 | NS |
| miR-124 | −1.66 | NS | −5.29 | 0.006 | −7.48 | 0.001 | −4.41 | 0.016 | −5.64 | 5 × 10−7 |
| miR-221 | −1.23 | NS | −1.71 | NS | −5.19 | NS | −1.97 | 0.045 | −5.66 | 2 × 10−4 |
| miR-181a | 1.02 | NS | −1.30 | NS | −2.42 | NS | −1.48 | NS | −2.10 | 2 × 10−4 |
| miR-30a | 2.00 | NS | 1.28 | NS | −1.17 | NS | −1.52 | NS | −1.27 | NS |
| miR-182 | −1.04 | NS | −1.05 | NS | 1.21 | NS | −1.52 | 0.007 | −1.15 | NS |
| miR-7 | 1.27 | NS | −1.31 | NS | −1.14 | NS | −3.49 | NS | −2.09 | 0.008 |
| let-7a | 2.85 | NS | 6.46 | NS | 3.18 | NS | 2.73 | 1 × 10−4 | 6.29 | 1 × 10−8 |
| miR-196b | −1.99 | 0.012 | −2.09 | NS | −1.82 | 1 × 10−4 | −2.87 | 1 × 10−8 | −2.22 | 2 × 10−8 |
| miR-20a | 1.48 | NS | 1.39 | 0.046 | 1.34 | NS | −1.11 | NS | −1.03 | NS |
| miR-23b | 1.28 | NS | −1.10 | NS | −2.02 | NS | −1.69 | NS | −1.43 | 0.015 |
| miR-26a | −2.75 | NS | −1.37 | NS | −1.01 | NS | 1.00 | NS | 1.05 | NS |
| miR-29b | 2.07 | NS | −1.40 | NS | −1.28 | NS | 1.05 | NS | −1.53 | NS |
| miR-145 | 1.96 | NS | −1.10 | NS | 1.46 | NS | 1.10 | NS | 1.22 | NS |
| miR-15a | −2.51 | NS | −1.25 | NS | −2.45 | 0.014 | −4.72 | 0.005 | −5.78 | 5 × 10−8 |
| miR-10b | 2.65 | N S | −2.20 | N S | −2.32 | N S | −2.33 | NS | −1.65 | NS |
| SLL vs. ALs | Corrected p-Value | MCL vs. ALs | Corrected p-Value | FL vs. ALs | Corrected p-Value | MZL vs. ALs | Corrected p-Value | DLBCL vs. ALs | Corrected p-Value | |
|---|---|---|---|---|---|---|---|---|---|---|
| miR-16 | 1.34 | NS | 1.74 | NS | 1.59 | NS | −1.21 | NS | 1.53 | NS |
| miR-155 | 5.81 | NS | 3.75 | NS | −1.98 | NS | −1.08 | NS | −1.36 | NS |
| miR-124 | −2.74 | NS | −8.73 | 0.013 | −1.98 | 0.001 | −7.28 | 0.002 | −9.31 | 1 × 10−8 |
| miR-221 | −2.13 | NS | −2.96 | NS | −12.35 | NS | −3.41 | 2 × 10−4 | −9.77 | 1 × 10−7 |
| miR-181a | 1.11 | NS | −1.19 | NS | −8.97 | NS | −1.35 | NS | −1.92 | 1 × 10−4 |
| miR-30a | 2.46 | NS | 1.58 | NS | −2.22 | NS | −1.23 | NS | −1.03 | NS |
| miR-182 | 1.02 | NS | 1.00 | NS | 1.27 | NS | −1.44 | NS | −1.09 | NS |
| miR-7 | 1.31 | NS | −1.28 | NS | −1.10 | NS | −3.39 | NS | −2.03 | 0.036 |
| let-7a | 3.55 | NS | 8.05 | NS | 3.96 | 0.013 | 3.40 | 3 × 10−8 | 7.83 | 1 × 10−14 |
| miR-196b | −1.63 | NS | −1.71 | NS | −1.49 | NS | −2.35 | 5 × 10−5 | −1.82 | 1 × 10−4 |
| miR-20a | 1.74 | NS | 1.63 | 0.009 | 1.57 | NS | 1.06 | NS | 1.14 | NS |
| miR-23b | 1.17 | NS | −1.21 | NS | −2.22 | NS | −1.85 | 0.030 | −1.57 | 0.003 |
| miR-26a | −1.96 | NS | −1.53 | NS | 1.11 | NS | 1.12 | NS | 1.17 | NS |
| miR-29b | 3.43 | NS | 1.18 | NS | 1.30 | NS | 1.74 | NS | 1.08 | NS |
| miR-145 | 2.45 | NS | 1.14 | NS | 1.83 | NS | 1.37 | NS | 1.52 | NS |
| miR-15a | −2.55 | NS | −1.27 | NS | −2.49 | NS | −4.79 | 0.004 | −5.87 | 2 × 10−8 |
| miR-10b | 2.39 | NS | −2.44 | NS | −2.58 | N S | −2.58 | NS | −1.83 | NS |
| AUC | Sensitivity | Specificity | |
|---|---|---|---|
| miR-124 | 0.835 | 74% | 78% |
| miR-15a | 0.816 | 81% | 71% |
| miR-221 | 0.769 | 71% | 64% |
| let-7a | 0.852 | 79% | 80% |
| let-7a/miR-124 | 0.908 | 91% | 80% |
| KEGG Pathway | Genes in the Pathway, Total | p Value | |
|---|---|---|---|
| miRNA-124 | |||
| Cell biology | |||
| Cell cycle (hsa04110) | 26 | 5 × 10−3 | |
| Cancer-associated pathways | |||
| Pathways in cancer (hsa05200) | 80 | 5 × 10−3 | |
| Proteoglycans in cancer (hsa05205) | 41 | 1.25 × 10−5 | |
| Chronic myeloid leukemia (hsa05220) | 20 | 1 × 10−2 | |
| Bladder cancer (hsa05219) | 15 | 1 × 10−3 | |
| Acute myeloid leukemia (hsa05221) | 16 | 1 × 10−2 | |
| Let-7a | |||
| Cell biology | |||
| Cell cycle (hsa04110) | 43 | 5.6 × 10−9 | |
| Cancer-associated pathways | |||
| Pathways in cancer (hsa05200) | 97 | 3 × 10−3 | |
| Proteoglycans in cancer (hsa05205) | 49 | 4 × 10−4 | |
| Chronic myeloid leukemia (hsa05220) | 20 | 2 × 10−3 | |
| Bladder cancer (hsa05219) | 13 | 3 × 10−2 | |
| Transcriptional misregulation in cancer (hsa05202) | 40 | 4 × 10−3 | |
| Signaling pathway | |||
| Hippo signaling pathway (hsa04390) | 42 | 2 × 10−7 | |
| p53 signaling pathway (hsa04115) | 23 | 1 × 10−3 | |
| FoxO signaling pathway (hsa04068) | 35 | 9 × 10−3 | |
| MAPK signaling pathway (hsa04010) | 51 | 7 × 10−3 | |
| TGF-beta signaling pathway (hsa04350) | 27 | 2 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veryaskina, Y.A.; Titov, S.E.; Kovynev, I.B.; Pospelova, T.I.; Zhimulev, I.F. The Profile of MicroRNA Expression in Bone Marrow in Non-Hodgkin’s Lymphomas. Diagnostics 2022, 12, 629. https://doi.org/10.3390/diagnostics12030629
Veryaskina YA, Titov SE, Kovynev IB, Pospelova TI, Zhimulev IF. The Profile of MicroRNA Expression in Bone Marrow in Non-Hodgkin’s Lymphomas. Diagnostics. 2022; 12(3):629. https://doi.org/10.3390/diagnostics12030629
Chicago/Turabian StyleVeryaskina, Yuliya A., Sergei E. Titov, Igor B. Kovynev, Tatiana I. Pospelova, and Igor F. Zhimulev. 2022. "The Profile of MicroRNA Expression in Bone Marrow in Non-Hodgkin’s Lymphomas" Diagnostics 12, no. 3: 629. https://doi.org/10.3390/diagnostics12030629