Quantitative Multiplex Real-Time Reverse Transcriptase–Polymerase Chain Reaction with Fluorescent Probe Detection of Killer Immunoglobulin-Like Receptors, KIR2DL4/3DL3



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Analyses and Probe Design
2.2. Reverse Transcription, Polymerase Chain Reaction and Plasmid Constructs
(molecules/mole)
(Length of DNA fragment) bp (× 660) Daltons
2.3. Optimization of Real Time Quantitative PCR (qRT–PCR)
2.4. Sensitivity and Reproducibility
2.5. Specificity
2.6. Human Sample Collection and RNA Extraction
2.7. Sensitivity and Specificity in Human Samples
2.8. Statistical Analysis
3. Results
3.1. Sequence Analysis and Probe Design
3.2. Optimization of Annealing Temperature for qRT-PCR
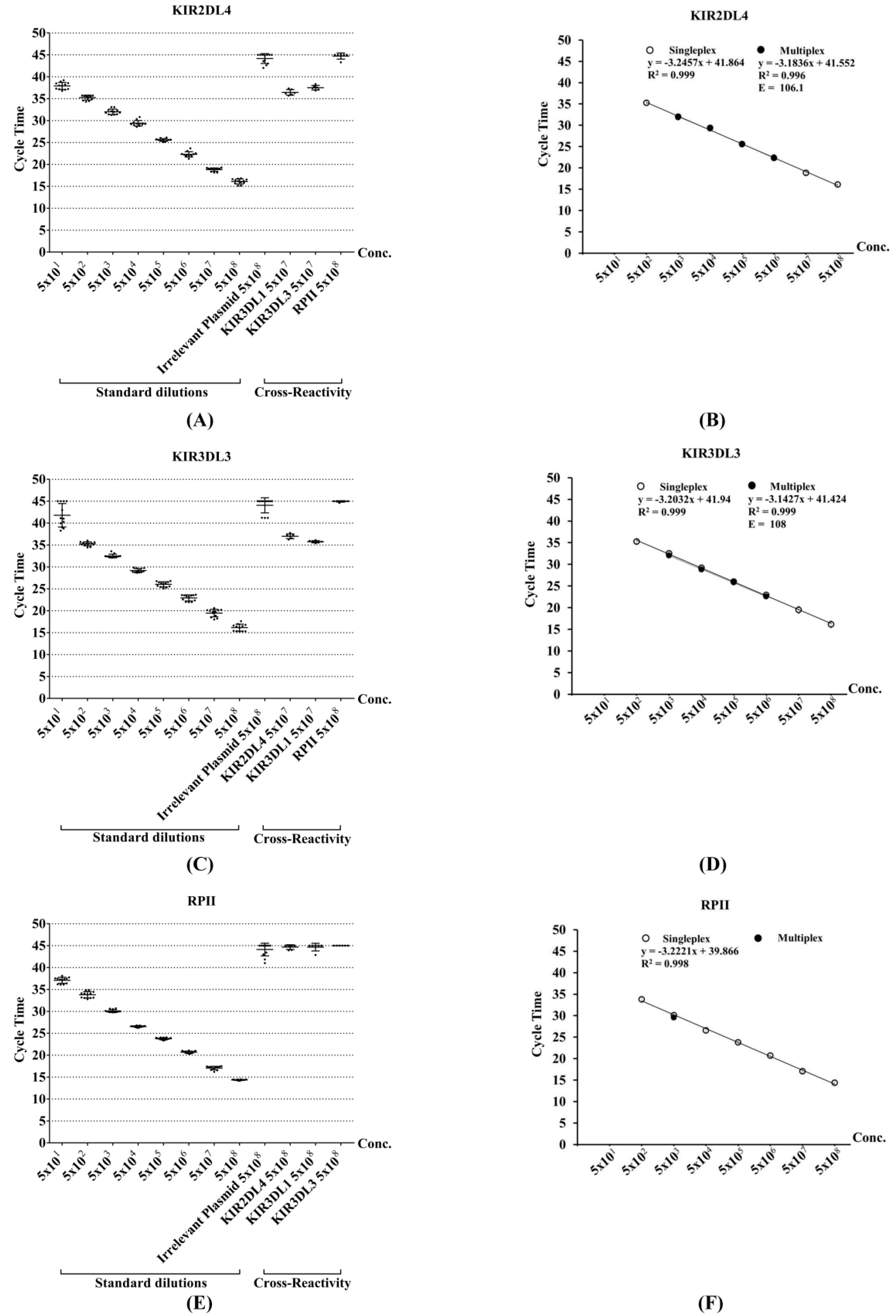
3.3. Assessment of Sensitivity and Reproducibility for Singleplex qRT-PCR
3.4. Assessment of Specificity for Singleplex qRT-PCR
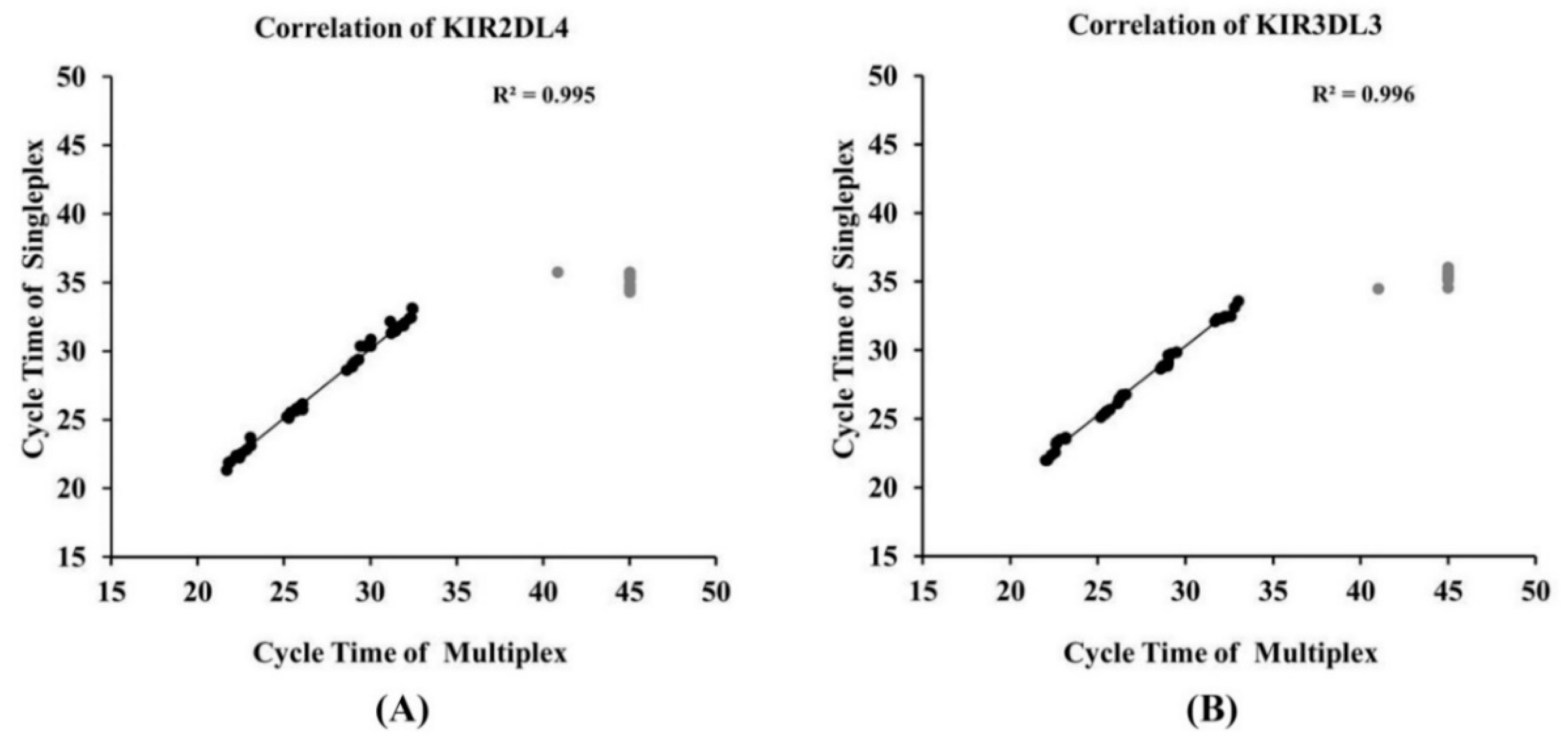
3.5. Development of Multiplex qRT-PCR
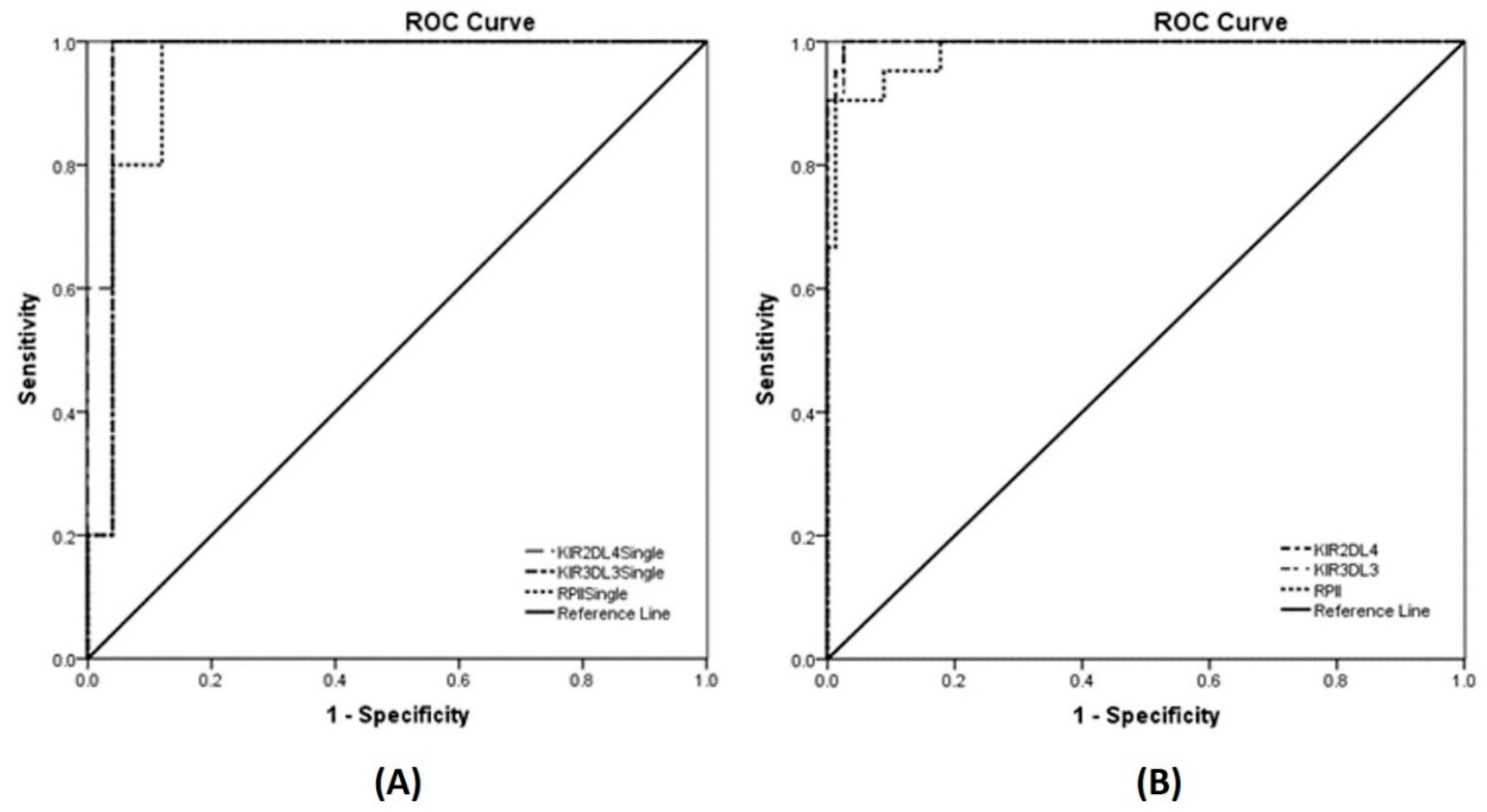
3.6. Detection of KIR Gene Expression in Human Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kärre, K.; Ljunggren, H.-G.; Piontek, G.; Kiessling, R. Selective rejection of H–2-deficient lymphoma variants suggests alternative immune defence strategy. Nature 1986, 319, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Algarra, I.; Cabrera, T.; Garrido, F. The HLA crossroad in tumor immunology. Hum. Immunol. 2000, 61, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.M.; Kulski, J.; Witt, C.; Pontarotti, P.; Christiansen, F. Leukocyte Ig-like receptor complex (LRC) in mice and men. Trends Immunol. 2002, 23, 81–88. [Google Scholar] [CrossRef]
- Lazetic, S.; Chang, C.; Houchins, J.P.; Lanier, L.L.; Phillips, J.H. Human natural killer cell receptors involved in MHC class I recognition are disulfide-linked heterodimers of CD94 and NKG2 subunits. J. Immunol. 1996, 157, 4741–4745. [Google Scholar]
- Vilches, C.; Parham, P. KIR: Diverse, Rapidly Evolving Receptors of Innate and Adaptive Immunity. Annu. Rev. Immunol. 2002, 20, 217–251. [Google Scholar] [CrossRef]
- Macfarlane, A.; Campbell, K.S. Signal Transduction in Natural Killer Cells. Curr. Top. Microbiol. Immunol. 2006, 298, 23–57. [Google Scholar]
- Hiby, S.E.; Walker, J.J.; O’Shaughnessy, K.M.; Redman, C.W.; Carrington, M.; Trowsdale, J.; Moffett, A. Combinations of Maternal KIR and Fetal HLA-C Genes Influence the Risk of Preeclampsia and Reproductive Success. J. Exp. Med. 2004, 200, 957–965. [Google Scholar] [CrossRef]
- Ruggeri, L.; Capanni, M.; Urbani, E.; Perruccio, K.; Shlomchik, W.D.; Tosti, A.; Posati, S.; Rogaia, D.; Frassoni, F.; Aversa, F.; et al. Effectiveness of Donor Natural Killer Cell Alloreactivity in Mismatched Hematopoietic Transplants. Science 2002, 295, 2097–2100. [Google Scholar] [CrossRef] [Green Version]
- Khakoo, I.S.; Thio, L.C.; Martin, P.M.; Brooks, R.C.; Gao, X.; Astemborski, J.; Cheng, J.; Goedert, J.J.; Vlahov, D.; Hilgartner, M.; et al. HLA and NK Cell Inhibitory Receptor Genes in Resolving Hepatitis C Virus Infection. Science 2004, 305, 872–874. [Google Scholar] [CrossRef]
- Hsu, K.C.; Liu, X.-R.; Selvakumar, A.; Mickelson, E.; O’Reilly, R.J.; Dupont, B. Killer Ig-like receptor haplotype analysis by gene content: Evidence for genomic diversity with a minimum of six basic framework haplotypes, each with multiple subsets. J. Immunol. 2002, 169, 5118–5129. [Google Scholar] [CrossRef]
- Chaisri, S.; Kitcharoen, K.; Romphruk, A.V.; Romphruk, A.; Witt, C.S.; Leelayuwat, C. Polymorphisms of killer immunoglobulin-like receptors (KIRs) and HLA ligands in northeastern Thais. Immunogenetics 2013, 65, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Chaisri, S.; A Traherne, J.; Jayaraman, J.; Romphruk, A.; Trowsdale, J.; Leelayuwat, C. Novel KIR genotypes and gene copy number variations in northeastern Thais. Immunology 2017, 153, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Valiante, N.M.; Uhrberg, M.; Shilling, H.G.; Lienert-Weidenbach, K.; Arnett, K.L.; D’Andrea, A.; Phillips, J.H.; Lanier, L.L.; Parham, P. Functionally and structurally distinct NK cell receptor repertoires in the peripheral blood of two human donors. Immunity 1997, 7, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Faure, M.; Long, E.O. KIR2DL4 (CD158d), an NK Cell-Activating Receptor with Inhibitory Potential. J. Immunol. 2002, 168, 6208–6214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Houser, B.L.; Nicotra, M.L.; Strominger, J.L. HLA-G homodimer-induced cytokine secretion through HLA-G receptors on human decidual macrophages and natural killer cells. Proc. Natl. Acad. Sci. USA 2009, 106, 5767–5772. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, S.; Antoine, M.; Uzan, S.; McMaster, M.; Dausset, J.; Carosella, E.D.; Paul, P. Specific activation of the non-classical class I histocompatibility HLA-G antigen and expression of the ILT2 inhibitory receptor in human breast cancer. J. Pathol. 2002, 196, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Crisa, L.; McMaster, M.T.; Ishii, J.K.; Fisher, S.J.; Salomon, D.R. Identification of a Thymic Epithelial Cell Subset Sharing Expression of the Class Ib HLA-G Molecule with Fetal Trophoblasts. J. Exp. Med. 1997, 186, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.C.; Hiby, S.E.; Moffett, A.; Trowsdale, J.; Young, N.T. Nature of allelic sequence polymorphism at the KIR3DL3 locus. Immunogenetics 2006, 58, 614–627. [Google Scholar] [CrossRef]
- Trundley, A.E.; Hiby, S.E.; Chang, C.; Sharkey, A.M.; Santourlidis, S.; Uhrberg, M.; Trowsdale, J.; Moffett, A. Molecular characterization of KIR3DL3. Immunogenetics 2006, 57, 904–916. [Google Scholar] [CrossRef]
- IPD-KIR Database. Available online: https://www.ebi.ac.uk/ipd/kir/ (accessed on 20 June 2020).
- Clustal Omega: Multiple Sequence Alignment. Available online: https://www.ebi.ac.uk/Tools/msa/clustalo/ (accessed on 20 June 2020).
- Tajik, N.; Shahsavar, F.; Mousavi, T.; Radjabzadeh, M.F. Distribution of KIRgenes in the Iranian population. Tissue Antigens 2009, 74, 22–31. [Google Scholar] [CrossRef]
- Godornes, C.; Leader, B.T.; Molini, B.J.; Centurion-Lara, A.; Lukehart, S.A. Quantitation of rabbit cytokine mRNA by real-time RT-PCR. Cytokine 2007, 38, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, C.R.; Elliott, T.; Parham, P.; Khakoo, S.I. The Inhibitory Receptor NKG2A Determines Lysis of Vaccinia Virus-Infected Autologous Targets by NK Cells. J. Immunol. 2006, 176, 1141–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraman, J.; Kirgizova, V.; Di, D.; Johnson, C.; Jiang, W.; Traherne, J.A. qKAT: Quantitative Semi-automated Typing of Killer-cell Immunoglobulin-like Receptor Genes. J. Vis. Exp. 2019, 145, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Radonic, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef]
- Rajagopalan, S.; Long, E.O. A Human Histocompatibility Leukocyte Antigen (HLA)-G–specific Receptor Expressed on All Natural Killer Cells. J. Exp. Med. 1999, 189, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Rajalingam, R.; Hong, M.; Adams, E.J.; Shum, B.P.; Guethlein, L.A.; Parham, P. Short KIR Haplotypes in Pygmy Chimpanzee (Bonobo) Resemble the Conserved Framework of Diverse Human KIR Haplotypes. J. Exp. Med. 2001, 193, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Goodridge, J.P.; Lathbury, L.J.; Steiner, N.K.; Shulse, C.N.; Pullikotil, P.; Seidah, N.G.; Hurley, C.K.; Christiansen, F.T.; Witt, C.S. Three common alleles of KIR2DL4 (CD158d) encode constitutively expressed, inducible and secreted receptors in NK cells. Eur. J. Immunol. 2007, 37, 199–211. [Google Scholar] [CrossRef]
- Kikuchi-Maki, A.; Yusa, S.-I.; Catina, T.L.; Campbell, K.S. KIR2DL4 Is an IL-2-Regulated NK Cell Receptor That Exhibits Limited Expression in Humans but Triggers Strong IFN-γ Production. J. Immunol. 2003, 171, 3415–3425. [Google Scholar] [CrossRef]
- Takei, Y.; Ueshima, C.; Kataoka, T.; Hirata, M.; Sugimoto, A.; Rokutan-Kurata, M.; Moriyoshi, K.; Ono, K.; Murakami, I.; Iwamoto, S.; et al. Killer cell immunoglobulin-like receptor 2DL4 is expressed in and suppresses the cell growth of Langerhans cell histiocytosis. Oncotarget 2017, 8, 36964–36972. [Google Scholar] [CrossRef] [Green Version]
- LeMaoult, J.; Zafaranloo, K.; Le Danff, C.; Carosella, E.D. HLA-G up-regulates ILT2, ILT3, ILT4, and KIR2DL4 in antigen presenting cells, NK cells, and T cells. FASEB J. 2005, 19, 1–23. [Google Scholar] [CrossRef]
- Roe, D.; Vierra-Green, C.; Pyo, C.-W.; Eng, K.; Hall, R.; Kuang, R.; Spellman, S.; Ranade, S.; Geraghty, D.E.; Maiers, M. Revealing complete complex KIR haplotypes phased by long-read sequencing technology. Genes Immun. 2017, 18, 127–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Probe | 5′ Fluorophore–Sequence–Quencher 3′ | Position (Exon) | Product Size (bp) | Reference |
|---|---|---|---|---|
| Specific probes for quantitative real–time RT–PCR | ||||
| P-2DL4 | FAM–TGGGTTTAACATCTTCACGCTGTAC–BHQ1 | 89–113 (E3) | This study | |
| P-3DL3 | TexasRed–TGCGGGTTCCCAGGTCAACTATTCCATGG–BHQ2 | 164–193 (E4) | This study | |
| P-RPII | TET–TGCTGGACCCACCGGCATGTTCTTTGG–BHQ1 | 180–187 (E26),1–19 (E27) | This study | |
| Primer | Primer sequences 5′–3′ | |||
| Specific primers for quantitative real-time RT–PCR | ||||
| 2DL4 | F: TCAGGACAAGCCCTTCTGC | 5–23 (E3) | 130 | [20] |
| R: GACAGGGACCCCATCTTTC | 116–134 (E3) | |||
| 3DL3 | F: GCAATGTTGGTCAGATGTCAG | 71–91 (E4) | 199 | [20] |
| R: AGCCGACAACTCATAGGGTA | 250–269 (E4) | |||
| RPII | F: AGTATGGCATGGAGATCCCC | 137–156 (E26) | 143 | This study |
| R: ATAGGCAGGGGTTGCACC | 75–92 (E27) | |||
| Genes | AUC (95% CI) | SE | p-Value |
|---|---|---|---|
| Singleplex qRT–PCR | |||
| KIR2DL4 | 0.984 (0.945, 1.000) | 0.020 | 0.001 |
| KIR3DL3 | 0.968 (0.903, 1.000) | 0.033 | 0.001 |
| RPII (reference gene) | 0.952 (0.876, 1.000) | 0.039 | 0.002 |
| Multiplex qRT–PCR | |||
| KIR2DL4 | 0.998 (0.994, 1.000) | 0.002 | <0.001 |
| KIR3DL3 | 0.998 (0.992, 1.000) | 0.003 | <0.001 |
| RPII (reference gene) | 0.984 (0.964, 1.000) | 0.011 | <0.001 |
| Genes | Sensitivity (%) | Specificity (%) | +LR | −LR | YI |
|---|---|---|---|---|---|
| Singleplex qRT-PCR | |||||
| KIR2DL4 | 100.000 | 99.960 | 25 | 0.040 | 99.960 |
| KIR3DL3 | 100.000 | 99.960 | 25 | 0.040 | 99.960 |
| RPII (reference gene) | 100.000 | 99.880 | 8.33 | 0.120 | 99.880 |
| Multiplex qRT-PCR | |||||
| KIR2DL4 | 100.000 | 99.975 | 40 | 0.025 | 99.975 |
| KIR3DL3 | 100.000 | 99.975 | 40 | 0.025 | 99.975 |
| RPII (reference gene) | 100.000 | 99.855 | 6.896 | 0.145 | 99.855 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wongfieng, W.; Nutalai, R.; Jumnainsong, A.; Leelayuwat, C. Quantitative Multiplex Real-Time Reverse Transcriptase–Polymerase Chain Reaction with Fluorescent Probe Detection of Killer Immunoglobulin-Like Receptors, KIR2DL4/3DL3. Diagnostics 2020, 10, 588. https://doi.org/10.3390/diagnostics10080588
Wongfieng W, Nutalai R, Jumnainsong A, Leelayuwat C. Quantitative Multiplex Real-Time Reverse Transcriptase–Polymerase Chain Reaction with Fluorescent Probe Detection of Killer Immunoglobulin-Like Receptors, KIR2DL4/3DL3. Diagnostics. 2020; 10(8):588. https://doi.org/10.3390/diagnostics10080588
Chicago/Turabian StyleWongfieng, Wipaporn, Rungtiwa Nutalai, Amonrat Jumnainsong, and Chanvit Leelayuwat. 2020. "Quantitative Multiplex Real-Time Reverse Transcriptase–Polymerase Chain Reaction with Fluorescent Probe Detection of Killer Immunoglobulin-Like Receptors, KIR2DL4/3DL3" Diagnostics 10, no. 8: 588. https://doi.org/10.3390/diagnostics10080588