Phosphates as Energy Sources to Expand Metabolic Networks

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Reconstruction of the Background Metabolism Pool
2.3. Network Expansion Simulation
2.4. Thermodynamically Constrained Network Expansion Simulation
2.5. Scale-Limiting Reaction Detection
- (1).
- Obtaining the potential reactions: Reaction set R1 and the corresponding metabolite set M1 at the thermodynamic threshold τ1 limiting the network expansion and reaction set R2 and corresponding metabolite set M2 at τ2 (τ2 = τ1 + 1) were obtained
- (2).
- Identifying the reactions triggering the dramatic expansion of networks: At first, a metabolite m derived from difference set (M3) of M2 and M1 (M3 = M2 - M1) was added into the metabolite set M1 to form a new metabolite set M4 (M4 = M1 + m). Then, new networks at τ1 were constructed with M4. Once the network expands dramatically, the corresponding reactions of m were considered to cause the dramatic expansion of networks. These steps were repeated for every metabolite in M3, and all of the expansion-triggering reactions were identified as the potential scale-limiting reactions.
- (3).
- Identifying the scale-limiting reactions: Since there may be more than one reaction limiting the dramatic expansion of the network, all combinations of potential scale-limiting reactions should be tested. Every combination of potential scale-limiting reactions was blocked in turn. The networks at the thermodynamic constraint τ2 were constructed based on these abridged reaction sets. When dramatic network expansion was no longer observed, the reaction combinations were considered as scale-limiting ones. Thus, the final scale-limiting reactions were identified by analyzing all the combinations.
2.6. Protein Domain Age Estimation
3. Results
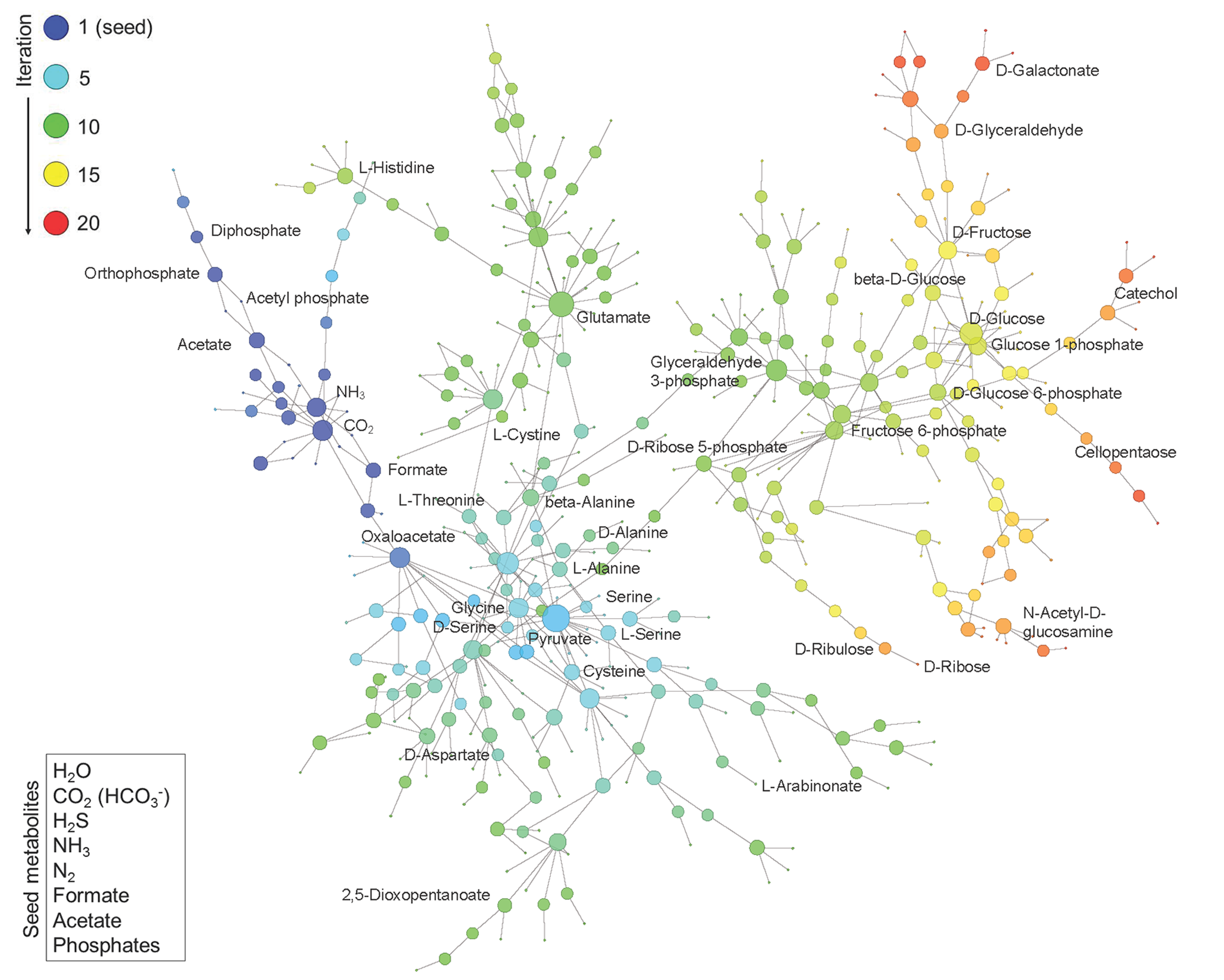
3.1. Construction of Phosphorus-Dependent Metabolic Network
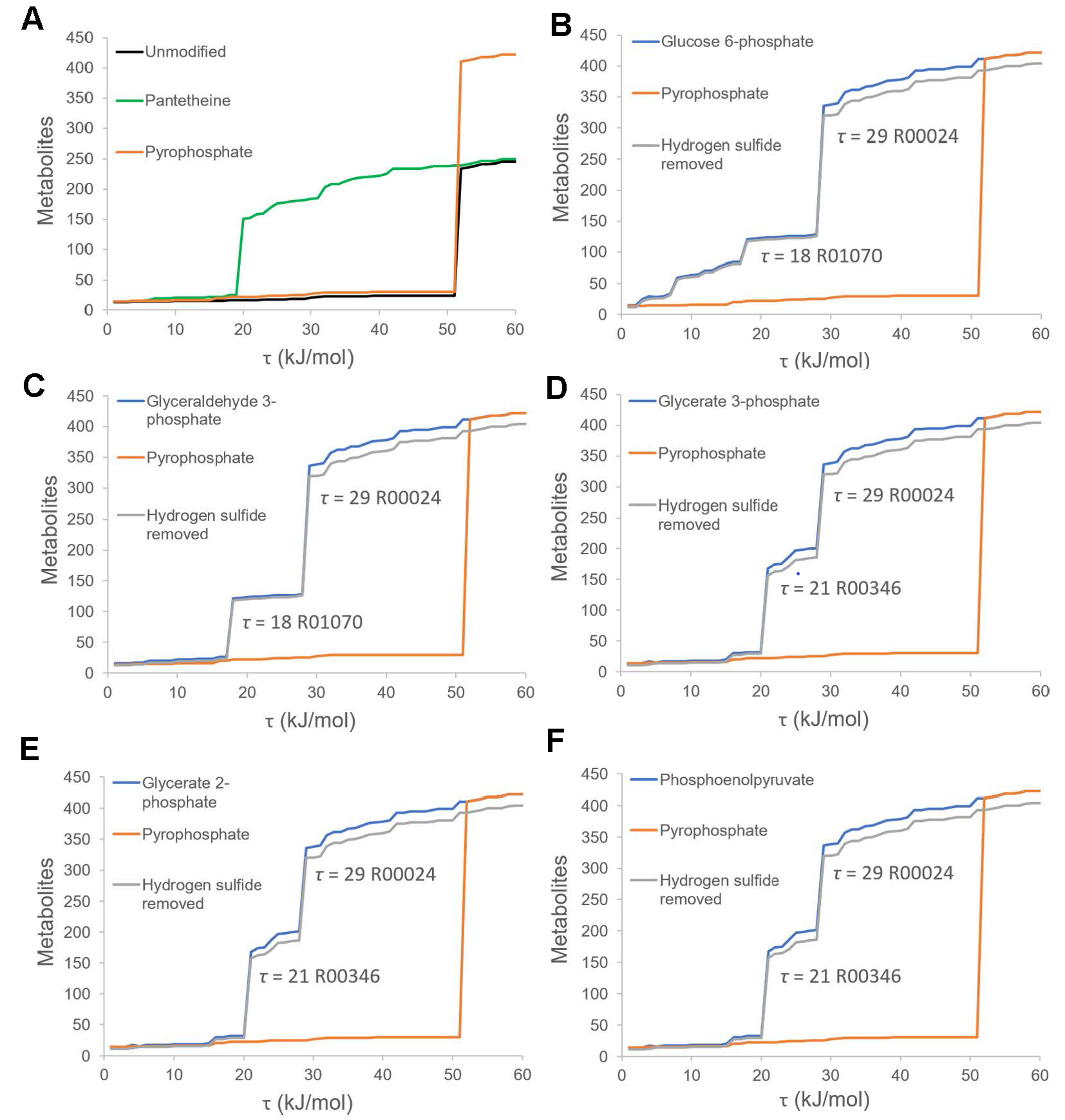
3.2. Thermodynamic Bottleneck Alleviation by Primitive Phosphates
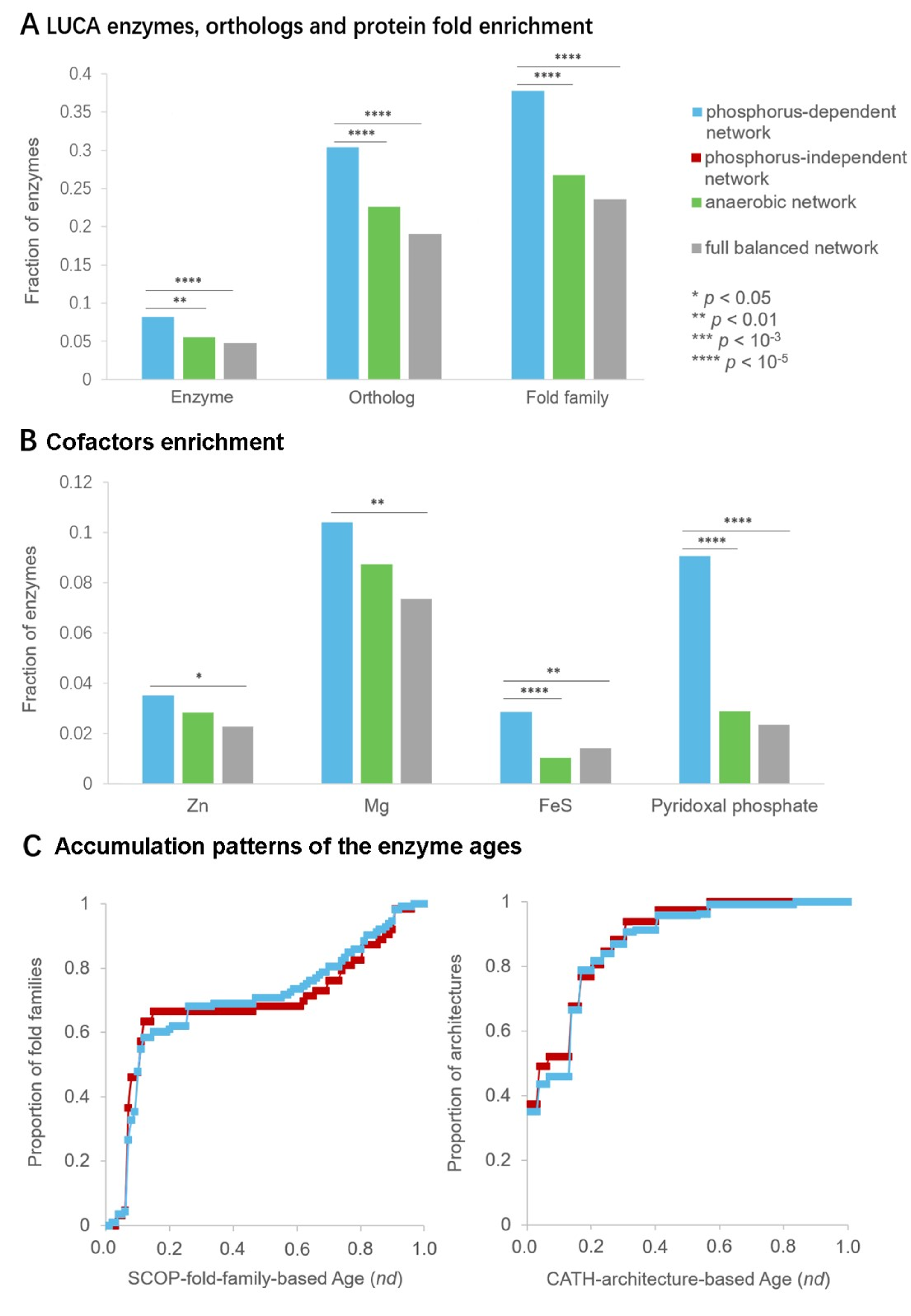
3.3. Ancient Origin of Phosphorus-Dependent Metabolic Network
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schwartz, A.W. Phosphorus in prebiotic chemistry. Philos. Trans. R. Soc. B 2006, 361, 1743–1749. [Google Scholar] [CrossRef] [Green Version]
- Gedulin, B.; Arrhenius, G. Sources and geochemical evolution of RNA precursor molecules-the role of phosphate. Early Life Earth 1994, 84, 91–110. [Google Scholar]
- Handschuh, G.J.; Orgel, L.E. Struvite and prebiotic phosphorylation. Science 1973, 179, 483–484. [Google Scholar] [CrossRef]
- Goldford, J.E.; Hartman, H.; Smith, T.F.; Segrè, D. Remnants of an ancient metabolism without phosphate. Cell 2017, 168, 1126–1134. [Google Scholar] [CrossRef]
- Islam, S.; Bučar, D.K.; Powner, M.W. Prebiotic selection and assembly of proteinogenic amino acids and natural nucleotides from complex mixtures. Nat. Chem. 2017, 9, 584–589. [Google Scholar] [CrossRef]
- Zhang, W.; Tam, C.P.; Walton, T.; Fahrenbach, A.C.; Birrane, G.; Szostak, J.W. Insight into the mechanism of nonenzymatic RNA primer extension from the structure of an RNA-GpppG complex. Proc. Natl. Acad. Sci. USA 2017, 114, 7659–7664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Liu, Y.; Xu, P.X.; Cai, M.Y.; Zhao, Y.F. Alpha-amino acid behaves differently from beta- or gamma-amino acids as treated by trimetaphosphate. Amino Acids 2008, 34, 47–53. [Google Scholar] [CrossRef]
- Pascal, R.; Poitevin, F.; Boiteau, L. Energy sources for prebiotic chemistry and early life: Constraints and availability. Orig. Life Evol. Biosph. 2009, 39, 260–261. [Google Scholar]
- Martin, W.; Russell, M.J. On the origin of biochemistry at an alkaline hydrothermal vent. Philos. Trans. R. Soc. B 2007, 362, 1887–1925. [Google Scholar] [CrossRef]
- Piast, R.W.; Wieczorek, R.M. Origin of life and the phosphate transfer catalyst. Astrobiology 2017, 17, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Pasek, M.A. Schreibersite on the early Earth: Scenarios for prebiotic phosphorylation. Geosci. Front. 2017, 8, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Pasek, M.A.; Harnmeijer, J.P.; Buick, R.; Gull, R.; Atlas, Z. Evidence for reactive reduced phosphorus species in the early Archean ocean. Proc. Natl. Acad. Sci. USA 2013, 110, 10089–10094. [Google Scholar] [CrossRef] [Green Version]
- Pasek, M.; Kee, T.; Bryant, D.; Pavlov, A.A.; Lunine, J.I. Production of potentially prebiotic condensed phosphates by phosphorus redox chemistry. Angew. Chem. 2008, 120, 8036–8038. [Google Scholar] [CrossRef]
- Gibard, C.; Bhowmik, S.; Karki, M.; Kim, E.-K.; Krishnamurthy, R. Phosphorylation, oligomerization and self-assembly in water under potential prebiotic conditions. Nat. Chem. 2018, 10, 212–217. [Google Scholar] [CrossRef]
- Ebenhöh, O.; Handorf, T.; Heinrich, R. Structural analysis of expanding metabolic networks. Genome Inf. 2004, 15, 35–45. [Google Scholar]
- Handorf, T.; Ebenhöh, O.; Heinrich, R. Expanding metabolic networks: Scopes of compounds, robustness, and evolution. J. Mol. Evol. 2005, 61, 498–512. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.; Segrè, D. The effect of oxygen on biochemical networks and the evolution of complex life. Science 2006, 311, 1764–1767. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Goldman, A.D.; Bernhard, T.M.; Dolzhenko, E.; Landweber, L.F. LUCApedia: A database for the study of ancient life. Nucleic Acids Res. 2013, 41, D1079–D1082. [Google Scholar] [CrossRef]
- Hubbard, T.J.; Murzin, A.G.; Brenner, S.E.; Chothia, C. SCOP: A structural classification of proteins database. Nucleic Acids Res. 1997, 25, 236–239. [Google Scholar] [CrossRef]
- Dawson, N.L.; Lewis, T.E.; Das, S.; Lees, J.G.; Lee, D.; Ashford, P.; Orengo, C.A.; Sillitoe, I. CATH: An expanded resource to predict protein function through structure and sequence. Nucleic Acids Res. 2017, 45, D289–D295. [Google Scholar] [CrossRef]
- Osterberg, R.; Orgel, L.E. Polyphosphate and trimetaphosphate formation under potentially prebiotic conditions. J. Mol. Evol. 1972, 1, 241–248. [Google Scholar] [CrossRef]
- Yamagata, Y.; Watanabe, H.; Saitoh, M.; Namba, T. Volcanic production of polyphosphates and its relevance to prebiotic evolution. Nature 1991, 352, 516–519. [Google Scholar] [CrossRef]
- Hermes-Lima, M.; Vieyra, A. Pyrophosphate synthesis from phosphor-(enol)pyruvate catalyzed by precipitated magnesium phosphate with “enzyme-like activity”. J. Mol. Evol. 1992, 35, 277–285. [Google Scholar] [CrossRef]
- Zwart, I.I.D.; Meade, S.J.; Pratt, A.J. Biomimetic phosphoryl transfer catalysed by iron(II)-mineral precipitates. Geochem. Cosmochim. Acta 2004, 68, 4093–4098. [Google Scholar] [CrossRef]
- Dodd, M.S.; Papineau, D.; Grenne, T.; Slack, J.F.; Pirajno, F.; O’Neil, J.; Little, C.T.S. Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature 2017, 543, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Flamholz, A.; Noor, E.; Bar-Even, A.; Milo, R. eQuilibrator-the biochemical thermodynamics calculator. Nucleic Acids Res. 2012, 40, D770–D775. [Google Scholar] [CrossRef]
- Maheen, G.; Wang, Y.; Wang, Y.; Shi, Z.; Tian, G.; Feng, S. Mimicking the prebiotic acidic hydrothermal environment: One-pot prebiotic hydrothermal synthesis of glucose phosphates. Heteroat. Chem. 2011, 22, 186–191. [Google Scholar] [CrossRef]
- Coggins, A.J.; Powner, M.W. Prebiotic synthesis of phosphoenol pyruvate by α-phosphorylation-controlled triose glycolysis. Nat. Chem. 2017, 9, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Noor, E.; Haraldsdóttir, H.S.; Milo, R.; Fleming, R.M. Consistent estimation of Gibbs energy using component contributions. PLoS Comput. Biol. 2013, 9, e1003098. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jiang, Y.Y.; Kim, K.M.; Qu, G.; Ji, H.-F.; Mittenthal, J.E.; Zhang, H.-Y.; Caetano-Anolles, G. A universal molecular clock of protein folds and its power in tracing the early history of aerobic metabolism and planet oxygenation. Mol. Biol. Evol. 2011, 28, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, C.A.; Gustavo, C.A. Structural phylogenomics reveals gradual evolutionary replacement of abiotic chemistries by protein enzymes in purine metabolism. PLoS ONE 2013, 8, e59300. [Google Scholar]
- Kim, K.M.; Qin, T.; Jiang, Y.Y.; Chen, L.-L.; Xiong, M.; Caetano-Anollés, D.; Zhang, H.-Y.; Caetano-Anollés, G. Protein domain structure uncovers the origin of aerobic metabolism and the rise of planetary oxygen. Structure 2012, 20, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Caetano-Anollés, G.; Kim, K.M.; Caetano-Anollés, D. The phylogenomic roots of modern biochemistry: Origins of proteins, cofactors and protein biosynthesis. J. Mol. Evol. 2012, 74, 1–34. [Google Scholar] [CrossRef]
- Bukhari, S.A.; Caetano-Anollés, G. Origin and evolution of protein fold designs inferred from phylogenomic analysis of CATH domain structures in proteomes. PLoS Comput. Biol. 2013, 9, e1003009. [Google Scholar] [CrossRef] [PubMed]
- Bar-Even, A.; Flamholz, A.; Noor, E.; Milo, R. Thermodynamic constraints shape the structure of carbon fixation pathways. Biochim. Biophys. Acta 2012, 1817, 1646–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, M.A.; Turchyn, A.V.; Ralser, M. Non-enzymatic glycolysis and pentose phosphate pathway-like reactions in a plausible Archean ocean. Mol. Syst. Biol. 2014, 10, 725. [Google Scholar] [CrossRef]
- Ślesak, I.; Ślesak, H.; Kruk, J. RubisCO early oxygenase activity: A kinetic and evolutionary perspective. Bioessays 2017, 39, 1700071. [Google Scholar] [CrossRef]
- Caetano-Anollés, G. RubisCO and the search for biomolecular culprits of planetary change. Bioessays 2017, 39, 1700074. [Google Scholar] [CrossRef]
- Nascimento, J.M.; Shi, L.Z.; Tam, J.; Chandsawangbhuwana, C.; Durrant, B.; Botvinick, E.L.; Berns, M.W. Comparison of glycolysis and oxidative phosphorylation as energy sources for mammalian sperm motility, using the combination of fluorescence imaging, laser tweezers, and real-time automated tracking and trapping. J. Cell. Physiol. 2008, 217, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Ashida, H.; Sakiyama, T.; de Marsac, N.T.; Danchin, A.; Sekowska, A.; Yokota, A. Structural and functional similarities between a ribulose-1,5-bisphosphate carboxylase/oxygenase (rubisco)-like protein from bacillus subtilis and photosynthetic rubisco. J. Biol. Chem. 2009, 284, 13256–13264. [Google Scholar] [CrossRef]
- Kai, T. Sweet siblings with different faces: The mechanisms of FBP and F6P aldolase, transaldolase, transketolase and phosphoketolase revisited in light of recent structural data. Bioorg. Chem. 2014, 57, 263–280. [Google Scholar]
- Nitschke, W.; McGlynn, S.E.; Milner-White, E.J.; Russell, M.J. On the antiquity of metalloenzymes and their substrates in bioenergetics. Biochim. Biophys. Acta 2013, 1827, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Sousa, F.L.; Martin, W.F. Biochemical fossils of the ancient transition from geoenergetics to bioenergetics in prokaryotic one carbon compound metabolism. Biochim. Biophys. Acta 2014, 1837, 964–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabinski, R.F.; Toney, M.D. Metal ion inhibition of nonenzymatic pyridoxal phosphate catalyzed decarboxylation and transamination. J. Am. Chem. Soc. 2001, 123, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Waite, J.H.; Glein, C.R.; Perryman, R.S.; Teolis, B.D.; Magee, B.A.; Miller, G.; Grimes, J.; Perry, M.E.; Miller, K.E.; Bouquet, A.; et al. Cassini finds molecular hydrogen in the Enceladus plume: Evidence for hydrothermal processes. Science 2017, 356, 155–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, T.; Chu, X.-Y.; Yang, Y.; Zhang, X.; Liu, Y.-M.; Gao, J.; Ma, B.-G.; Zhang, H.-Y. Phosphates as Energy Sources to Expand Metabolic Networks. Life 2019, 9, 43. https://doi.org/10.3390/life9020043
Tian T, Chu X-Y, Yang Y, Zhang X, Liu Y-M, Gao J, Ma B-G, Zhang H-Y. Phosphates as Energy Sources to Expand Metabolic Networks. Life. 2019; 9(2):43. https://doi.org/10.3390/life9020043
Chicago/Turabian StyleTian, Tian, Xin-Yi Chu, Yi Yang, Xuan Zhang, Ye-Mao Liu, Jun Gao, Bin-Guang Ma, and Hong-Yu Zhang. 2019. "Phosphates as Energy Sources to Expand Metabolic Networks" Life 9, no. 2: 43. https://doi.org/10.3390/life9020043