
Aerobic Exercise Modulates Proteomic Profiles in Gastrocnemius Muscle of db/db Mice, Ameliorating Sarcopenia

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Animal Model Design and Exercise Intervention
2.2. Blood Glucose Levels
2.3. Histological Analysis
2.4. Sample Preparation for Proteomics Analysis
2.5. Proteomic Analysis Using LC-MS/MS
2.6. Bioinformatic Tools
2.7. Statistical Analysis
3. Results
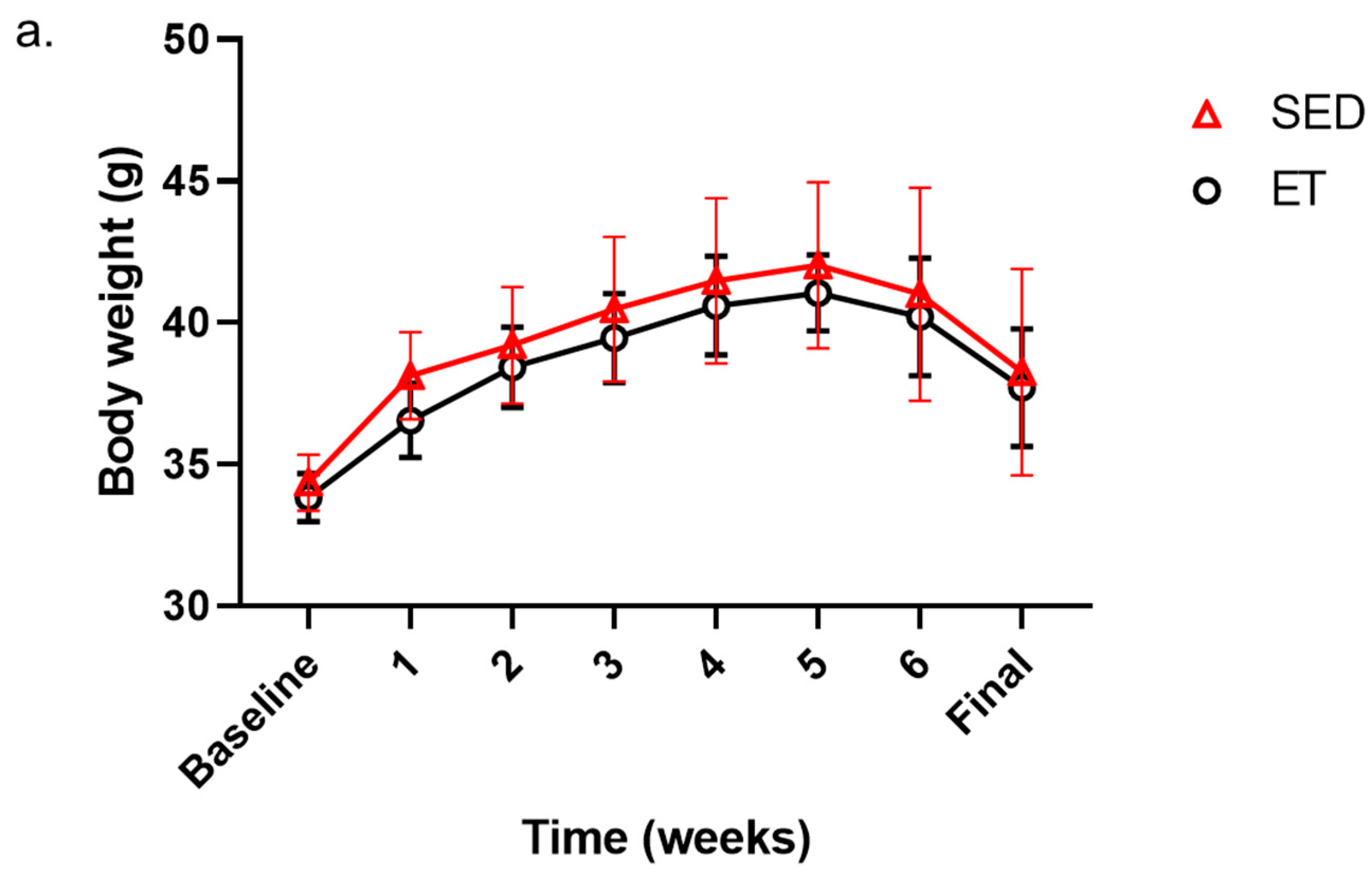
3.1. The Impact of Aerobic Exercise on Body Weight and Blood Glucose Levels
3.2. The Impact of Aerobic Exercise on Skeletal Muscle Composition
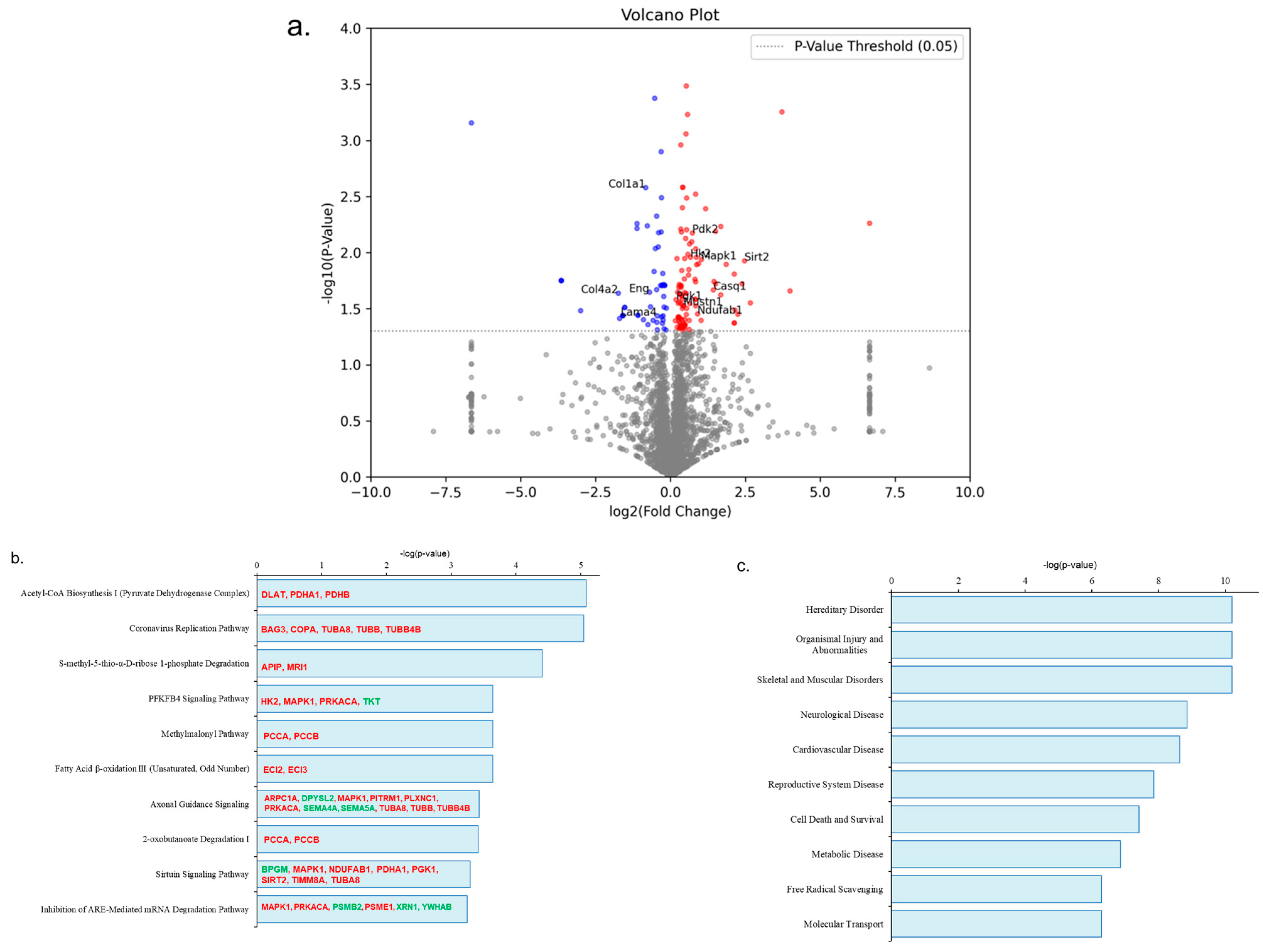
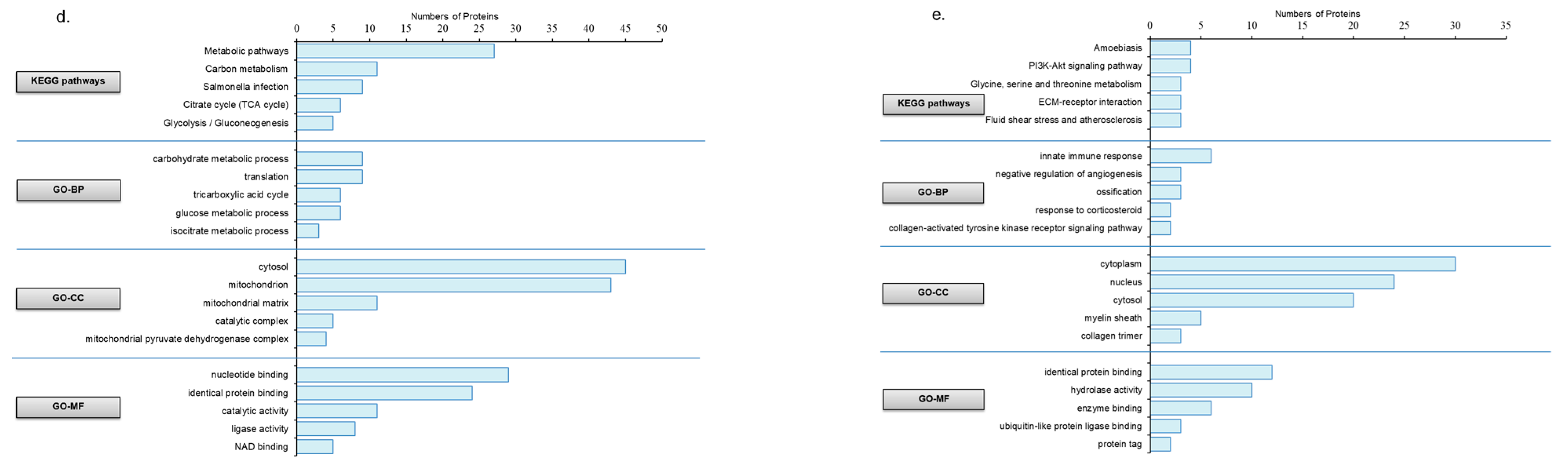
3.3. The Impact of Aerobic Exercise on the Proteomic Analysis of Gastrocnemius Muscle
3.4. The Impact of Aerobic Exercise on Proteomic Analysis of Soleus Muscle
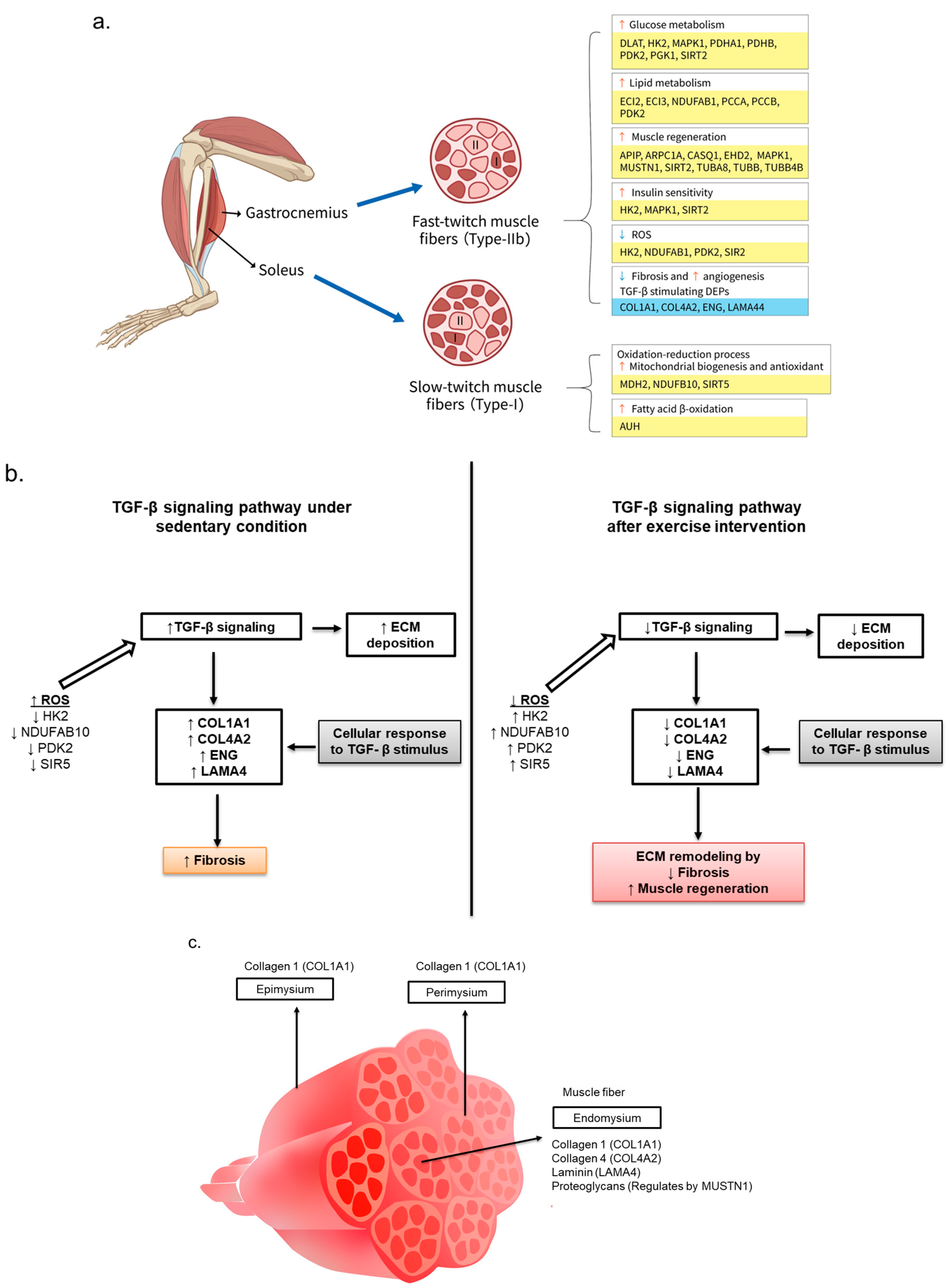
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trierweiler, H.; Kisielewicz, G.; Hoffmann Jonasson, T.; Rasmussen Petterle, R.; Aguiar Moreira, C.; Zeghbi Cochenski Borba, V. Sarcopenia: A chronic complication of type 2 diabetes mellitus. Diabetol. Metab. Syndr. 2018, 10, 25. [Google Scholar] [CrossRef]
- International Diabetes Federation. Diabetes Atlas-Taiwan Diabetes Report 2000–2045. 2021. Available online: https://diabetesatlas.org/data/en/ (accessed on 13 March 2024).
- Yang, J.; Qian, F.; Chavarro, J.E.; Ley, S.H.; Tobias, D.K.; Yeung, E.; Hinkle, S.N.; Bao, W.; Li, M.; Liu, A.; et al. Modifiable risk factors and long term risk of type 2 diabetes among individuals with a history of gestational diabetes mellitus: Prospective cohort study. BMJ 2022, 378, e070312. [Google Scholar] [CrossRef]
- Seidu, S.; Khunti, K.; Yates, T.; Almaqhawi, A.; Davies, M.J.; Sargeant, J. The importance of physical activity in management of type 2 diabetes and COVID-19. Ther. Adv. Endocrinol. Metab. 2021, 12, 20420188211054686. [Google Scholar] [CrossRef]
- Colberg, S.R.; Sigal, R.J.; Yardley, J.E.; Riddell, M.C.; Dunstan, D.W.; Dempsey, P.C.; Horton, E.S.; Castorino, K.; Tate, D.F. Physical Activity/Exercise and Diabetes: A Position Statement of the American Diabetes Association. Diabetes Care 2016, 39, 2065–2079. [Google Scholar] [CrossRef]
- Kirwan, J.P.; Sacks, J.; Nieuwoudt, S. The essential role of exercise in the management of type 2 diabetes. Clevel. Clin. J. Med. 2017, 84, S15–S21. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, A.S.; Steenberg, D.E.; Hostrup, M.; Birk, J.B.; Larsen, J.K.; Santos, A.; Kjøbsted, R.; Hingst, J.R.; Schéele, C.C.; Murgia, M.; et al. Deep muscle-proteomic analysis of freeze-dried human muscle biopsies reveals fiber type-specific adaptations to exercise training. Nat. Commun. 2021, 12, 304. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Niu, Y.; Liu, X.; Yang, F.; Niu, W.; Fu, L. Proteomic analysis of skeletal muscle in insulin-resistant mice: Response to 6-week aerobic exercise. PLoS ONE 2013, 8, e53887. [Google Scholar] [CrossRef]
- Kleinert, M.; Parker, B.L.; Jensen, T.E.; Raun, S.H.; Pham, P.; Han, X.; James, D.E.; Richter, E.A.; Sylow, L. Quantitative proteomic characterization of cellular pathways associated with altered insulin sensitivity in skeletal muscle following high-fat diet feeding and exercise training. Sci. Rep. 2018, 8, 10723. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.J.; Lu, C.W.; Lin, L.Y.; Hsu, Y.J.; Huang, C.C.; Huang, K.C. Proteomic Analysis of Skeletal Muscle and White Adipose Tissue after Aerobic Exercise Training in High Fat Diet Induced Obese Mice. Int. J. Mol. Sci. 2023, 24, 5743. [Google Scholar] [CrossRef] [PubMed]
- Murgia, M.; Nogara, L.; Baraldo, M.; Reggiani, C.; Mann, M.; Schiaffino, S. Protein profile of fiber types in human skeletal muscle: A single-fiber proteomics study. Skelet. Muscle 2021, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Ostler, J.E.; Maurya, S.K.; Dials, J.; Roof, S.R.; Devor, S.T.; Ziolo, M.T.; Periasamy, M. Effects of insulin resistance on skeletal muscle growth and exercise capacity in type 2 diabetic mouse models. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E592–E605. [Google Scholar] [CrossRef]
- Hsu, Y.J.; Wu, M.F.; Lee, M.C.; Huang, C.C. Exercise training combined with Bifidobacterium longum OLP-01 treatment regulates insulin resistance and physical performance in db/db mice. Food Funct. 2021, 12, 7728–7740. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Marcotte-Chénard, A.; Oliveira, B.; Little, J.P.; Candow, D.G. Sarcopenia and type 2 diabetes: Pathophysiology and potential therapeutic lifestyle interventions. Diabetes Metab. Syndr. Clin. Res. Rev. 2023, 17, 102835. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Takahashi, F.; Okamura, T.; Hamaguchi, M.; Fukui, M. Diet, exercise, and pharmacotherapy for sarcopenia in people with diabetes. Metabolism 2023, 144, 155585. [Google Scholar] [CrossRef] [PubMed]
- Tung, Y.T.; Hsu, Y.J.; Liao, C.C.; Ho, S.T.; Huang, C.C.; Huang, W.C. Physiological and Biochemical Effects of Intrinsically High and Low Exercise Capacities Through Multiomics Approaches. Front. Physiol. 2019, 10, 1201. [Google Scholar] [CrossRef] [PubMed]
- Mahdy, M. Skeletal muscle fibrosis: An overview. Cell Tissue Res. 2019, 375, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, L. Diabetes and Collagen: Interrelations. Avicenna J. Med. Biochem. 2019, 7, 64–71. [Google Scholar] [CrossRef]
- Heyne, E.; Zeeb, S.; Junker, C.; Petzinna, A.; Schrepper, A.; Doenst, T.; Koch, L.G.; Britton, S.L.; Schwarzer, M. Exercise Training Differentially Affects Skeletal Muscle Mitochondria in Rats with Inherited High or Low Exercise Capacity. Cells 2024, 13, 393. [Google Scholar] [CrossRef]
- Lee, E.-J.; Lee, M.-M.; Park, S.; Jeong, K.-S. Sirt2 positively regulates muscle regeneration after Notexin-induced muscle injury. Exp. Mol. Pathol. 2022, 127, 104798. [Google Scholar] [CrossRef]
- Misawa, T.; Takahama, M.; Kozaki, T.; Lee, H.; Zou, J.; Saitoh, T.; Akira, S. Microtubule-driven spatial arrangement of mitochondria promotes activation of the NLRP3 inflammasome. Nat. Immunol. 2013, 14, 454–460. [Google Scholar] [CrossRef]
- Kitada, M.; Ogura, Y.; Monno, I.; Koya, D. Sirtuins and Type 2 Diabetes: Role in Inflammation, Oxidative Stress, and Mitochondrial Function. Front. Endocrinol. 2019, 10, 187. [Google Scholar] [CrossRef]
- Posey, A.D., Jr.; Pytel, P.; Gardikiotes, K.; Demonbreun, A.R.; Rainey, M.; George, M.; Band, H.; McNally, E.M. Endocytic recycling proteins EHD1 and EHD2 interact with fer-1-like-5 (Fer1L5) and mediate myoblast fusion. J. Biol. Chem. 2011, 286, 7379–7388. [Google Scholar] [CrossRef]
- Tomasi, M.; Canato, M.; Paolini, C.; Dainese, M.; Reggiani, C.; Volpe, P.; Protasi, F.; Nori, A. Calsequestrin (CASQ1) rescues function and structure of calcium release units in skeletal muscles of CASQ1-null mice. Am. J. Physiol. Cell Physiol. 2012, 302, C575–C586. [Google Scholar] [CrossRef]
- Paolini, C.; Quarta, M.; Nori, A.; Boncompagni, S.; Canato, M.; Volpe, P.; Allen, P.D.; Reggiani, C.; Protasi, F. Reorganized stores and impaired calcium handling in skeletal muscle of mice lacking calsequestrin-1. J. Physiol. 2007, 583, 767–784. [Google Scholar] [CrossRef]
- Bhat, A.; Abu, R.; Jagadesan, S.; Vellichirammal, N.N.; Pendyala, V.V.; Yu, L.; Rudebush, T.L.; Guda, C.; Zucker, I.H.; Kumar, V.; et al. Quantitative Proteomics Identifies Novel Nrf2-Mediated Adaptative Signaling Pathways in Skeletal Muscle Following Exercise Training. Antioxidants 2023, 12, 151. [Google Scholar] [CrossRef] [PubMed]
- Prior, S.J.; Blumenthal, J.B.; Katzel, L.I.; Goldberg, A.P.; Ryan, A.S. Increased skeletal muscle capillarization after aerobic exercise training and weight loss improves insulin sensitivity in adults with IGT. Diabetes Care 2014, 37, 1469–1475. [Google Scholar] [CrossRef] [PubMed]
- Ambery, P.; Adamsson Eryd, S.; Jenkins-Jones, S.; Heywood, B.; Berni, E.; Brown, M.N.; Astbury, C.; Hunt, P.; Currie, C.J. The absolute risk of incident type 2 diabetes following exposure to systemic corticosteroids in selected steroid-related and phenotypic groups. Diabetes Obes. Metab. 2022, 24, 2222–2231. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.J.; Lin, I.H.; Lee, C.W.; Chen, Y.F. Aged Skeletal Muscle Retains the Ability to Remodel Extracellular Matrix for Degradation of Collagen Deposition after Muscle Injury. Int. J. Mol. Sci. 2021, 22, 2123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, Y.; Zhang, H. Extracellular matrix: An important regulator of cell functions and skeletal muscle development. Cell Biosci. 2021, 11, 65. [Google Scholar] [CrossRef] [PubMed]
- Robertson, I.B.; Rifkin, D.B. Regulation of the Bioavailability of TGF-β and TGF-β-Related Proteins. Cold Spring Harb. Perspect. Biol. 2016, 8, a021907. [Google Scholar] [CrossRef] [PubMed]
- Liu, n.; He, S.; Ma, L.; Ponnusamy, M.; Tang, J.; Tolbert, E.; Bayliss, G.; Zhao, T.; Yan, H.; Zhuang, S. Blocking the Class I Histone Deacetylase Ameliorates Renal Fibrosis and Inhibits Renal Fibroblast Activation via Modulating TGF-Beta and EGFR Signaling. PLoS ONE 2013, 8, e54001. [Google Scholar] [CrossRef] [PubMed]
- Shi, A.; Hillege, M.M.G.; Wüst, R.C.I.; Wu, G.; Jaspers, R.T. Synergistic short-term and long-term effects of TGF-β1 and 3 on collagen production in differentiating myoblasts. Biochem. Biophys. Res. Commun. 2021, 547, 176–182. [Google Scholar] [CrossRef]
- Karakilic, A.; Yuksel, O.; Kizildag, S.; Hosgorler, F.; Topcugil, B.; Ilgin, R.; Gumus, H.; Guvendi, G.; Koc, B.; Kandis, S.; et al. Regular aerobic exercise increased VEGF levels in both soleus and gastrocnemius muscles correlated with hippocampal learning and VEGF levels. Acta Neurobiol. Exp. 2021, 81, 1–9. [Google Scholar] [CrossRef]
- Hong, S.M.; Lee, E.Y.; Park, J.; Kim, J.; Kim, S.Y. Aerobic Exercise Ameliorates Muscle Atrophy Induced by Methylglyoxal via Increasing Gastrocnemius and Extensor Digitorum Longus Muscle Sensitivity. Biomol. Ther. 2023, 31, 573–582. [Google Scholar] [CrossRef]
- Plotkin, D.L.; Roberts, M.D.; Haun, C.T.; Schoenfeld, B.J. Muscle Fiber Type Transitions with Exercise Training: Shifting Perspectives. Sports 2021, 9, 127. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.-C.; Sanotra, M.R.; Huang, C.-C.; Hsu, Y.-J.; Liao, C.-C. Aerobic Exercise Modulates Proteomic Profiles in Gastrocnemius Muscle of db/db Mice, Ameliorating Sarcopenia. Life 2024, 14, 412. https://doi.org/10.3390/life14030412
Huang Y-C, Sanotra MR, Huang C-C, Hsu Y-J, Liao C-C. Aerobic Exercise Modulates Proteomic Profiles in Gastrocnemius Muscle of db/db Mice, Ameliorating Sarcopenia. Life. 2024; 14(3):412. https://doi.org/10.3390/life14030412
Chicago/Turabian StyleHuang, Yen-Chun, Monika Renuka Sanotra, Chi-Chang Huang, Yi-Ju Hsu, and Chen-Chung Liao. 2024. "Aerobic Exercise Modulates Proteomic Profiles in Gastrocnemius Muscle of db/db Mice, Ameliorating Sarcopenia" Life 14, no. 3: 412. https://doi.org/10.3390/life14030412