2.1. Cultivation of Hairy Roots
H. neglectum seeds purchased from Sady Rossii (Chibanovo village, Krasnoarmeisky district, Chelyabinsk region, Russia) were used to obtain hairy roots. Seeds were sterilized in 5 cm
3 of solution (3% hydrogen peroxide and 96% ethanol in a 1:1 ratio) for 10 min at 25 ± 2 °C, then washed repeatedly with sterile distilled water and transferred for germination in Petri dishes on Murashige–Skoog solid medium (MS) [
35] in dark conditions at 25 ± 0.5 °C for two days [
35]. To produce hairy roots, 25–35 explants were used.
The seedlings were perforated with a sterile needle and incubated for 24 h in liquid MS medium containing a suspension of
Agrobacterium rhizogenes ATCC 15834 [
19]. A. rhizogenes bacteria were purchased from the collection of the Siberian Institute of Plant Physiology and Biochemistry, Siberian Branch of the Russian Academy of Sciences (Irkutsk, Russia).
The agrobacterial strains used in this study were grown on YEB medium for 48 h at 28 ± 0.5 °C [
36]; a suspension of agrobacteria with an optical density of 0.8–1 determined at a wavelength of 600 nm was used to infect seedlings [
37]. The composition of the YEB medium is presented in the article of G. Vervliet [
38]. Spectrophotometric measurements of bacterial density were performed on a ClaioStar device (BMG, Offenburg, Germany).
After the indicated time,
H. neglectum infected with agrobacteria were washed with sterile distilled water and placed on solid nutrient media, the composition of which is shown in
Table 1, with the addition of 500 mg/dm
3 of claforan (Thermo Fisher Scientific Inc., Bourgoin, France). Cultivation parameters: 25 ± 1 °C with 16.0 h photoperiod (cold white fluorescent lamps, illumination—2–3 klx). Experiments were transferred every three days until complete elimination of agrobacteria. Sterility of cultured roots was checked by incubating root samples in liquid YEB medium for 48 h at 26 ± 2 °C. The appearance of primary roots was observed 14–28 days after transformation.
Primary roots were separated and transplanted to agar medium with 250 mg/dm
3 of claforan. The hairy roots were then transferred to antibiotic-free liquid medium and transplanted every three weeks by inoculating the crude root mass into conical flasks containing liquid nutrient medium [
24,
36,
39]. Plant material was cultivated in the dark at 23 ± 0.5 °C on an orbital shaker (shaker incubator ES-20/80, BioSan, Riga, Latvia) at 90 rpm.
The compositions of the MS and B5 (Gamborg) media are presented in the articles by T. Murashige and F. Skoog [
35] and O. L. Gamborg et al. [
40], respectively. The literature review suggests that the ratios of inoculant to nutrient medium should be 1:100 and of flask volume to medium should be 5:1. When the hairy roots reached a size of 10–15 cm, they were separated from the plant and transferred to a new medium with the same components and subcultured every 30 days. The cultivation cycle was 5 weeks [
24,
36,
39].
Explants punctured with a sterile needle but not infected with agrobacteria served as controls. Infected explants cultured on nutrient media without the addition of hormones were also used as controls. All experiments were performed in triplicate.
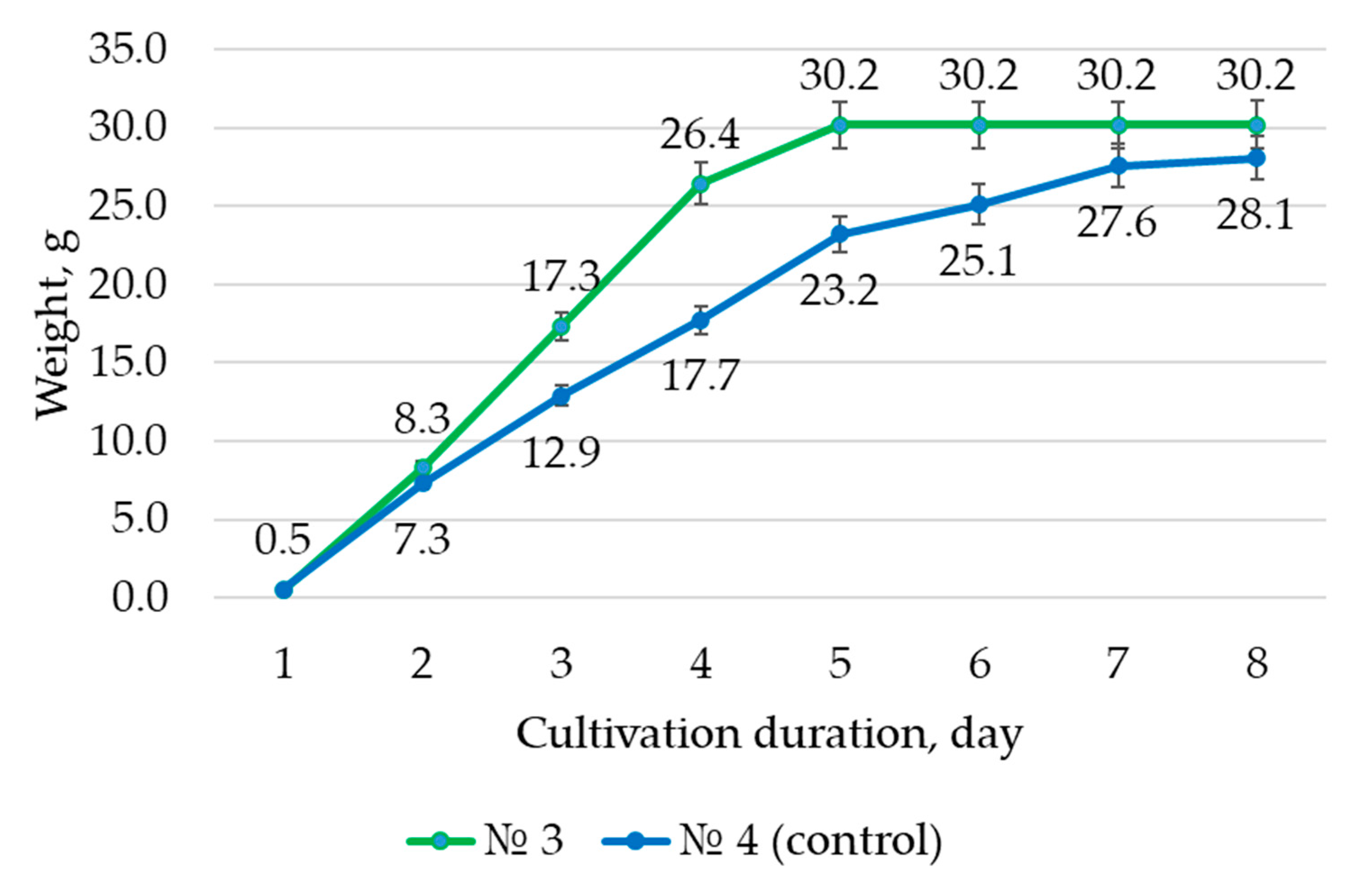
The difference in root dry mass at the end of cultivation and at the start of cultivation was used to calculate the growth index (I), which was calculated using Formula (1) [
24]. Hairy root biomass was dried to constant weight in an air stream at 30 ± 0.5 °C in a desiccator (ShSvL-80—Kasimov (Kasimov priborniy zavod, Kasimov, Russia).
where X
max—mass at the end of the cultivation cycle, g;
X0—mass at the beginning of cultivation, g.
Appropriate nutrient media, indicated in
Table 1, were used to maintain grown cultures.
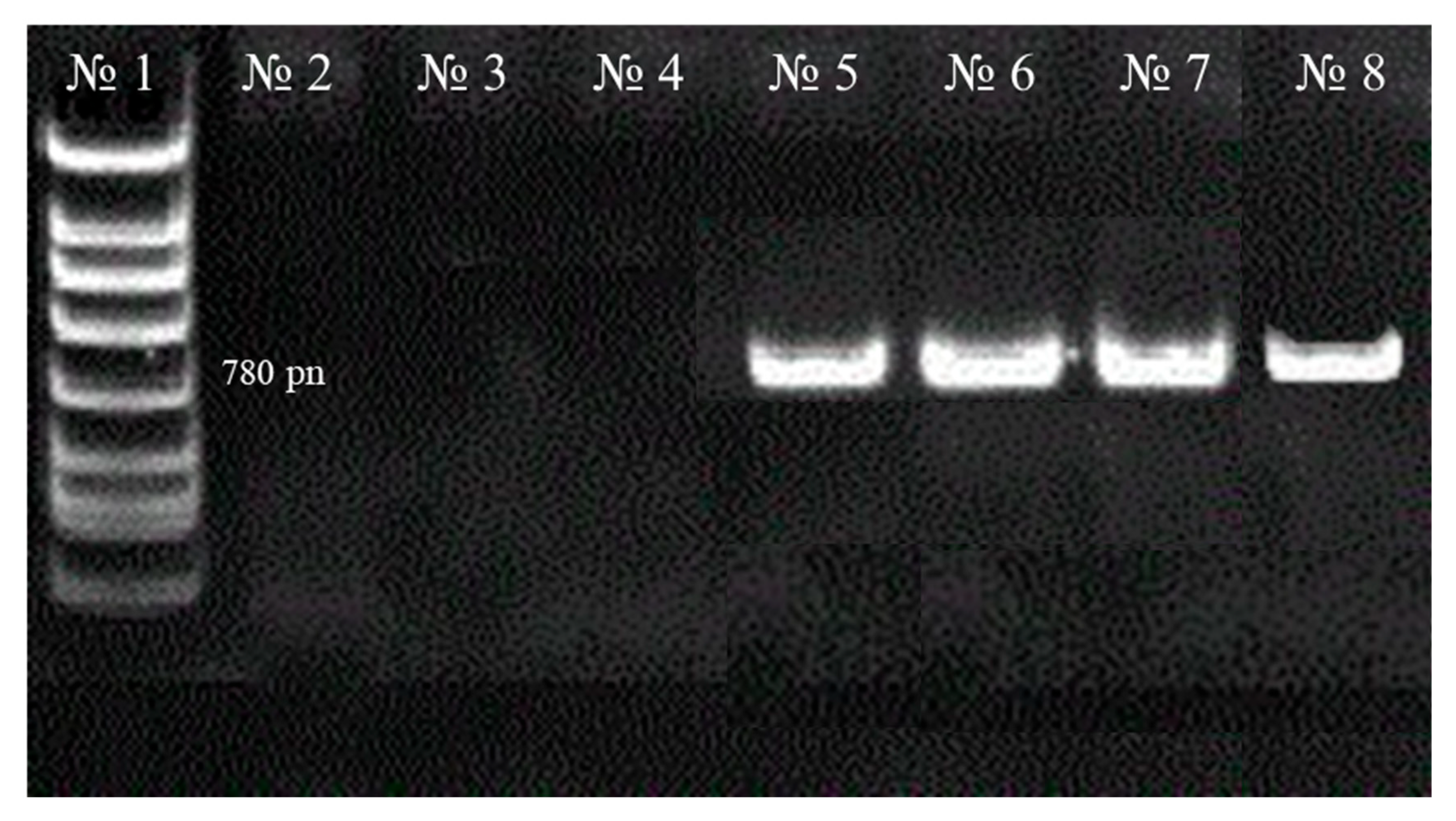
Total genomic DNA was isolated from biosamples for molecular genetic analysis of the obtained hairy roots (to detect the integration of T-DNA plasmid Ri into the transformed roots (
rolB gene). For DNA isolation, a commercial DNA-EXTRAN-3 kit (NPF Sintol, Moscow, Russia) was used. PCR analysis was performed on a TP4-PCR-01—Tertsik DNA amplifier (NPO DNK-Tekhnologiya, Moscow, Russia). Primers and specific amplification parameters for the
rolB gene were previously reported by R. K. Tiwari et al. [
37] and A. Stojakowska et al. [
36] (
Table 2).
Non-transformed DNA from the seedlings of the studied plants was used as a negative control for PCR analysis. PCR products (amplified DNA) were analyzed using electrophoretic separation on a 1% agarose gel and visualized in ultraviolet light after staining with ethidium bromide. Complex consisting of mixture A and B for amplification and primers was purchased from OOO NPF Sintol, Russia.
The biomass of hairy roots for further studies was accumulated by cultivating on liquid nutrient media, i.e., media of similar composition without the addition of agar–agar.
2.2. Preparation of Extracts from Hairy Roots Biomass, Evaluation of the Qualitative and Quantitative Composition of Metabolites
Filtration through a sterile paper filter (Yellow Ribbon GOST 12026-76 (Ekokhim, Novosibirsk, Russia) using a Buechner funnel under vacuum was used to separate the hairy roots from the culture liquid. Hairy root biomass was dried to constant weight in an air stream at 30 ± 0.5 °C in a desiccator (ShSvL-80—Kasimov (Kasimov priborniy zavod, Russia)). The dried biomass was ground in an LZM-1M mill (Olis, Barnaul, Russia) and sieved through a sieve with 1–3 mm apertures.
The plant material was extracted using a PE-4310 water bath (Ekroshim, Novosibirsk, Russia) with a reflux condenser. The extractant raw material was first exposed to ultrasound (US) for 15 min at a frequency of 40 kHz without allowing the water to heat above 40 °C, followed by extraction in a water bath with a reflux condenser. Ultrasonic (US) treatment was performed using a Stegler 3DT ultrasound bath (STEGLER, Taiwan, China) at an ultrasound frequency of 40 kHz. The extract was separated from the sediment of plant material using filtration with a sterile paper filter (Yellow Ribbon GOST 12026-76 (Ecohim, Saint Petersburg, Russia) and a Buechner funnel under vacuum.
Table 3 presents the extraction parameters.
The mechanical effect of ultrasonic treatment promotes diffusion (penetration of the extractant into the cells of raw materials), destruction of cell walls, and accelerated release of their contents, which was discovered to be a relatively simple method of green chemistry. That is, when compared to the treatment without ultrasound, the method allows for a reduction in the duration and temperature of the BAS extraction process (without changing/increasing the yield of the extract) from plant raw materials [
44,
45].
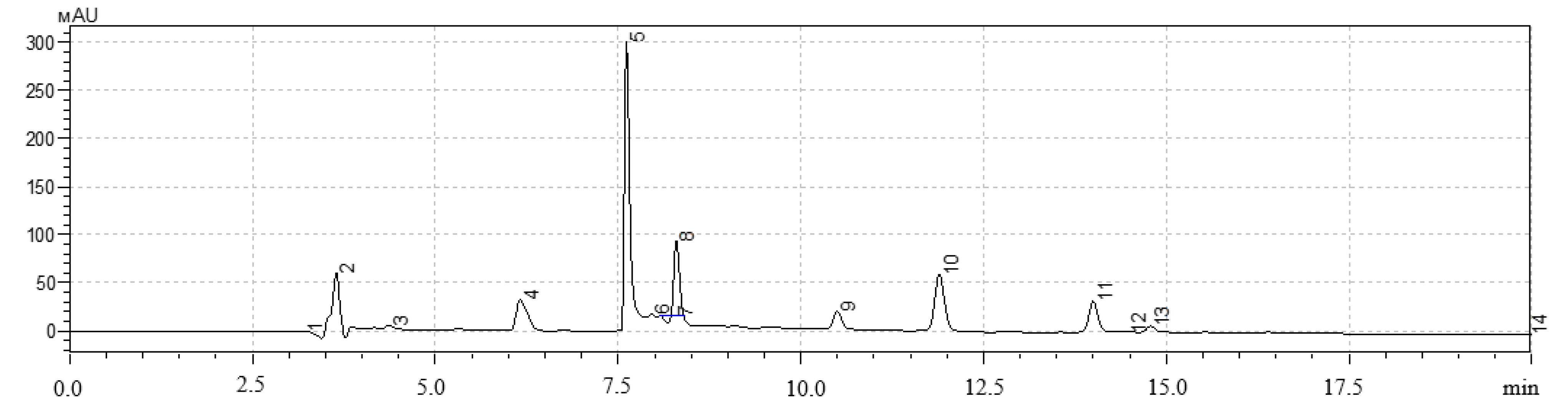
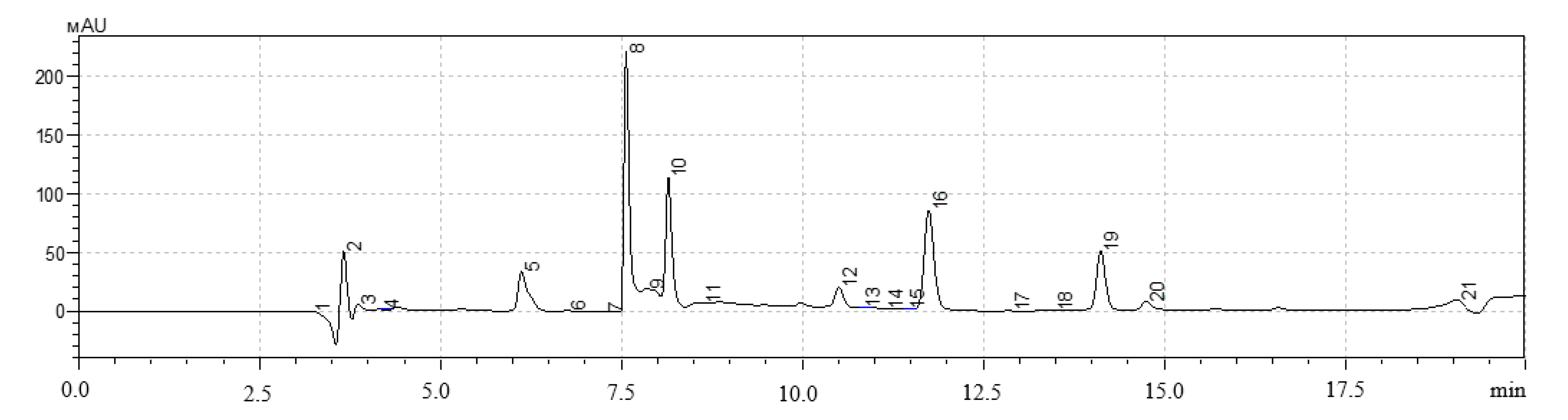
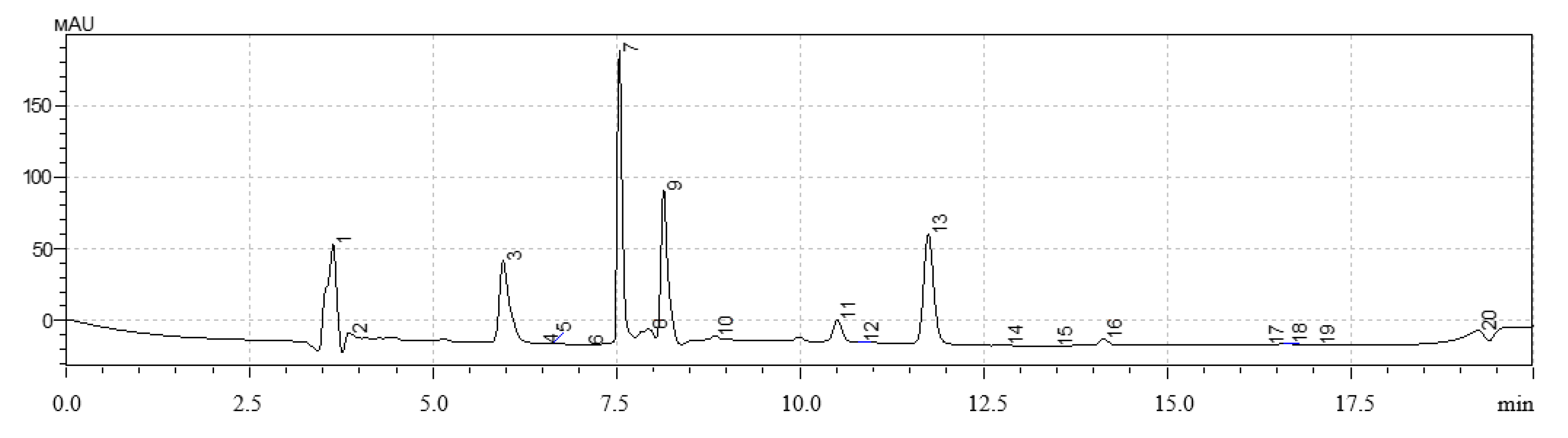
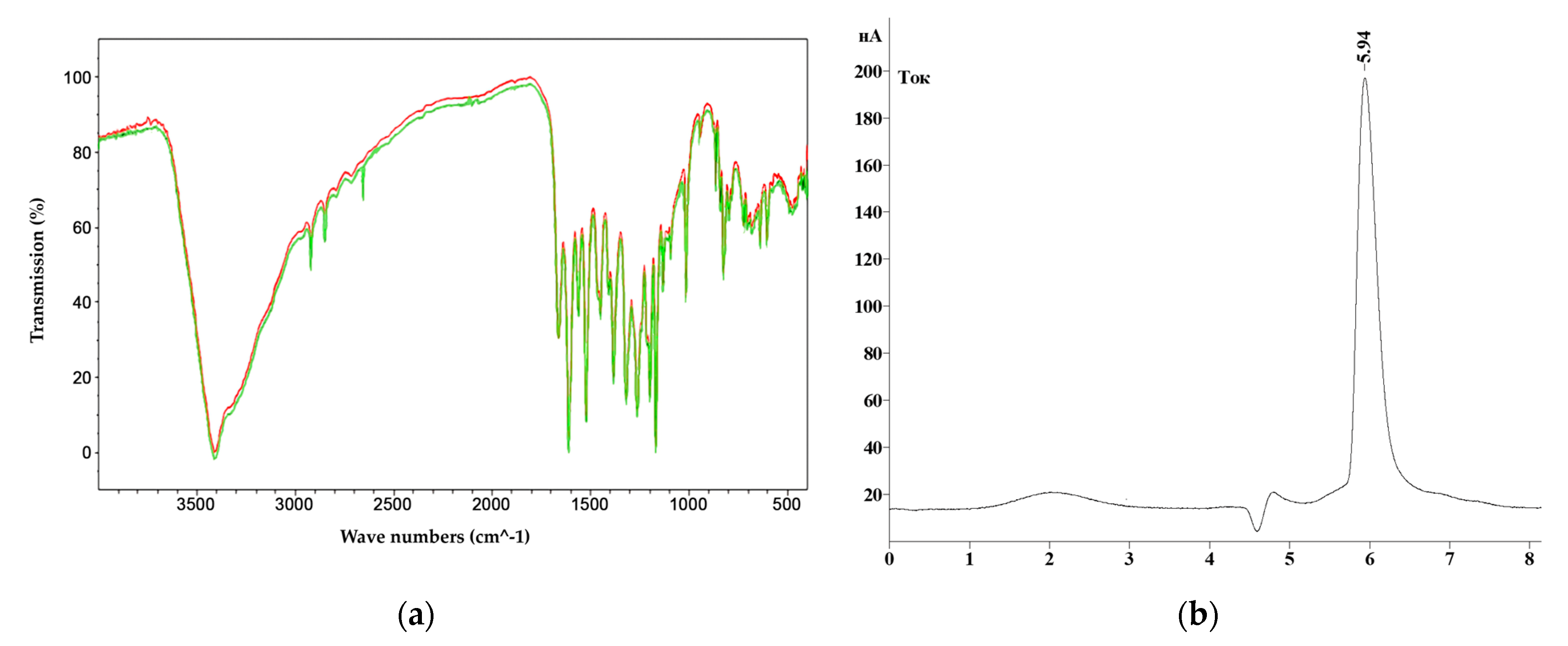
A high-performance liquid chromatography (HPLC) method was used to determine the qualitative and quantitative content of polyphenols in extracts of hairy roots. The analysis was performed on a Shimadzu LC-20 Prominence chromatograph with a Shimadzu SPD20MA diode-matrix detector and a Phenomenex Gemini C-18 250 × 4.6 mm column (Shimadzu, Kyoto, Japan).
H. neglectum hairy root extracts were vacuum dried to a dry residue on an IKA RV 8 rotary evaporator (IKA®-Werke GmbH & Co. KG, Germany) at 45 ± 0.5 °C. Dry extract in the amount of 1 ± 0.05 g was added to 10 ± 0.01 cm3 of 1.5 M HCl solution. The resulting mixture was subjected to acid hydrolysis for 60.0 ± 0.5 min in a water bath with a reflux condenser (95 ± 5 °C). Then, the mixture was neutralized to pH = 7.0 by adding 1.0 M NaOH solution, filtered through a Yellow Ribbon GOST 12026-76 paper filter (Ecohim, Saint Petersburg, Russia), and then through a membrane filter with a pore size of 0.45 μm (Membrane Solutions, Auburn, WA, USA).
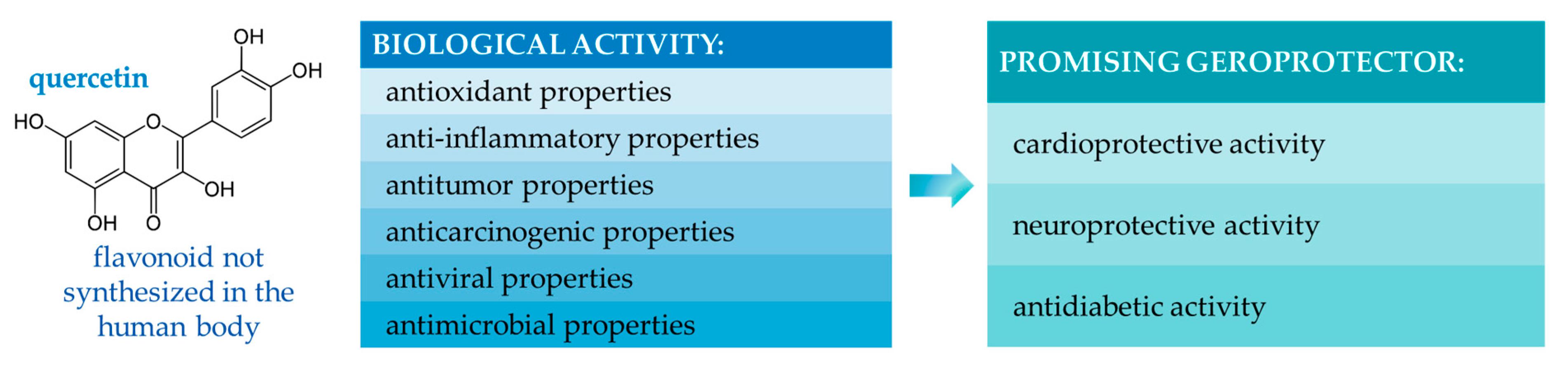
The gradient chromatography mode was used for all samples. Components of the mobile phase: acetonitrile, isopropyl alcohol, deionized water with the addition of orthophosphoric acid to pH = 3.5. Injection volume was 20 mm3, elution rate was 0.8 cm3/min, column temperature was 30 °C. The quantitative content was determined by the absolute graduation method using 98% purity standards: gallic acid, quercetin, protocatechic acid, chlorogenic acid, and caffeic acid (Sigma-Aldrich, Saint-Louis, MO, USA).
Quantitative analysis of the polyphenols under study was performed using calibration curves plotted in the concentration range of 1.9–235 µg/cm
3. The equation for the calibration curves is presented in Formula (2).
where X—standard concentration (µg/cm
3);
Y—corresponding peak area according to HPLC results;
a—factor of proportionality.
Formula (3) relates the peak area to a unit of dry mass.
where C—flavone content in a sample of dry material (mg/g);
m—mass of dry material (g);
a—factor of proportionality from the equation of the calibration curve; factor 1000 is necessary to convert C to mg/g.
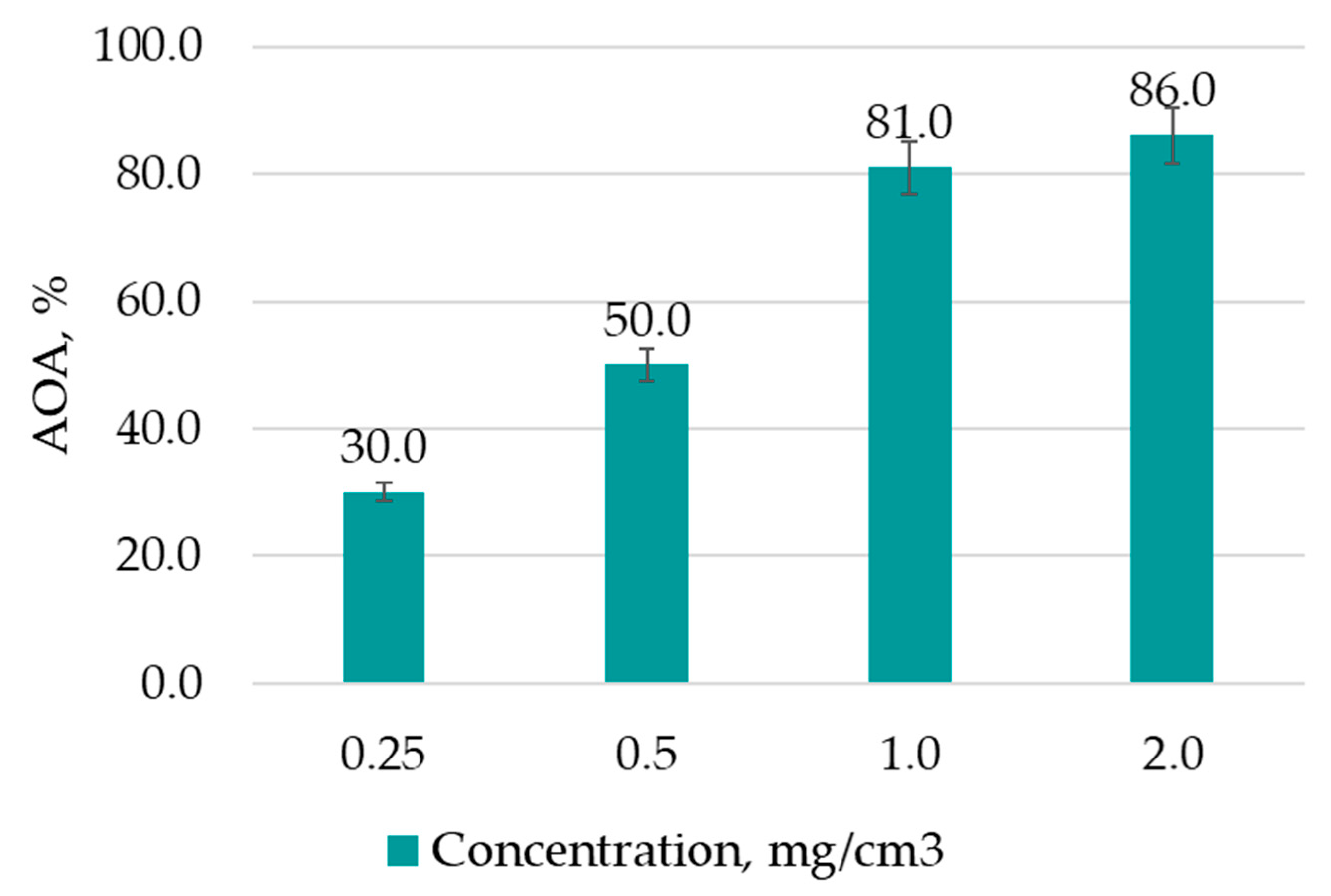
2.4. Study of the Antioxidant Activity of Hairy Root Extracts and the Bioactivity of Quercetin Solutions
The antioxidant activity (AOA) of the obtained extracts and quercetin solutions was evaluated using the 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) ABTS•+ cation radical decolorization method [
46] on a UV 1800 spectrophotometer (Shimadzu, Japan):
Reagent ABTS (AppliChem GmbH, Darmstadt, Germany) was dissolved in water to a concentration of 7.0 mM (0.18011 g in 50 cm3 of water in a volumetric flask).
ABTS cation radical (ABTS•+) was obtained by reacting the stock solution of ABTS with 2.45 mM potassium persulfate (0.03311 g in 50 cm3 water in a measuring flask) and incubating the mixture in the dark at room temperature for 12–16 h before use. They were mixed in the ratio of ABTS:persulfate—2:1.
The ABTS•+ solution was diluted with ethanol to an optical density of 0.70 ± 0.02 at 734 nm.
To study the AOA, 3 cm3 of ABTS•+ was mixed with 30 mm3 of sample solution (100:1 ratio).
Control for ABTS•+ density determination—pure solvent. Control for samples—ABTS•+ without antioxidant. Formula (4) was used to calculate the AOA.
where A
ABTS—optical density of the initial ABTS solution;
Ax—optical density of ABTS solution + test sample.
The obtained AOA results were compared to the activity of different concentrations of vitamin C solutions (control). The extract with the highest AOA was chosen based on the results, and the target BAS were isolated from it.
The BAS under study were dissolved in a 70% aqueous–ethanol solution to obtain a 1 M stock solution [
47], and solutions with concentrations of 800 μM, 600 μM, 400 μM, and 200 μM were prepared by dilution in water.
The bioactivity of the studied BAS isolated from extracts of plant hairy roots was tested on model organisms of C. elegans nematodes. The C. elegans N2 Bristol strain was provided by the Laboratory of Innovative Drug Development and Agrobiotechnology of the Moscow Institute of Physics and Technology (National Research University, Russia).
For the study, stock solutions of BAS in dimethyl sulfoxide (DMSO) at a concentration of 10 mM were prepared. Then, in distilled water, solutions with concentrations of 2000 μM, 1000 μM, 500 μM, and 100 μM were obtained. These solutions in an amount of 15 mm3 were added to the wells of the plate, thus obtaining test solutions of 200 µM, 100 µM, 50 µM, and 10 µM BAS concentration.
Pregnant adult nematodes were removed after 3 h of egg-laying on nematode growth medium (NGM) dishes in order to produce synchronized larvae by the age of 5–10. The dishes with eggs were treated with an aqueous NaCLO solution (salt:water ratio of 1:5). Eggs were cultivated on S medium at 20 ± 0.2 °C until hatching. The obtained synchronized L1-stage worms were placed on standard NGM agar dishes with
Escherichia coli OP50 and incubated until the larval L4 stage [
48]. The composition of the NGM and S medium is reflected in the work by T. Stiernagle [
49].
E. coli strain OP50 was used as food for nematodes, provided by the V. Engelhardt Institute of Molecular Biology RAS (Russia). To obtain a starter culture of E. coli strain OP50, one colony was grown in 5 cm3 of LB medium (AppliChem GmbH, Germany) for 12 ± 0.5 h at 37 ± 0.5 ℃ under vigorous stirring (120 rpm).
Synchronized L4-stage larvae were transferred to NGM agar plates containing
E. coli OP50 suspension at 20 ± 0.5 °C for 3 days. After the specified time, the larvae are in the L4 stage, to which 0.5 mg/cm
3 of 5-fluoro-2-deoxyuridine (FUdR), a substance that inhibits nematode reproduction, is added, eliminating the possibility of obtaining a mixed worm population. After cultivating for 1 day at 20 ± 0.5 °C, the worms were transferred to liquid nutrient medium—S-medium. The experiments were performed in 96-well plates with a flat transparent bottom (Merck KGaA, Darmstadt, Germany), to which 150 mm
3 of nematode and
E. coli OP50 suspensions (bacteria concentration in the suspension: 0.5 mg/cm
3) and 15 mm
3 of different concentrations of tested BAS solutions were added [
50,
51].
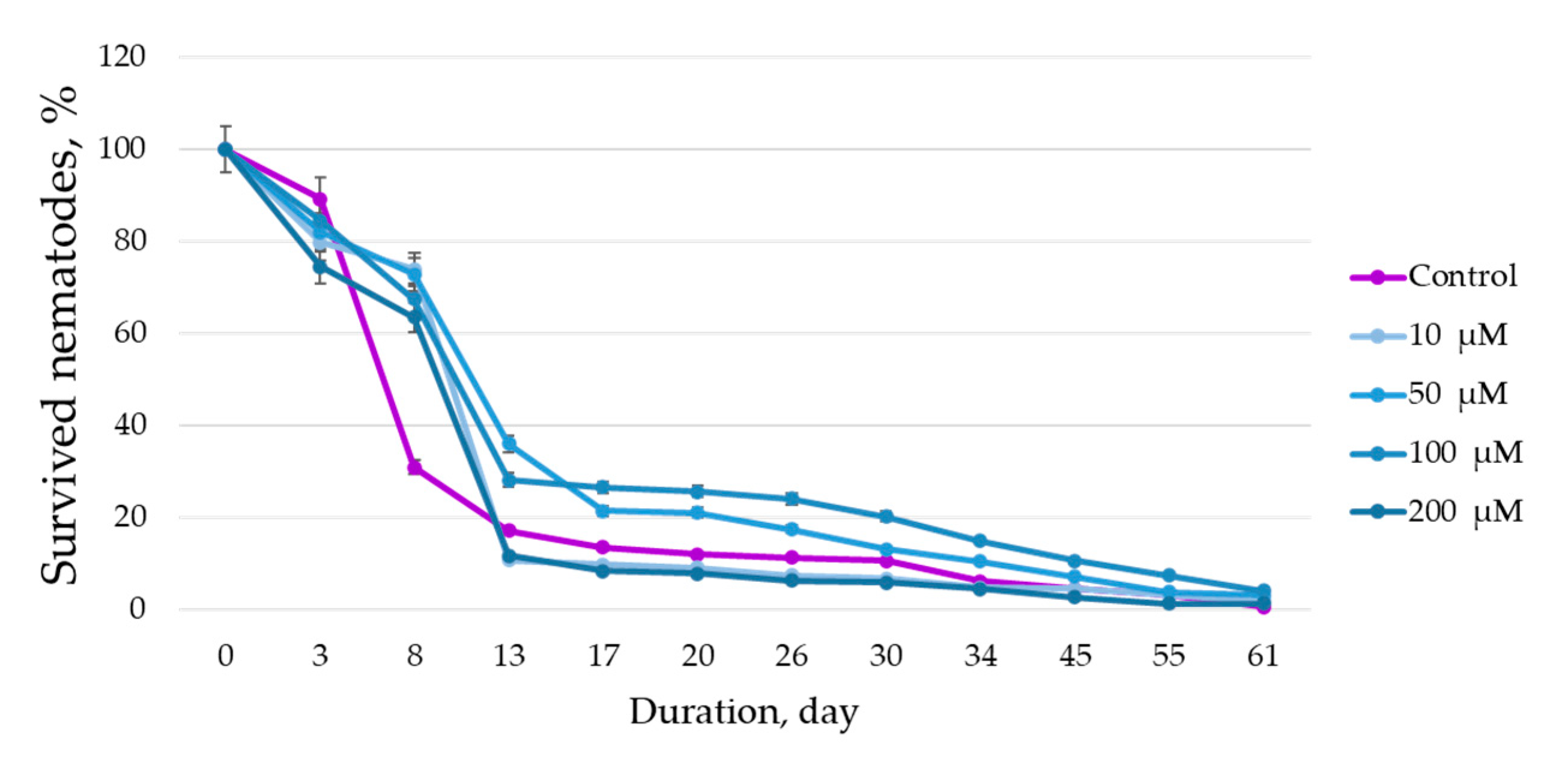
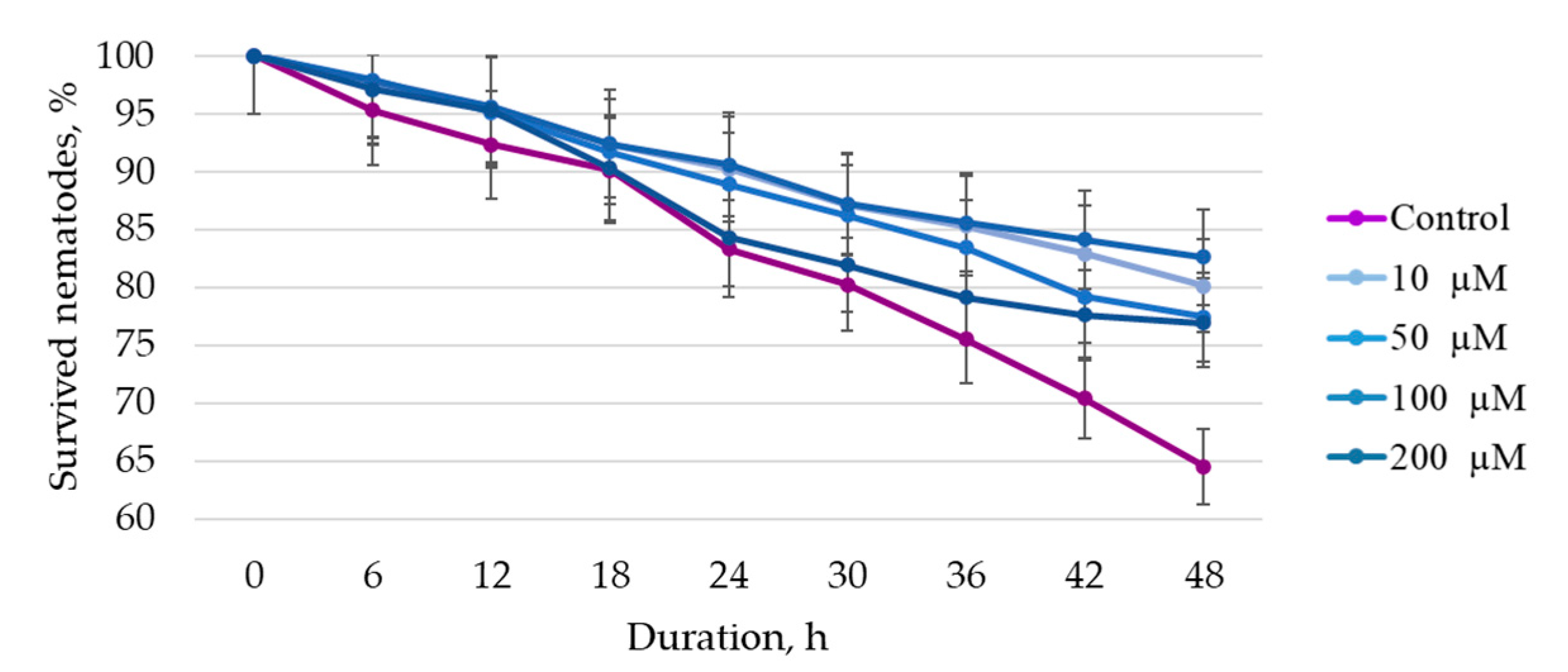
To assess the life span of the nematode plates, they were cultivated at 20 ± 0.5 °C for 61 days. To assess the effect of BAS solutions on the stress tolerance of
C. elegans, 15 mm
3 of 1 M paraquat (a substance that simulates oxidative stress) were added to each well of the plate and the cultivation was continued at 20 ± 0.5 °C for 48 h in 6 h increments. After the specified time, live and dead nematodes were counted. Assessment was performed using an Axio Observer Z1 microscope (Karl Zeiss, Oberkochen, Germany). The criterion for the death of nematodes was the complete absence of motor activity when they were exposed to the bright light of the microscope beam. Nematodes suffering from internal hatching (embryos hatching inside an adult hermaphrodite) that escaped from the cups were censored (removed from the experiment). Experiments were performed in sterile conditions of a UVC/T-AR box (BioSan, Latvia). In more detail, the methodology of life span and stress tolerance assessment, the composition of nutrient media, etc., are presented in the paper by F. R. Amrit et al. [
27].
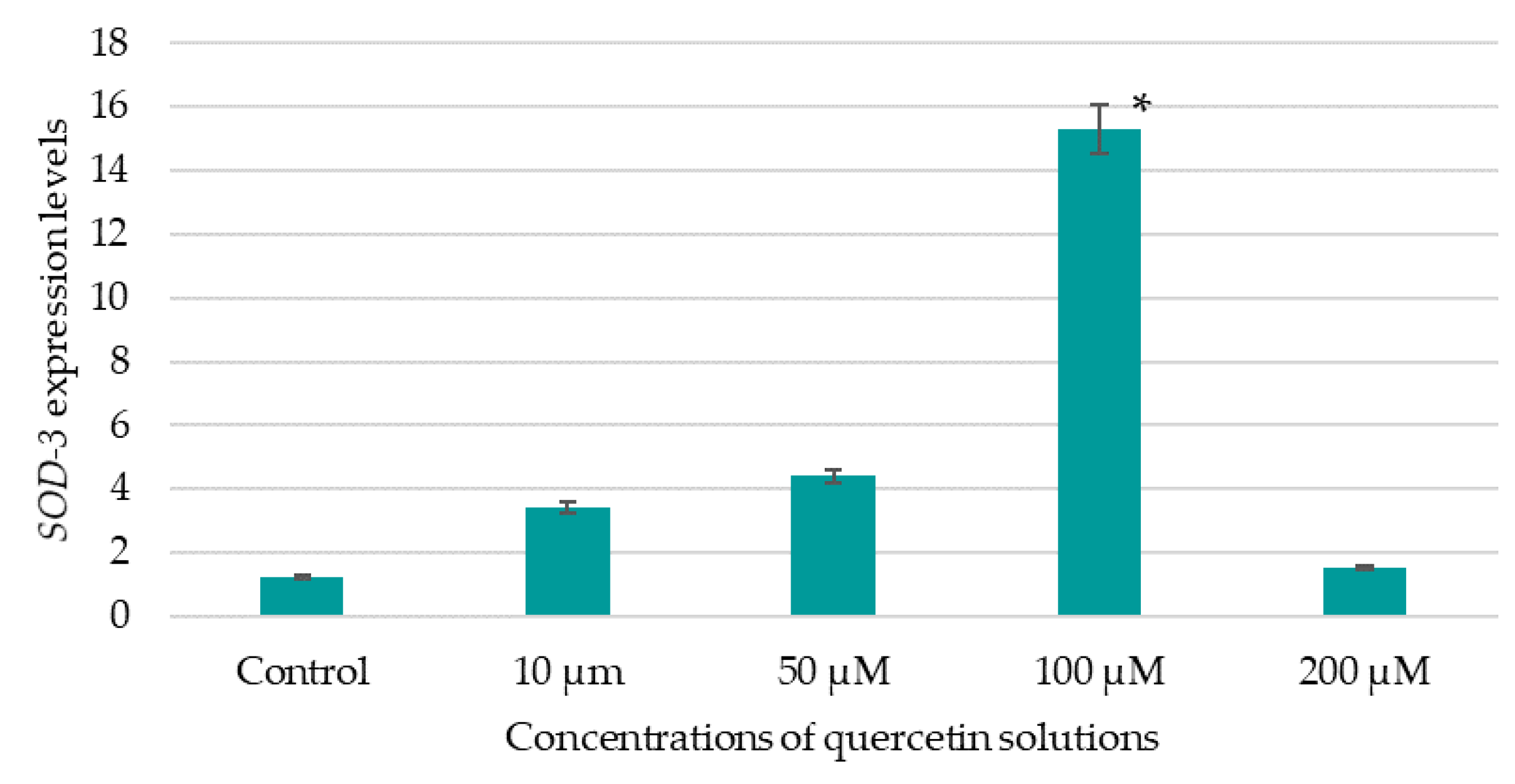
A 24-well plate with 900 mm
3 of L1-stage nematode suspension and 100 mm
3 of BAS solution was used to evaluate the expression level of the
SOD-3 gene. The plate was left for 72 h at 20.0 ± 0.5 °C until the nematodes reached the L4 stage of development, and then the plate was placed in an incubator at 35 ± 0.5 °C for 5.0 h. After the specified time, the plate was cooled on ice and the content of the well was transferred to an Eppendorf tube and centrifuged for 2 min at 1000
g to remove the supernatant. The resulting precipitate was immediately frozen at 86 °C below zero until ribonucleic acid (RNA) was isolated. RNA was isolated from nematodes using the Exstract RNA kit according to the manufacturer’s instructions (Eurogen, Moscow, Russia). The quality of isolated RNA was assessed using a Nanodrop 2000C RNA concentration meter (Thermo Scientific, Waltham, MA, USA). Synthesis of the first cDNA chain was performed using the MMLV RT kit according to the manufacturer’s instructions (Eurogen, Russia). RNA amplification was performed on a CFX96 RealTime System (BioRad, USA) using a one-step approach with a 5X qPCRmix-HS SYBR reaction mixture designed for real-time PCR with SYBER Green I dye. The amplification steps are shown in
Table 4.
The expression of the SOD-3 gene in nematodes treated with the tested BAS solutions was compared to the expression of these genes in control samples—nematodes that had not been treated with the tested compounds. The quantitative gene expression levels were normalized relative to the expression levels of the AMA or ACT reference genes, which were determined using the same real-time amplification conditions and amount of cDNA tested.
Relative gene expression (RE) was calculated using Formula (5).
where E—amplification efficiency (equal to 2.0).
Normalization by the mean of the reference genes is presented in Formula (6).
where C(t)
reference—geometric mean (C(t)) of reference genes (for mRNA);
C(t)target—C(t) of target genes.
The calculation of RE is presented in more detail in the paper by T.A. Dimitriadi et al. [
52].
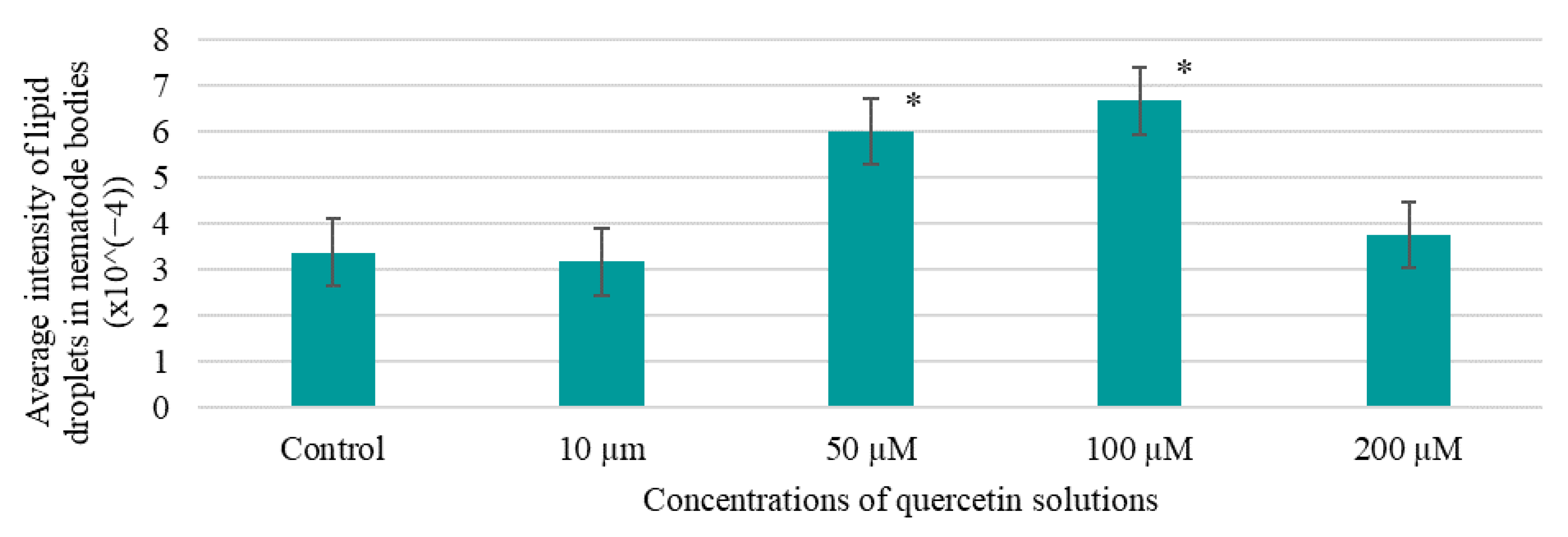
Fluorescent staining of lipid inclusions using the BDP 505/515 reagent bordipyrromethane fluorophore (Lumiprobe, Moscow, Russia) to visualize lipid membranes was used to assess the effect of BAS on fat accumulation in nematodes [
34]. At the end of 10 days of nematode incubation in the presence of BAS, the contents of the 96-well flat-bottomed plate were transferred to the corresponding wells of the 96-well V-well plate. The plate was centrifuged at 1000 rpm for 3 min, and then the supernatant was removed without affecting the nematode precipitate. To wash the nematodes from the culture fluid, 150.0 mm
3 of sterile PBS (8.0 g NaCl + 0.2 g KCl + 1.44 g Na
2HPO
4 + 0.24 g KH
2PO
4 dissolved in 1.0 dm
3 water) were added to each cell and centrifuged at 1000 rpm for 3 min. The washing process was repeated twice. In each cell, 150 mm
3 of fixative solution (40.0% isopropanol solution) were added to the washed nematode precipitate. The plates with nematodes were incubated for 20 min at 24.0 ± 2.0 °C. After this time, the plate was centrifuged at 1000 rpm for 3 min. Then, 150 mm
3 of BDP 505/515 fat staining reagent working solution (2 mm
3/cm
3 of stock solution in 40% isopropanol) were added to each cell of the plate. The plate with nematodes was incubated for 15 min at 50 rpm 24.0 ± 2.0 °C. Then, it was centrifuged at 1000 rpm for 3 min and the supernatant was removed. The nematode precipitate was washed with sterile M9T solution (50 cm
3 of M9 buffer (3 g KH
2PO
4 + 6 g Na
2HPO
4 + 5 g NaCl, autoclaving 20 min, 120 °C, then adding 1 cm
3 of 1 M MgSO
4 solution) with 5 mm
3 Triton X-100) three times. After washing was completed, 100 mm
3 of M9T were added to the wells and the stained lipid inclusions in the nematode bodies were scanned using an ImageXpress Mico XL automated fluorescence microscope (Molecular Devices, Silicon Valley, CA, USA) [
34].




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}