
Coastal Wetland Species Rumex hydrolapathum: Tolerance against Flooding, Salinity, and Heavy Metals for Its Potential Use in Phytoremediation and Environmental Restoration Technologies
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Cultivation, and Experimental Design
2.2. Physiological Measurements
2.3. Termination of the Experiments and Measurements
2.4. Data Analysis
3. Results
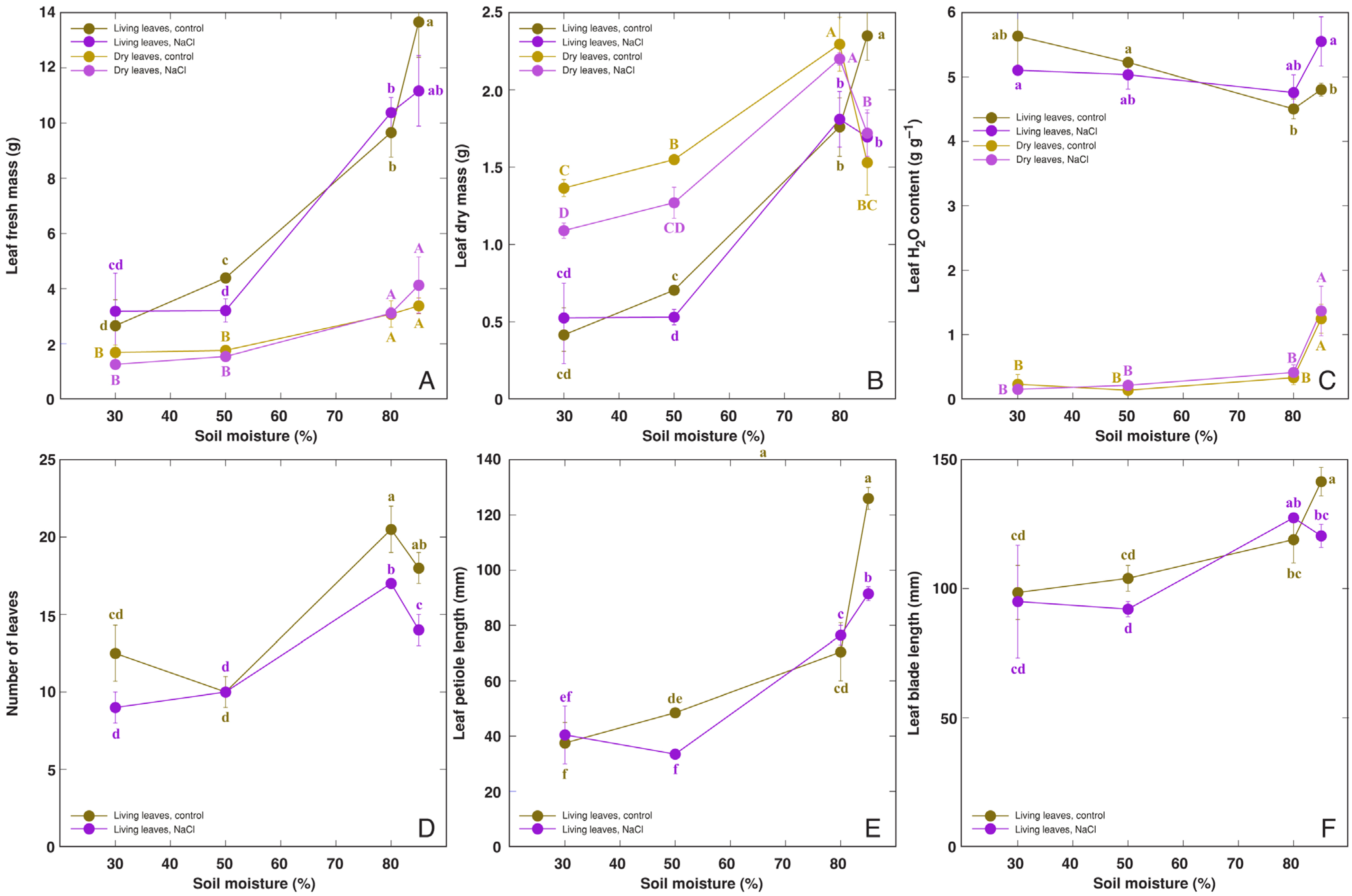
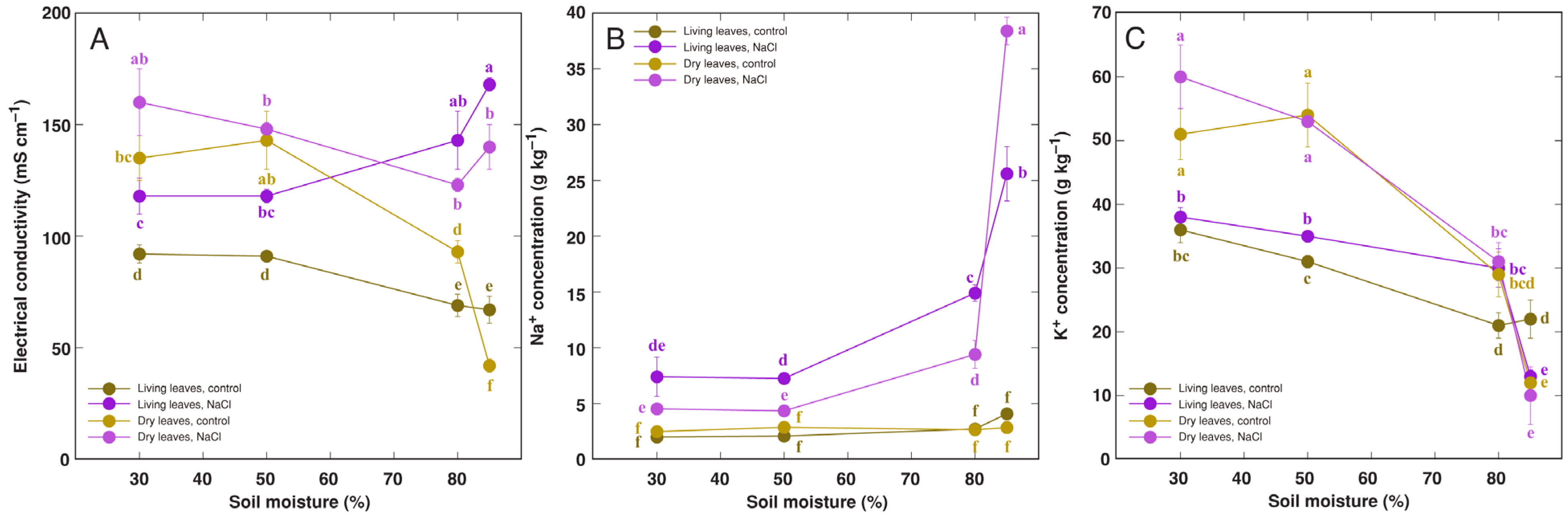
3.1. Effect of Soil Moisture and NaCl
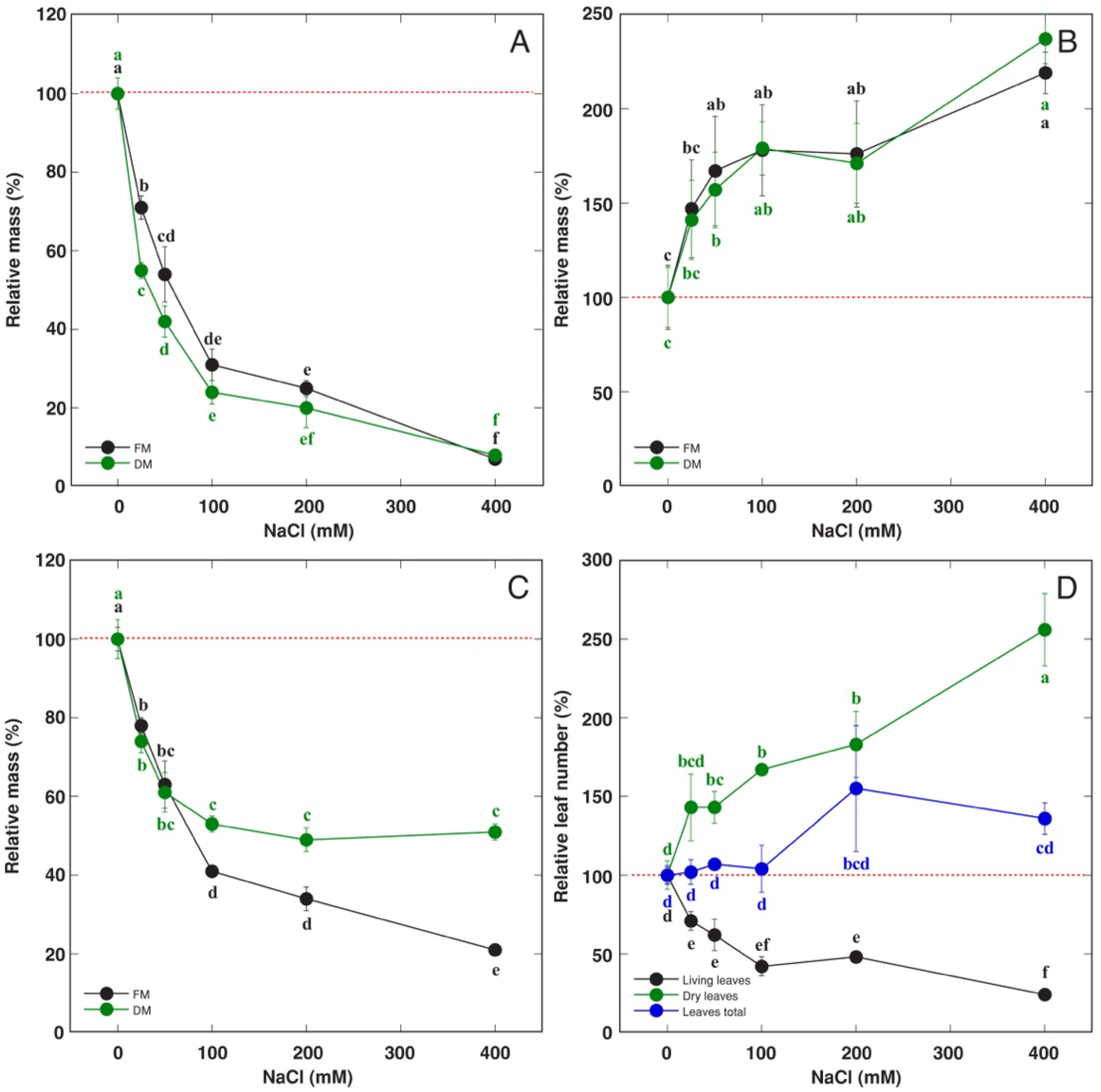
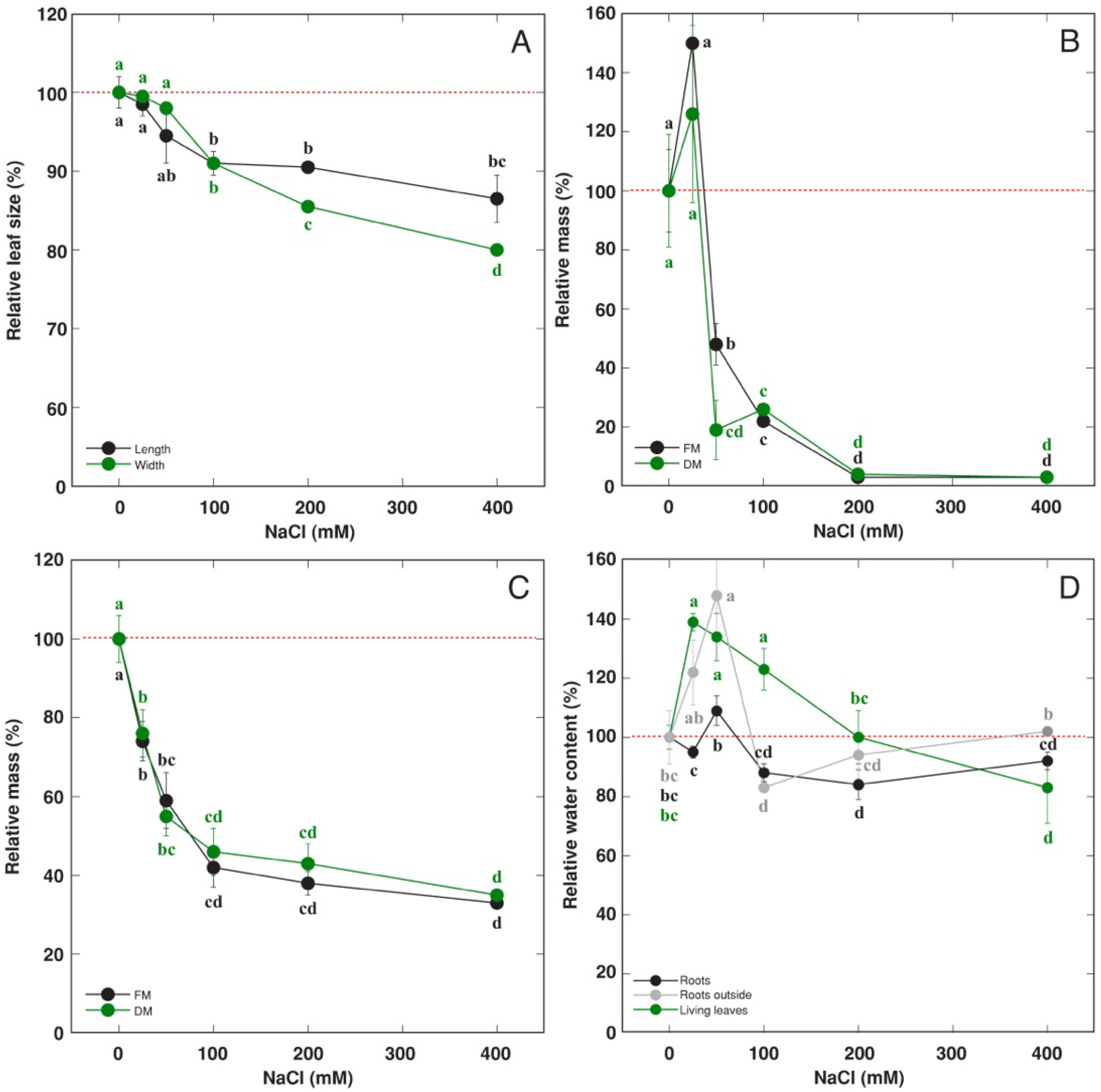
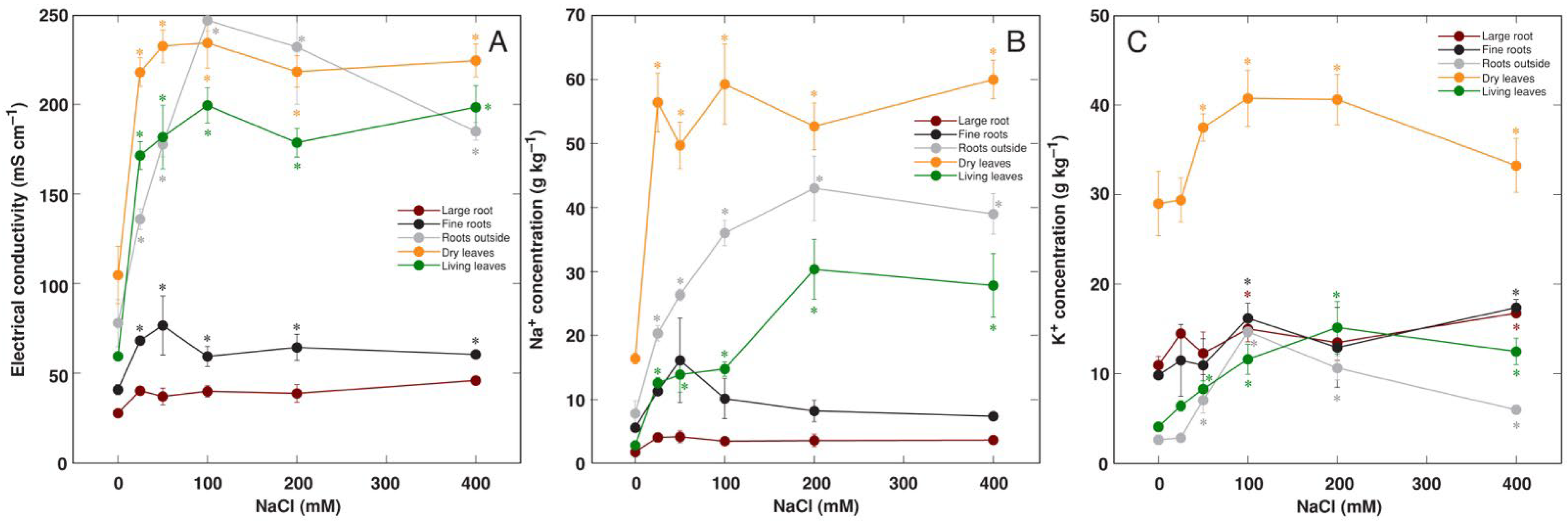
3.2. Effect of Soil Waterlogging with NaCl
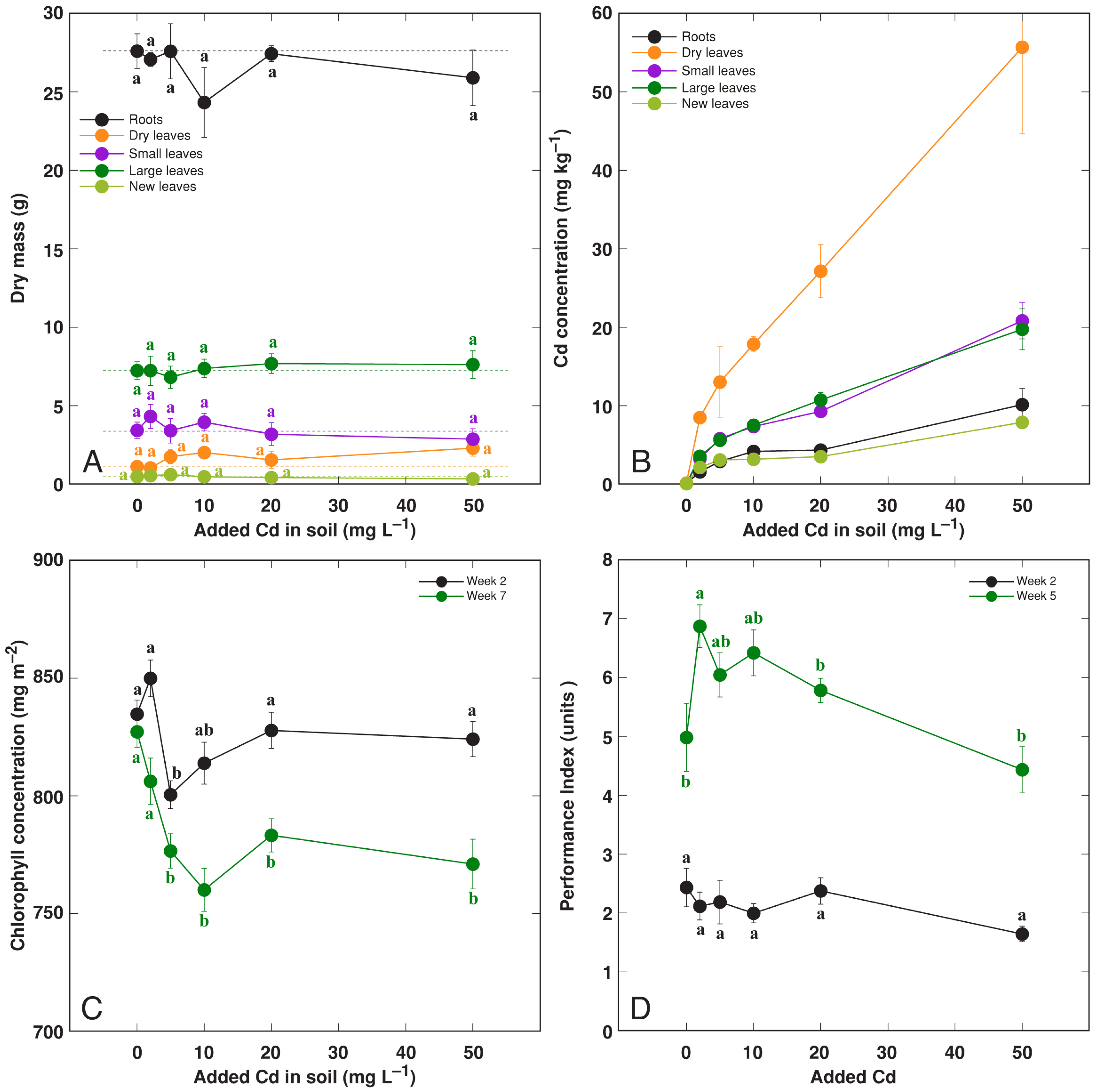
3.3. Effect of Heavy Metals: Cd
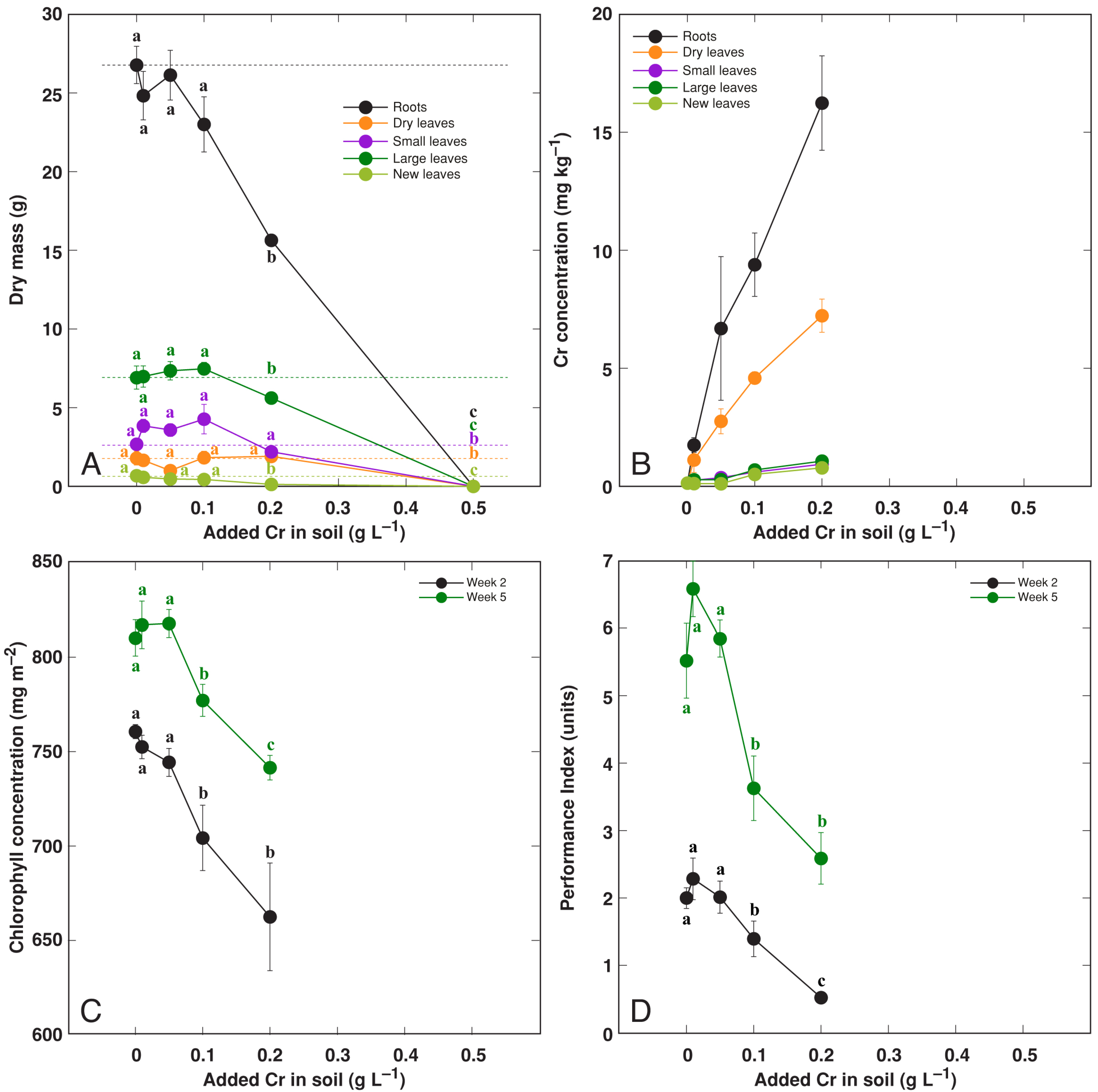
3.4. Effect of Heavy Metals: Cr
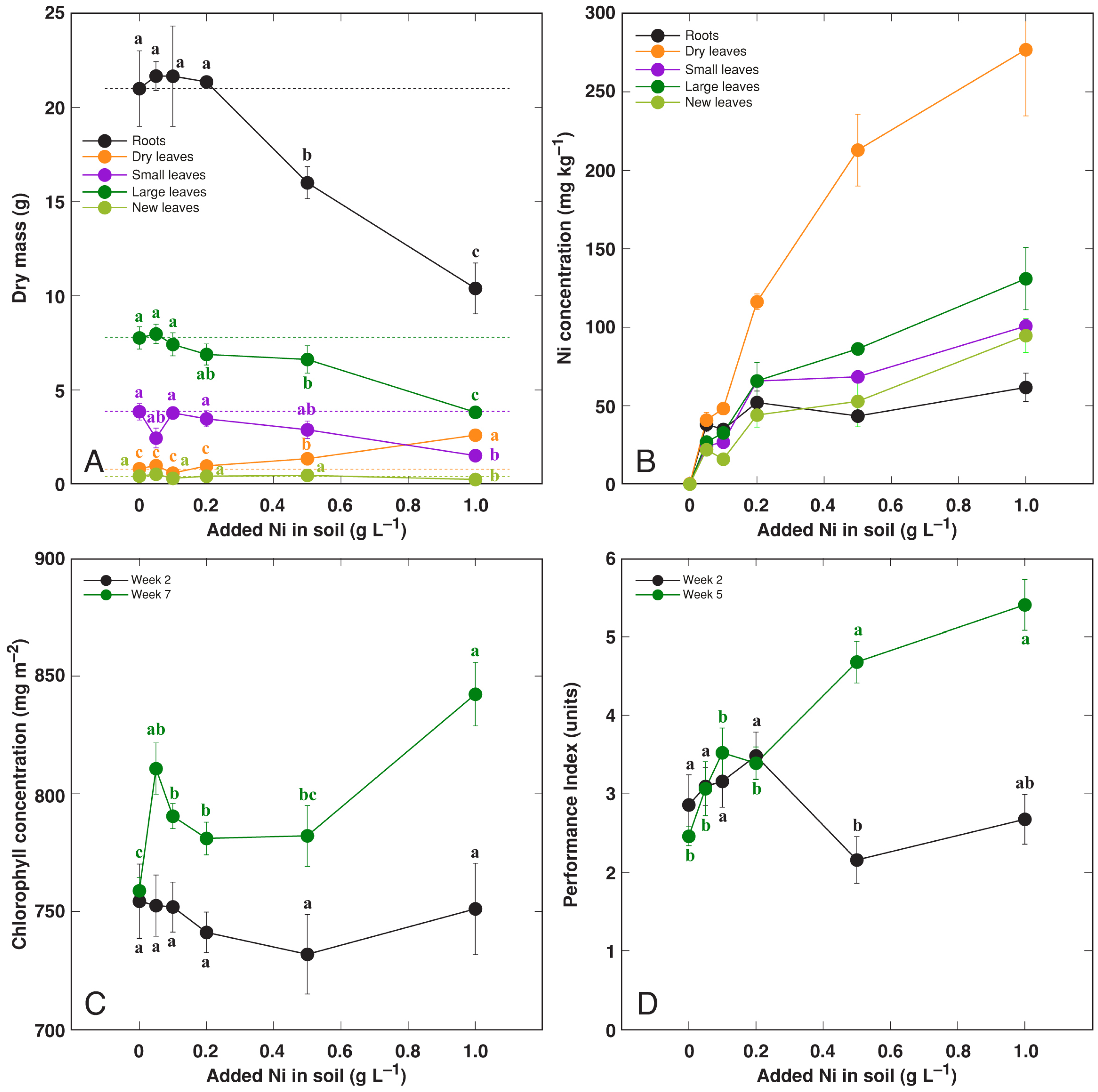
3.5. Effect of Heavy Metals: Ni
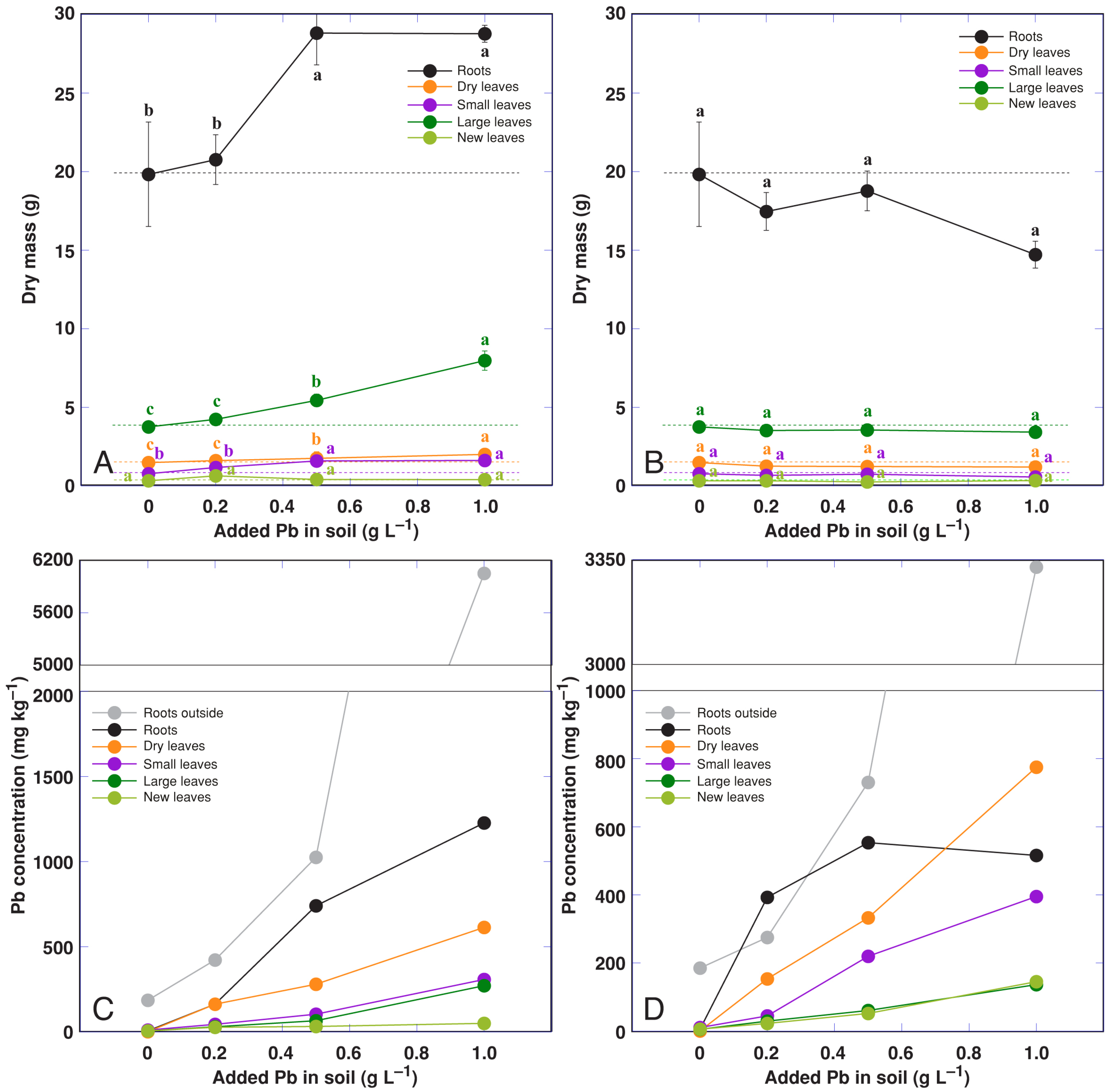
3.6. Effect of Heavy Metals: Pb in the Form of Nitrate and Acetate
4. Discussion
4.1. Abiotic Stress Tolerance
4.2. Tolerance against Heavy Metals
4.3. Metal Accumulation Potential
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marín-Muñiz, J.L.; Zitácuaro-Contreras, I.; Ortega-Pineda, G.; Álvarez-Hernández, L.M.; Martínez-Aguilar, K.E.; López-Roldán, A.; Zamora, S. Bibliometric analysis of constructed wetlands with ornamental flowering plants: The importance of green technology. Processes 2023, 11, 1253. [Google Scholar] [CrossRef]
- Mocek-Płóciniak, A.; Mencel, J.; Zakrzewski, W.; Roszkowski, S. Phytoremediation as an effective remedy for removing trace elements from ecosystems. Plants 2023, 12, 1653. [Google Scholar] [CrossRef] [PubMed]
- Priya, A.K.; Muruganandam, M.; Ali, S.S.; Kornaros, M. Clean-up of heavy metals from contaminated soil by phytoremediation: A multidisciplinary and eco-friendly approach. Toxics 2023, 11, 422. [Google Scholar] [CrossRef] [PubMed]
- Moray, C.; Goolsbay, E.W.; Bromham, L. The phylogenetic association between salt tolerance and heavy metal hyperaccumulation in Angiosperms. Evol. Biol. 2016, 43, 119–130. [Google Scholar] [CrossRef]
- Sruthi, P.; Shackira, A.M.; Puthur, J.T. Heavy metal detoxification mechanisms in halophytes: An overview. Wetlands Ecol. Manag. 2017, 25, 129–148. [Google Scholar] [CrossRef]
- Waly, M.M.; Ahmed, T.; Abunada, Z.; Miskovski, S.B.; Thomson, C. Constructed wetland for sustainable and low-cost wastewater treatment: Review article. Land 2022, 11, 1388. [Google Scholar] [CrossRef]
- Agaton, C.B.; Guila, P.M.C. Ecosystem services valuation of constructed wetlands as a nature-based solution to wastewater treatment. Earth 2023, 4, 78–92. [Google Scholar] [CrossRef]
- Sager, L.; Clerc, C. Factors influencing the distribution of Hydrocharis morsus-ranae L. and Rumex hydrolapathum Huds. in a mowed low-lying marshland, Réserve de Cheyres, lac de Neuchâtel, Switzerland. In Macrophytes in Aquatic Ecosystems: From Biology to Management; Caffrey, J.M., Dutartre, A., Haury, J., Murphy, K.J., Wade, P.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 223–229. [Google Scholar]
- Visser, E.J.W.; Blom, C.W.P.M.; Voesenek, L.A.C.J. Flooding-induced adventitious rooting in Rumex: Morphology and development in an ecological perspective. Acta Bot. Neerl. 1996, 45, 17–28. [Google Scholar] [CrossRef]
- Grevilliot, F.; Muller, S. Grassland ecotopes of the upper Meuse as references for habitats and biodiversity restoration: A synthesis. Landsc. Ecol. 2002, 17, 19–33. [Google Scholar] [CrossRef]
- Grevilliot, F.; Krebs, L.; Muller, S. Comparative importance and interference of hydrological conditions and soil nutrient gradients in floristic biodiversity in flood meadows. Biodiv. Conserv. 1998, 7, 1495–1520. [Google Scholar] [CrossRef]
- Voesenek, L.A.C.J.; Benschop, J.J.; Bou, J.; Cox, M.C.H.; Groeneveld, H.W.; Millenaar, F.F.; Vreeburg, R.A.M.; Peeters, A.J.M. Interactions between plant hormones regulate submergence-induced shoot elongation in the flooding-tolerant dicot Rumex palustris. Ann. Bot. 2003, 91, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Fang, Y. Copper accumulation by Polygonum microcephalum D. Don and Rumex hastatus D. Don from copper mine spoils in Yunnan Province, P.R. China. Environ. Geol. 2001, 40, 902–907. [Google Scholar] [CrossRef]
- Zhuang, P.; Wang, Q.W.; Wang, H.B.; Shu, W.S. Phytoextraction of heavy metals by eight plant species in field. Water Air Soil Pollut. 2007, 184, 235–242. [Google Scholar] [CrossRef]
- Xue, X.; Liu, G. Resistance and distribution to heavy metals of Zoysia sinica Hance and Rumex crispus. Adv. Mater. Res. 2014, 1010–1012, 117–120. [Google Scholar] [CrossRef]
- Vondráková, S.; Száková, J.; Drábek, O.; Tejnecký, V.; Hejcman, M.; Müllerová, V.; Tlustoð, P. Aluminium uptake and translocation in Al hyperaccumulator Rumex obtusifolius is affected by low-molecular weight organic acids content and soil pH. PLoS ONE 2015, 10, e0123351. [Google Scholar] [CrossRef]
- Zhao, Y.-H.; Jing, J.-W.; Wang, X.-T.; Yue, H.-M.; Niu, X.-Y.; Fang, J.-P. Study on heavy metals bioaccumulation characteristics and tolerance to pioneer plants from Central Tibet mining area. Acta Agrestia Sin. 2016, 24, 598–603. [Google Scholar]
- Wen, W.; Zhao, H.; Ma, J.; Li, Z.; Li, H.; Zhu, X.; Shao, J.; Yang, Z.; Yang, Y.; He, F.; et al. Effects of mutual intercropping on Pb and Zn accumulation of accumulator plants Rumex nepalensis, Lolium perenne and Trifolium repens. Chem. Ecol. 2018, 34, 259–271. [Google Scholar] [CrossRef]
- Barrutia, O.; Epelde, L.; García-Plazaola, J.I.; Garbisu, C.; Becerril, J.M. Phytoextraction potential of two Rumex acetosa L. accessions collected from metalliferous and non-metalliferous sites: Effect of fertilization. Chemosphere 2009, 74, 259–264. [Google Scholar] [CrossRef]
- Ye, M.; Liao, B.; Li, J.T.; Mengoni, A.; Hu, M.; Luo, W.C.; Shu, W.S. Contrasting patterns of genetic divergence in two sympatric pseudo-metallophytes: Rumex acetosa L. and Commelina communis L. BMC Evol. Biol. 2012, 12, 84. [Google Scholar] [CrossRef] [Green Version]
- Čiamporová, M.; Nadubinská, M.; Banásová, V.; Ďurišová, E.; Zelinová, V.; Horak, O.; Gruber, D.; Lichtscheidl, I.K. Structural traits of leaf epidermis correspond to metal tolerance in Rumex acetosella populations growing on metal-contaminated soils. Protoplasma 2021, 258, 1277–1290. [Google Scholar] [CrossRef]
- Jutila, H. How does grazing by cattle modify the vegetation of coastal grasslands along the Baltic Sea? Ann. Bot. Fenn. 2001, 38, 181–200. [Google Scholar]
- Tyler, T.; Herbertsson, L.; Olofsson, J.; Olsson, P.A. Ecological indicator and traits values for Swedish vascular plants. Ecol. Indic. 2021, 120, 106923. [Google Scholar] [CrossRef]
- Ievinsh, G.; Dišlere, E.; Karlsons, A.; Osvalde, A.; Vikmane, M. Physiological responses of wetland species Rumex hydrolapathum to increased concentration of biogenous heavy metals Zn and Mn in substrate. Proc. Latv. Acad. Sci. B 2020, 74, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Landorfa-Svalbe, Z.; Andersone-Ozola, U.; Ievinsh, G. Type of anion largely determines salinity tolerance in four Rumex species. Plants 2023, 12, 92. [Google Scholar] [CrossRef]
- Snowden, R.E.D.; Wheeler, B.D. Iron toxicity to fen plant species. J. Ecol. 1993, 81, 35–46. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterise and screen photosynthetic samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor & Francis: London, UK, 2000; pp. 445–483. [Google Scholar]
- Gailite, A.; Andersone-Ozola, U.; Samsone, I.; Karlsons, A.; Ievinsh, G. Ecophysiology of endangered plant species Saussurea esthonica: Effect of mineral nutrient availability and soil moisture. Plants 2023, 12, 888. [Google Scholar] [CrossRef] [PubMed]
- Schat, H. A comparative ecophysiological study on the effects of waterlogging and submergence on dune slack plants: Growth, survival and mineral nutrition in sand culture experiments. Oecologia 1984, 62, 279–286. [Google Scholar] [CrossRef]
- Fukao, T.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Submergence and waterlogging stress in plants: A review highlighting research opportunities and understudied aspects. Front. Plant Sci. 2019, 10, 340. [Google Scholar] [CrossRef]
- Samsone, I.; Ievinsh, G. Different plant species accumulate various concentration of Na+ in a sea-affected coastal wetland during a vegetation season. Environ. Exp. Biol. 2018, 16, 117–127. [Google Scholar]
- Shani, U.; Ben-Gal, A.; Tripler, E.; Dudley, L.M. Plant response to the soil environment: An analytical model integrating yield, water, soil type, and salinity. Water Resour. Res. 2007, 43, W08418. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Hniličková, H.; Hnilička, F.; Martinková, J.; Kraus, K. Effects of salt stress on water status, photosynthesis and chlorophyll fluorescence of rocket. Plant Soil Environ. 2017, 63, 362–367. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wei, G.; Jie, Y.; Wang, L.; Zhou, H.; Ran, C.; Anjum, S.A. Effects of concentrations of sodium chloride on photosynthesis, antioxidative enzymes, growth and fiber yield of hybrid ramie. Plant Physiol. Biochem. 2014, 76, 86–93. [Google Scholar] [CrossRef]
- Najar, R.; Aydi, S.; Sassi-Aydi, S.; Zarai, A.; Abdelly, C. Effect of salt stress on photosynthesis and chlorophyll fluorescence in Medicago truncatula. Plant Biosyst. 2019, 153, 88–97. [Google Scholar] [CrossRef]
- Qiu, N.; Lu, Q.; Lu, C. Photosynthesis, photosystem II efficiency and the xanthophyll cycle in the salt-adapted halophyte Atriplex centralasiatica. New Phytol. 2003, 159, 479–486. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Mateos-Naranjo, E.; Figueroa, M.E.; Davy, A.J. Salt stimulation of growth and photosynthesis in an extreme halophyte, Arthrocnemum macrostachyum. Plant Biol. 2010, 12, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Redondo-Gómez, S.; Wharmby, C.; Castillo, J.M.; Mateos-Naranjo, E.; Luque, C.J.; de Cires, A.; Luque, T.; Davy, A.J.; Figueroa, M.E. Growth and photosynthetic responses to salinity in an extreme halophyte, Sarcocornia fruticosa. Physiol. Plant. 2006, 128, 116–124. [Google Scholar] [CrossRef]
- Moravčiková, D.; Žiarovská, J. The effect of cadmium on plants in terms of the response of gene expression level and activity. Plants 2023, 12, 1848. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Mir, R.A.; Tyagi, A.; Manzar, N.; Kashyap, A.S.; Mushtaq, M.; Raina, A.; Park, S.; Sharma, S.; Mir, Z.A.; et al. Chromium toxicity in plants: Signaling, mitigation, and future perspectives. Plants 2023, 12, 1502. [Google Scholar] [CrossRef] [PubMed]
- van der Pas, L.; Ingle, R.A. Towards an understanding of the molecular basis of nickel hyperaccumulation in plants. Plants 2019, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Afzal, M.R.; Naz, M.; Wan, J.; Dai, Z.; Ullah, R.; ur Rehman, S.; Du, D. Insights into the mechanisms involved in lead (Pb) tolerance in invasive plants—The current status of understanding. Plants 2023, 12, 2084. [Google Scholar] [CrossRef] [PubMed]
- Ievinsh, G.; Landorfa-Svalbe, Z.; Andersone-Ozola, U.; Karlsons, A.; Osvalde, A. Salinity and heavy metal tolerance, and phytoextraction potential of Ranunculus sceleratus plants from a sandy coastal beach. Life 2022, 12, 1959. [Google Scholar] [CrossRef] [PubMed]
- Wyszkowski, M.; Radziemska, M. Assessment of tri- and hexavalent chromium phytotoxicity on oats (Avena sativa L.) biomass and content of nitrogen compounds. Water Air Soil Pollut. 2013, 224, 1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Mfarrej, M.F.B.; E-Esawi, M.A.; Waseem, M.; Alatawi, A.; Nafees, M.; Saleem, M.H.; Rizwan, M.; Yasmeen, T.; Anayat, A.; et al. Chromium-resistant Staphylococcus aureus alleviates chromium toxicity by developing synergistic relationships with zinc oxide nanoparticles in wheat. Ecotoxicol. Environ. Safety 2022, 230, 113142. [Google Scholar] [CrossRef] [PubMed]
- Bera, A.K.; Kanta-Bokaria, A.K.; Bokaria, K. Effect of tannery effluent on seed germination, seedling growth and chloroplast pigment content in mungbean (Vigna radiata). Environ. Ecol. 1999, 17, 958–961. [Google Scholar]
- Istenič, D.; Arias, C.A.; Vollertsen, J.; Nielsen, A.H.; Wium-Andersen, T.; Hvitved-Jacobsen, T.; Brix, H. Improved urban stormwater treatment and pollutant removal pathways in amended wet detention ponds. J. Environ. Sci. Health A 2012, 47, 1466–1477. [Google Scholar] [CrossRef]
- Gawęda, M. Heavy metal content in common sorrel plants (Rumex acetosa L.) obtained from natural sites in Małopolska province. Pol. J. Environ. Stud. 2009, 18, 213–218. [Google Scholar]
- Reeves, R.D.; Baker, A.J.M.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; van der Ent, A. A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Ullah, R.; Hadi, F.; Ahmad, S.; Jan, A.U.; Rongliang, Q. Phytoremediation of lead and chromium contaminated soil improves with the endogenous phenolics and proline production in Parthenium, Cannabis, Euphorbia, and Rumex species. Water Air Soil Pollut. 2019, 230, 40. [Google Scholar] [CrossRef]
- Bah, A.M.; Dai, H.; Zhao, J.; Sun, H.; Cao, F.; Zhang, G.; Wu, F. Effects of cadmium, chromium and lead on growth, metal uptake and antioxidative capacity in Typha angustifolia. Biol. Trace Elem. Res. 2011, 142, 77–92. [Google Scholar] [CrossRef]
- Massoura, S.T.; Echevarria, G.; Leclerc-Cessac, E.; Morel, J.L. Response of excluder, indicator, and hyperaccumulator plants to nickel availability in soils. Austr. J. Soil Res. 2004, 42, 933–938. [Google Scholar] [CrossRef]
- Ievinsh, G.; Andersone-Ozola, U.; Samsone, I. Alyssum montanum subsp. gmelinii, a rare plant species from coastal sand dunes, as a potential Ni accumulator: Comparison with Alyssum murale. Environ. Exp. Biol. 2020, 18, 107–115. [Google Scholar]
- Zulfiqar, U.; Farooq, M.; Hussain, S.; Maqsood, M.; Hussain, M.; Ishfaq, M.; Ahmad, M.; Anjum, M.Z. Lead toxicity in plants: Impacts and remediation. J. Environ. Manag. 2019, 250, 109557. [Google Scholar] [CrossRef] [PubMed]
- Ievinsh, G.; Ieviņa, S.; Andersone-Ozola, U.; Samsone, I. Leaf sodium, potassium and electrolyte accumulation capacity of plant species from salt-affected coastal habitats of the Baltic Sea: Towards a definition of Na hyperaccumulation. Flora 2021, 274, 151748. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M. Dual role of metallic trace elements in stress biology—From negative to beneficial impacts on plants. Int. J. Mol. Sci. 2019, 20, 20–24. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Soil Moisture | Treatment | Fertilization |
|---|---|---|---|
| Soil Moisture | 30% | – | Before treatment: Kristalon Green Label (0.5 g L–1), 50 mL per plant |
| 30% | 3 × 100 mL 200 mM NaCl | ||
| 50% | – | ||
| 50% | 3 × 100 mL 200 mM NaCl | ||
| Waterlogged | – | ||
| Waterlogged | 3 × 100 mL 200 mM NaCl | ||
| Flooded | – | ||
| Flooded | 3 × 100 mL 200 mM NaCl | ||
| Waterlogging with NaCl | Waterlogged | Stepwise increase to 0, 25, 50, 100, 200, 400 mM NaCl in water used for waterlogging | Before treatment: Kristalon Green Label (0.5 g L–1), 50 mL per plant |
| Increasing Soil Cd | 60–70% | Stepwise increase to 0, 2, 5, 10, 20, 100 mg Cd L–1 of soil in a form of solubilized CdSO4 after the final transplanting of seedlings | Kristalon Green Label (0.5 g L–1), 50 mL per plant, once every two weeks |
| Increasing Soil Cr | 60–70% | Stepwise increase to 0, 2, 5, 10, 20, 100 mg Cr L–1 of soil in the form of solubilized K2Cr2O7 after the final transplanting of seedlings | Kristalon Green Label (0.5 g L–1), 50 mL per plant, once every two weeks |
| Increasing Soil Ni | 60–70% | Ni 0, 50, 100, 200, 500, 1000 mg L–1 of soil in the dry form of NiSO4 before the final transplanting of seedlings | Kristalon Green Label (0.5 g L–1), 50 mL per plant, once every two weeks |
| Increasing Soil Pb Nitrate and Pb Acetate | 60–70% | Stepwise increase to 0, 200, 500, 1000 mg Pb L–1 of soil in the form of solubilized Pb(NO3)2 or Pb(CH3COO)2 after the final transplanting of seedlings | Kristalon Green Label (0.5 g L–1), weeks 50 mL per plant, once every two |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ieviņa, S.; Karlsons, A.; Osvalde, A.; Andersone-Ozola, U.; Ievinsh, G. Coastal Wetland Species Rumex hydrolapathum: Tolerance against Flooding, Salinity, and Heavy Metals for Its Potential Use in Phytoremediation and Environmental Restoration Technologies. Life 2023, 13, 1604. https://doi.org/10.3390/life13071604
Ieviņa S, Karlsons A, Osvalde A, Andersone-Ozola U, Ievinsh G. Coastal Wetland Species Rumex hydrolapathum: Tolerance against Flooding, Salinity, and Heavy Metals for Its Potential Use in Phytoremediation and Environmental Restoration Technologies. Life. 2023; 13(7):1604. https://doi.org/10.3390/life13071604
Chicago/Turabian StyleIeviņa, Silvija, Andis Karlsons, Anita Osvalde, Una Andersone-Ozola, and Gederts Ievinsh. 2023. "Coastal Wetland Species Rumex hydrolapathum: Tolerance against Flooding, Salinity, and Heavy Metals for Its Potential Use in Phytoremediation and Environmental Restoration Technologies" Life 13, no. 7: 1604. https://doi.org/10.3390/life13071604