
Seed Treatment with Sodium Nitroprusside Ensures a Long-Term Physiological and Protective Effect on Wheat under Salinity
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Treatment
2.1.1. Experiments on the Conditions of a Laboratory
2.1.2. Pot Experiments under Controlled Conditions
2.1.3. Field Experiments
2.2. Estimation of Leaf Surface Area
2.3. Yield Components
2.4. Grain Quality Analysis (Amino Acids, Carbohydrates, and Raw Fat)
2.5. Pigment Analysis
2.6. Oxidative Stress Markers
2.7. Estimation of Nonenzymatic Antioxidants
2.8. Antioxidant Enzyme Activities
2.9. Endogenous NO Measurement
3. Statistical Analysis
4. Results
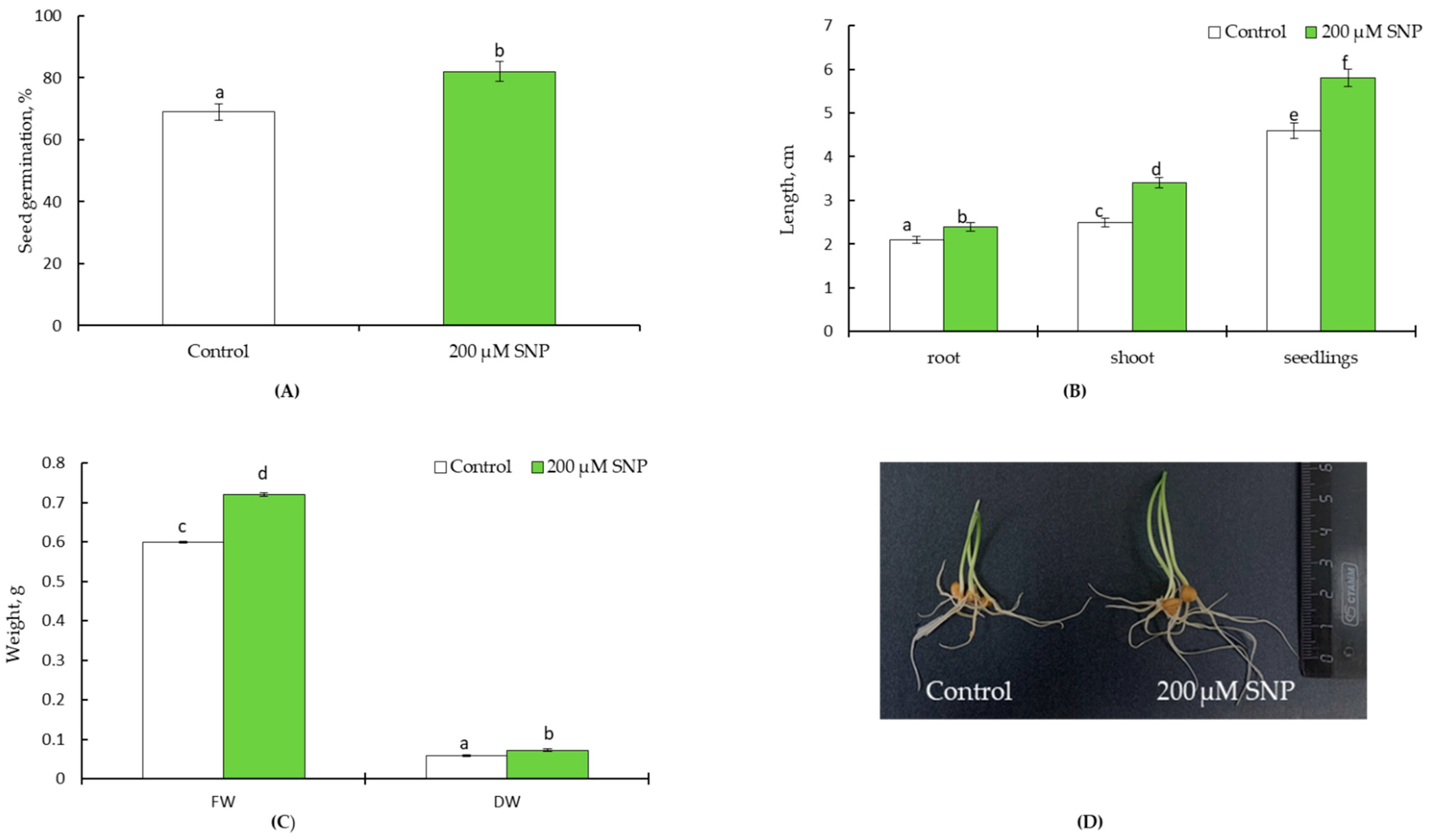
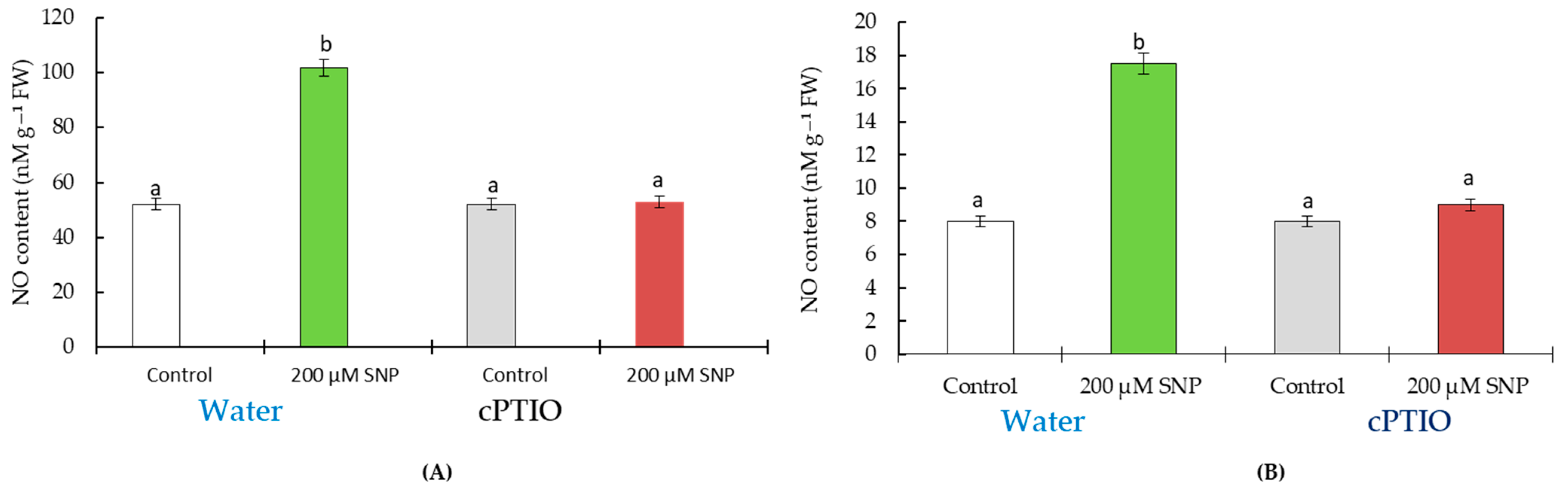
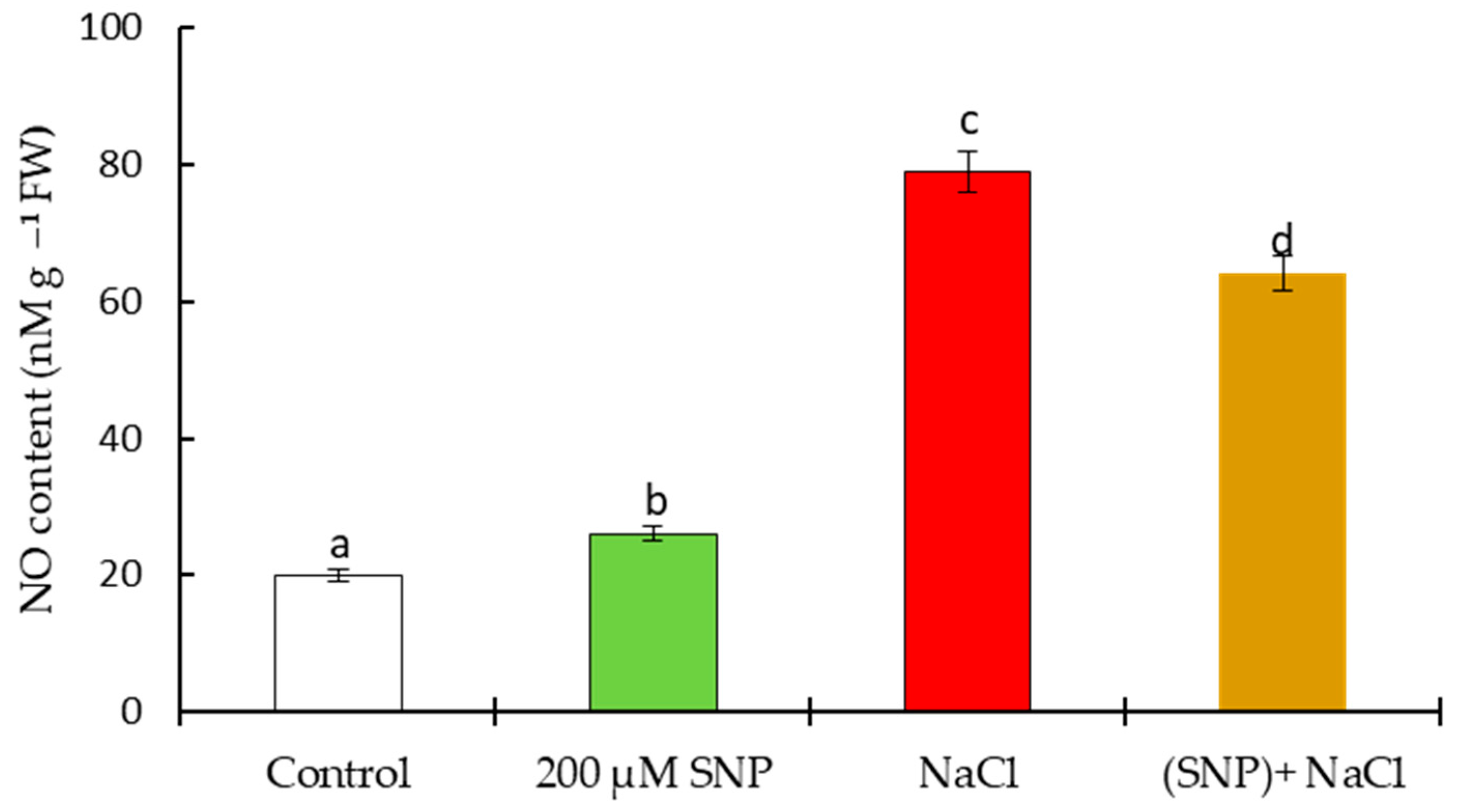
4.1. Laboratory Experiment
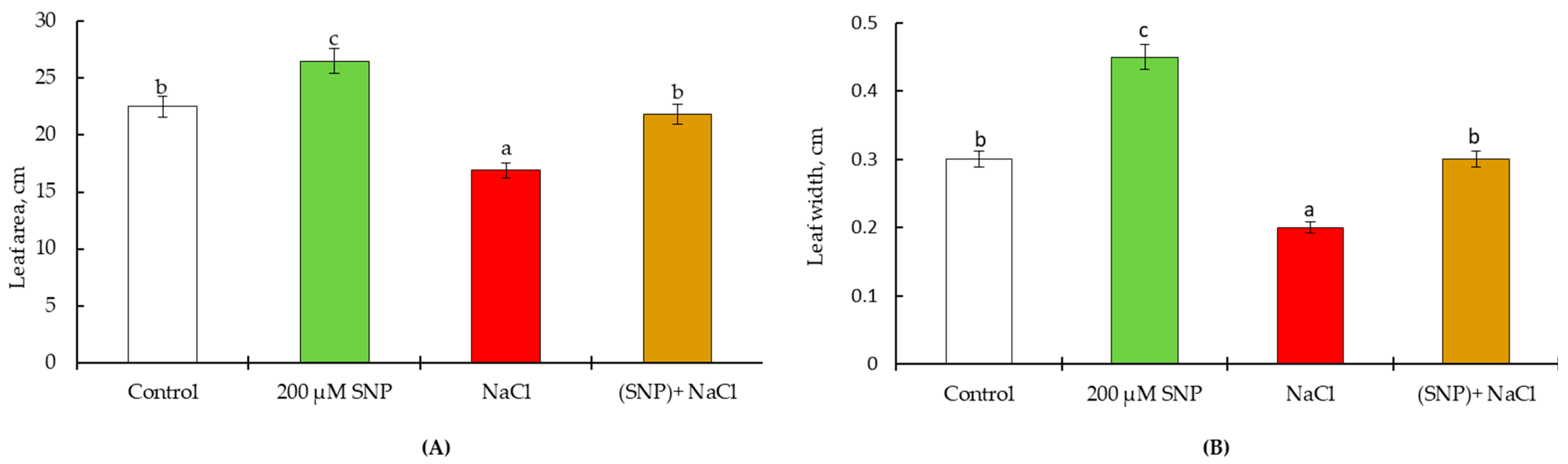
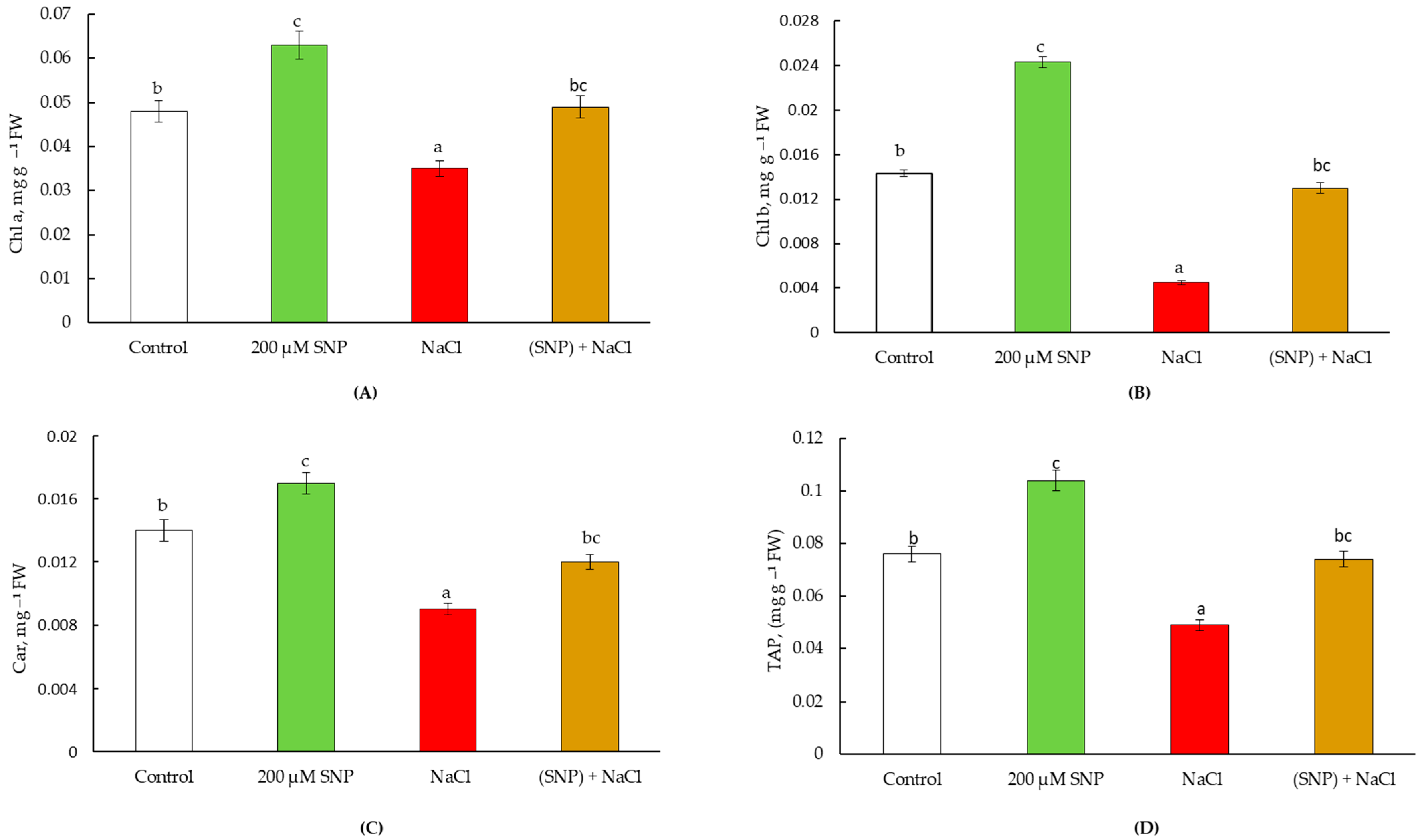
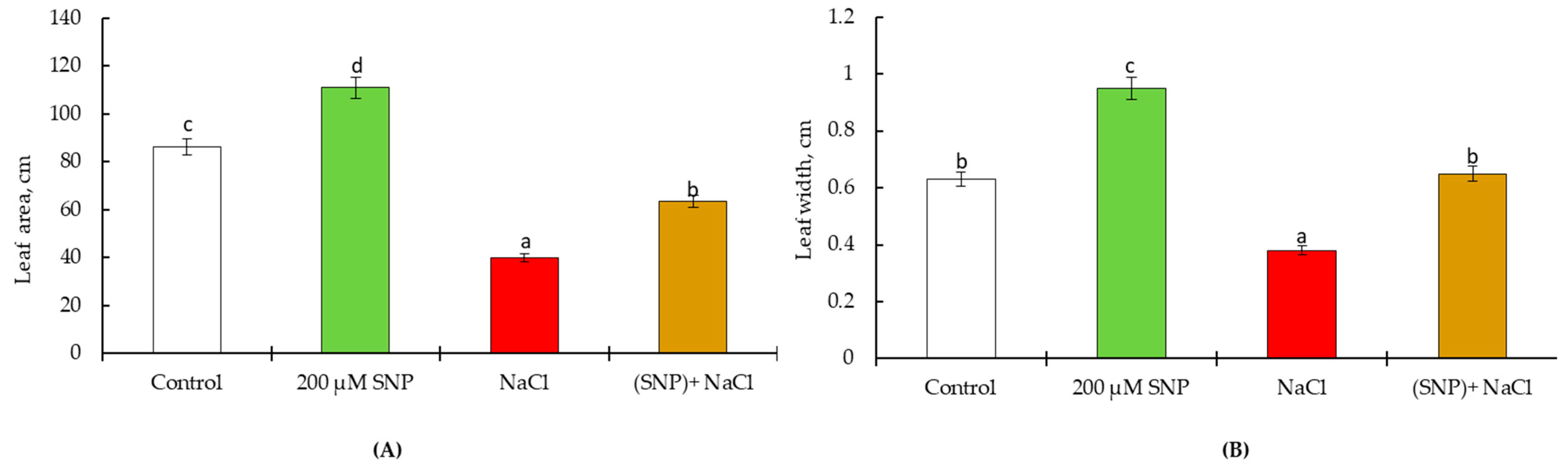
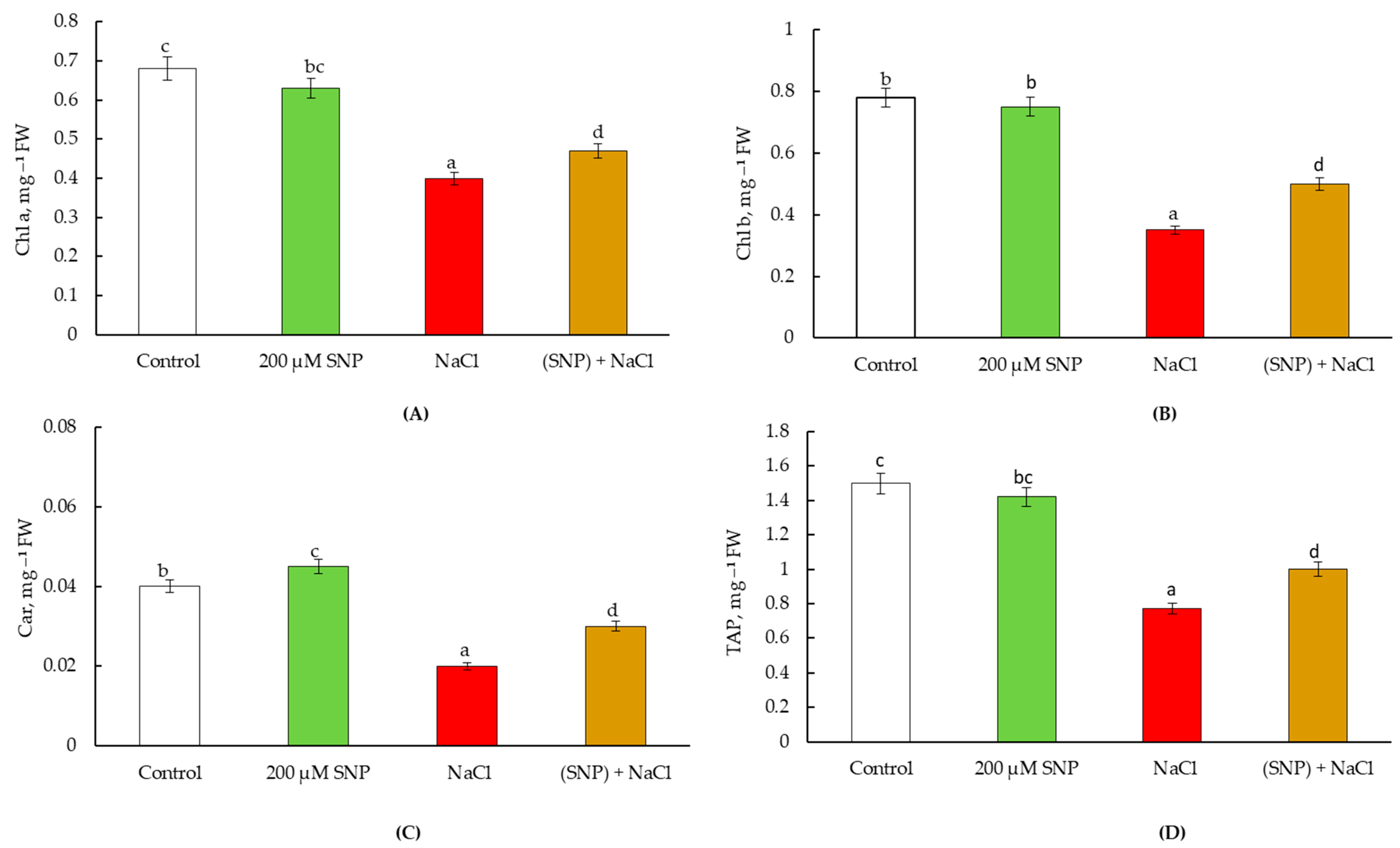
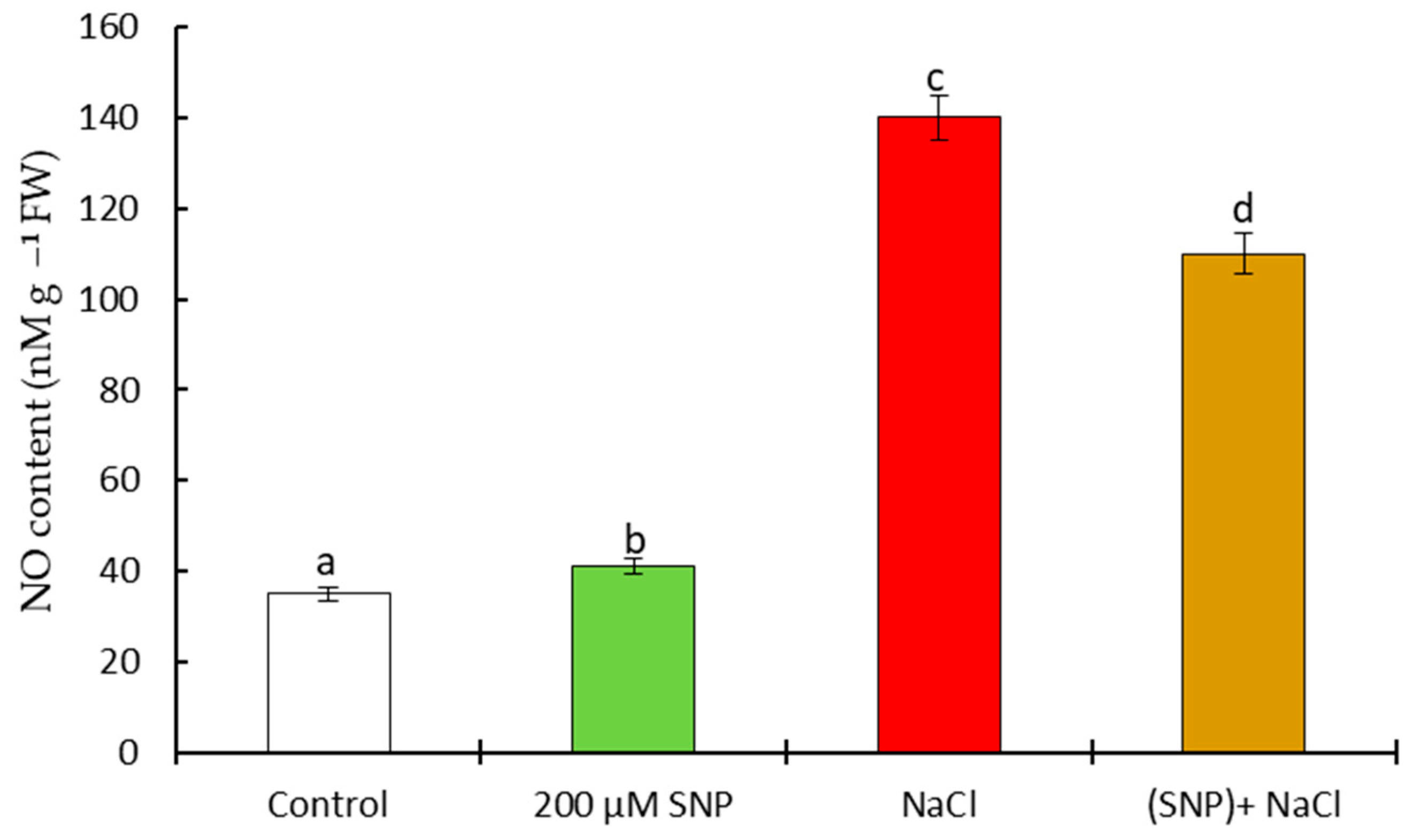
4.2. Pot Experiments
4.3. Small-Scale Field Experiments
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hossain, M.S. Present scenario of global salt affected soils, its management and importance of salinity research. Int. Res. J. Biol. Sci. 2019, 1, 1–3. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant. Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragaey, M.M.; Sadak, M.S.; Dawood, M.F.A.; Mousa, N.H.S.; Hanafy, R.S.; Latef, A.A.H.A. Role of signaling molecules sodium nitroprusside and arginine in alleviating salt-induced oxidative stress in wheat. Plants 2022, 11, 1786. [Google Scholar] [CrossRef] [PubMed]
- Zahra, N.; Sulaiman, M.; Hinai, A.; Hafeez, M.B.; Rehman, A.; Wahid, A.; Siddique, K.H.M.; Farooq, M. Regulation of photosynthesis under salt stress and associated tolerance mechanisms. Plant Physiol. Biochem. 2022, 178, 55–69. [Google Scholar] [CrossRef]
- Ali, Q.; Daud, M.K.; Haider, M.Z.; Ali, S.; Rizwan, M.; Aslam, N.; Noman, A.; Iqbal, N.; Shahzad, F.; Deeba, F.; et al. Seed priming by sodium nitroprusside improves salt tolerance in wheat (Triticum aestivum L. ) by enhancing physiological and biochemical parameters. Plant Physiol. Biochem. 2017, 119, 50–58. [Google Scholar] [CrossRef]
- Habib, N.; Ali, Q.; Ali, S.; Haider, M.Z.; Javed, M.T.; Khalid, M.; Perveen, R.; Alsahli, A.A.; Alyemeni, M.N. Seed Priming with Sodium Nitroprusside and H2O2 Confers Better Yield in Wheat Under Salinity: Water Relations, Antioxidative Defense Mechanism and Ion Homeostasis. J. Plant Growth Regul. 2021, 40, 2433–2453. [Google Scholar] [CrossRef]
- Sami, F.; Siddiqui, H.; Alam, P.; Hayat, S. Nitric oxide mitigates the salt-induced oxidative damage in mustard by upregulating the activity of various enzymes. J. Plant Growth Regul. 2021, 40, 2409–2432. [Google Scholar] [CrossRef]
- Quamruzzaman, M.; Manik, S.M.N.; Shabala, S.; Zhou, M. Improving performance of salt-grown crops by exogenous application of plant growth regulators. Biomolecules 2021, 11, 788. [Google Scholar] [CrossRef]
- Habib, N.; Ali, Q.; Ali, S.; Javed, M.T.; Zulqurnain Haider, M.; Perveen, R.; Shahid, M.R.; Rizwan, M.; Abdel-Daim, M.M.; Elkelish, A.; et al. Use of nitric oxide and hydrogen peroxide for better yield of wheat (Triticum aestivum L.) under water deficit conditions: Growth, osmoregulation, and antioxidative defense mechanism. Plants 2020, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Fatima, A.; Husain, T.; Suhel, M.; Prasad, S.M.; Singh, V.P. Implication of nitric oxide under salinity stress: The possible interaction with other signaling molecules. J Plant Growth Regul. 2022, 41, 163–177. [Google Scholar] [CrossRef]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Yun, B.-W. Nitric oxide acts as a key signaling molecule in plant development under stressful conditions. Int. J. Mol. Sci. 2023, 24, 4782. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, Y.; Liu, L.; Liu, X.; Li, B.; Jin, C.; Lin, X. Molecular functions of nitric oxide and its potential applications in horticultural crops. Hortic Res. 2021, 8, 71. [Google Scholar] [CrossRef]
- Shang, J.-X.; Li, X.; Li, C.; Zhao, L. The role of nitric oxide in plant responses to salt stress. Int. J. Mol. Sci. 2022, 23, 6167. [Google Scholar] [CrossRef]
- Yasir, T.A.; Khan, A.; Skalicky, M.; Wasaya, A.; Rehmani, M.I.A.; Sarwar, N.; Mubeen, K.; Aziz, M.; Hassan, M.M.; Hassan, F.A.S. Exogenous sodium nitroprusside mitigates salt stress in lentil (Lens culinaris Medik.) by affecting the growth, yield, and biochemical properties. Molecules 2021, 26, 2576. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Liu, G.; Xu, S.; Dai, J.; Li, W.; Li, Z.; Zhang, D.; Li, C.; Dong, H. Nitric oxide increases the biomass and lint yield of field-grown cotton under temporary waterlogging through physiological and molecular regulation. Field Crop. Res. 2021, 261, 107989. [Google Scholar] [CrossRef]
- Badem, A.; Söylemez, S. Effects of nitric oxide and silicon application on growth and productivity of pepper under salinity stress. J. King Saud Univ. 2022, 34, 102189. [Google Scholar] [CrossRef]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef] [Green Version]
- Hmidi, D.; Abdelly, C.; Athar, H.U.; Ashraf, M.; Messedi, D. Effect of salinity on osmotic adjustment, proline accumulation and possible role of ornithine-δ-aminotransferase in proline biosynthesis in Cakile maritima. Physiol. Mol. Biol. Plants 2018, 24, 1017–1033. [Google Scholar] [CrossRef]
- Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savouré, A. How does proline treatment promote salt stress tolerance during crop plant development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef]
- Maslennikova, D.R.; Allagulova, C.h.R.; Fedorova, K.A.; Plotnikov, A.A.; Avalbaev, A.M.; Shakirova, F.M. Cytokinins contribute to realization of nitric oxide growth-stimulating and protective effects on wheat plants. Rus. J. Plant Physiol. 2017, 64, 665–671. [Google Scholar] [CrossRef]
- Maslennikova, D.R.; Plotnikov, A.A.; Shakirova, F.M. Comparative analysis of the physiological effect of nitric oxide and 6-benzylaminopurine on the state of the components of the glutathione complex in the roots of wheat seedlings. Agrochem 2019, 3, 37–43. [Google Scholar] [CrossRef]
- Maslennikova, D.R.; Lastochkina, O.V.; Shakirova, F.M. Exogenous sodium nitroprusside improves salt stress tolerance of wheat (Triticum aestivum L.) via regulating the components of ascorbate-glutathione cycle, chlorophyll content and stabilization of cell membranes state. Russ. J. Plant Physiol. 2022, 69, 130. [Google Scholar] [CrossRef]
- Maslennikova, D.; Ivanov, S.; Petrova, S.; Burkhanova, G.; Maksimov, I.; Lastochkina, O. Components of the phenylpropanoid pathway in the implementation of the protective effect of sodium nitroprusside on wheat under salinity. Plants 2023, 12, 2123. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Nile, S.H.; Thiruvengadam, M.; Wang, Y.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Nile, A.; Sun, M.; Venkidasamy, B.; Xiao, J.; et al. Nano-priming as emerging seed priming technology for sustainable agriculture—Recent developments and future perspectives. J. Nanobiotechnol. 2022, 20, 254. [Google Scholar] [CrossRef]
- Tahjib-Ul-Arif, M.; Wei, X.; Jahan, I.; Hasanuzzaman, M.; Sabuj, Z.H.; Zulfiqar, F.; Chen, J.; Iqbal, R.; Dastogeer, K.M.G.; Sohag, A.A.M.; et al. Exogenous nitric oxide promotes salinity tolerance in plants: A meta-analysis. Front. Plant Sci. 2022, 13, 957735. [Google Scholar] [CrossRef]
- Choudhary, V.K.; Chander, S.; Chethan, C.R.; Kumar, B. Effect of seed priming on abiotic stress tolerance in plants. In Plant Tolerance to Environmental Stress. Role of Phytoprotectants; Hasanuzzaman, M., Fujita, M., Oku, H., Islam, M.T., Eds.; CRC Press: Boca Raton, FL, USA, 2019; p. 488. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, R.; Wang, X.; Zhao, C.; Shen, H.; Yang, L. Nitric by integrating multiple signalling pathways. Int. J. Mol. Sci. 2023, 24, 9052. [Google Scholar] [CrossRef]
- Wang, H.; Tang, S.; Wang, J.; Shen, H.; Yang, L. Interaction between reactive oxygen species and hormones during the breaking of embryo dormancy in Sorbus pohuashanensis by exogenous nitric oxide. J. For. Res. 2022, 33, 435–444. [Google Scholar] [CrossRef]
- Zykin, V.A.; Kiraev, R.S. Ecologically sustainable varieties of spring soft wheat Salvata Yulaev and Vatan. Bull. Altai State Agrar. Univ. 2011, 8, 5–8. [Google Scholar]
- Mokronosova, A.T. Small Workshop on Plant Physiology; Moscow State University: Moscow, Russia, 1994; 184p. [Google Scholar]
- Arkhipova, T.; Martynenko, E.; Sharipova, G.; Kuzmina, L.; Ivanov, I.; Garipova, M.; Kudoyarova, G. Effects of plant growth promoting rhizobacteria on the content of abscisic acid and salt resistance of wheat plants. Plants 2020, 9, 1429. [Google Scholar] [CrossRef]
- Aldesuquy, H.; Baka, Z.; Mickket, B. Kinetin and spermine mediated induction of salt tolerance in wheat plants: Leaf area, photosynthesis and chloroplast ultrastructure of flag leaf at ear emergence. Egypt. J. Basic Appl. Sci. 2014, 1, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Schrader, J.; Shi, P.; Royer, D.L.; Peppe, D.J.; Gallagher, R.V.; Li, Y.; Wang, R.; Wright, I.J. Leaf size estimation based on leaf length, width and shape. Ann. Bot. 2021, 128, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Dospechov, B.A. Field Experiment Methodology; Agropromizdat: Moscow, Russia, 1985; 351p. [Google Scholar]
- Jefferey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1, and c2 in higher plants, algae, and natural phytoplankton. Biochem. Physiol. Pfl. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Maslennikova, D.; Lastochkina, O. Contribution of ascorbate and glutathione in endobacteria Bacillus subtilis-mediated drought tolerance in two Triticum aestivum L. genotypes contrasting in drought sensitivity. Plants 2021, 10, 2557. [Google Scholar] [CrossRef]
- Hissin, P.J.; Hilf, R.A. A fluorometric method for determination of oxidize and reduced glutathione in tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldern, R.P.; Teare, D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Kalinkina, L.G. Proline accumulation in cells of marine and freshwater chlorella in depending on the concentration of NaCl in the medium and the growth rate of algae. Plant Physiol. 1985, 32, 42–52. [Google Scholar]
- Bradford, M.M. A rapid and sensitive methods for quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Vishwakarma, A.; Wany, A.; Pandey, S.; Bulle, M.; Kumari, A.; Kishorekumar, R.; Igamberdiev, A.U.; Mur, L.A.J.; Gupta, K.J. Current approaches to measure nitric oxide in plants. J. Exp. Bot. 2019, 70, 4333–4343. [Google Scholar] [CrossRef] [Green Version]
- Lubyanova, A.R.; Bezrukova, M.V.; Shakirova, F.M. Involvement of nitric oxide in methyl jasmonate-mediated regulation of water metabolism in wheat plants under drought stress. Stresses 2022, 2, 477–492. [Google Scholar] [CrossRef]
- Sherin, G.; Aswathi, K.P.R.; Puthur, J.T. Photosynthetic functions in plants subjected to stresses are positively influenced by priming. Plant Stress 2022, 4, 100079. [Google Scholar] [CrossRef]
- Azmat, A.; Yasmin, H.; Hassan, M.N.; Nosheen, A.; Naz, R.; Sajjad, M.; Ilyas, N.; Akhtar, M.N. Co-application of bio-fertilizer and salicylic acid improves growth, photosynthetic pigments and stress tolerance in wheat under drought stress. PeerJ 2020, 8, e9960. [Google Scholar] [CrossRef]
- Nawaz, F.; Rafeeq, R.; Majeed, S. Biochar amendment in combination with endophytic bacteria stimulates photosynthetic activity and antioxidant enzymes to improve soybean yield under drought stress. J. Soil Sci. Plant Nutr. 2023, 23, 746–760. [Google Scholar] [CrossRef]
- Considine, M.J.; Foyer, C.H. Redox regulation of plant development. Antioxid. Redox Signal. 2014, 21, 1305–1326. [Google Scholar] [CrossRef] [Green Version]
- Nasir, N.M.; Khandaker, N.N.; Mohd, M.M.; Badaluddin, N.A.; Mat, N. Effect of hydrogen peroxide on plant growth, photosynthesis, leaf histology and rubisco gene expression of the Ficus deltoidei Jack Var. deltoidei Jack. J Plant Growth Regul. 2021, 40, 1950–1971. [Google Scholar] [CrossRef]
- Wieser, H.; Koehler, P.; Scherf, K.A. The two faces of wheat. Front. Nutr. 2020, 7, 517313. [Google Scholar] [CrossRef]
- Sułek, A.; Cacak-Pietrzak, G.; Rózewicz, M.; Nieróbca, A.; Grabiński, J.; Studnicki, M.; Sujka, K.; Dziki, D. Effect of production technology intensity on the grain yield, protein content and amino acid profile in common and durum wheat grain. Plants 2023, 12, 364. [Google Scholar] [CrossRef]
- Hu, X.; Ma, J.; Qian, W.; Cao, Y.; Zhang, Y.; Liu, B.; Tang, L.; Cao, W.; Zhu, Y.; Liu, L. Effects of low temperature on the amino acid composition of wheat grains. Agronomy 2022, 12, 1171. [Google Scholar] [CrossRef]
- Siddiqi, R.A.; Singh, T.P.; Rani, M.; Sogi, D.S.; Bhat, M.A. Diversity in grain, flour, amino acid composition, protein profiling, and proportion of total flour proteins of different wheat cultivars of North India. Front. Nutr. 2020, 7, 141. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Satheesh, N.; Kherawat, B.S.; Kumar, A.; Kim, H.-U.; Chung, S.-M.; Kumar, M. Regulation of reactive oxygen species during salt stress in plants and their crosstalk with other signaling molecules—Current perspectives and future directions. Plants 2023, 12, 864. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Glutathione in plants: Biosynthesis and physiological role in environmental stress tolerance. Physiol. Mol. Biol. Plants 2017, 23, 249–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliniaeifard, S.; Hajilou, J.; Tabatabaei, S.J.; Sifi-Kalhor, M. Effects of ascorbic acid and reduced glutathione on the alleviation of salinity stress in olive plants. Int. J. Fruit Sci. 2016, 16, 395–409. [Google Scholar] [CrossRef]
- Dorion, S.; Ouellet, J.C.; Rivoal, J. Glutathione metabolism in plants under stress: Beyond reactive oxygen species detoxification. Metabolites 2021, 11, 641. [Google Scholar] [CrossRef]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012. [CrossRef] [Green Version]
- González-Gordo, S.; Rodríguez-Ruiz, M.; López-Jaramillo, J.; Muñoz-Vargas, M.A.; Palma, J.M.; Corpas, F.J. Nitric oxide (NO) differentially modulates the ascorbate peroxidase (APX) isozymes of sweet pepper (Capsicum annuum L.) fruits. Antioxidants 2022, 11, 765. [Google Scholar] [CrossRef]
- Lastochkina, O.; Garshina, D.; Ivanov, S.; Yuldashev, R.; Khafizova, R.; Allagulova, C.; Fedorova, K.; Avalbaev, A.; Maslennikova, D.; Bosacchi, M. Seed priming with endophytic Bacillus subtilis modulates physiological responses of two different Triticum aestivum L. cultivars under drought stress. Plants 2020, 9, 1810. [Google Scholar] [CrossRef]
- López-Carrión, A.I.; Castellano, R.; Rosales, M.A.; Ruiz, J.M.; Romero, L. Role of nitric oxide under saline stress: Implications on proline metabolism. Biol. Plant 2008, 52, 587–591. [Google Scholar] [CrossRef]
- Sehar, Z.; Mir, I.R.; Khan, S.; Masood, A.; Khan, N.A. Nitric oxide and proline modulate redox homeostasis and photosynthetic metabolism in wheat plants under high temperature stress acclimation. Plants 2023, 12, 1256. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | GSH | GSSG | GSH/ GSSG | ASA | GR | APX | H2O2 | Proline | MDA |
|---|---|---|---|---|---|---|---|---|---|
| Control | 59 ± 2.5 a | 2.5 ± 0.11 a | 23.6 ± 0.9 a | 1.9 ± 0.08 a | 3.59 ± 0.14 a | 5.45 ± 0.22 a | 3.5 ± 0.13 a | 13 ± 0.52 a | 5.5 ± 0.22 a |
| 200 µM SNP | 65 ± 2.5 b | 2.3 ± 0.09 a | 28 ± 1.1 b | 2 ± 0.08 a | 4.9 ± 0.19 b | 5.78 ± 0.23 b | 3.9 ± 0.14 b | 18 ± 0.75 b | 5.56 ± 0.22 a |
| NaCl | 25 ± 1.1 c | 5.8 ± 0.23 b | 4.1 ± 0.16 c | 0.6 ± 0.02 b | 7.8 ± 0.32 c | 11.08 ± 0.44 | 6.1 ± 0.24 c | 30 ± 1.2 c | 7.8 ± 0.32 b |
| (SNP) + NaCl | 54 ± 2.16 ab | 2.8 ± 0.11 ac | 19.2 ± 0.8 d | 1.4 ± 0.06 c | 9.2 ± 0.36 d | 7.58 ± 0.3 d | 4.8 ± 0.19 d | 34 ± 1.36 d | 7.1 ± 0.29 c |
| Treatment | GSH | GSSG | GSH/ GSSG | ASA | GR | APX | H2O2 | Proline | MDA |
|---|---|---|---|---|---|---|---|---|---|
| Control | 90 ± 3.6 b | 8.9 ± 0.36 a | 10.1 ± 0.4 b | 2.85 ± 0.11 a | 5.35 ± 0.21 a | 8.17 ± 0.32 a | 5.25 ± 0.21 a | 20.4 ± 0.82 a | 8.25 ± 0.33 a |
| 200µM SNP | 93.9 ± 3.6 b | 9.1 ± 0.37 a | 10.3 ± 1.41 b | 3 ± 0.12 a | 6.2 ± 0.25 b | 8.6 ± 0.34 b | 5.89 ± 0.23 b | 21.3 ± 0.85 ab | 8.34 ± 0.33 a |
| NaCl | 30 ± 1.2 a | 12.5 ± 0.5 b | 2.4 ± 0.09 a | 1 ± 0.04 b | 9.3 ± 0.37 c | 15 ± 0.61 c | 9.1 ± 0.37 c | 35.8 ± 1.4 c | 16.2 ± 0.64 b |
| (SNP) + NaCl | 54 ± 2.2 c | 9 ± 0.36 a | 6 ± 0.24 c | 2.1 ± 0.08 c | 9.8 ± 0.39 d | 9.8 ± 0.39 d | 7.2 ± 0.29 d | 37.8 ± 1.5 d | 9.8 ± 0.39 c |
| Year | Treatments | Shoot Length (cm) | Spike Length (cm) | 1000 Grain Weight (g) |
|---|---|---|---|---|
| 2021 | Control | 101 ± 4.1 ᵃ | 9.3 ± 0.37 ᵃ | 36.8 ± 1.48 ᵃ |
| 200 µM SNP | 114 ± 4.6 ᵇ | 12 ± 0.48 ᵇ | 44.2 ± 1.76 ᵇ | |
| 2022 | Control | 109 ± 4.4 ᵃ | 10.4 ± 0.4 ᵃ | 39.3 ± 1.57 ᵃ |
| 200 µM SNP | 123 ± 4.9 ᵇ | 13 ± 0.52 ᵇ | 47.1 ± 1.88 ᵇ |
| Parameters | Control | 200 µM SNP |
|---|---|---|
| Valine | 408.07 ± 16.32 a | 511.75 ± 20.50 b |
| Leucine + Isoleucine | 995.42 ± 39.80 a | 1140.39 ± 45.60 b |
| Methionine | 151.42 ± 6.04 a | 233.48 ± 9.36 b |
| Tryptophan | 180.81 ± 7.20 a | 255.61 ± 10.2 b |
| Phenylalanine | 426.43 ± 17.04 a | 556.31 ± 22.24 b |
| Lysine | 327.39 ± 13.08 a | 455.09 ± 18.2 b |
| Threonine | 315.10 ± 12.6 | 421.09 ± 16.84 b |
| Histidine | 289.92 ± 11.55 a | 391.89 ± 15.67 b |
| Essential AA | 3091 ± 123.65 a | 3962 ± 158.42 b |
| Arginine | 610.67 ± 24.4 a | 800.21 ± 32.90 b |
| Serine | 557.33 ± 22.28 a | 700.28 ± 28.65 b |
| Proline | 1054.35 ± 42.15 a | 1270.28 ± 50.8 b |
| Glycine | 419.93 ± 16.74 a | 500.76 ± 20.86 b |
| Alanine | 357.47 ± 14.67 a | 420.49 ± 16.8 b |
| Aspartic acid + Asparagine | 401.72 ± 16.01 a | 422.53 ± 16.91 a |
| Glutamic acid | 3101.82 ± 124.04 a | 3275.39 ± 131.22 a |
| Cysteine | 262.26 ± 10.60 a | 261.45 ± 10.17 a |
| Non-essential AA | 6761 ± 270.45 a | 7693 ± 307.75 b |
| Total AA | 9852 ± 392.26 a | 11655 ± 466.65 b |
| Essential AA/Non-essential AA | 0.45 ± 0.02 a | 0.52 ± 0.03 b |
| Fructose | 108.67 ± 4.32 a | 116.93 ± 4.62 a |
| Glucose | 271.23 ± 10.84 a | 278.38 ± 11.19 a |
| Sucrose | 1204.09 ± 48.15 a | 1248.32 ± 49.28 a |
| Raw fat | 2.54 ± 0.10 a | 2.57 ± 0.10 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maslennikova, D.; Knyazeva, I.; Vershinina, O.; Titenkov, A.; Lastochkina, O. Seed Treatment with Sodium Nitroprusside Ensures a Long-Term Physiological and Protective Effect on Wheat under Salinity. Life 2023, 13, 1499. https://doi.org/10.3390/life13071499
Maslennikova D, Knyazeva I, Vershinina O, Titenkov A, Lastochkina O. Seed Treatment with Sodium Nitroprusside Ensures a Long-Term Physiological and Protective Effect on Wheat under Salinity. Life. 2023; 13(7):1499. https://doi.org/10.3390/life13071499
Chicago/Turabian StyleMaslennikova, Dilara, Inna Knyazeva, Oksana Vershinina, Andrey Titenkov, and Oksana Lastochkina. 2023. "Seed Treatment with Sodium Nitroprusside Ensures a Long-Term Physiological and Protective Effect on Wheat under Salinity" Life 13, no. 7: 1499. https://doi.org/10.3390/life13071499