
An Overview of the Microbiota of the Human Urinary Tract in Health and Disease: Current Issues and Perspectives

 , ,
, , 
Abstract
:1. Introduction
- Current microbiology diagnostic tests
- The sample can be observed: (a) after staining, for the targeted search for specific microbial/pathogen groups (i.e., Gram staining)., (b) “fresh observation” without staining. They are both suitable for the study of some biological properties (shape, organization, motility, chemical reactivity);
- The choice of the microscopic technique to be used depends on the pathogen whose presence is suspected: (a) bright field microscopy (frequent use), (b) dark field microscopy (e.g., for Treponema pallidum), (c) phase contrast microscopy (frequent use), and (d) fluorescence microscopy (greater sensitivity because antibodies are used to search the microorganism)
- With advanced cultivation techniques and molecular approaches, wide numbers and variations have been identified types of microorganisms in healthy and asymptomatic individuals.
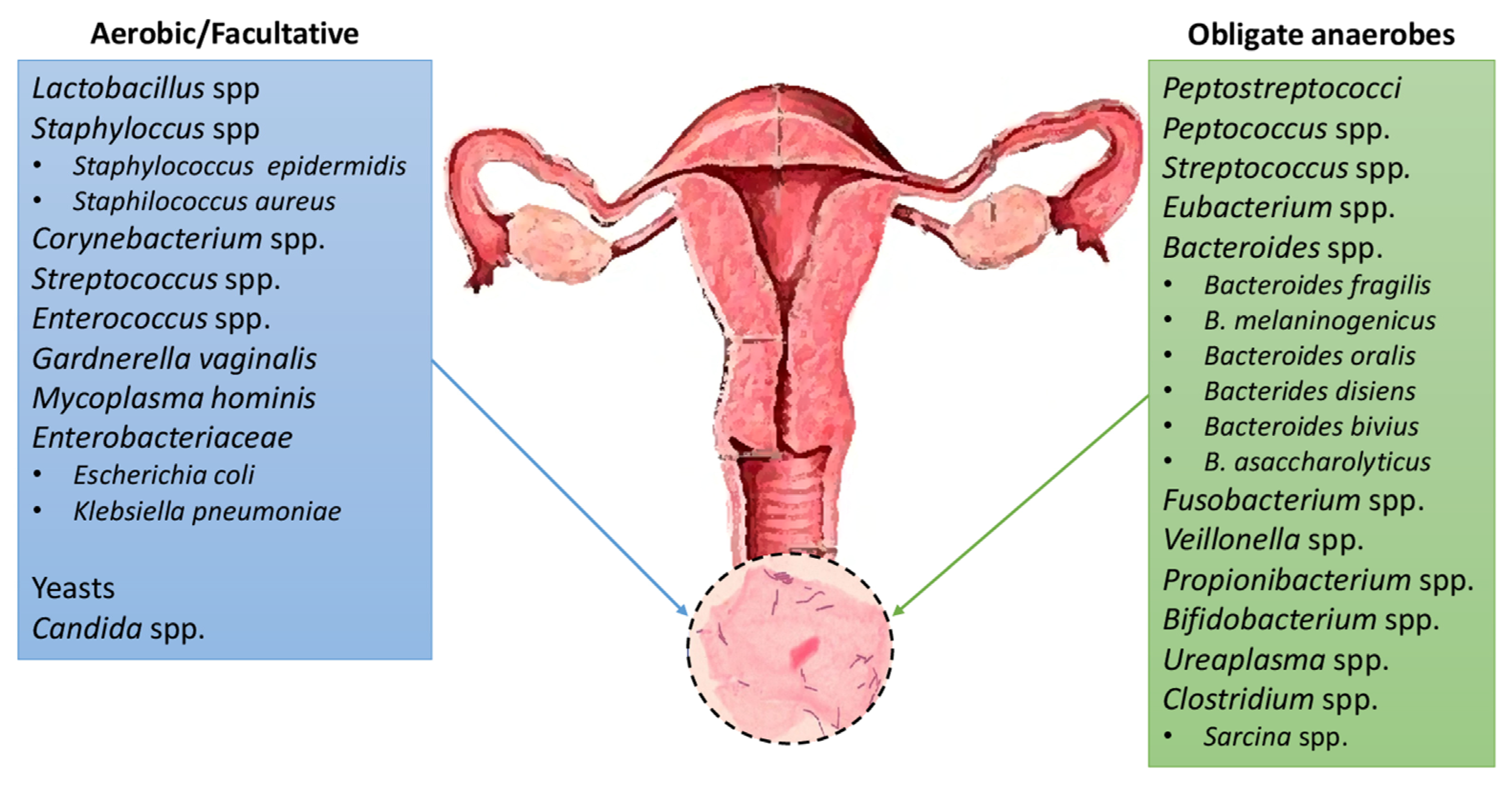
2. The Microbiota Composition of the Urinary Tract
3. The “Behavior” of the Microbiota in Urinary Tract Diseases
3.1. Role of the Microbiota Gut/Kidney and Bladder/Gut/Brain Axes
3.2. Urinary Tract Infections (UTIs)
3.3. Urgency Urinary Incontinence (UUI)
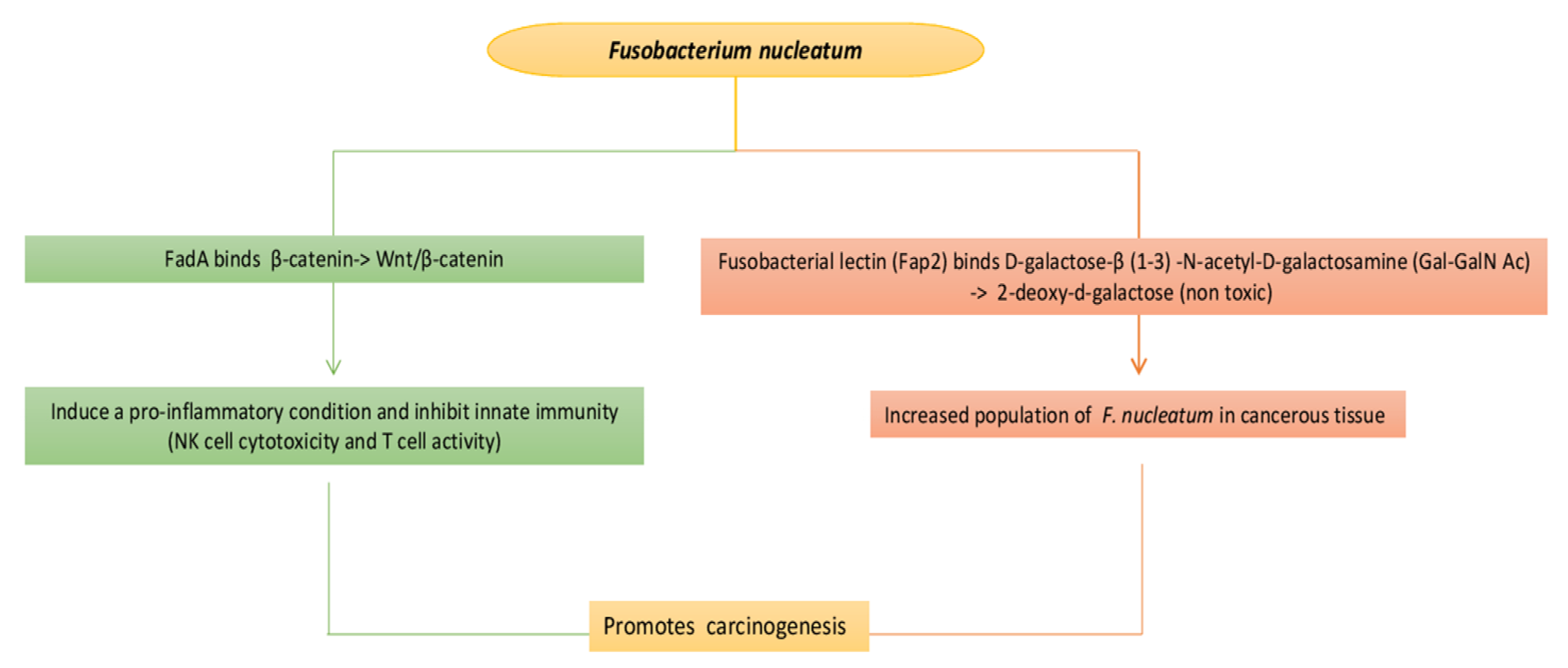
3.4. Bladder Cancer
4. The Microbiota of Female and Male Reproductive System
4.1. Reproductive System Microbiota and Immune Functions during Conception
4.2. The Role of the Genital Microbiota on Fetus Health
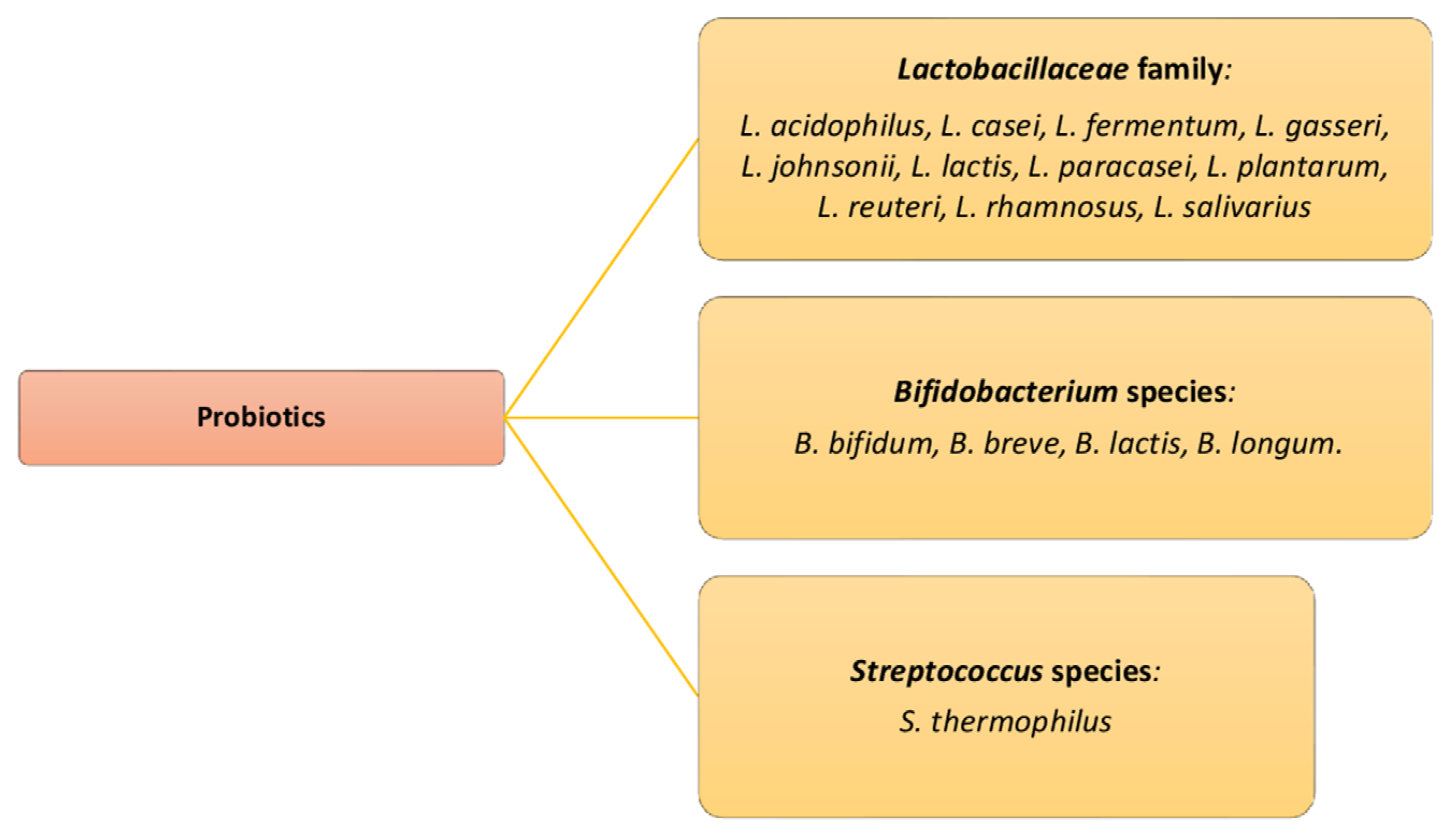
5. The Role of Probiotics as Adjuvants for the Eubiosis in the Urinary Tract
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aragón, I.M.; Herrera-Imbroda, B.; Queipo-Ortuño, M.I.; Castillo, E.; Del Moral, J.S.; Gómez-Millán, J.; Yucel, G.; Lara, M.F. The Urinary Tract Microbiome in Health and Disease. Eur. Urol. Focus 2018, 4, 128–138. [Google Scholar] [CrossRef]
- Ipe, D.S.; Horton, E.; Ulett, G.C. The Basics of Bacteriuria: Strategies of Microbes for Persistence in Urine. Front. Cell. Infect. Microbiol. 2016, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas-White, K.; Brady, M.; Wolfe, A.J.; Mueller, E.R. The bladder is not sterile: History and current discoveries on the urinary microbiome. Curr. Bladder Dysfunct. Rep. 2016, 11, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, A.J.; Toh, E.; Shibata, N.; Rong, R.; Kenton, K.; Fitzgerald, M.; Mueller, E.R.; Schreckenberger, P.; Dong, Q.; Nelson, D.E.; et al. Evidence of uncultivated bacteria in the adult female bladder. J. Clin. Microbiol. 2012, 50, 1376–1383. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Rautava, S.; Aakko, J.; Isolauri, E.; Salminen, S. Human gut colonisation may be initiated in utero by distinct microbial communities in the placenta and amniotic fluid. Sci. Rep. 2016, 6, 23129. [Google Scholar] [CrossRef] [Green Version]
- Franasiak, J.M.; Scott, R.T., Jr. Introduction: Microbiome in human reproduction. Fertil. Steril. 2015, 104, 1341–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, E.R.; Wolfe, A.J.; Brubaker, L. Female urinary microbiota. Curr. Opin. Urol. 2017, 27, 282–286. [Google Scholar] [CrossRef]
- Whiteside, S.A.; Razvi, H.; Dave, S.; Reid, G.; Burton, J.P. The microbiome of the urinary tract—A role beyond infection. Nat. Rev. Urol. 2015, 12, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Mody, L.; Juthani-Mehta, M. Urinary tract infections in older women: A clinical review. JAMA 2014, 26, 844–854. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, H.; Nederbragt, A.J.; Lagesen, K.; Jeansson, S.L.; Jakobsen, K.S. Assessing diversity of the female urine microbiota by high throughput sequencing of 16S rDNA amplicons. BMC Microbiol. 2011, 11, 244. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.E.; Dong, Q.; Van der Pol, B.; Toh, E.; Fan, B.; Katz, B.P.; Mi, D.; Rong, R.; Weinstock, G.M.; Sodergren, E.; et al. Bacterial communities of the coronal sulcus and distal urethra of adolescent males. PLoS ONE 2012, 7, e36298. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D.A.; Brown, R.; Williams, J.; White, P.; Jacobson, S.K.; Marchesi, J.R.; Drake, M.J. The human urinary microbiome; bacterial DNA in voided urine of asymptomatic adults. Front. Cell. Infect. Microbiol. 2013, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Roth, R.S.; Liden, M.; Huttner, A. The urobiome in men and women: A clinical review. Clin. Microbiol. Infect. 2022, 24. in press. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Santacroce, L.; Imbimbo, C.; Ballini, A.; Crocetto, F.; Scacco, S.; Cantore, S.; Di Zazzo, E.; Colella, M.; Jirillo, E. Testicular Immunity and Its Connection with the Microbiota. Physiological and Clinical Implications in the Light of Personalized Medicine. J. Pers. Med. 2022, 12, 1335. [Google Scholar] [CrossRef] [PubMed]
- Brubaker, L.; Wolfe, A. The urinary microbiota: A paradigm shift for bladder disorders? Curr. Opin. Obstet. Gynecol. 2016, 28, 407–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepor, H. Pathophysiology of lower urinary tract symptoms in the aging male population. Rev. Urol. 2005, 7 (Suppl. S7), S3–S11. [Google Scholar]
- Lee, K.W.; Song, H.Y.; Kim, Y.H. The microbiome in urological diseases. Investig. Clin. Urol. 2020, 61, 338–348. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [Green Version]
- Stavropoulou, E.; Kantartzi, K.; Tsigalou, C.; Konstantinidis, T.; Romanidou, G.; Voidarou, C.; Bezirtzoglou, E. Focus on the Gut-Kidney Axis in Health and Disease. Front. Med. 2021, 7, 620102. [Google Scholar] [CrossRef]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The Gut Microbiome as a Major Regulator of the Gut-Skin Axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Domenico, M.; Ballini, A.; Boccellino, M.; Scacco, S.; Lovero, R.; Charitos, I.A.; Santacroce, L. The Intestinal Microbiota May Be a Potential Theranostic Tool for Personalized Medicine. J. Pers. Med. 2022, 12, 523. [Google Scholar] [CrossRef] [PubMed]
- Santacroce, L.; Man, A.; Charitos, I.A.; Haxhirexha, K.; Topi, S. Current knowledge about the connection between health status and gut microbiota from birth to elderly. A narrative review. Front. Biosci. 2021, 26, 135–148. [Google Scholar] [CrossRef]
- Khoury, T.; Tzukert, K.; Abel, R.; Abu Rmeileh, A.; Levi, R.; Ilan, Y. The gut-kidney axis in chronic renal failure: A new potential target for therapy. Hemodial. Int. 2017, 21, 323–334. [Google Scholar] [CrossRef]
- Evenepoel, P.; Poesen, R.; Meijers, B. The gut-kidney axis. Pediatr. Nephrol. 2017, 32, 2005–2014. [Google Scholar] [CrossRef]
- Leue, C.; Kruimel, J.; Vrijens, D.; Masclee, A.; van Os, J.; van Koeveringe, G. Functional urological disorders: A sensitized defence response in the bladder-gut-brain axis. Nat. Rev. Urol. 2017, 14, 153–163. [Google Scholar] [CrossRef]
- Cataldi, S.; Poli, L.; Şahin, F.N.; Patti, A.; Santacroce, L.; Bianco, A.; Greco, G.; Ghinassi, B.; Di Baldassarre, A.; Fischetti, F. The Effects of Physical Activity on the Gut Microbiota and the Gut-Brain Axis in Preclinical and Human Models: A Narrative Review. Nutrients 2022, 14, 3293. [Google Scholar] [CrossRef]
- Wegierska, A.E.; Charitos, I.A.; Topi, S.; Potenza, M.A.; Montagnani, M.; Santacroce, L. The Connection between Physical Exercise and Gut Microbiota: Implications for Competitive Sports Athletes. Sports Med. 2022, 52, 2355–2369. [Google Scholar] [CrossRef]
- Miller, L.G.; Tang, A.W. Treatment of uncomplicated urinary tract infections in an era of increasing antimicrobial resistance. Mayo. Clin. Proc. 2004, 79, 1048–1053. [Google Scholar] [CrossRef] [Green Version]
- Brubaker, L.; Wolfe, A.J. The new world of the urinary microbiota in women. Am. J. Obstet. Gynecol. 2015, 213, 644–649. [Google Scholar] [CrossRef]
- Khasriya, R.; Sathiananthamoorthy, S.; Ismail, S.; Kelsey, M.; Wilson, M.; Rohn, J.L.; Malone-Lee, J. Spectrum of bacterial colonization associated with urothelial cells from patients with chronic lower urinary tract symptoms. J. Clin. Microbiol. 2013, 51, 2054–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J. Microbiome in the urinary system—A review. AIMS Microbiol. 2017, 3, 143. [Google Scholar] [CrossRef]
- Prete, R.D.; Ronga, L.; Addati, G.; Magrone, R.; Abbasciano, A.; Carlo, D.D.; Santacroce, L. A Retrospective Study about the Impact of Switching from Nested PCR to Multiplex Real-Time PCR on the Distribution of the Human Papillomavirus (HPV) Genotypes. Medicina 2019, 55, 418. [Google Scholar] [CrossRef] [Green Version]
- Gatt, M.; Reddy, B.S.; MacFie, J. Bacterial translocation in the critically ill—Evidence and methods of prevention. Aliment. Pharmacol. Ther. 2007, 25, 741–757. [Google Scholar] [CrossRef]
- Nagpal, R.; Yadav, H. Bacterial Translocation from the Gut to the Distant Organs: An Overview. Ann. Nutr. Metab. 2017, 71 (Suppl. S1), 11–16. [Google Scholar] [CrossRef] [PubMed]
- Krentz, T.; Allen, S. Bacterial translocation in critical illness. J. Small Anim. Pract. 2017, 58, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Gogos, C.A. The role of the intestinal immune system in health and disease. Arch. Hell. Med. 2003, 20 (Suppl. A), 30–32. [Google Scholar]
- Ungaro, F.; Tacconi, C.; D’Alessio, S. Beyond Intestinal Barrier: The Blood Endothelium as a Second Wall of Defense against Bacterial Invasion. Gastroenterology 2016, 150, 1678–1686. [Google Scholar] [CrossRef] [Green Version]
- Lugea, A.; Salas, A.; Casalot, J.; Guarner, F.; Malagelada, J.R. Surface hydrophobicity of the rat colonic mucosa is a defensive barrier against macromolecules and toxins. Gut 2000, 46, 515–521. [Google Scholar] [CrossRef] [Green Version]
- Schirinzi, A.; Cazzolla, A.P.; Mascolo, E.; Palmieri, G.; Pesce, F.; Gesualdo, L.; Santacroce, L.; Ballini, A.; Lovero, R.; Di Serio, F. Determination of the Upper Reference Limit of Human Epididymis Secretory Protein 4 (HE4) in Healthy Male Individuals and Correlation with Renal and Fertility Markers. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 912–918. [Google Scholar] [CrossRef]
- Hart, A.L.; Stagg, A.J.; Frame, M.; Graffner, H.; Glise, H.; Falk, P.; Kamm, M.A. The role of the gut flora in health and disease, and its modification as therapy. Aliment. Pharmacol. Ther. 2002, 16, 1383–1393. [Google Scholar] [CrossRef]
- Vaishnavi, C. Translocation of gut flora and its role in sepsis. Indian J. Med. Microbiol. 2013, 31, 334–342. [Google Scholar] [CrossRef]
- Barnett, C.; Nazzal, L.; Goldfarb, D.S.; Blaser, M.J. The Presence of Oxalobacter formigenes in the Microbiome of Healthy Young Adults. J. Urol. 2016, 195, 499–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, D.W.; Kelly, J.P.; Curhan, G.C.; Anderson, T.E.; Dretler, S.P.; Preminger, G.M.; Cave, D.R. Oxalobacter formigenes may reduce the risk of calcium oxalate kidney stones. J. Am. Soc. Nephrol. 2008, 19, 1197–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santacroce, L.; Bottalico, L.; Topi, S.; Castellaneta, F.; Charitos, I.A. The “Scourge of the Renaissance”. A Short Review about Treponema pallidum infection. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Nelson, D.E.; Toh, E.; Diao, L.; Gao, X.; Fortenberry, J.D.; Van der Pol, B. The microbial communities in male first catch urine are highly similar to those in paired urethral swab specimens. PLoS ONE 2011, 6, e19709. [Google Scholar] [CrossRef] [Green Version]
- Malmsten, U.G.; Molander, U.; Peeker, R.; Irwin, D.E.; Milsom, I. Urinary incontinence, overactive bladder, and other lower urinary tract symptoms: A longitudinal population-based survey in men aged 45–103 years. Eur. Urol. 2010, 58, 149–156. [Google Scholar] [CrossRef]
- Martínez-Agulló, E.; Ruiz-Cerdá, J.L.; Arlandis, S.; Rebollo, P.; Pérez, M.; Chaves, J.; Grupo de Estudio Cooperativo EPICC. Análisis del síndrome de vejiga hiperactiva y de la incontinencia urinaria en mujeres laboralmente activas entre 25–64 años. Estudio EPICC [Analysis of overactive bladder and urinary incontinence in working women aged between 25 and 64 years. EPICC study]. Actas Urológicas Españolas 2010, 34, 618–624. (In Spanish) [Google Scholar]
- Pearce, M.M.; Hilt, E.E.; Rosenfeld, A.B.; Zilliox, M.J.; Thomas-White, K.; Fok, C.; Kliethermes, S.; Schreckenberger, P.C.; Brubaker, L.; Gai, X.; et al. The female urinary microbiome: A comparison of women with and without urgency urinary incontinence. mBio 2014, 5, e01283-14. [Google Scholar] [CrossRef] [Green Version]
- Papavasileiou, G.; Tsilingiris, D.; Spyrou, N.; Vallianou, N.G.; Karampela, I.; Magkos, F.; Dalamaga, M. Obesity and main urologic cancers: Current systematic evidence, novel biological mechanisms, perspectives and challenges. Semin. Cancer Biol. 2023, 91, 70–98. [Google Scholar] [CrossRef]
- Metts, M.C.; Metts, J.C.; Milito, S.J.; Thomas, C.R., Jr. Bladder cancer: A review of diagnosis and management. J. Natl. Med. Assoc. 2000, 92, 285–294. [Google Scholar] [PubMed]
- Topi, S.; Bottalico, L.; Charitos, I.A.; Colella, M.; Di Domenico, M.; Palmirotta, R.; Santacroce, L. Biomolecular Mechanisms of Autoimmune Diseases and Their Relationship with the Resident Microbiota: Friend or Foe? Pathophysiology 2022, 29, 507–536. [Google Scholar] [CrossRef] [PubMed]
- Angelini, K. Pelvic Floor Muscle Training to Manage Overactive Bladder and Urinary Incontinence. Nurs. Womens Health 2017, 21, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Friedell, G.H.; Jacobs, J.B.; Nagy, G.K.; Cohen, S.M. The pathogenesis of bladder cancer. Am. J. Pathol. 1977, 89, 431–442. [Google Scholar] [PubMed]
- Bajic, P.; Wolfe, A.J.; Gupta, G.N. The Urinary Microbiome: Implications in Bladder Cancer Pathogenesis and Therapeutics. Urology 2019, 126, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Stone, L. Urine microbiota differ in bladder cancer. Nat. Rev. Urol. 2023, 20, 7. [Google Scholar] [CrossRef]
- Bučević Popović, V.; Šitum, M.; Chow, C.T.; Chan, L.S.; Roje, B.; Terzić, J. The urinary microbiome associated with bladder cancer. Sci. Rep. 2018, 8, 12157. [Google Scholar] [CrossRef] [Green Version]
- Hourigan, S.K.; Zhu, W.; SW Wong, W.; Clemency, N.C.; Provenzano, M.; Vilboux, T.; Niederhuber, J.E.; Deeken, J.; Chung, S.; McDaniel-Wiley, K.; et al. Studying the urine microbiome in superficial bladder cancer: Samples obtained by midstream voiding versus cystoscopy. BMC Urol. 2020, 20, 5. [Google Scholar] [CrossRef]
- Polimeno, L.; Barone, M.; Mosca, A.; Viggiani, M.T.; Joukar, F.; Mansour-Ghanaei, F.; Mavaddati, S.; Daniele, A.; Debellis, L.; Bilancia, M.; et al. Soy Metabolism by Gut Microbiota from Patients with Precancerous Intestinal Lesions. Microorganisms 2020, 8, 469. [Google Scholar] [CrossRef] [Green Version]
- Al-Nasiry, S.; Ambrosino, E.; Schlaepfer, M.; Morré, S.A.; Wieten, L.; Voncken, J.W.; Spinelli, M.; Mueller, M.; Kramer, B.W. The Interplay between Reproductive Tract Microbiota and Immunological System in Human Reproduction. Front. Immunol. 2020, 11, 378. [Google Scholar] [CrossRef]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen-gut microbiome axis: Physiological and clinical implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepargneur, J.P.; Rousseau, V. Rôle protecteur de la flore de Doderleïn [Protective role of the Doderleïn flora]. J. Gynecol. Obstet. Biol. Reprod. 2002, 31, 485–494. [Google Scholar]
- García-Velasco, J.A.; Budding, D.; Campe, H.; Malfertheiner, S.F.; Hamamah, S.; Santjohanser, C.; Schuppe-Koistinen, I.; Nielsen, H.S.; Vieira-Silva, S.; Laven, J. The reproductive microbiome—Clinical practice recommendations for fertility specialists. Reprod. Biomed. Online 2020, 41, 443–453. [Google Scholar] [CrossRef]
- Witkin, S.S.; Linhares, I.M. Why do lactobacilli dominate the human vaginal microbiota? BJOG 2017, 124, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Lu, Y.; Chen, T.; Li, R. The Female Vaginal Microbiome in Health and Bacterial Vaginosis. Front. Cell. Infect. Microbiol. 2021, 11, 631972. [Google Scholar] [CrossRef]
- Green, K.A.; Zarek, S.M.; Catherino, W.H. Gynecologic health and disease in relation to the microbiome of the female reproductive tract. Fertil. Steril. 2015, 104, 1351–1357. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.S.; Li, L. Quantifying the human vaginal community state types (CSTs) with the species specificity index. PeerJ. 2017, 5, e3366. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Lopez, V.; Cook, R.L.; Sobel, J.D. Emerging role of lactobacilli in the control and maintenance of the vaginal bacterial microflora. Rev. Infect. Dis. 1990, 12, 856–872. [Google Scholar] [CrossRef]
- Nakama, C.; Thompson, B.; Szybala, C.; McBeth, A.; Dobner, P.; Zwickey, H. The Continuum of Microbial Ecosystems along the Female Reproductive Tract: Implications for Health and Fertility. Pathogens 2022, 11, 1244. [Google Scholar] [CrossRef]
- Jepsen, I.E.; Saxtorph, M.H.; Englund, A.L.M.; Petersen, K.B.; Wissing, M.L.M.; Hviid, T.V.F.; Macklon, N. Probiotic treatment with specific lactobacilli does not improve an unfavorable vaginal microbiota prior to fertility treatment-A randomized, double-blinded, placebo-controlled trial. Front. Endocrinol. 2022, 13, 1057022. [Google Scholar] [CrossRef]
- Zhou, X.; Bent, S.J.; Schneider, M.G.; Davis, C.C.; Islam, M.R.; Forney, L.J. Characterization of vaginal microbial communities in adult healthy women using cultivation-independent methods. Microbiology 2004, 150, 2565–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsson, T.; Forsum, U. Changes in the predominant human Lactobacillus flora during in vitro fertilisation. Ann. Clin. Microbiol. Antimicrob. 2008, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4680–4687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajer, P.; Brotman, R.M.; Bai, G.; Sakamoto, J.; Schütte, U.M.; Zhong, X.; Koenig, S.S.; Fu, L.; Ma, Z.S.; Zhou, X.; et al. Temporal dynamics of the human vaginal microbiota. Sci. Transl. Med. 2012, 4, 132ra52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santacroce, L.; Carlaio, R.G.; Bottalico, L. Does it make sense that diabetes is reciprocally associated with periodontal disease? Endocr. Metab. Immune Disord. Drug Targets 2010, 10, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Selman, H.; Mariani, M.; Barnocchi, N.; Mencacci, A.; Bistoni, F.; Arena, S.; Pizzasegale, S.; Brusco, G.F.; Angelini, A. Examination of bacterial contamination at the time of embryo transfer, and its impact on the IVF/pregnancy outcome. J. Assist. Reprod. Genet. 2007, 24, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Tsonis, O.; Gkrozou, F.; Paschopoulos, M. Microbiome affecting reproductive outcome in ARTs. J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 102036. [Google Scholar] [CrossRef]
- Kaur, H.; Merchant, M.; Haque, M.M.; Mande, S.S. Crosstalk between Female Gonadal Hormones and Vaginal Microbiota Across Various Phases of Women’s Gynecological Lifecycle. Front. Microbiol. 2020, 11, 551. [Google Scholar] [CrossRef]
- Bardos, J.; Fiorentino, D.; Longman, R.E.; Paidas, M. Immunological Role of the Maternal Uterine Microbiome in Pregnancy: Pregnancies Pathologies and Alterated Microbiota. Front. Immunol. 2020, 10, 2823. [Google Scholar] [CrossRef] [Green Version]
- van Oostrum, N.; De Sutter, P.; Meys, J.; Verstraelen, H. Risks associated with bacterial vaginosis in infertility patients: A systematic review and meta-analysis. Hum. Reprod. 2013, 28, 1809–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haahr, T.; Jensen, J.S.; Thomsen, L.; Duus, L.; Rygaard, K.; Humaidan, P. Abnormal vaginal microbiota may be associated with poor reproductive outcomes: A prospective study in IVF patients. Hum. Reprod. 2016, 31, 795–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angela, A.; Raffaele, D.P.; Federica, R.; Adriana, M.; Luigi, S.; Luigi, R. Multi-year prevalence and macrolide resistance of Mycoplasma genitalium in clinical samples from a southern Italian hospital. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 893–895. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.; Franasiak, J.M. Endometrial microbiota-new player in town. Fertil. Steril. 2017, 108, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Hemsell, D.L.; Obregon, V.L.; Heard, M.C.; Nobles, B.J. Endometrial bacteria in asymptomatic, nonpregnant women. J. Reprod. Med. 1989, 34, 872–874. [Google Scholar]
- Rampersaud, R.; Randis, T.M.; Ratner, A.J. Microbiota of the upper and lower genital tract. Semin. Fetal Neonatal Med. 2012, 17, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, C.M.; Haick, A.; Nkwopara, E.; Garcia, R.; Rendi, M.; Agnew, K.; Fredricks, D.N.; Eschenbach, D. Colonization of the upper genital tract by vaginal bacterial species in nonpregnant women. Am. J. Obstet. Gynecol. 2015, 212, e1–e9. [Google Scholar] [CrossRef] [Green Version]
- Moreno, I.; Codoñer, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazán, J.; Alonso, R.; Alamá, P.; Remohí, J.; Pellicer, A.; et al. Evidence that the endometrial microbiota has an effect on implantation success or failure. Am. J. Obstet. Gynecol. 2016, 215, 684–703. [Google Scholar] [CrossRef] [Green Version]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef] [Green Version]
- Williams, N.; Vella, R.; Zhou, Y.; Gao, H.; Mass, K.; Townsel, C.; Campbell, W.; Luo, G. Investigating the origin of the fetal gut and placenta microbiome in twins. J. Matern. Fetal Neonatal Med. 2021, 35, 7025–7035. [Google Scholar] [CrossRef]
- van Nimwegen, F.A.; Penders, J.; Stobberingh, E.E.; Postma, D.S.; Koppelman, G.H.; Kerkhof, M.; Reijmerink, N.E.; Dompeling, E.; van den Brandt, P.A.; Ferreira, I.; et al. Mode and place of delivery, gastrointestinal microbiota, and their influence on asthma and atopy. J. Allergy Clin. Immunol. 2011, 128, e1–e3. [Google Scholar] [CrossRef]
- Steel, J.H.; Malatos, S.; Kennea, N.; Edwards, A.D.; Miles, L.; Duggan, P.; Reynolds, P.R.; Feldman, R.G.; Sullivan, M.H. Bacteria and inflammatory cells in fetal membranes do not always cause preterm labor. Pediatr. Res. 2005, 57, 404–411. [Google Scholar] [CrossRef] [Green Version]
- Gschwind, R.; Fournier, T.; Butel, M.J.; Wydau-Dematteis, S. Établissement du microbiote—Une colonisation in utero déterminante pour la santé future? [Microbiota establishment: An in utero colonization decisive for future health?]. Med. Sci. 2018, 34, 331–337. (In French) [Google Scholar] [CrossRef] [Green Version]
- Kroon, B.; Hart, R.J.; Wong, B.M.; Ford, E.; Yazdani, A. Antibiotics prior to embryo transfer in ART. Cochrane Database Syst. Rev. 2012, 3, CD008995. [Google Scholar] [CrossRef] [PubMed]
- Salim, R.; Ben-Shlomo, I.; Colodner, R.; Keness, Y.; Shalev, E. Bacterial colonization of the uterine cervix and success rate in assisted reproduction: Results of a prospective survey. Hum. Reprod. 2002, 17, 337–340. [Google Scholar] [CrossRef] [Green Version]
- Egbase, P.E.; al-Sharhan, M.; al-Othman, S.; al-Mutawa, M.; Udo, E.E.; Grudzinskas, J.G. Incidence of microbial growth from the tip of the embryo transfer catheter after embryo transfer in relation to clinical pregnancy rate following in-vitro fertilization and embryo transfer. Hum. Reprod. 1996, 11, 1687–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyagi, M.; Mekaru, K.; Tanaka, S.E.; Arai, W.; Ashikawa, K.; Sakuraba, Y.; Nakamura, R.; Oishi, S.; Akamine, K.; Aoki, Y. Endometrial and vaginal microbiomes influence assisted reproductive technology outcomes. JBRA Assist. Reprod. 2023, 27, 267–281. [Google Scholar] [CrossRef]
- Peric, A.; Weiss, J.; Vulliemoz, N.; Baud, D.; Stojanov, M. Bacterial Colonization of the Female Upper Genital Tract. Int. J. Mol. Sci. 2019, 20, 3405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heeren, A.M.; van Iperen, L.; Klootwijk, D.B.; de Melo Bernardo, A.; Roost, M.S.; Gomes Fernandes, M.M.; Louwe, L.A.; Hilders, C.G.; Helmerhorst, F.M.; van der Westerlaken, L.A.; et al. Development of the follicular basement membrane during human gametogenesis and early folliculogenesis. BMC Dev. Biol. 2015, 15, 4. [Google Scholar] [CrossRef] [Green Version]
- Usman, S.F.; Shuaibu, I.R.; Durojaiye, K.; Medugu, N.; Iregbu, K.C. The presence of microorganisms in follicular fluid and its effect on the outcome of in vitro fertilization-embryo transfer (IVF-ET) treatment cycles. PLoS ONE 2021, 16, e0246644. [Google Scholar] [CrossRef]
- Da Broi, M.G.; Giorgi, V.S.I.; Wang, F.; Keefe, D.L.; Albertini, D.; Navarro, P.A. Influence of follicular fluid and cumulus cells on oocyte quality: Clinical implications. J. Assist. Reprod. Genet. 2018, 35, 735–751. [Google Scholar] [CrossRef]
- Pelzer, E.S.; Allan, J.A.; Cunningham, K.; Mengersen, K.; Allan, J.M.; Launchbury, T.; Beagley, K.; Knox, C.L. Microbial colonization of follicular fluid: Alterations in cytokine expression and adverse assisted reproduction technology outcomes. Hum. Reprod. 2011, 26, 1799–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiecka, A.; Macura, B.; Szczepanik, M. Can Lactobacillus spp. be a Factor Reducing the Risk of Miscarriage? Pol. J. Microbiol. 2021, 70, 431–446. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Song, X.; Wei, W.; Zhong, H.; Dai, J.; Lan, Z.; Li, F.; Yu, X.; Feng, Q.; Wang, Z.; et al. The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat. Commun. 2017, 8, 875. [Google Scholar] [CrossRef] [Green Version]
- Mändar, R.; Punab, M.; Borovkova, N.; Lapp, E.; Kiiker, R.; Korrovits, P.; Metspalu, A.; Krjutškov, K.; Nõlvak, H.; Preem, J.K.; et al. Complementary seminovaginal microbiome in couples. Res. Microbiol. 2015, 166, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, A.A.; Desmarais, B.M.; Yin, H.Z.; Loverde, J.; Eyre, R.C. Detection and identification of bacterial DNA in semen. Fertil. Steril. 2008, 90, 1744–1756. [Google Scholar] [CrossRef]
- Weng, S.L.; Chiu, C.M.; Lin, F.M.; Huang, W.C.; Liang, C.; Yang, T.; Yang, T.L.; Liu, C.Y.; Wu, W.Y.; Chang, Y.A.; et al. Bacterial communities in semen from men of infertile couples: Metagenomic sequencing reveals relationships of seminal microbiota to semen quality. PLoS ONE 2014, 9, e110152. [Google Scholar] [CrossRef] [Green Version]
- Puerta Suárez, J.; Cardona Maya, W.D. Microbiota, Prostatitis, and Fertility: Bacterial Diversity as a Possible Health Ally. Adv. Urol. 2021, 2021, 1007366. [Google Scholar] [CrossRef]
- Baud, D.; Pattaroni, C.; Vulliemoz, N.; Castella, V.; Marsland, B.J.; Stojanov, M. Sperm Microbiota and Its Impact on Semen Parameters. Front. Microbiol. 2019, 10, 234. [Google Scholar] [CrossRef] [Green Version]
- Santacroce, L.; Colella, M.; Charitos, I.A. The Persistence and Increase in Sexually Transmitted Diseases (STDs) to Pandemic Levels. Venereology 2022, 1, 2–8. [Google Scholar] [CrossRef]
- Zozaya, M.; Ferris, M.J.; Siren, J.D.; Lillis, R.; Myers, L.; Nsuami, M.J.; Eren, A.M.; Brown, J.; Taylor, C.M.; Martin, D.H. Bacterial communities in penile skin, male urethra, and vaginas of heterosexual couples with and without bacterial vaginosis. Microbiome 2016, 4, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbonetti, A.; Cinque, B.; Vassallo, M.R.; Mineo, S.; Francavilla, S.; Cifone, M.G.; Francavilla, F. Effect of vaginal probiotic lactobacilli on in vitro-induced sperm lipid peroxidation and its impact on sperm motility and viability. Fertil. Steril. 2011, 95, 2485–2488. [Google Scholar] [CrossRef]
- Barbonetti, A.; Vassallo, M.R.; Cinque, B.; Filipponi, S.; Mastromarino, P.; Cifone, M.G.; Francavilla, S.; Francavilla, F. Soluble products of Escherichia coli induce mitochondrial dysfunction-related sperm membrane lipid peroxidation which is prevented by lactobacilli. PLoS ONE 2013, 8, e83136. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Zhou, X.; Zhong, X.; Settles, M.L.; Herring, J.; Wang, L.; Abdo, Z.; Forney, L.J.; Xu, C. Microbiota of the seminal fluid from healthy and infertile men. Fertil. Steril. 2013, 100, 1261–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundy, S.D.; Vij, S.C.; Rezk, A.H.; Cohen, J.A.; Bajic, P.; Ramasamy, R. The microbiome of the infertile male. Curr. Opin. Urol. 2020, 30, 355–362. [Google Scholar] [CrossRef]
- Koren, O.; Goodrich, J.K.; Cullender, T.C.; Spor, A.; Laitinen, K.; Bäckhed, H.K.; Gonzalez, A.; Werner, J.J.; Angenent, L.T.; Knight, R.; et al. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell 2012, 150, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Amabebe, E.; Anumba, D.O.C. Female Gut and Genital Tract Microbiota-Induced Crosstalk and Differential Effects of Short-Chain Fatty Acids on Immune Sequelae. Front. Immunol. 2020, 11, 2184. [Google Scholar] [CrossRef]
- Aagaard, K.; Riehle, K.; Ma, J.; Segata, N.; Mistretta, T.A.; Coarfa, C.; Raza, S.; Rosenbaum, S.; Van den Veyver, I.; Milosavljevic, A.; et al. A metagenomic approach to characterization of the vaginal microbiome signature in pregnancy. PLoS ONE 2012, 7, e36466. [Google Scholar] [CrossRef]
- Romero, R.; Hassan, S.S.; Gajer, P.; Tarca, A.L.; Fadrosh, D.W.; Nikita, L.; Galuppi, M.; Lamont, R.F.; Chaemsaithong, P.; Miranda, J.; et al. The composition and stability of the vaginal microbiota of normal pregnant women is different from that of non-pregnant women. Microbiome 2014, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Robertson, S.A.; Chin, P.Y.; Glynn, D.J.; Thompson, J.G. Peri-conceptual cytokines--setting the trajectory for embryo implantation, pregnancy and beyond. Am. J. Reprod. Immunol. 2011, 66 (Suppl. S1), 2–10. [Google Scholar] [CrossRef]
- Robertson, S.A. Immune regulation of conception and embryo implantation-all about quality control? J. Reprod. Immunol. 2010, 85, 51–57. [Google Scholar] [CrossRef]
- Hsu, P.; Nanan, R. Foetal immune programming: Hormones, cytokines, microbes and regulatory T cells. J. Reprod. Immunol. 2014, 104–105, 2–7. [Google Scholar] [CrossRef]
- Tal, R.; Taylor, H.S. Endocrinology of Pregnancy. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Wang, W.; Sung, N.; Gilman-Sachs, A.; Kwak-Kim, J. T Helper (Th) Cell Profiles in Pregnancy and Recurrent Pregnancy Losses: Th1/Th2/Th9/Th17/Th22/Tfh Cells. Front. Immunol. 2020, 11, 2025. [Google Scholar] [CrossRef] [PubMed]
- Busse, M.; Campe, K.J.; Nowak, D.; Schumacher, A.; Plenagl, S.; Langwisch, S.; Tiegs, G.; Reinhold, A.; Zenclussen, A.C. IL-10 producing B cells rescue mouse fetuses from inflammation-driven fetal death and are able to modulate T cell immune responses. Sci. Rep. 2019, 9, 9335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantos, K.; Grigoriadis, S.; Maziotis, E.; Pistola, K.; Xystra, P.; Pantou, A.; Kokkali, G.; Pappas, A.; Lambropoulou, M.; Sfakianoudis, K.; et al. The Role of Interleukins in Recurrent Implantation Failure: A Comprehensive Review of the Literature. Int. J. Mol. Sci. 2022, 23, 2198. [Google Scholar] [CrossRef]
- Simopoulou, M.; Sfakianoudis, K.; Maziotis, E.; Tsioulou, P.; Giannelou, P.; Grigoriadis, S.; Pantou, A.; Anifandis, G.; Christopoulos, P.; Pantos, K.; et al. Investigating the Optimal Time for Intrauterine Human Chorionic Gonadotropin Infusion in Order to Improve IVF Outcome: A Systematic Review and Meta-Analysis. Vivo 2019, 33, 1737–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Hao, J.; Li, Y. Individualized luteal phase support after fresh embryo transfer: Unanswered questions, a review. Reprod. Health 2022, 19, 19. [Google Scholar] [CrossRef] [PubMed]
- Bezirtzoglou, E.; Voidarou Ch Papadaki, A.; Tsiotsias, A.; Kotsovolou, O.; Konstandi, M. Hormone therapy alters the composition of the vaginal microflora in ovariectomized rats. Microb. Ecol. 2008, 55, 751–759. [Google Scholar] [CrossRef]
- Hyman, R.W.; Herndon, C.N.; Jiang, H.; Palm, C.; Fukushima, M.; Bernstein, D.; Vo, K.C.; Zelenko, Z.; Davis, R.W.; Giudice, L.C. The dynamics of the vaginal microbiome during infertility therapy with in vitro fertilization-embryo transfer. J. Assist. Reprod. Genet. 2012, 29, 105–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carosso, A.; Revelli, A.; Gennarelli, G.; Canosa, S.; Cosma, S.; Borella, F.; Tancredi, A.; Paschero, C.; Boatti, L.; Zanotto, E.; et al. Controlled ovarian stimulation and progesterone supplementation affect vaginal and endometrial microbiota in IVF cycles: A pilot study. J. Assist. Reprod. Genet. 2020, 37, 2315–2326. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Fujishita, A.; Masumoto, H.; Muto, H.; Kitajima, M.; Masuzaki, H.; Kitawaki, J. Molecular detection of intrauterine microbial colonization in women with endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 199, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Chen, H.; Xu, Z.; Wang, B.; Sun, H.; Hu, Y. Pregnancy, Delivery, and Neonatal Outcomes of In Vitro Fertilization-Embryo Transfer in Patient with Previous Cesarean Scar. Med. Sci. Monit. 2016, 22, 3288–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, E.T.; Ramos, L.; Vyas, N.; Bhasin, G.; Simmons, C.F.; Pisarska, M.D. Maternal and neonatal outcomes associated with infertility. J. Matern. Fetal Neonatal Med. 2019, 32, 2820–2823. [Google Scholar] [CrossRef] [PubMed]
- Charitos, I.A.; Topi, S.; Gagliano-Candela, R.; De Nitto, E.; Polimeno, L.; Montagnani, M.; Santacroce, L. The Toxic Effects of Endocrine Disrupting Chemicals (EDCs) on Gut Microbiota: Bisphenol A (BPA) A Review. Endocr. Metab. Immune Disord. Drug Targets 2022, 22, 716–727. [Google Scholar] [CrossRef]
- Yamamoto, T.; Zhou, X.; Williams, C.J.; Hochwalt, A.; Forney, L.J. Bacterial populations in the vaginas of healthy adolescent women. J. Pediatr. Adolesc. Gynecol. 2009, 22, 11–18. [Google Scholar] [CrossRef]
- Eschenbach, D.A.; Davick, P.R.; Williams, B.L.; Klebanoff, S.J.; Young-Smith, K.; Critchlow, C.M.; Holmes, K.K. Prevalence of hydrogen peroxide-producing Lactobacillus species in normal women and women with bacterial vaginosis. J. Clin. Microbiol. 1989, 27, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Hawes, S.E.; Hillier, S.L.; Benedetti, J.; Stevens, C.E.; Koutsky, L.A.; Wolner-Hanssen, P.; Holmes, K.K. Hydrogen peroxide-producing lactobacilli and acquisition of vaginal infections. J. Infect. Dis. 1996, 174, 1058–1063. [Google Scholar] [CrossRef] [Green Version]
- Grönlund, M.M.; Lehtonen, O.P.; Eerola, E.; Kero, P. Fecal microflora in healthy infants born by different methods of delivery: Permanent changes in intestinal flora after cesarean delivery. J. Pediatr. Gastroenterol. Nutr. 1999, 28, 19–25. [Google Scholar] [CrossRef]
- Kataoka, S.; Yamada, T.; Chou, K.; Nishida, R.; Morikawa, M.; Minami, M.; Yamada, H.; Sakuragi, N.; Minakami, H. Association between preterm birth and vaginal colonization by mycoplasmas in early pregnancy. J. Clin. Microbiol. 2006, 44, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Santacroce, L.; Bottalico, L.; Haxhirexha, K.; Topi, S.; Charitos, I.A. Pre-Chemistry Concepts and Medical Therapy among Ancient Physicians through the Pre-Socratic Philosophers. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 1470–1477. [Google Scholar] [CrossRef]
- Ballini, A.; Charitos, I.A.; Cantore, S.; Topi, S.; Bottalico, L.; Santacroce, L. About Functional Foods: The Probiotics and Prebiotics State of Art. Antibiotics 2023, 12, 635. [Google Scholar] [CrossRef] [PubMed]
- Santacroce, L.; Sardaro, N.; Topi, S.; Pettini, F.; Bottalico, L.; Cantore, S.; Cascella, G.; Del Prete, R.; Dipalma, G.; Inchingolo, F. The pivotal role of oral microbiota in health and disease. J. Biol. Regul. Homeost. Agents 2020, 34, 733–737. [Google Scholar] [CrossRef]
- Petricevic, L.; Unger, F.M.; Viernstein, H.; Kiss, H. Randomized, double-blind, placebo-controlled study of oral lactobacilli to improve the vaginal flora of postmenopausal women. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 141, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.; Dols, J.; Miller, W. Targeting the vaginal microbiota with probiotics as a means to counteract infections. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Mack, D.R.; Michail, S.; Wei, S.; McDougall, L.; Hollingsworth, M.A. Probiotics inhibit enteropathogenic E. coli adherence in vitro by inducing intestinal mucin gene expression. Am. J. Physiol. 1999, 276, G941–G950. [Google Scholar]
- Anukam, K.; Osazuwa, E.; Ahonkhai, I.; Ngwu, M.; Osemene, G.; Bruce, A.W.; Reid, G. Augmentation of antimicrobial metronidazole therapy of bacterial vaginosis with oral probiotic Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri RC-14: Randomized, double-blind, placebo-controlled trial. Microbes Infect. 2006, 8, 1450–1454. [Google Scholar] [CrossRef]
- Gargiulo Isacco, C.; Balzanelli, M.G.; Garzone, S.; Lorusso, M.; Inchingolo, F.; Nguyen, K.C.D.; Santacroce, L.; Mosca, A.; Del Prete, R. Alterations of Vaginal Microbiota and Chlamydia trachomatis as Crucial Co-Causative Factors in Cervical Cancer Genesis Procured by HPV. Microorganisms 2023, 11, 662. [Google Scholar] [CrossRef]
- Mastromarino, P.; Vitali, B.; Mosca, L. Bacterial vaginosis: A review on clinical trials with probiotics. New Microbiol. 2013, 36, 229–238. [Google Scholar]
- García-Velasco, J.A.; Menabrito, M.; Catalán, I.B. What fertility specialists should know about the vaginal microbiome: A review. Reprod. Biomed. Online 2017, 35, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, P.; Prabha, V. Evaluation of profertility effect of probiotic Lactobacillus plantarum 2621 in a murine model. Indian J. Med. Res. 2015, 142, 79–84. [Google Scholar] [CrossRef]
- Baud, A.; Hillion, K.H.; Plainvert, C.; Tessier, V.; Tazi, A.; Mandelbrot, L.; Poyart, C.; Kennedy, S.P. Microbial diversity in the vaginal microbiota and its link to pregnancy outcomes. Sci. Rep. 2023, 13, 9061. [Google Scholar] [CrossRef] [PubMed]
- Babu, G.; Singaravelu, B.G.; Srikumar, R.; Reddy, S.V.; Kokan, A. Comparative Study on the Vaginal Flora and Incidence of Asymptomatic Vaginosis among Healthy Women and in Women with Infertility Problems of Reproductive Age. J. Clin. Diagn. Res. 2017, 11, DC18–DC22. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbiology Laboratory Diagnostic Tests | ||
|---|---|---|
| Direct Diagnosis | Rapid Diagnosis | Indirect Diagnosis |
Aimed to establish the presence of the pathogen, its identity and its sensitivity to antibiotics, directly in the sample by:
| Useful in severe infections as it allows identification in a short time by:
| The objective is to detect the host’s immune response (antibodies) against the infective agent. |
| Urinary Microbiome and Homeostasis | |||||
|---|---|---|---|---|---|
| It can produce neurotransmitters that interact with the CNS | It can compete with pathogens for common energy sources | It can play a role in the regulation of epithelial molecular biomechanisms and in maintaining the correct structure between its cells | It can produce antimicrobial molecules that kill pathogens | It can strengthen the epithelial defenses and thus the immune system | It can degrade harmful factors (such as some microbial toxins and other). |
| The Microbiota of the Urinary Tract | ||
|---|---|---|
| * Healthy women | ** Healthy men | * Healthy men and women |
|
|
|
| Uropathogenic Microorganisms | ||
|---|---|---|
| Gram (−) | Gram (+) | Fungi |
| Escherichia coli | Enterococcus spp. | Candida spp. |
| Proteus spp. | Staphylococcus saprophyticus | |
| Klebsiella spp. | Staphylococcus aureus | |
| Citrobacter spp. | Streptococcus spp. | |
| Serratia spp. | ||
| Acinetobacter spp. | ||
| Pseudomonas aeruginosa | ||
| Sample Source | Species |
|---|---|
| Voided urine and Transurethral catheter urine sample | Uncultured bacterium BF0002B042, Uncultured Prevotella spp., Uncultured bacterium clone HRX_H16 |
| Frequency of Detected Vaginal Bacteria | |
|---|---|
| Community State Type (CST) | Dominant Bacteria |
| 1 | Lactobacillus crispatus 25% |
| 2 | Lactobacillus gasseri 5% |
| 3 | Lactobacillus iners 35% |
| 4 | Poor in species from Lactobacillaceae family, Gardnerella 30% |
| 5 | Lactobacillus jensenii 5% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colella, M.; Topi, S.; Palmirotta, R.; D’Agostino, D.; Charitos, I.A.; Lovero, R.; Santacroce, L. An Overview of the Microbiota of the Human Urinary Tract in Health and Disease: Current Issues and Perspectives. Life 2023, 13, 1486. https://doi.org/10.3390/life13071486
Colella M, Topi S, Palmirotta R, D’Agostino D, Charitos IA, Lovero R, Santacroce L. An Overview of the Microbiota of the Human Urinary Tract in Health and Disease: Current Issues and Perspectives. Life. 2023; 13(7):1486. https://doi.org/10.3390/life13071486
Chicago/Turabian StyleColella, Marica, Skender Topi, Raffaele Palmirotta, Donato D’Agostino, Ioannis Alexandros Charitos, Roberto Lovero, and Luigi Santacroce. 2023. "An Overview of the Microbiota of the Human Urinary Tract in Health and Disease: Current Issues and Perspectives" Life 13, no. 7: 1486. https://doi.org/10.3390/life13071486