
The Urinary Microbiome and Bladder Cancer

Abstract
:1. Introduction
2. Possible Mechanisms: Urinary Microbiome, Inflammation and Bladder Tumor Microenvironment
3. Literature Review
3.1. Bladder Cancer versus Controls
3.2. Males versus Females with Bladder Cancer
3.3. Bladder Cancer Stage/Severity
3.4. BCG Responsiveness
3.5. Urethral Microbiota
3.6. Bladder Cancer Tissue
4. Discussion
5. Limitations of the Current Literature
6. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Key Statistics for Bladder Cancer. 2022. Available online: https://www.cancer.org/cancer/bladder-cancer/about/key-statistics.html#:~:text=The%20American%20Cancer%20Society’s%20estimates,men%20and%204%2C980%20in%20women) (accessed on 5 December 2022).
- Cambier, S.; Sylvester, R.J.; Collette, L.; Gontero, P.; Brausi, M.A.; van Andel, G.; Kirkels, W.J.; Silva, F.C.; Oosterlinck, W.; Prescott, S.; et al. EORTC Nomograms and Risk Groups for Predicting Recurrence, Progression, and Disease-specific and Overall Survival in Non-Muscle-invasive Stage Ta-T1 Urothelial Bladder Cancer Patients Treated with 1-3 Years of Maintenance Bacillus Calmette-Guerin. Eur. Urol. 2016, 69, 60–69. [Google Scholar] [CrossRef]
- Schrier, B.P.; Hollander, M.P.; van Rhijn, B.W.G.; Kiemeney, L.A.L.M.; Alfred Witjes, J. Prognosis of Muscle-Invasive Bladder Cancer: Difference between Primary and Progressive Tumours and Implications for Therapy. Eur. Urol. 2004, 45, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, A.J.; Brubaker, L. “Sterile Urine” and the Presence of Bacteria. Eur. Urol. 2015, 68, 173–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragón, I.M.; Herrera-Imbroda, B.; Queipo-Ortuño, M.I.; Castillo, E.; Del Moral, J.S.; Gómez-Millán, J.; Yucel, G.; Lara, M.F. The Urinary Tract Microbiome in Health and Disease. Eur. Urol. Focus 2018, 4, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Brubaker, L.; Wolfe, A. The urinary microbiota: A paradigm shift for bladder disorders? Curr. Opin. Obstet. Gynecol. 2016, 28, 407–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackerman, A.L.; Underhill, D.M. The mycobiome of the human urinary tract: Potential roles for fungi in urology. Ann. Transl. Med. 2017, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Pearce, M.M.; Hilt, E.E.; Rosenfeld, A.B.; Zilliox, M.J.; Thomas-White, K.; Fok, C.; Kliethermes, S.; Schreckenberger, P.C.; Brubaker, L.; Gai, X.; et al. The female urinary microbiome: A comparison of women with and without urgency urinary incontinence. mBio 2014, 5, e01283-01214. [Google Scholar] [CrossRef] [Green Version]
- Santiago-Rodriguez, T.M.; Ly, M.; Bonilla, N.; Pride, D.T. The human urine virome in association with urinary tract infections. Front. Microbiol. 2015, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Jones-Freeman, B.; Chonwerawong, M.; Marcelino, V.R.; Deshpande, A.V.; Forster, S.C.; Starkey, M.R. The microbiome and host mucosal interactions in urinary tract diseases. Mucosal Immunol. 2021, 14, 779–792. [Google Scholar] [CrossRef]
- El-Deeb, O.S.; Atef, M.M.; Hafez, Y.M. The interplay between microbiota-dependent metabolite trimethylamine N-oxide, Transforming growth factor beta/SMAD signaling and inflammasome activation in chronic kidney disease patients: A new mechanistic perspective. J. Cell. Biochem. 2019, 120, 14476–14485. [Google Scholar] [CrossRef] [PubMed]
- Fouts, D.E.; Pieper, R.; Szpakowski, S.; Pohl, H.; Knoblach, S.; Suh, M.J.; Huang, S.T.; Ljungberg, I.; Sprague, B.M.; Lucas, S.K.; et al. Integrated next-generation sequencing of 16S rDNA and metaproteomics differentiate the healthy urine microbiome from asymptomatic bacteriuria in neuropathic bladder associated with spinal cord injury. J. Transl. Med. 2012, 10, 174. [Google Scholar] [CrossRef] [Green Version]
- Abernethy, M.G.; Tsuei, A. The bladder microbiome and interstitial cystitis: Is there a connection? Curr. Opin. Obstet. Gynecol. 2021, 33, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Pederzoli, F.; Ferrarese, R.; Amato, V.; Locatelli, I.; Alchera, E.; Luciano, R.; Nebuloni, M.; Briganti, A.; Gallina, A.; Colombo, R.; et al. Sex-specific Alterations in the Urinary and Tissue Microbiome in Therapy-naive Urothelial Bladder Cancer Patients. Eur. Urol. Oncol. 2020, 3, 784–788. [Google Scholar] [CrossRef]
- Peek, R.M., Jr.; Blaser, M.J. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2002, 2, 28–37. [Google Scholar] [CrossRef]
- Moore, P.S.; Chang, Y. Why do viruses cause cancer? Highlights of the first century of human tumour virology. Nat. Rev. Cancer 2010, 10, 878–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, M.; Buc, E.; Sauvanet, P.; Darcha, C.; Dubois, D.; Pereira, B.; Déchelotte, P.; Bonnet, R.; Pezet, D.; Darfeuille-Michaud, A. Colonization of the human gut by E. coli and colorectal cancer risk. Clin. Cancer Res. 2014, 20, 859–867. [Google Scholar] [CrossRef] [Green Version]
- Oliva, M.; Mulet-Margalef, N.; Ochoa-De-Olza, M.; Napoli, S.; Mas, J.; Laquente, B.; Alemany, L.; Duell, E.J.; Nuciforo, P.; Moreno, V. Tumor-Associated Microbiome: Where Do We Stand? Int. J. Mol. Sci. 2021, 22, 1446. [Google Scholar] [CrossRef]
- Hussein, A.A.; Smith, G.; Guru, K.A. The Association Between the Urinary Microbiome and Bladder Cancer. Urol. Clin. N. Am. 2023, 50, 81–89. [Google Scholar] [CrossRef]
- Gakis, G. The role of inflammation in bladder cancer. Adv. Exp. Med. Biol. 2014, 816, 183–196. [Google Scholar] [CrossRef]
- Sadrekarimi, H.; Gardanova, Z.R.; Bakhshesh, M.; Ebrahimzadeh, F.; Yaseri, A.F.; Thangavelu, L.; Hasanpoor, Z.; Zadeh, F.A.; Kahrizi, M.S. Emerging role of human microbiome in cancer development and response to therapy: Special focus on intestinal microflora. J. Transl. Med. 2022, 20, 301. [Google Scholar] [CrossRef] [PubMed]
- Nadler, N.; Kvich, L.; Bjarnsholt, T.; Jensen, J.B.; Gögenur, I.; Azawi, N. The discovery of bacterial biofilm in patients with muscle invasive bladder cancer. APMIS 2021, 129, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- El-Mosalamy, H.; Salman, T.M.; Ashmawey, A.M.; Osama, N. Role of chronic E. coli infection in the process of bladder cancer- an experimental study. Infect. Agents Cancer 2012, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Wang, J.; Zhou, K.; Lv, J.; Wang, L.; Gao, S.; Keller, E.T.; Zhang, Z.S.; Wang, Q.; Yao, Z. Cytotoxic necrotizing factor 1 promotes bladder cancer angiogenesis through activating RhoC. FASEB J. 2020, 34, 7927–7940. [Google Scholar] [CrossRef] [Green Version]
- Sears, C.L.; Garrett, W.S. Microbes, microbiota, and colon cancer. Cell Host Microbe 2014, 15, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Cosseau, C.; Devine, D.A.; Dullaghan, E.; Gardy, J.L.; Chikatamarla, A.; Gellatly, S.; Yu, L.L.; Pistolic, J.; Falsafi, R.; Tagg, J.; et al. The commensal Streptococcus salivarius K12 downregulates the innate immune responses of human epithelial cells and promotes host-microbe homeostasis. Infect. Immun. 2008, 76, 4163–4175. [Google Scholar] [CrossRef] [Green Version]
- Larsen, E.S.; Joensen, U.N.; Poulsen, A.M.; Goletti, D.; Johansen, I.S. Bacillus Calmette-Guerin immunotherapy for bladder cancer: A review of immunological aspects, clinical effects and BCG infections. APMIS 2020, 128, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Hussein, A.A.; Elsayed, A.S.; Durrani, M.; Jing, Z.; Iqbal, U.; Gomez, E.C.; Singh, P.K.; Liu, S.; Smith, G.; Tang, L.; et al. Investigating the association between the urinary microbiome and bladder cancer: An exploratory study. Urol. Oncol. 2021, 39, 370.e9–370.e19. [Google Scholar] [CrossRef]
- Oresta, B.; Braga, D.; Lazzeri, M.; Frego, N.; Saita, A.; Faccani, C.; Fasulo, V.; Colombo, P.; Guazzoni, G.; Hurle, R.; et al. The Microbiome of Catheter Collected Urine in Males with Bladder Cancer According to Disease Stage. J. Urol. 2021, 205, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Moynihan, M.; Sullivan, T.; Provenzano, K.; Rieger-Christ, K. Urinary Microbiome Evaluation in Patients Presenting with Hematuria with a Focus on Exposure to Tobacco Smoke. Res. Rep. Urol. 2019, 11, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Bucevic Popovic, V.; Situm, M.; Chow, C.T.; Chan, L.S.; Roje, B.; Terzic, J. The urinary microbiome associated with bladder cancer. Sci. Rep. 2018, 8, 12157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, H.; Tian, Y.; Song, C.; Li, J.; Liu, T.; Chen, Z.; Chen, C.; Huang, Y.; Zhang, Y. Urinary microbiota—A potential biomarker and therapeutic target for bladder cancer. J. Med. Microbiol. 2019, 68, 1471–1478. [Google Scholar] [CrossRef]
- Liu, F.; Liu, A.; Lu, X.; Zhang, Z.; Xue, Y.; Xu, J.; Zeng, S.; Xiong, Q.; Tan, H.; He, X.; et al. Dysbiosis signatures of the microbial profile in tissue from bladder cancer. Cancer Med. 2019, 8, 6904–6914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.; Zhang, G.; Zhao, J.; Chen, J.; Chen, Y.; Huang, W.; Zhong, J.; Zeng, J. Profiling the Urinary Microbiota in Male Patients with Bladder Cancer in China. Front. Cell. Infect. Microbiol. 2018, 8, 167. [Google Scholar] [CrossRef]
- Mai, G.; Chen, L.; Li, R.; Liu, Q.; Zhang, H.; Ma, Y. Common Core Bacterial Biomarkers of Bladder Cancer Based on Multiple Datasets. BioMed Res. Int. 2019, 2019, 4824909. [Google Scholar] [CrossRef] [Green Version]
- Pederzoli, F.; Bandini, M.; Raggi, D.; Marandino, L.; Basile, G.; Alfano, M.; Colombo, R.; Salonia, A.; Briganti, A.; Gallina, A.; et al. Is There a Detrimental Effect of Antibiotic Therapy in Patients with Muscle-invasive Bladder Cancer Treated with Neoadjuvant Pembrolizumab? Eur. Urol. 2021, 80, 319–322. [Google Scholar] [CrossRef]
- Xu, W.; Yang, L.; Lee, P.; Huang, W.C.; Nossa, C.; Ma, Y.; Deng, F.M.; Zhou, M.; Melamed, J.; Pei, Z. Mini-review: Perspective of the microbiome in the pathogenesis of urothelial carcinoma. Am. J. Clin. Exp. Urol. 2014, 2, 57–61. [Google Scholar]
- Zeng, J.; Zhang, G.; Chen, C.; Li, K.; Wen, Y.; Zhao, J.; Wu, P. Alterations in Urobiome in Patients With Bladder Cancer and Implications for Clinical Outcome: A Single-Institution Study. Front. Cell. Infect. Microbiol. 2020, 10, 555508. [Google Scholar] [CrossRef]
- Mansour, B.; Monyók, Á.; Makra, N.; Gajdács, M.; Vadnay, I.; Ligeti, B.; Juhász, J.; Szabó, D.; Ostorházi, E. Bladder cancer-related microbiota: Examining differences in urine and tissue samples. Sci. Rep. 2020, 10, 11042. [Google Scholar] [CrossRef] [PubMed]
- Modena, B.D.; Milam, R.; Harrison, F.; Cheeseman, J.A.; Abecassis, M.M.; Friedewald, J.J.; Kirk, A.D.; Salomon, D.R. Changes in Urinary Microbiome Populations Correlate in Kidney Transplants With Interstitial Fibrosis and Tubular Atrophy Documented in Early Surveillance Biopsies. Am. J. Transplant. 2017, 17, 712–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, T.K.; Hilt, E.E.; Thomas-White, K.; Mueller, E.R.; Wolfe, A.J.; Brubaker, L. The urobiome of continent adult women: A cross-sectional study. BJOG 2020, 127, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Hourigan, S.K.; Zhu, W.; Wong, W.S.W.; Clemency, N.C.; Provenzano, M.; Vilboux, T.; Niederhuber, J.E.; Deeken, J.; Chung, S.; McDaniel-Wiley, K.; et al. Studying the urine microbiome in superficial bladder cancer: Samples obtained by midstream voiding versus cystoscopy. BMC Urol. 2020, 20, 5. [Google Scholar] [CrossRef]
- Ishaq, A.; Zahid, S.; Mukhtar, S.; Azim, M.K.; Akhtar, S.; Irfan, M.; Malik, K.A. Comparative Analysis of Microbiome of Bladder Cancer Patients. 2020. [Google Scholar] [CrossRef]
- Chipollini, J.; Wright, J.R.; Nwanosike, H.; Kepler, C.Y.; Batai, K.; Lee, B.R.; Spiess, P.E.; Stewart, D.B.; Lamendella, R. Characterization of urinary microbiome in patients with bladder cancer: Results from a single-institution, feasibility study. Urol. Oncol. Semin. Orig. Investig. 2020, 38, 615–621. [Google Scholar] [CrossRef]
- Garrett, W.S. Cancer and the microbiota. Science 2015, 348, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, S.A.; Razvi, H.; Dave, S.; Reid, G.; Burton, J.P. The microbiome of the urinary tract--a role beyond infection. Nat. Rev. Urol. 2015, 12, 81–90. [Google Scholar] [CrossRef]
- Dejea, C.M.; Wick, E.C.; Hechenbleikner, E.M.; White, J.R.; Mark Welch, J.L.; Rossetti, B.J.; Peterson, S.N.; Snesrud, E.C.; Borisy, G.G.; Lazarev, M.; et al. Microbiota organization is a distinct feature of proximal colorectal cancers. Proc. Natl. Acad. Sci. USA 2014, 111, 18321–18326. [Google Scholar] [CrossRef] [Green Version]
- Kunze, E.; Chang-Claude, J.; Frentzel-Beyme, R. Life style and occupational risk factors for bladder cancer in Germany. A case-control study. Cancer 1992, 69, 1776–1790. [Google Scholar] [CrossRef]
- Jiang, X.; Castelao, J.E.; Groshen, S.; Cortessis, V.K.; Shibata, D.; Conti, D.V.; Yuan, J.M.; Pike, M.C.; Gago-Dominguez, M. Urinary tract infections and reduced risk of bladder cancer in Los Angeles. Br. J. Cancer 2009, 100, 834–839. [Google Scholar] [CrossRef] [Green Version]
- Bayne, C.E.; Farah, D.; Herbst, K.W.; Hsieh, M.H. Role of urinary tract infection in bladder cancer: A systematic review and meta-analysis. World J. Urol. 2018, 36, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, Y.; Nakai, S.; Tsukamoto, T.; Masumori, N.; Akaza, H.; Miyanaga, N.; Kitamura, T.; Kawabe, K.; Kotake, T.; Kuroda, M.; et al. Habitual intake of lactic acid bacteria and risk reduction of bladder cancer. Urol. Int. 2002, 68, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Hara, T.; Nagaoka, M.; Mike, A.; Mitsuyama, K.; Sako, T.; Yamamoto, M.; Kado, S.; Takada, T. A component of polysaccharide peptidoglycan complex on Lactobacillus induced an improvement of murine model of inflammatory bowel disease and colitis-associated cancer. Immunology 2009, 128, e170–e180. [Google Scholar] [CrossRef]
- Zhang, K.; Dai, H.; Liang, W.; Zhang, L.; Deng, Z. Fermented dairy foods intake and risk of cancer. Int. J. Cancer 2019, 144, 2099–2108. [Google Scholar] [CrossRef] [PubMed]
- Hoesl, C.E.; Altwein, J.E. The probiotic approach: An alternative treatment option in urology. Eur. Urol. 2005, 47, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Akazan, H. Prophylactic effect of a Lactobacillus casei preparation on the recurrence of superficial bladder cancer. BLP Study Group. Urol. Int. 1992, 49, 125–129. [Google Scholar] [CrossRef]
- Ingersoll, M.A.; Albert, M.L. From infection to immunotherapy: Host immune responses to bacteria at the bladder mucosa. Mucosal Immunol. 2013, 6, 1041–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redelman-Sidi, G.; Glickman, M.S.; Bochner, B.H. The mechanism of action of BCG therapy for bladder cancer—A current perspective. Nat. Rev. Urol. 2014, 11, 153–162. [Google Scholar] [CrossRef]
- Mortimer, T.D.; Annis, D.S.; O’Neill, M.B.; Bohr, L.L.; Smith, T.M.; Poinar, H.N.; Mosher, D.F.; Pepperell, C.S. Adaptation in a Fibronectin Binding Autolysin of Staphylococcus saprophyticus. mSphere 2017, 2, e00511–e00517. [Google Scholar] [CrossRef] [Green Version]
- Shah, G.; Zhang, G.; Chen, F.; Cao, Y.; Kalyanaraman, B.; See, W.A. iNOS expression and NO production contribute to the direct effects of BCG on urothelial carcinoma cell biology. Urol. Oncol. 2014, 32, 45.e1–45.e9. [Google Scholar] [CrossRef] [Green Version]
- Bajic, P.; Wolfe, A.J.; Gupta, G.N. Old instillations and new implications for bladder cancer: The urinary microbiome and intravesical BCG. BJU Int. 2019, 124, 7–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seow, S.W.; Rahmat, J.N.; Mohamed, A.A.; Mahendran, R.; Lee, Y.K.; Bay, B.H. Lactobacillus species is more cytotoxic to human bladder cancer cells than Mycobacterium Bovis (bacillus Calmette-Guerin). J. Urol. 2002, 168, 2236–2239. [Google Scholar] [CrossRef] [PubMed]
- McMillan, A.; Macklaim, J.M.; Burton, J.P.; Reid, G. Adhesion of Lactobacillus iners AB-1 to human fibronectin: A key mediator for persistence in the vagina? Reprod. Sci. 2013, 20, 791–796. [Google Scholar] [CrossRef]
- Role of Microbiome in BCG Responsiveness Prediction (SILENTEMPIRE). 2023. Available online: https://clinicaltrials.gov/ct2/show/NCT05204199 (accessed on 12 January 2023).
- Sweis, R.F.; Golan, S.; Barashi, N.; Hill, E.; Andolfi, C.; Werntz, R.P.; Bloodworth, J.; Steinberg, G.D. Association of the commensal urinary microbiome with response to Bacillus Calmette-Guérin (BCG) immunotherapy in nonmuscle invasive bladder cancer. J. Clin. Oncol. 2019, 37, 423. [Google Scholar] [CrossRef]
- Larsson, S.C.; Andersson, S.O.; Johansson, J.E.; Wolk, A. Cultured milk, yogurt, and dairy intake in relation to bladder cancer risk in a prospective study of Swedish women and men. Am. J. Clin. Nutr. 2008, 88, 1083–1087. [Google Scholar] [CrossRef] [Green Version]
- Curtiss, N.; Balachandran, A.; Krska, L.; Peppiatt-Wildman, C.; Wildman, S.; Duckett, J. Age, menopausal status and the bladder microbiome. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 228, 126–129. [Google Scholar] [CrossRef]
- Luthje, P.; Hirschberg, A.L.; Brauner, A. Estrogenic action on innate defense mechanisms in the urinary tract. Maturitas 2014, 77, 32–36. [Google Scholar] [CrossRef]
- Lewis, D.A.; Brown, R.; Williams, J.; White, P.; Jacobson, S.K.; Marchesi, J.R.; Drake, M.J. The human urinary microbiome; bacterial DNA in voided urine of asymptomatic adults. Front. Cell. Infect. Microbiol. 2013, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Eto, D.S.; Sundsbak, J.L.; Mulvey, M.A. Actin-gated intracellular growth and resurgence of uropathogenic Escherichia coli. Cell. Microbiol. 2006, 8, 704–717. [Google Scholar] [CrossRef]
- Dizman, N.; Meza, L.; Bergerot, P.; Alcantara, M.; Dorff, T.; Lyou, Y.; Frankel, P.; Cui, Y.; Mira, V.; Llamas, M.; et al. Nivolumab plus ipilimumab with or without live bacterial supplementation in metastatic renal cell carcinoma: A randomized phase 1 trial. Nat. Med. 2022, 28, 704–712. [Google Scholar] [CrossRef]


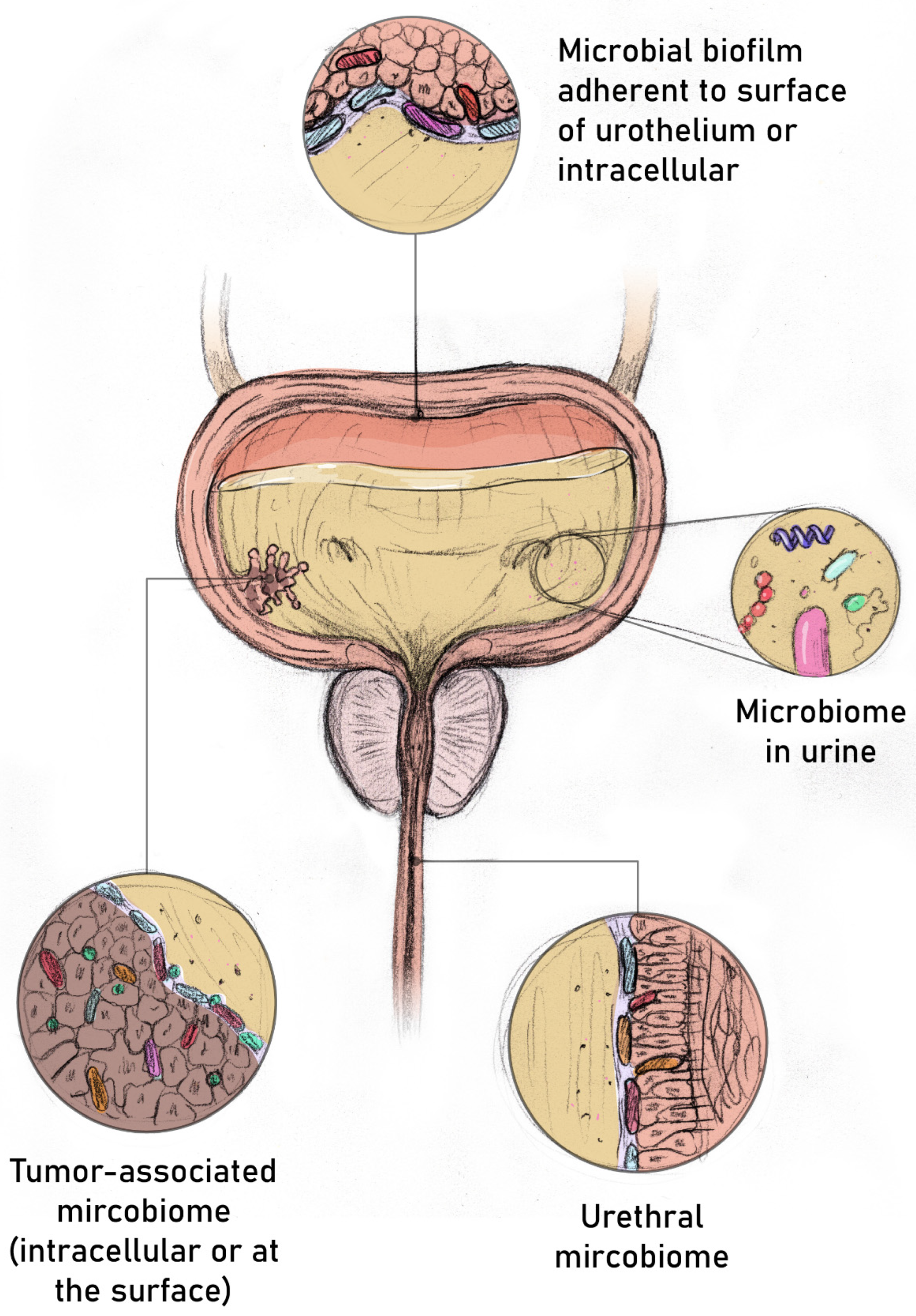{kind=link}
| Study, Year | BCa, n | Non-BCa, n | Females n (%) | Sample | Alpha-Diversity | Beta-Diversity | Relative Abundance, Phyla | Relative Abundance, Genera | Other |
|---|---|---|---|---|---|---|---|---|---|
| Xu et al., 2014 [40] | 8 | 6 | NA | Urine—clean catch | Not reported | Not reported | BCa: Streptococcus Non-BCa: Streptococcus abundance near 0 | N/A | -- |
| Wu et al., 2018 [37] | 31 | 18 | 0 | Urine—clean catch | Different | Different | BCa: Proteobacteria Firmicutes Actinobacteria Non-BCa: Proteobacteria Firmicutes Bacteroidetes | BCa: Escherichia-Shigella Staphylococcus Streptococcus Aeromonas Non-BCa: Escherichia-Shigella Staphylococcus Streptococcus Lactobacillus | -- |
| Bucevic Popovic et al., 2018 [34] | 12 | 11 | Urine—clean catch | NS | NS | Firmicutes, Actinobacteria, Bacteroidetes and Proteobacteria | Streptococcus, Prevotella, Peptoniphilus, Campylobacter, Veillonella, Anaerococcus, Finegoldia | ||
| Mai et al., 2019 [38] | 24 | 0 | 6 (25%) | Urine—clean catch | Not reported | Not reported | The five most abundant phyla are Proteobacteria, Firmicutes, Actinobacteria, Tenericutes, and Bacteroidetes | There are 31 bacterial genera (Core31) in all of these 24 samples, including Clostridiales_f_g, Peptoniphilus, Mycoplasma, Cupriavidus, Lachnospiraceae, Ureaplasma, Delftia, o_Rhizobiales_f_g, Acinetobacter, Enterococcus, Hydrogenophilus, Prevotella, Bacillus | |
| Bi et al., 2019 [35] | 29 | 26 | 20 (36%) | Urine—clean catch | different | different | Tenericutes and Proteobacteria was higher in the cancer group versus the control group | Non-BCa: Streptococcus, Bifidobacterium, Lactobacillus and Veillonella BCa: Actinomyces | |
| Moynihan et al., 2019 [33] | 8 | 33 | 0 | Mid-stream urine | NS | NS | Firmicutes, Proteobacteria, and Bacteroides | Turicibacter, Lactobacillus, and Bacteroides | |
| Hourigan et al., 2020 [45] | 22 | 0 | 8 | Cystoscopy and mid-stream urine | NS | NS | Firmicutes (increased in voided samples) and Proteobacteria (increased in males) | Sternotrophomonas (increased in cystoscopy samples) and tepidimonas (increased in males) | |
| Ishaq et al., 2020 [46] | 10 | 0 | 3 | Midstream urine and tissue | NS | NS | Enterobacteriaceae, Bacillus, Meiothermus and Methylotenera | ||
| Chipollini et al., 2020 [47] | 27 | 10 | Mid-stream urine | Different | Different | NA | NA | ||
| Mansour et al., 2020 [42] | 10 | 0 | 5 | Mid-stream urine and tissue | NS | NS | In urine samples, the most abundant phyla detected were Firmicutes, Proteobacteria, Actinobacteria, Cyanobacteria, and Bacteroidetes. In the tissue samples, Firmicutes, Actinobacteria, Proteobacteria, Bacteroidetes, Cyanobacteria | urine samples: Lactobacillus, Corynebacterium, Streptococcus and Staphylococcus. Tissue samples: Bacteroides, Akkermansia, Klebsiella and Clostridium | Akkermansia, Bacteroides, Clostridium sensu stricto, Enterobacter and Klebsiella genera showed remarkably higher compositional abundance in tissue than in urine samples |
| Pederzoli et al., 2020 [39] | 49 | 59 | 38 | Mid-stream urine Tissue | NS | In urine: Different In tissue: NS | Proteobacteria, Firmicutes, and Bacteroidetes | Sought to find shared BCa microbiome between urine and bladder tissue | |
| Zeng et al., 2020 [41] | 62 | 19 | 0 | Mid-strean urine | Different | Different in males only | Firmicutes, Proteobacteria, and Actinobacteria | Staphylococcus, Streptococcus, Prevotella, and Corynebacterium | |
| Hussein et al., 2021 [31] | 43 | 10 | 12 | Mid-stream urine (healthy) Cath/cystoscopy from cancer patients | NS | Different | BCa: Actinobacteria, Proteobacteria Non- BCa: Firmicutes, Deinococcus-Thermus | BCa: Actinomyces, Achromobacter, Brevibacterium, and Brucella Non-BCa: Salinococcus, Jeotgalicoccus, Escherichia-Shigella, Faecalibacterium, Thermus, and Lactobacillus | MIBC vs. NMIBC BCG responders vs. non responders Male vs. females with BCa |
| Oresta et al., 2021 [32] | 51 | 10 | 0 | Mid-stream urine and catheterized urine | Different | NS | Firmicutes, Actinobacteria, Bacteroidetes and Pro-teobacteria | Veillonella and Corynebacterium were enriched in the BC group | Veillonella was increased in pTa/T1 HG, CIS and T2 tumors compared to controls and pTa LG tumors; Corynebacterium and Staphylococcus were specifically enriched in HG NMIBC and pTa LG tumors, respectively |
| Study, Year | BCa, n | Non-BCa, n | Females n (%) | Sample | Alpha-Diversity | Beta-Diversity | Relative Abundance, Phyla | Relative Abundance, Genera | Other |
|---|---|---|---|---|---|---|---|---|---|
| Liu et al., 2019 [36] | 22 | 12 | 0 | BCa and non-BCa tissue | NS (except for Shannon Index) | Different | BCa: Proteobacteria, Actinobacteria, Cyanobacteria, Chloroflexi, Deinococcus-Thermus, Armatimonadetes Non- BCa: Firmicutes, Bacteroidetes | BCa: Cupriavidus, Uncl Brucellaceae, Acinebacter, Escherichia-Shigella, Sphingomonas, Pelomonas, Ralstonia, Anoxybacillus Non-BCa: lactobacillus, Prevotella, Ruminococcaceae | High Grade vs. Low Grade High risk for recurrence and progression |
| Ishaq et al., 2020 [46] | 10 | 0 | 3 | Midstream urine and tissue | NS | NS | Enterobacteriaceae, Bacillus, Meiothermus and Methylotenera | ||
| Mansour et al., 2020 [42] | 10 | 0 | 5 | Mid-stream urine and tissue | NS | NS | In urine samples, the most abundant phyla detected were Firmicutes, Proteobacteria, Actinobacteria, Cyanobacteria, and Bacteroidetes. In the tissue samples, Firmicutes, Actinobacteria, Proteobacteria, Bacteroidetes, Cyanobacteria | urine samples: Lactobacillus, Corynebacterium, Streptococcus and Staphylococcus. Tissue samples: Bacteroides, Akkermansia, Klebsiella and Clostridium | Akkermansia, Bacteroides, Clostridium sensu stricto, Enterobacter and Klebsiella genera showed remarkably higher compositional abundance in tissue than in urine samples |
| Pederzoli et al., 2020 [39] | 49 | 59 | 38 | Mid-stream urine Tissue | NS | In urine: Different In tissue: NS | Proteobacteria, Firmicutes, and Bacteroidetes | Sought to find shared BCa microbiome between urine and bladder tissue |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heidar, N.A.; Bhat, T.A.; Shabir, U.; Hussein, A.A. The Urinary Microbiome and Bladder Cancer. Life 2023, 13, 812. https://doi.org/10.3390/life13030812
Heidar NA, Bhat TA, Shabir U, Hussein AA. The Urinary Microbiome and Bladder Cancer. Life. 2023; 13(3):812. https://doi.org/10.3390/life13030812
Chicago/Turabian StyleHeidar, Nassib Abou, Tariq A. Bhat, Usma Shabir, and Ahmed A. Hussein. 2023. "The Urinary Microbiome and Bladder Cancer" Life 13, no. 3: 812. https://doi.org/10.3390/life13030812