
Sphingosine-1-phosphate Treatment Improves Cryopreservation Efficiency in Human Mesenchymal Stem Cells
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of DP-MSCs
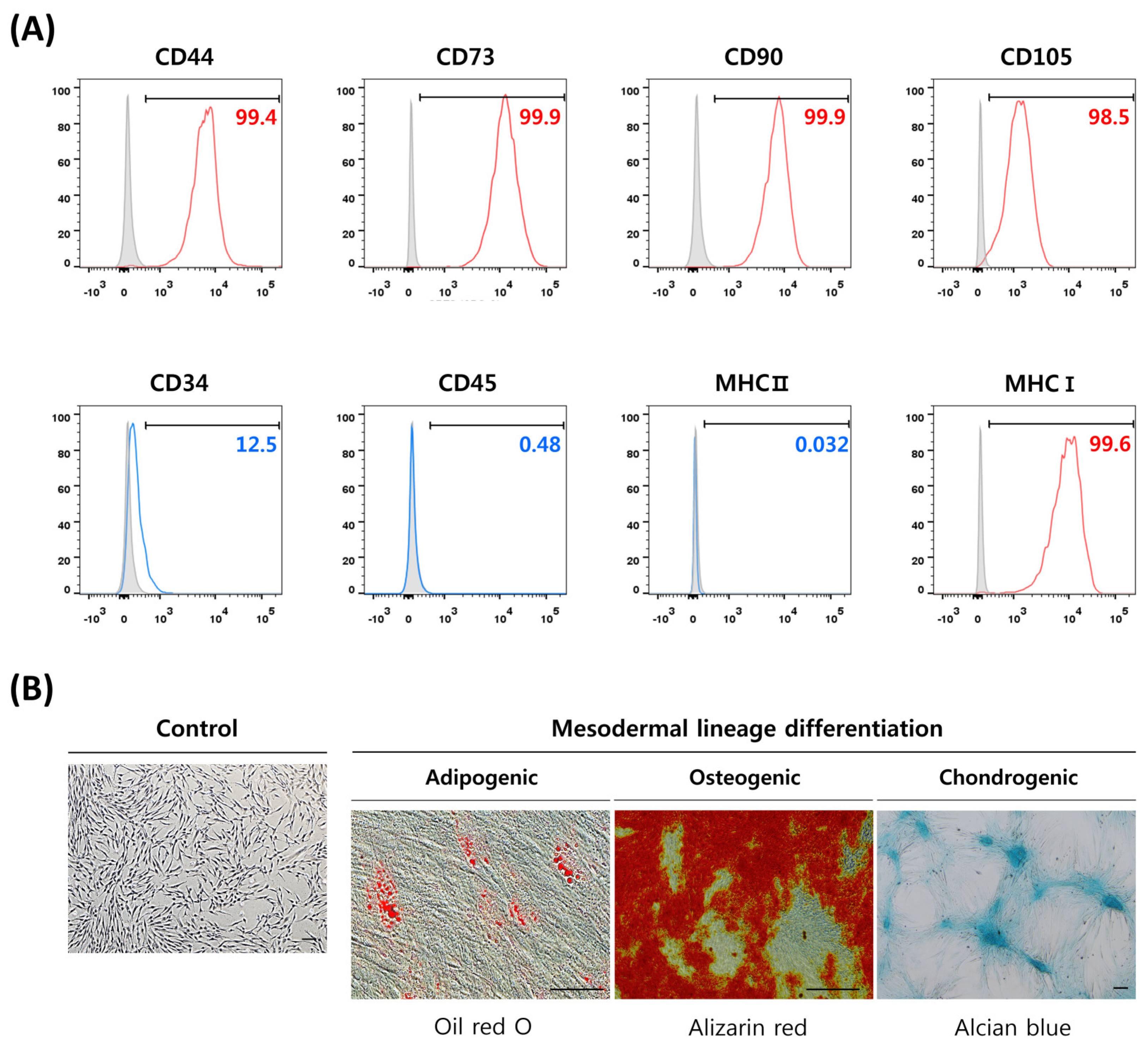
2.2. Characterization of DP-MSCs
2.3. qRT-PCR
2.4. Cryopreservation and Thawing of MSCs
2.5. Cell Viability and Proliferation Analysis
2.6. F-Actin Staining
2.7. Cell Adhesion Assay
2.8. Statistical Analysis
3. Results
3.1. Characterization of DP-MSCs by Surface Markers Expression and Differentiation Potential
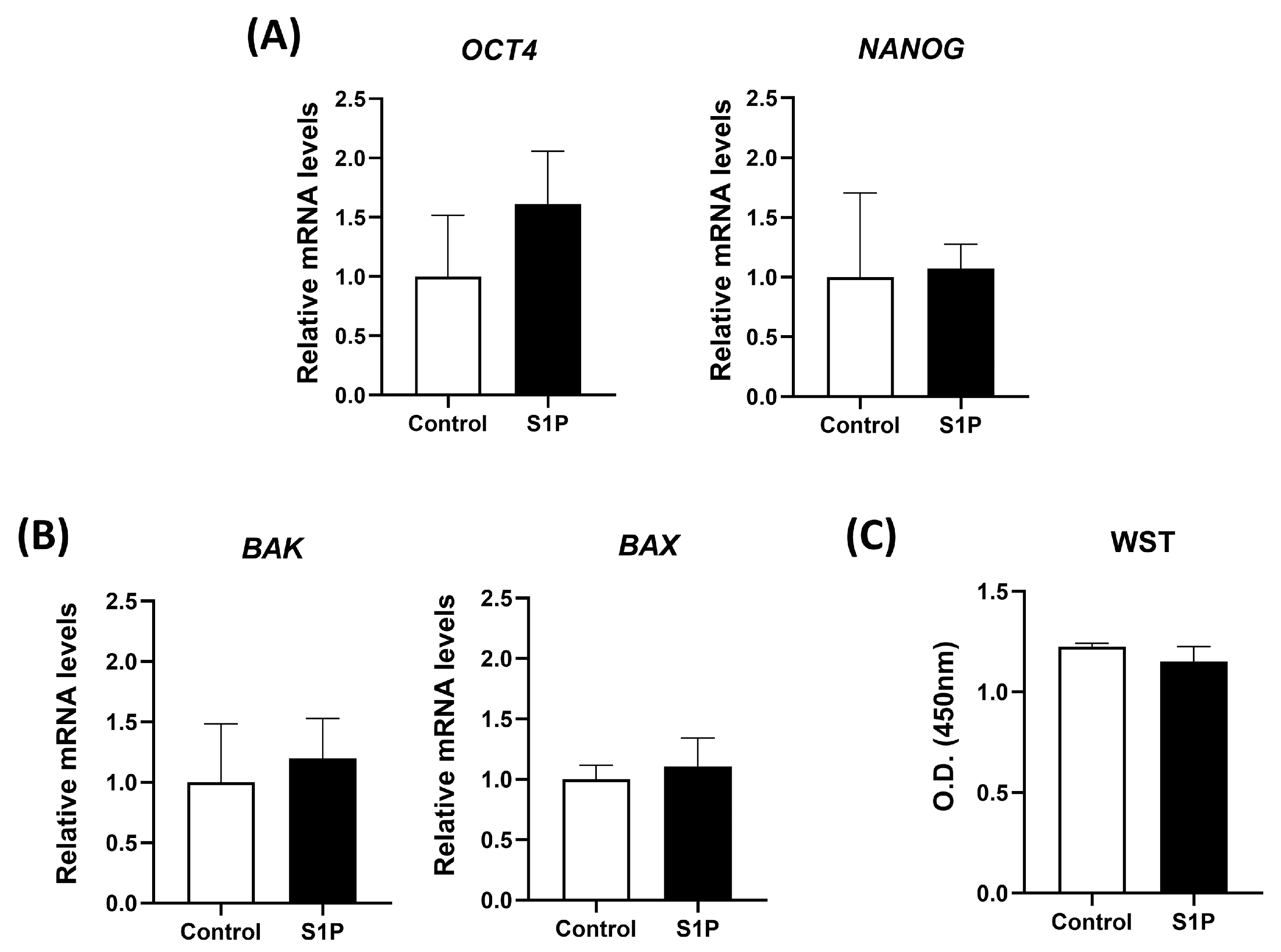
3.2. S1P Treatment Does Not Induce a Negative Impact on the Viability and Stemness of DP-MSCs
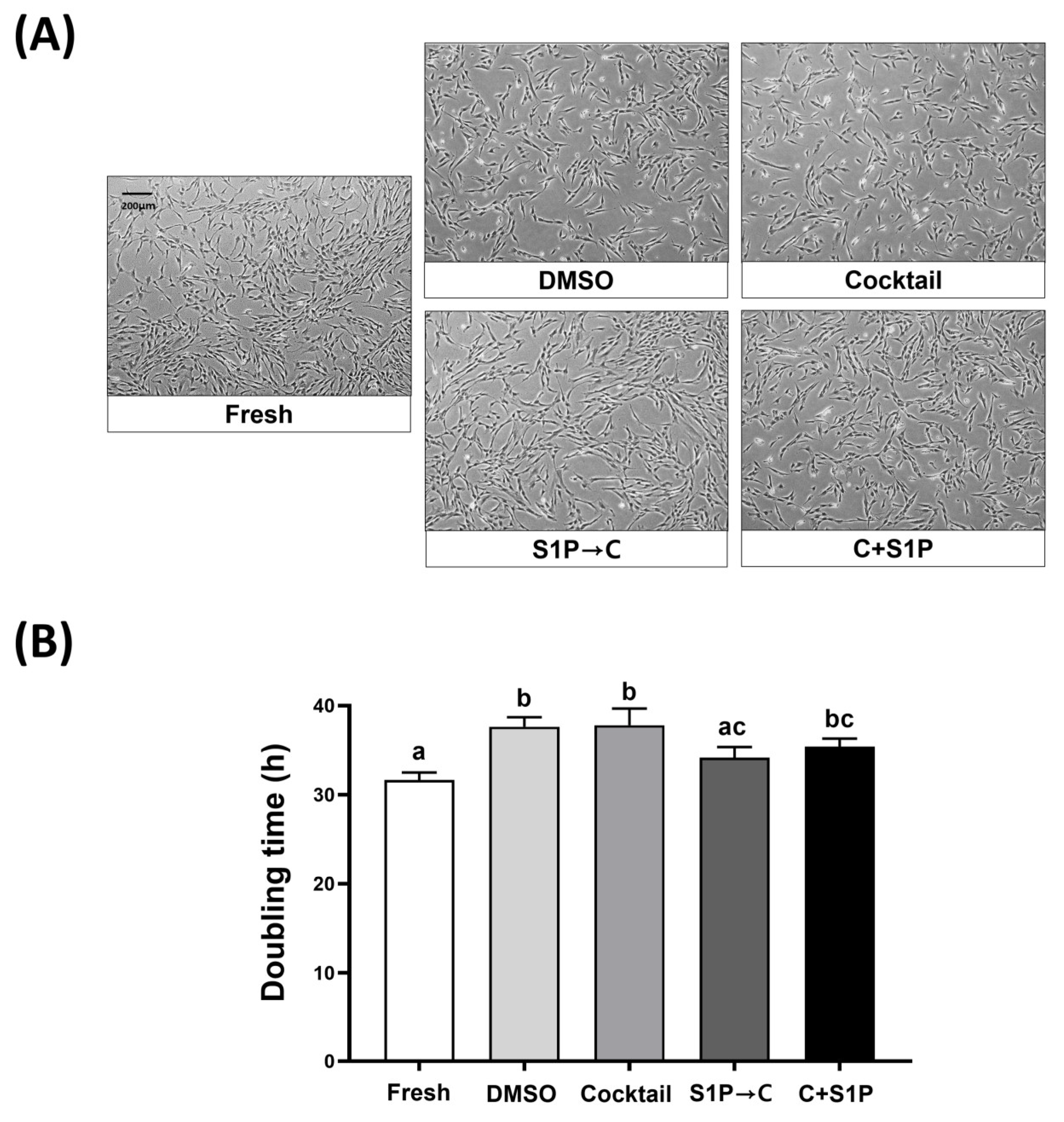
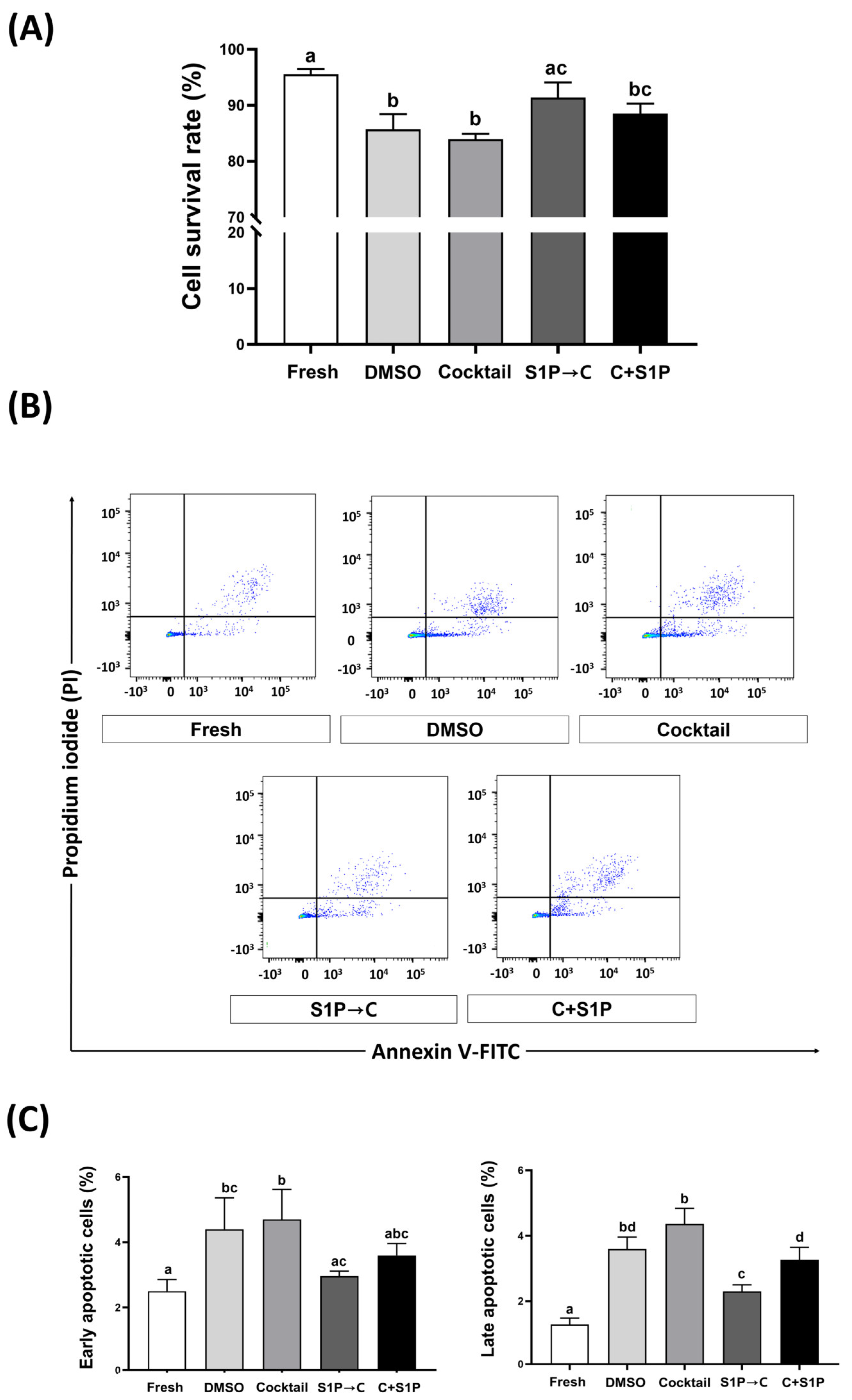
3.3. S1P Pretreatment Enhanced the Cell Viability of Post-Freeze/Thaw DP-MSCs
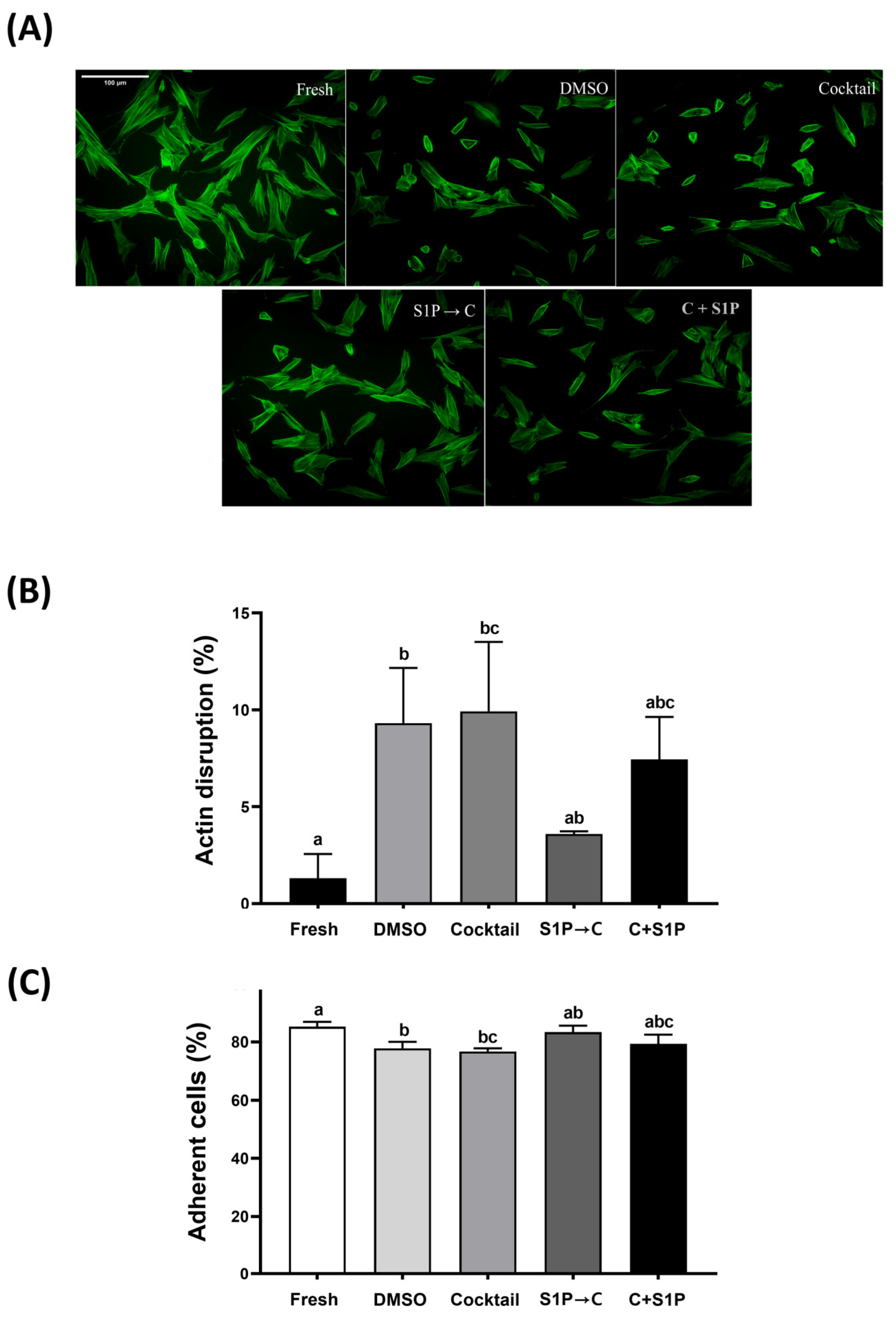
3.4. S1P Pre-Treatment Reduced Actin Cytoskeletal Disruption and Improved Adhesion Ability
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arthur, A.; Zannettino, A.; Gronthos, S. The Therapeutic Applications of Multipotential Mesenchymal/Stromal Stem Cells in Skeletal Tissue Repair. J. Cell. Physiol. 2009, 218, 237–245. [Google Scholar] [CrossRef]
- de Castro, L.L.; Lopes-Pacheco, M.; Weiss, D.J.; Cruz, F.F.; Rocco, P.R.M. Current Understanding of the Immunosuppressive Properties of Mesenchymal Stromal Cells. J. Mol. Med. 2019, 97, 605–618. [Google Scholar] [CrossRef]
- Casagrande, L.; Cordeiro, M.M.; Nör, S.A.; Nör, J.E. Dental Pulp Stem Cells in Regenerative Dentistry. Odontology 2011, 99, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ledesma-Martínez, E.; Mendoza-Núñez, V.M.; Santiago-Osorio, E. Mesenchymal Stem Cells Derived from Dental Pulp: A Review. Stem Cells Int. 2016, 2016, 4709572. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, T.W. Dental Pulp Stem Cells: Advances to Applications. Stem Cells Cloning 2020, 13, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Pacini, S. Deterministic and Stochastic Approaches in the Clinical Application of Mesenchymal Stromal Cells (MSCs). Front. Cell Dev. Biol. 2014, 2, 50. [Google Scholar] [CrossRef] [PubMed]
- Marquez-Curtis, L.A.; Janowska-Wieczorek, A.; McGann, L.E.; Elliott, J.A.W. Mesenchymal Stromal Cells Derived from Various Tissues: Biological, Clinical and Cryopreservation Aspects. Cryobiology 2015, 71, 181–197. [Google Scholar] [CrossRef]
- Meryman, H.T. Cryoprotective Agents. Cryobiology 1971, 8, 173–183. [Google Scholar] [CrossRef]
- Patel, M.; Park, J.K.; Jeong, B. Rediscovery of poly(ethylene glycol)s as a cryoprotectant for mesenchymal stem cells. Biomater Res. 2023, 27, 17. [Google Scholar] [CrossRef]
- Haack-Sørensen, M.; Kastrup, J. Cryopreservation and Revival of Mesenchymal Stromal Cells. Methods Mol. Biol. 2011, 698, 161–174. [Google Scholar] [CrossRef]
- Davies, O.G.; Smith, A.J.; Cooper, P.R.; Shelton, R.M.; Scheven, B.A. The Effects of Cryopreservation on Cells Isolated from Adipose, Bone Marrow and Dental Pulp Tissues. Cryobiology 2014, 69, 342–347. [Google Scholar] [CrossRef]
- Hunt, C.J. Cryopreservation of Human Stem Cells for Clinical Application: A Review. Transfus. Med. Hemother. 2011, 38, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Jang, T.H.; Park, S.C.; Yang, J.H.; Kim, J.Y.; Seok, J.H.; Park, U.S.; Choi, C.W.; Lee, S.R.; Han, J. Cryopreservation and Its Clinical Applications. Integr. Med. Res. 2017, 6, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Yong, K.W.; Wan Safwani, W.K.Z.; Xu, F.; Wan Abas, W.A.B.; Choi, J.R.; Pingguan-Murphy, B. Cryopreservation of Human Mesenchymal Stem Cells for Clinical Applications: Current Methods and Challenges. Biopreserv. Biobank. 2015, 13, 231–239. [Google Scholar] [CrossRef]
- Karlsson, J.O.; Toner, M. Long-Term Storage of Tissues by Cryopreservation: Critical Issues. Biomaterials 1996, 17, 243–256. [Google Scholar] [CrossRef]
- Zambelli, A.; Poggi, G.; Da Prada, G.; Pedrazzoli, P.; Cuomo, A.; Miotti, D.; Perotti, C.; Preti, P.; Robustelli della Cuna, G. Clinical Toxicity of Cryopreserved Circulating Progenitor Cells Infusion. Anticancer Res. 1998, 18, 4705–4708. [Google Scholar] [PubMed]
- Ekpo, M.D.; Boafo, G.F.; Xie, J.; Liu, X.; Chen, C.; Tan, S. Strategies in developing dimethyl sulfoxide (DMSO)-free cryopreservation protocols for biotherapeutics. Front. Immunol. 2022, 13, 1030965. [Google Scholar] [CrossRef] [PubMed]
- Traversari, G.; Delogu, F.; Aparicio, S.; Cincotti, A. hMSCs in contact with DMSO for cryopreservation: Experiments and modeling of osmotic injury and cytotoxic effect. Biotechnol. Bioeng. 2022, 119, 2890–2907. [Google Scholar] [CrossRef]
- Higman, M.A.; Port, J.D.; Beauchamp, N.J.J.; Chen, A.R. Reversible Leukoencephalopathy Associated with Re-Infusion of DMSO Preserved Stem Cells. Bone Marrow Transplant. 2000, 26, 797–800. [Google Scholar] [CrossRef]
- Bruserud, O.; Tronstad, K.J.; Berge, R. In Vitro Culture of Human Osteosarcoma Cell Lines: A Comparison of Functional Characteristics for Cell Lines Cultured in Medium without and with Fetal Calf Serum. J. Cancer Res. Clin. Oncol. 2005, 131, 377–384. [Google Scholar] [CrossRef]
- Lange, C.; Cakiroglu, F.; Spiess, A.-N.; Cappallo-Obermann, H.; Dierlamm, J.; Zander, A.R. Accelerated and Safe Expansion of Human Mesenchymal Stromal Cells in Animal Serum-Free Medium for Transplantation and Regenerative Medicine. J. Cell. Physiol. 2007, 213, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Latinwo, L.M.; Badisa, V.L.D.; Odewumi, C.O.; Ikediobi, C.O.; Badisa, R.B.; Brooks-Walter, A.; Lambert, A.-T.T.; Nwoga, J. Comparative Evaluation of Cytotoxicity of Cadmium in Rat Liver Cells Cultured in Serum-Containing Medium and Commercially Available Serum-Free Medium. Int. J. Mol. Med. 2008, 22, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Grein, T.A.; Freimark, D.; Weber, C.; Hudel, K.; Wallrapp, C.; Czermak, P. Alternatives to Dimethylsulfoxide for Serum-Free Cryopreservation of Human Mesenchymal Stem Cells. Int. J. Artif. Organs 2010, 33, 370–380. [Google Scholar] [CrossRef]
- Petrenko, Y.A.; Rogulska, O.Y.; Mutsenko, V.V.; Petrenko, A.Y. A Sugar Pretreatment as a New Approach to the Me2SO- and Xeno-Free Cryopreservation of Human Mesenchymal Stromal Cells. Cryo Lett. 2014, 35, 239–246. [Google Scholar]
- Shivakumar, S.B.; Bharti, D.; Jang, S.-J.; Hwang, S.-C.; Park, J.-K.; Shin, J.-K.; Byun, J.-H.; Park, B.-W.; Rho, G.-J. Cryopreservation of Human Wharton’s Jelly-Derived Mesenchymal Stem Cells Following Controlled Rate Freezing Protocol Using Different Cryoprotectants: A Comparative Study. Int. J. Stem Cells 2015, 8, 155–169. [Google Scholar] [CrossRef]
- Xu, X.; Liu, Y.; Cui, Z.; Wei, Y.; Zhang, L. Effects of Osmotic and Cold Shock on Adherent Human Mesenchymal Stem Cells during Cryopreservation. J. Biotechnol. 2012, 162, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Ragoonanan, V.; Hubel, A.; Aksan, A. Response of the Cell Membrane-Cytoskeleton Complex to Osmotic and Freeze/Thaw Stresses. Cryobiology 2010, 61, 335–344. [Google Scholar] [CrossRef]
- Ragoonanan, V.; Less, R.; Aksan, A. Response of the Cell Membrane-Cytoskeleton Complex to Osmotic and Freeze/Thaw Stresses. Part 2: The Link between the State of the Membrane-Cytoskeleton Complex and the Cellular Damage. Cryobiology 2013, 66, 96–104. [Google Scholar] [CrossRef]
- Chinnadurai, R.; Garcia, M.A.; Sakurai, Y.; Lam, W.A.; Kirk, A.D.; Galipeau, J.; Copland, I.B. Actin Cytoskeletal Disruption Following Cryopreservation Alters the Biodistribution of Human Mesenchymal Stromal Cells in Vivo. Stem Cell Rep. 2014, 3, 60–72. [Google Scholar] [CrossRef]
- Heng, N.; Zhao, Z.X.; Guo, Y.; Gao, S.; Cai, D.L.; Fu, B.F.; Sheng, X.H.; Wang, X.G.; Xing, K.; Xiao, L.F.; et al. RhoA improves cryopreservation of rooster sperm through the Rho/RhoA-associated kinase/cofilin pathway. Poult. Sci. 2022, 101, 102051. [Google Scholar] [CrossRef]
- Spiegel, S.; Milstien, S. Sphingosine-1-Phosphate: An Enigmatic Signalling Lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Chen, Z.; Zhao, X.; Pan, F.; Cai, M.; Wang, T.; Zhang, H.; Lu, J.R.; Lei, M. Sphingosine-1-Phosphate Promotes the Differentiation of Human Umbilical Cord Mesenchymal Stem Cells into Cardiomyocytes under the Designated Culturing Conditions. J. Biomed. Sci. 2011, 18, 37. [Google Scholar] [CrossRef]
- Messias, C.V.; Santana-Van-Vliet, E.; Lemos, J.P.; Moreira, O.C.; Cotta-de-Almeida, V.; Savino, W.; Mendes-da-Cruz, D.A. Sphingosine-1-Phosphate Induces Dose-Dependent Chemotaxis or Fugetaxis of T-ALL Blasts through S1P1 Activation. PLoS ONE 2016, 11, e0148137. [Google Scholar] [CrossRef] [PubMed]
- Sassoli, C.; Pierucci, F.; Zecchi-Orlandini, S. Sphingosine 1-Phosphate (S1P)/S1P Receptor Signaling and Mechanotransduction: Implications for Intrinsic Tissue Repair/Regeneration. Int. J. Mol. Sci. 2019, 20, 5545. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Zheng, Y.; Hu, M.; Xu, P.; Lin, L.; Liu, X.; Wu, Y.; Huang, B.; Ye, X.; Li, S.; et al. Impaired Sphingosine-1-Phosphate Synthesis Induces Preeclampsia by Deactivating Trophoblastic YAP (Yes-Associated Protein) Through S1PR2 (Sphingosine-1-Phosphate Receptor-2)-Induced Actin Polymerizations. Hypertension 2022, 79, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Duan, M.; Gao, P.; Chen, S.X.; Novák, P.; Yin, K.; Zhu, X. Sphingosine-1-phosphate in mitochondrial function and metabolic diseases. Obes. Rev. 2022, 23, e13426. [Google Scholar] [CrossRef] [PubMed]
- Guzel, Y.; Bildik, G.; Dilege, E.; Oktem, O. Sphingosine-1-Phosphate Reduces Atresia of Primordial Follicles Occurring during Slow-Freezing and Thawing of Human Ovarian Cortical Strips. Mol. Reprod. Dev. 2018, 85, 858–864. [Google Scholar] [CrossRef]
- Onions, V.J.; Mitchell, M.R.P.; Campbell, B.K.; Webb, R. Ovarian Tissue Viability Following Whole Ovine Ovary Cryopreservation: Assessing the Effects of Sphingosine-1-Phosphate Inclusion. Hum. Reprod. 2008, 23, 606–618. [Google Scholar] [CrossRef]
- Son, Y.-B.; Kang, Y.-H.; Lee, H.-J.; Jang, S.-J.; Bharti, D.; Lee, S.-L.; Jeon, B.-G.; Park, B.-W.; Rho, G.-J. Evaluation of Odonto/Osteogenic Differentiation Potential from Different Regions Derived Dental Tissue Stem Cells and Effect of 17β-Estradiol on Efficiency. BMC Oral Health 2021, 21, 15. [Google Scholar] [CrossRef]
- Thakur, G.; Bok, E.-Y.; Kim, S.-B.; Jo, C.-H.; Oh, S.-J.; Baek, J.-C.; Park, J.-E.; Kang, Y.-H.; Lee, S.-L.; Kumar, R.; et al. Scaffold-Free 3D Culturing Enhance Pluripotency, Immunomodulatory Factors, and Differentiation Potential of Wharton’s Jelly-Mesenchymal Stem Cells. Eur. J. Cell Biol. 2022, 101, 151245. [Google Scholar] [CrossRef]
- Vorotelyak, E.A.; Vasiliev, A.V.; Terskikh, V.V. On the Origin of Pluripotent Embryonic Stem Cells in the Mouse (Survey). Adv. Biol. Earth Sci. 2016, 1, 1–20. [Google Scholar]
- Woods, E.J.; Perry, B.C.; Hockema, J.J.; Larson, L.; Zhou, D.; Goebel, W.S. Optimized Cryopreservation Method for Human Dental Pulp-Derived Stem Cells and Their Tissues of Origin for Banking and Clinical Use. Cryobiology 2009, 59, 150–157. [Google Scholar] [CrossRef]
- Paes, S.M.; Pupo, Y.M.; Cavenago, B.C.; Fonseca-Silva, T.; Santos, C.C.d.O. Cryopreservation of Mesenchymal Stem Cells Derived from Dental Pulp: A Systematic Review. Restor. Dent. Endod. 2021, 46, e26. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-L.; Chang, W.-J.; Lin, C.-Y.; Hsieh, S.-C.; Lee, S.-Y.; Fan, K.-H.; Lin, C.-T.; Huang, H.-M. Static Magnetic Field Increases Survival Rate of Dental Pulp Stem Cells during DMSO-Free Cryopreservation. Electromagn. Biol. Med. 2015, 34, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Cottle, C.; Orter, A.P.; Lipat, A.; Turner-Lyles, C.; Nguyen, J.; Moll, G.; Chinnadurai, R. Impact of Cryopreservation and Freeze-Thawing on Therapeutic Properties of Mesenchymal Stromal/Stem Cells and Other Common Cellular Therapeutics. Curr. Stem Cell Rep. 2022, 8, 72–92. [Google Scholar] [CrossRef]
- Park, B.-W.; Jang, S.-J.; Byun, J.-H.; Kang, Y.-H.; Choi, M.-J.; Park, W.-U.; Lee, W.-J.; Rho, G.-J. Cryopreservation of Human Dental Follicle Tissue for Use as a Resource of Autologous Mesenchymal Stem Cells. J. Tissue Eng. Regen. Med. 2017, 11, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human Mesenchymal Stem Cells—Current Trends and Future Prospective. Biosci. Rep. 2015, 35, e00191. [Google Scholar] [CrossRef]
- Ezquer, F.E.; Ezquer, M.E.; Vicencio, J.M.; Calligaris, S.D. Two Complementary Strategies to Improve Cell Engraftment in Mesenchymal Stem Cell-Based Therapy: Increasing Transplanted Cell Resistance and Increasing Tissue Receptivity. Cell Adh. Migr. 2017, 11, 110–119. [Google Scholar] [CrossRef]
- Ullah, M.; Liu, D.D.; Thakor, A.S. Mesenchymal Stromal Cell Homing: Mechanisms and Strategies for Improvement. iScience 2019, 15, 421–438. [Google Scholar] [CrossRef]
- Donati, C.; Bruni, P. Sphingosine 1-Phosphate Regulates Cytoskeleton Dynamics: Implications in Its Biological Response. Biochim. Biophys. Acta 2006, 1758, 2037–2048. [Google Scholar] [CrossRef]
- Chamberlain, G.; Fox, J.; Ashton, B.; Middleton, J. Concise Review: Mesenchymal Stem Cells: Their Phenotype, Differentiation Capacity, Immunological Features, and Potential for Homing. Stem Cells 2007, 25, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Romani, R.; Manni, G.; Donati, C.; Pirisinu, I.; Bernacchioni, C.; Gargaro, M.; Pirro, M.; Calvitti, M.; Bagaglia, F.; Sahebkar, A.; et al. S1P Promotes Migration, Differentiation and Immune Regulatory Activity in Amniotic-Fluid-Derived Stem Cells. Eur. J. Pharmacol. 2018, 833, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Cai, X.; Liu, J.; Bai, B.; Li, X. Sphingosine 1-Phosphate Promotes Mesenchymal Stem Cell-Mediated Cardioprotection against Myocardial Infarction via ERK1/2-MMP-9 and Akt Signaling Axis. Life Sci. 2018, 215, 31–42. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Product Size (bp) | Accession No. |
|---|---|---|---|
| OCT4 | F: AAGCAGCGACTATGCACAAC R: AGTACAGTGCAGTGAAGTGAGG | 140 | NM_002701.5 |
| NANOG | F: GCAGATGCAAGAACTCTCCAAC R: CTGCGTCACACCATTGCTATTC | 133 | AB093576.1 |
| BAX | F: TCTGACGGCAACTTCAACTG R: AGTCCAATGTCCAGCCCATG | 127 | NM_001291428.1 |
| BAK | F: GGCACCTCAACATTGCATGG R: CAGTCTCTTGCCTCCCCAAG | 144 | NM_001188.3 |
| GAPDH | F: AGTCAGCCGCATCTTCTTTT R: CCAATACGACCAAATCCGTT | 97 | NM_002046.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, S.-J.; Jo, C.-H.; Kim, T.-S.; Hong, C.-Y.; Lee, S.-L.; Kang, Y.-H.; Rho, G.-J. Sphingosine-1-phosphate Treatment Improves Cryopreservation Efficiency in Human Mesenchymal Stem Cells. Life 2023, 13, 1286. https://doi.org/10.3390/life13061286
Oh S-J, Jo C-H, Kim T-S, Hong C-Y, Lee S-L, Kang Y-H, Rho G-J. Sphingosine-1-phosphate Treatment Improves Cryopreservation Efficiency in Human Mesenchymal Stem Cells. Life. 2023; 13(6):1286. https://doi.org/10.3390/life13061286
Chicago/Turabian StyleOh, Seong-Ju, Chan-Hee Jo, Tae-Seok Kim, Chae-Yeon Hong, Sung-Lim Lee, Young-Hoon Kang, and Gyu-Jin Rho. 2023. "Sphingosine-1-phosphate Treatment Improves Cryopreservation Efficiency in Human Mesenchymal Stem Cells" Life 13, no. 6: 1286. https://doi.org/10.3390/life13061286