
Epigenetic Modification Factors and microRNAs Network Associated with Differentiation of Embryonic Stem Cells and Induced Pluripotent Stem Cells toward Cardiomyocytes: A Review

 , ,
, , 
Abstract
:1. Introduction
2. Embryonic Stem Cells (ESCs)
3. Induced Pluripotent Stem Cells (iPSCs)
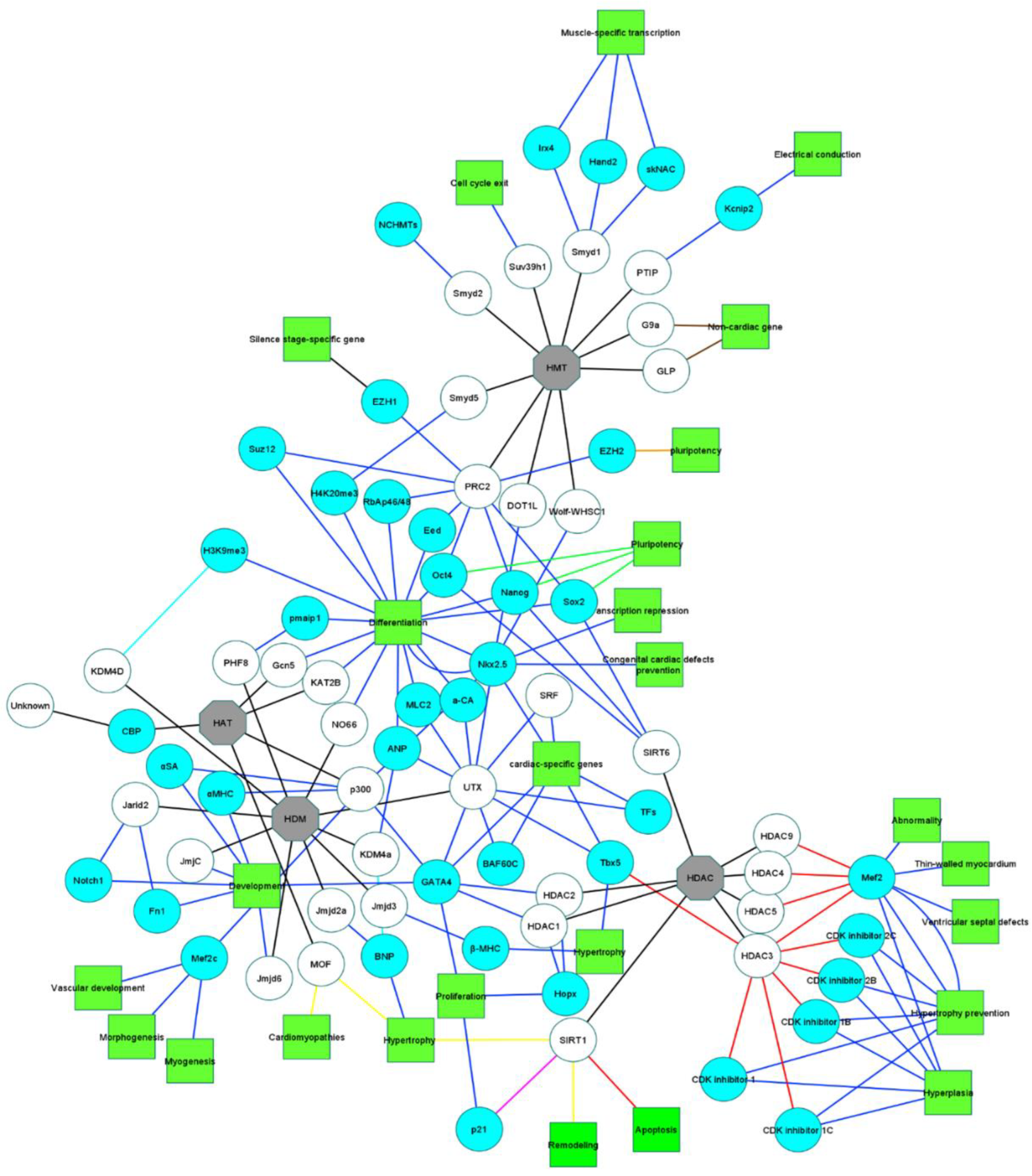
4. Epigenetics of Gene Expression and Silencing
4.1. Histone Modifications
4.1.1. Histone Acetylation of Stem Cells toward Cardiomyocyte Differentiation
4.1.2. Histone Deacetylation of Stem Cells toward Cardiomyocyte Differentiation
4.1.3. Histone Methylation of Stem Cells toward Cardiomyocyte Differentiation
4.1.4. Histone Demethylation of Stem Cells toward Cardiomyocyte Differentiation
4.2. Methylations
4.2.1. DNA Methylation
4.2.2. N6-Methyladenosine (m6a) RNA Methylation
4.3. ATP-Dependent Chromatin Remodeling Complexes
4.4. Cardiac Mitochondrial Mutations

{kind=link}
{kind=link}
| Epigenetic Factors | Function | Affected Gene | Effect(s) on Cardiac Tissue | Species | References | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Evolution | Physiology | Pathology | Anomaly | Unknown | ||||||
| HAT | ||||||||||
| p300 | Activation | αMHC | + | Mice | [74,197] | |||||
| Activation | αSA | + | Mice | |||||||
| Activation | GATA4 | + | Mice | [71,72,73] | ||||||
| Activation | Nkx2.5 | + | Mice | |||||||
| Activation | Mef2c | + | Mice | |||||||
| KAT2B | + | Human | [198] | |||||||
| Gcn5 | + | Rat | [80] | |||||||
| CBP | + | Mice | [78,79] | |||||||
| MOF | + | Mice | [77] | |||||||
| HDAC | HDAC1 | Activation | Hopx | + | Mice | [82,83] | ||||
| Activation | GATA4 | + | Mice | |||||||
| HDAC2 | Activation | Hopx | + | Mice | ||||||
| Activation | GATA4 | + | Mice | |||||||
| HDAC3 | Inhibition | Mef2 | + | Mice | [88,89] | |||||
| Inhibition | Cdkn1a | + | Mice | [84] | ||||||
| Inhibition | Cdkn1b | + | Mice | |||||||
| Inhibition | Cdkn1c | + | Mice | |||||||
| Inhibition | Cdkn2b | + | Mice | |||||||
| Inhibition | Cdkn2c | + | Mice | |||||||
| Inhibition | Tbx5 | + | + | Mice | [87] | |||||
| HDAC4 | Inhibition | Mef2 | + | Mice | [88,89] | |||||
| HDAC5 | Inhibition | Mef2 | + | + | Mice | [90,91] | ||||
| HDAC9 | Inhibition | Mef2 | + | + | Mice | |||||
| SIRT1 | Deacetylation | p21 | + | Mice | [199] | |||||
| SIRT1 | + | Mice | ||||||||
| SIRT6 | Activation | Oct4 | + | Mice | [109] | |||||
| Activation | Sox2 | + | Mice | |||||||
| Activation | Nanog | + | Mice | |||||||
| PTIP | Activation | Kcnip2 | + | Mice | [124] | |||||
| Smyd1 | Activation | skNAC | + | Mice | [117,128,129] | |||||
| Activation | Hand2 | + | Mice | |||||||
| Activation | Irx4 | + | Mice | |||||||
| Smyd2 | Activation | NCHMTs | + | Mice | [132] | |||||
| Smyd5 | Activation | H4K20me3 | + | + | Mice | [200] | ||||
| Wolf-WHSC1 | Activation | Nkx2.5 | + | + | Mice | [134] | ||||
| PRC2 | Activation | Oct4 | + | Human | [53] | |||||
| Activation | Sox2 | + | Human | |||||||
| Activation | Nanog | + | Human | |||||||
| Activation | EZH1 | + | Mice | [137,138,201] [138,139,202] | ||||||
| Activation | EZH2 | + | Mice | |||||||
| Activation | Eed | + | Mice | [136] | ||||||
| Activation | RbAp46/48 | + | Mice | |||||||
| Activation | Suz12 | + | Mice | |||||||
| G9a | + | + | Mice | [146] | ||||||
| GLP | + | + | Mice | |||||||
| Suv39h1 | + | Mice | [144] | |||||||
| DOT1L | Activation | Nkx2.5 | + | Human | [202] | |||||
| HDM | KDM4a | Activation | ANP | + | Mice | [89,156] | ||||
| Activation | BNP | + | Mice | |||||||
| KDM4D | Activation | H3K9me3 | + | Mice | [203] | |||||
| UTX | Activation | ANP | + | Mice | [122] | |||||
| Activation | MLC2 | + | Mice | |||||||
| Activation | a-CA | + | Mice | |||||||
| Activation | TFs | + | Mice | |||||||
| Activation | Nkx2.5 | + | Mice | |||||||
| Activation | Tbx5 | + | Mice | |||||||
| Activation | GATA4 | + | Mice | |||||||
| Activation | SRF | + | Mice | |||||||
| Activation | Baf60c | + | Mice | |||||||
| JmjC | NA | NA | + | Mice | [152] | |||||
| NO66 | + | Mice | [204] | |||||||
| Jmjd6 | + | + | Mice | [154] | ||||||
| Jmjd2a | Activation | BNP | + | Mice Human | [205] | |||||
| Jarid2 | Activation | Notch1 | + | Mice | [205,206] | |||||
| Activation | Fn1 | + | Mice | |||||||
| Jmjd3 | Activation | β-MHC | + | + | Mice Rat | [207,208] | ||||
| PHF8 | Activation | pmaip1 | + | Mice | [209] | |||||
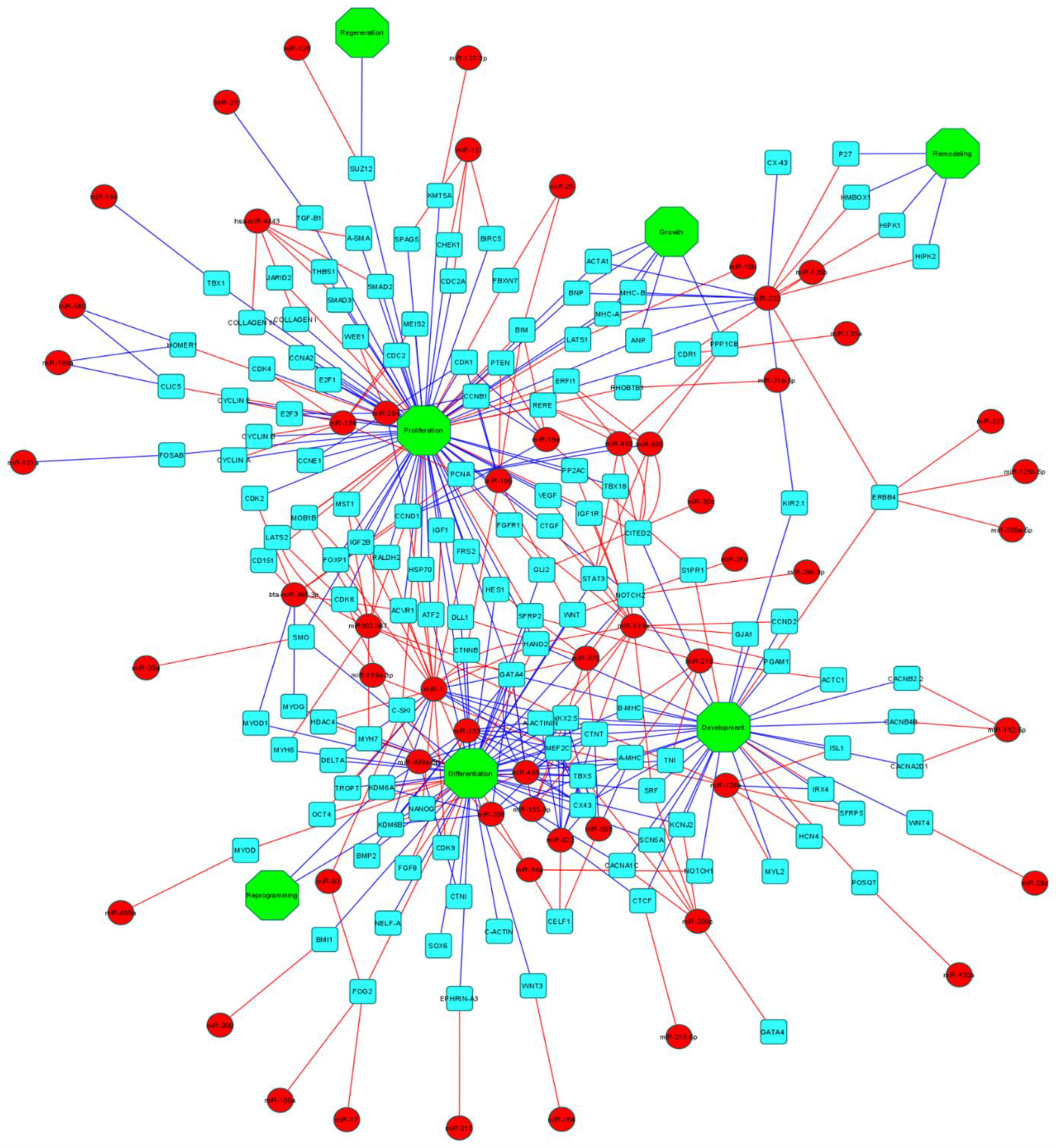
5. MicroRNAs (miRNAs)
5.1. miRNAs and Cardiomyocyte Differentiation
5.2. miRNAs and Congenital Heart Defects
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MSCs | Mesenchymal stem cells |
| hMSCs | Human mesenchymal stem cells |
| ESCs | Embryonic stem cells |
| iPSCs | Induced pluripotent stem cells |
| miRNAs | MicroRNAs |
| siRNA | Small interfering RNA |
| piRNA | Piwi-interacting RNA |
| HATs | Histone acetyltransferases |
| HDACs | Histone deacetylases |
| MOF | Males absent on the first |
| SIRTs | Sirtuins |
| SIRT1 | Sirtuin 1 |
| SIRT2 | Sirtuin 2 |
| SIRT3 | Sirtuin 3 |
| SIRT4 | Sirtuin 4 |
| SIRT5 | Sirtuin 5 |
| SIRT6 | Sirtuin 6 |
| SIRT7 | Sirtuin 7 |
| PTIP | PAX-interacting protein 1 |
| PRC2 | Polycomb repressive complex 2 |
| UTX | Ubiquitously transcribed tetratricopeptide repeat, X chromosome |
| CHD | Congenital heart defect |
| EVs | Extracellular vesicles |
| VEGF | vascular endothelial growth factor |
| 5-AZA | 5-azacytidine |
References
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2021 Update: A Report From the American Heart Association. Circulation 2021, 143, e254–e743. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.J.; Levant, S.; DeFrances, C.J. Hospitalization for Congestive Heart Failure: United States, 2000–2010; NCHS Data Brief: Washington, DC, USA; US Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Health Statistics: Washington, DC, USA, 2012; pp. 1–8.
- Benjamin, E.J.; Virani, S.S.; Callaway, C.W.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Chiuve, S.E.; Cushman, M.; Delling, F.N.; Deo, R.; et al. Heart Disease and Stroke Statistics-2018 Update: A Report From the American Heart Association. Circulation 2018, 137, e67–e492. [Google Scholar] [CrossRef] [PubMed]
- Verhestraeten, C.; Heggermont, W.A.; Maris, M. Clinical inertia in the treatment of heart failure: A major issue to tackle. Heart. Fail. Rev. 2021, 26, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Karbassi, E.; Fenix, A.; Marchiano, S.; Muraoka, N.; Nakamura, K.; Yang, X.; Murry, C.E. Cardiomyocyte maturation: Advances in knowledge and implications for regenerative medicine. Nat. Rev. Cardiol. 2020, 17, 341–359. [Google Scholar] [CrossRef]
- Yang, X.; Pabon, L.; Murry, C.E. Engineering adolescence: Maturation of human pluripotent stem cell-derived cardiomyocytes. Circ. Res. 2014, 114, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, P.; Sdek, P.; MacLellan, W.R. Cardiac myocyte cell cycle control in development, disease, and regeneration. Physiol. Rev. 2007, 87, 521–544. [Google Scholar] [CrossRef] [Green Version]
- Mollova, M.; Bersell, K.; Walsh, S.; Savla, J.; Das, L.T.; Park, S.Y.; Silberstein, L.E.; Dos Remedios, C.G.; Graham, D.; Colan, S.; et al. Cardiomyocyte proliferation contributes to heart growth in young humans. Proc. Natl. Acad. Sci. USA 2013, 110, 1446–1451. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, N.; Li, M.; Calvert, J.W.; Tejada, T.; Lambert, J.P.; Wu, J.; Kesteven, S.H.; Holman, S.R.; Matsuda, T.; Lovelock, J.D.; et al. A proliferative burst during preadolescence establishes the final cardiomyocyte number. Cell 2014, 157, 795–807. [Google Scholar] [CrossRef] [Green Version]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Dolati, S.; Yousefi, M.; Mahdipour, M.; Afrasiabi Rad, A.; Pishgahi, A.; Nouri, M.; Jodati, A.R. Mesenchymal stem cell and bone marrow mononuclear cell therapy for cardiomyopathy: From bench to bedside. J. Cell. Biochem. 2019, 120, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Bearzi, C.; Rota, M.; Hosoda, T.; Tillmanns, J.; Nascimbene, A.; De Angelis, A.; Yasuzawa-Amano, S.; Trofimova, I.; Siggins, R.W.; Lecapitaine, N.; et al. Human cardiac stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 14068–14073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cambria, E.; Pasqualini, F.S.; Wolint, P.; Gunter, J.; Steiger, J.; Bopp, A.; Hoerstrup, S.P.; Emmert, M.Y. Translational cardiac stem cell therapy: Advancing from first-generation to next-generation cell types. NPJ Regen. Med. 2017, 2, 17. [Google Scholar] [CrossRef]
- Evans, M.J.; Kaufman, M.H. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef] [PubMed]
- McLaren, A. Ethical and social considerations of stem cell research. Nature 2001, 414, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, H.S.; Srivastava, D. Stem cell therapy for cardiac disease. Pediatr. Res. 2012, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Mummery, C.; Ward-van Oostwaard, D.; Doevendans, P.; Spijker, R.; van den Brink, S.; Hassink, R.; van der Heyden, M.; Opthof, T.; Pera, M.; de la Riviere, A.B.; et al. Differentiation of human embryonic stem cells to cardiomyocytes: Role of coculture with visceral endoderm-like cells. Circulation 2003, 107, 2733–2740. [Google Scholar] [CrossRef] [Green Version]
- Graichen, R.; Xu, X.; Braam, S.R.; Balakrishnan, T.; Norfiza, S.; Sieh, S.; Soo, S.Y.; Tham, S.C.; Mummery, C.; Colman, A.; et al. Enhanced cardiomyogenesis of human embryonic stem cells by a small molecular inhibitor of p38 MAPK. Differentiation 2008, 76, 357–370. [Google Scholar] [CrossRef]
- Kawamura, M.; Miyagawa, S.; Miki, K.; Saito, A.; Fukushima, S.; Higuchi, T.; Kawamura, T.; Kuratani, T.; Daimon, T.; Shimizu, T.; et al. Feasibility, safety, and therapeutic efficacy of human induced pluripotent stem cell-derived cardiomyocyte sheets in a porcine ischemic cardiomyopathy model. Circulation 2012, 126, S29–S37. [Google Scholar] [CrossRef] [Green Version]
- Mummery, C.L.; Zhang, J.; Ng, E.S.; Elliott, D.A.; Elefanty, A.G.; Kamp, T.J. Differentiation of human embryonic stem cells and induced pluripotent stem cells to cardiomyocytes: A methods overview. Circ. Res. 2012, 111, 344–358. [Google Scholar] [CrossRef]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.J.; Tamadon, A.; Kim, Y.Y.; Kang, B.C.; Ku, S.Y. Epigenetic Regulation of Cardiomyocyte Differentiation from Embryonic and Induced Pluripotent Stem Cells. Int. J. Mol. Sci. 2021, 22, 8599. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Dent, S.Y. Chromatin modifiers and remodellers: Regulators of cellular differentiation. Nat. Rev. Genet. 2014, 15, 93–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Reinberg, D. Chromatin higher-order structures and gene regulation. Curr. Opin. Genet. Dev. 2011, 21, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kooistra, S.M.; Helin, K. Molecular mechanisms and potential functions of histone demethylases. Nat. Rev. Mol. Cell Biol. 2012, 13, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Paige, S.L.; Plonowska, K.; Xu, A.; Wu, S.M. Molecular regulation of cardiomyocyte differentiation. Circ. Res. 2015, 116, 341–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Castell-Auvi, A.; Cedo, L.; Movassat, J.; Portha, B.; Sanchez-Cabo, F.; Pallares, V.; Blay, M.; Pinent, M.; Ardevol, A. Procyanidins modulate microRNA expression in pancreatic islets. J. Agric Food Chem. 2013, 61, 355–363. [Google Scholar] [CrossRef]
- Plotnikova, O.; Baranova, A.; Skoblov, M. Comprehensive Analysis of Human microRNA-mRNA Interactome. Front. Genet. 2019, 10, 933. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Boon, R.A.; Dimmeler, S. MicroRNAs in myocardial infarction. Nat. Rev. Cardiol. 2015, 12, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Sluijter, J.P.; van Mil, A.; van Vliet, P.; Metz, C.H.; Liu, J.; Doevendans, P.A.; Goumans, M.J. MicroRNA-1 and -499 regulate differentiation and proliferation in human-derived cardiomyocyte progenitor cells. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 859–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivey, K.N.; Muth, A.; Arnold, J.; King, F.W.; Yeh, R.F.; Fish, J.E.; Hsiao, E.C.; Schwartz, R.J.; Conklin, B.R.; Bernstein, H.S.; et al. MicroRNA regulation of cell lineages in mouse and human embryonic stem cells. Cell Stem Cell 2008, 2, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, K.; Liu, D.; Lai, H.; Li, J.; Wang, C. Developing miRNA therapeutics for cardiac repair in ischemic heart disease. J. Thorac. Dis. 2016, 8, E918–E927. [Google Scholar] [CrossRef] [Green Version]
- Fard, M.; Akhavan-Tavakoli, M.; Khanjani, S.; Zare, S.; Edalatkhah, H.; Arasteh, S.; Mehrabani, D.; Zarnani, A.-H.; Kazemnejad, S.; Shirazi, R. Bilayer amniotic membrane/nano-fibrous fibroin scaffold promotes differentiation capability of menstrual blood stem cells into keratinocyte-like cells. Mol. Biotechnol. 2018, 60, 100–110. [Google Scholar] [CrossRef]
- Kashani, I.R.; Zarnani, A.H.; Soleimani, M.; Abdolvahabi, M.A.; Nayernia, K.; Shirazi, R. Retinoic acid induces mouse bone marrow-derived CD15+, Oct4+ and CXCR4+ stem cells into male germ-like cells in a two-dimensional cell culture system. Cell Biol. Int. 2014, 38, 782–789. [Google Scholar] [CrossRef]
- Kofidis, T.; de Bruin, J.L.; Yamane, T.; Tanaka, M.; Lebl, D.R.; Swijnenburg, R.J.; Weissman, I.L.; Robbins, R.C. Stimulation of paracrine pathways with growth factors enhances embryonic stem cell engraftment and host-specific differentiation in the heart after ischemic myocardial injury. Circulation 2005, 111, 2486–2493. [Google Scholar] [CrossRef] [Green Version]
- Rajasingh, J.; Bord, E.; Hamada, H.; Lambers, E.; Qin, G.; Losordo, D.W.; Kishore, R. STAT3-dependent mouse embryonic stem cell differentiation into cardiomyocytes: Analysis of molecular signaling and therapeutic efficacy of cardiomyocyte precommitted mES transplantation in a mouse model of myocardial infarction. Circ. Res. 2007, 101, 910–918. [Google Scholar] [CrossRef] [Green Version]
- Park, I.H.; Zhao, R.; West, J.A.; Yabuuchi, A.; Huo, H.; Ince, T.A.; Lerou, P.H.; Lensch, M.W.; Daley, G.Q. Reprogramming of human somatic cells to pluripotency with defined factors. Nature 2008, 451, 141–146. [Google Scholar] [CrossRef]
- Takahashi, K.; Okita, K.; Nakagawa, M.; Yamanaka, S. Induction of pluripotent stem cells from fibroblast cultures. Nat. Protoc. 2007, 2, 3081–3089. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Takahashi, K.; Ichisaka, T.; Aoi, T.; Okita, K.; Mochiduki, Y.; Takizawa, N.; Yamanaka, S. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat. Biotechnol. 2008, 26, 101–106. [Google Scholar] [CrossRef]
- Brambrink, T.; Foreman, R.; Welstead, G.G.; Lengner, C.J.; Wernig, M.; Suh, H.; Jaenisch, R. Sequential expression of pluripotency markers during direct reprogramming of mouse somatic cells. Cell Stem Cell 2008, 2, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadtfeld, M.; Nagaya, M.; Utikal, J.; Weir, G.; Hochedlinger, K. Induced pluripotent stem cells generated without viral integration. Science 2008, 322, 945–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woltjen, K.; Michael, I.P.; Mohseni, P.; Desai, R.; Mileikovsky, M.; Hamalainen, R.; Cowling, R.; Wang, W.; Liu, P.; Gertsenstein, M.; et al. piggyBac transposition reprograms fibroblasts to induced pluripotent stem cells. Nature 2009, 458, 766–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajasingh, J.; Lambers, E.; Hamada, H.; Bord, E.; Thorne, T.; Goukassian, I.; Krishnamurthy, P.; Rosen, K.M.; Ahluwalia, D.; Zhu, Y.; et al. Cell-free embryonic stem cell extract-mediated derivation of multipotent stem cells from NIH3T3 fibroblasts for functional and anatomical ischemic tissue repair. Circ. Res. 2008, 102, e107–e117. [Google Scholar] [CrossRef]
- Ban, H.; Nishishita, N.; Fusaki, N.; Tabata, T.; Saeki, K.; Shikamura, M.; Takada, N.; Inoue, M.; Hasegawa, M.; Kawamata, S.; et al. Efficient generation of transgene-free human induced pluripotent stem cells (iPSCs) by temperature-sensitive Sendai virus vectors. Proc. Natl. Acad. Sci. USA 2011, 108, 14234–14239. [Google Scholar] [CrossRef] [Green Version]
- Stadtfeld, M.; Hochedlinger, K. Induced pluripotency: History, mechanisms, and applications. Genes Dev. 2010, 24, 2239–2263. [Google Scholar] [CrossRef] [Green Version]
- Yagi, M.; Yamanaka, S.; Yamada, Y. Epigenetic foundations of pluripotent stem cells that recapitulate in vivo pluripotency. Lab. Investig. 2017, 97, 1133–1141. [Google Scholar] [CrossRef]
- Bernstein, B.E.; Mikkelsen, T.S.; Xie, X.; Kamal, M.; Huebert, D.J.; Cuff, J.; Fry, B.; Meissner, A.; Wernig, M.; Plath, K.; et al. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell 2006, 125, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Holliday, R. Epigenetics: A historical overview. Epigenetics 2006, 1, 76–80. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.I.; Jenner, R.G.; Boyer, L.A.; Guenther, M.G.; Levine, S.S.; Kumar, R.M.; Chevalier, B.; Johnstone, S.E.; Cole, M.F.; Isono, K.; et al. Control of developmental regulators by Polycomb in human embryonic stem cells. Cell 2006, 125, 301–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, P.; Schultz, R.M. Histone deacetylase 1 (HDAC1) regulates histone acetylation, development, and gene expression in preimplantation mouse embryos. Dev. Biol. 2008, 319, 110–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikkelsen, T.S.; Ku, M.; Jaffe, D.B.; Issac, B.; Lieberman, E.; Giannoukos, G.; Alvarez, P.; Brockman, W.; Kim, T.K.; Koche, R.P.; et al. Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 2007, 448, 553–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vakoc, C.R.; Sachdeva, M.M.; Wang, H.; Blobel, G.A. Profile of histone lysine methylation across transcribed mammalian chromatin. Mol. Cell. Biol. 2006, 26, 9185–9195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, S.; Geiman, T.M.; Briones, V.; Guang Tao, Y.; Xu, H.; Muegge, K. Lsh participates in DNA methylation and silencing of stem cell genes. Stem Cells 2009, 27, 2691–2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajkova, P. Epigenetic reprogramming in the germline: Towards the ground state of the epigenome. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 2266–2273. [Google Scholar] [CrossRef] [Green Version]
- Simon, J.A.; Kingston, R.E. Mechanisms of polycomb gene silencing: Knowns and unknowns. Nat. Rev. Mol. Cell Biol. 2009, 10, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Bibikova, M.; Laurent, L.C.; Ren, B.; Loring, J.F.; Fan, J.B. Unraveling epigenetic regulation in embryonic stem cells. Cell Stem Cell 2008, 2, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Mahdipour, M.; van Tol, H.T.; Stout, T.A.; Roelen, B.A. Validating reference microRNAs for normalizing qRT-PCR data in bovine oocytes and preimplantation embryos. BMC Dev. Biol. 2015, 15, 25. [Google Scholar] [CrossRef] [Green Version]
- Roovers, E.F.; Rosenkranz, D.; Mahdipour, M.; Han, C.T.; He, N.; Chuva de Sousa Lopes, S.M.; van der Westerlaken, L.A.; Zischler, H.; Butter, F.; Roelen, B.A.; et al. Piwi proteins and piRNAs in mammalian oocytes and early embryos. Cell Rep. 2015, 10, 2069–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paige, S.L.; Thomas, S.; Stoick-Cooper, C.L.; Wang, H.; Maves, L.; Sandstrom, R.; Pabon, L.; Reinecke, H.; Pratt, G.; Keller, G.; et al. A temporal chromatin signature in human embryonic stem cells identifies regulators of cardiac development. Cell 2012, 151, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wamstad, J.A.; Alexander, J.M.; Truty, R.M.; Shrikumar, A.; Li, F.; Eilertson, K.E.; Ding, H.; Wylie, J.N.; Pico, A.R.; Capra, J.A.; et al. Dynamic and coordinated epigenetic regulation of developmental transitions in the cardiac lineage. Cell 2012, 151, 206–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luger, K.; Mader, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef]
- Phillips, D.M. The presence of acetyl groups of histones. Biochem. J. 1963, 87, 258–263. [Google Scholar] [CrossRef] [Green Version]
- Xhemalce, B.; Dawson, M.A.; Bannister, A.J. Histone Modifications. In Encyclopedia of Molecular Cell Biology and Molecular Medicine; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2011. [Google Scholar] [CrossRef]
- Clayton, A.L.; Hebbes, T.R.; Thorne, A.W.; Crane-Robinson, C. Histone acetylation and gene induction in human cells. FEBS Lett. 1993, 336, 23–26. [Google Scholar] [CrossRef] [Green Version]
- Sartorelli, V.; Huang, J.; Hamamori, Y.; Kedes, L. Molecular mechanisms of myogenic coactivation by p300: Direct interaction with the activation domain of MyoD and with the MADS box of MEF2C. Mol. Cell. Biol. 1997, 17, 1010–1026. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, T.; Ono, K.; Morimoto, T.; Wada, H.; Hirai, M.; Hidaka, K.; Morisaki, T.; Heike, T.; Nakahata, T.; Kita, T.; et al. Acetylation of GATA-4 is involved in the differentiation of embryonic stem cells into cardiac myocytes. J. Biol. Chem. 2005, 280, 19682–19688. [Google Scholar] [CrossRef] [Green Version]
- Ma, K.; Chan, J.K.; Zhu, G.; Wu, Z. Myocyte enhancer factor 2 acetylation by p300 enhances its DNA binding activity, transcriptional activity, and myogenic differentiation. Mol. Cell. Biol. 2005, 25, 3575–3582. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Yang, X.; Zhu, J.; Lv, T.; Chen, Y.; Chen, G.; Zhong, L.; Li, Y.; Huang, X.; Huang, G.; et al. Inhibition of p300-HAT results in a reduced histone acetylation and down-regulation of gene expression in cardiac myocytes. Life Sci. 2010, 87, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Yao, T.P.; Oh, S.P.; Fuchs, M.; Zhou, N.D.; Ch’ng, L.E.; Newsome, D.; Bronson, R.T.; Li, E.; Livingston, D.M.; Eckner, R. Gene dosage-dependent embryonic development and proliferation defects in mice lacking the transcriptional integrator p300. Cell 1998, 93, 361–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikama, N.; Lutz, W.; Kretzschmar, R.; Sauter, N.; Roth, J.F.; Marino, S.; Wittwer, J.; Scheidweiler, A.; Eckner, R. Essential function of p300 acetyltransferase activity in heart, lung and small intestine formation. EMBO J. 2003, 22, 5175–5185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schueler, M.; Zhang, Q.; Schlesinger, J.; Tonjes, M.; Sperling, S.R. Dynamics of Srf, p300 and histone modifications during cardiac maturation in mouse. Mol. Biosyst. 2012, 8, 495–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, W.; Zhang, W.; Gai, Y.; Zhao, L.; Fan, J. The histone acetyltransferase MOF overexpression blunts cardiac hypertrophy by targeting ROS in mice. Biochem. Biophys. Res. Commun. 2014, 448, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Naruse, I.; Hongo, T.; Xu, M.; Nakahata, T.; Maekawa, T.; Ishii, S. Extensive brain hemorrhage and embryonic lethality in a mouse null mutant of CREB-binding protein. Mech. Dev. 2000, 95, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhu, J.; Lv, T.; Wu, G.; Sun, H.; Huang, X.; Tian, J. Spatiotemporal expression of histone acetyltransferases, p300 and CBP, in developing embryonic hearts. J. Biomed. Sci. 2009, 16, 24. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhu, J.; Tian, J.; Liu, X.; Feng, C. A role for Gcn5 in cardiomyocyte differentiation of rat mesenchymal stem cells. Mol. Cell. Biochem. 2010, 345, 309–316. [Google Scholar] [CrossRef]
- Montgomery, R.L.; Davis, C.A.; Potthoff, M.J.; Haberland, M.; Fielitz, J.; Qi, X.; Hill, J.A.; Richardson, J.A.; Olson, E.N. Histone deacetylases 1 and 2 redundantly regulate cardiac morphogenesis, growth, and contractility. Genes Dev. 2007, 21, 1790–1802. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, C.M.; Zhu, W.; Wang, Q.; Jia, C.; Kee, H.J.; Li, L.; Hannenhalli, S.; Epstein, J.A. Hopx and Hdac2 interact to modulate Gata4 acetylation and embryonic cardiac myocyte proliferation. Dev. Cell 2010, 19, 450–459. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Kook, H.; Milewski, R.; Gitler, A.D.; Lu, M.M.; Li, J.; Nazarian, R.; Schnepp, R.; Jen, K.; Biben, C.; et al. Hop is an unusual homeobox gene that modulates cardiac development. Cell 2002, 110, 713–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, C.M.; Lu, M.M.; Wang, Q.; Epstein, J.A. Transgenic overexpression of Hdac3 in the heart produces increased postnatal cardiac myocyte proliferation but does not induce hypertrophy. J. Biol. Chem. 2008, 283, 26484–26489. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, R.L.; Potthoff, M.J.; Haberland, M.; Qi, X.; Matsuzaki, S.; Humphries, K.M.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. Maintenance of cardiac energy metabolism by histone deacetylase 3 in mice. J. Clin. Investig. 2008, 118, 3588–3597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greulich, F.; Rudat, C.; Kispert, A. Mechanisms of T-box gene function in the developing heart. Cardiovasc Res. 2011, 91, 212–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowski, S.L.; Janardhan, H.P.; Smee, K.M.; Bachman, M.; Sun, Z.; Lazar, M.A.; Trivedi, C.M. Histone deacetylase 3 modulates Tbx5 activity to regulate early cardiogenesis. Hum. Mol. Genet. 2014, 23, 3801–3809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backs, J.; Worst, B.C.; Lehmann, L.H.; Patrick, D.M.; Jebessa, Z.; Kreusser, M.M.; Sun, Q.; Chen, L.; Heft, C.; Katus, H.A.; et al. Selective repression of MEF2 activity by PKA-dependent proteolysis of HDAC4. J. Cell Biol. 2011, 195, 403–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohl, M.; Wagner, M.; Reil, J.C.; Muller, S.A.; Tauchnitz, M.; Zimmer, A.M.; Lehmann, L.H.; Thiel, G.; Bohm, M.; Backs, J.; et al. HDAC4 controls histone methylation in response to elevated cardiac load. J. Clin. Investig. 2013, 123, 1359–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.L.; McKinsey, T.A.; Chang, S.; Antos, C.L.; Hill, J.A.; Olson, E.N. Class II histone deacetylases act as signal-responsive repressors of cardiac hypertrophy. Cell 2002, 110, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.; McKinsey, T.A.; Zhang, C.L.; Richardson, J.A.; Hill, J.A.; Olson, E.N. Histone deacetylases 5 and 9 govern responsiveness of the heart to a subset of stress signals and play redundant roles in heart development. Mol. Cell. Biol. 2004, 24, 8467–8476. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.; Heisig, J.; Kneitz, S.; Wolf, E.; Eilers, M.; Gessler, M. Mechanisms of epigenetic and cell-type specific regulation of Hey target genes in ES cells and cardiomyocytes. J. Mol. Cell. Cardiol. 2015, 79, 79–88. [Google Scholar] [CrossRef]
- Li, X.; Kazgan, N. Mammalian sirtuins and energy metabolism. Int. J. Biol. Sci 2011, 7, 575–587. Available online: https://www.ijbs.com/v07p0575.htm (accessed on 28 September 2022). [CrossRef] [PubMed] [Green Version]
- Herskovits, A.Z.; Guarente, L. Sirtuin deacetylases in neurodegenerative diseases of aging. Cell. Res. 2013, 23, 746–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Wang, L.; Jiang, W.; Xiong, Y.; Pang, L.; Zhong, Y.; Zhang, C.; Ou, W.; Tian, C.; Chen, X.; et al. Myocyte enhancer factor 2A delays vascular endothelial cell senescence by activating the PI3K/p-Akt/SIRT1 pathway. Aging (Albany NY) 2019, 11, 3768–3784. [Google Scholar] [CrossRef]
- Tsai, K.L.; Hung, C.H.; Chan, S.H.; Hsieh, P.L.; Ou, H.C.; Cheng, Y.H.; Chu, P.M. Chlorogenic Acid Protects Against oxLDL-Induced Oxidative Damage and Mitochondrial Dysfunction by Modulating SIRT1 in Endothelial Cells. Mol. Nutr. Food. Res. 2018, 62, e1700928. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.S.; Chuang, S.Y.; Shen, C.Y.; Ho, T.J.; Chang, R.L.; Yeh, Y.L.; Kuo, C.H.; Mahalakshmi, B.; Kuo, W.W.; Huang, C.Y. Antioxidant Sirt1/Akt axis expression in resveratrol pretreated adipose-derived stem cells increases regenerative capability in a rat model with cardiomyopathy induced by diabetes mellitus. J. Cell. Physiol. 2021, 236, 4290–4302. [Google Scholar] [CrossRef]
- Choudhery, M.S.; Khan, M.; Mahmood, R.; Mohsin, S.; Akhtar, S.; Ali, F.; Khan, S.N.; Riazuddin, S. Mesenchymal stem cells conditioned with glucose depletion augments their ability to repair-infarcted myocardium. J. Cell. Mol. Med. 2012, 16, 2518–2529. [Google Scholar] [CrossRef]
- Yeh, E.T.; Bickford, C.L. Cardiovascular complications of cancer therapy: Incidence, pathogenesis, diagnosis, and management. J. Am. Coll. Cardiol. 2009, 53, 2231–2247. [Google Scholar] [CrossRef] [Green Version]
- Hequet, O.; Le, Q.H.; Moullet, I.; Pauli, E.; Salles, G.; Espinouse, D.; Dumontet, C.; Thieblemont, C.; Arnaud, P.; Antal, D.; et al. Subclinical late cardiomyopathy after doxorubicin therapy for lymphoma in adults. J. Clin. Oncol. 2004, 22, 1864–1871. [Google Scholar] [CrossRef]
- Zhuang, L.; Xia, W.; Chen, D.; Ye, Y.; Hu, T.; Li, S.; Hou, M. Exosomal LncRNA-NEAT1 derived from MIF-treated mesenchymal stem cells protected against doxorubicin-induced cardiac senescence through sponging miR-221-3p. J. Nanobiotechnology. 2020, 18, 157. [Google Scholar] [CrossRef]
- Mao, Q.; Liang, X.L.; Zhang, C.L.; Pang, Y.H.; Lu, Y.X. LncRNA KLF3-AS1 in human mesenchymal stem cell-derived exosomes ameliorates pyroptosis of cardiomyocytes and myocardial infarction through miR-138-5p/Sirt1 axis. Stem. Cell. Res. Ther. 2019, 10, 393. [Google Scholar] [CrossRef] [Green Version]
- Denu, R.A. SIRT3 Enhances Mesenchymal Stem Cell Longevity and Differentiation. Oxid. Med. Cell. Longev. 2017, 2017, 5841716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellizzi, D.; Rose, G.; Cavalcante, P.; Covello, G.; Dato, S.; De Rango, F.; Greco, V.; Maggiolini, M.; Feraco, E.; Mari, V.; et al. A novel VNTR enhancer within the SIRT3 gene, a human homologue of SIR2, is associated with survival at oldest ages. Genomics 2005, 85, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Gao, T.; Xu, R.J.; Sun, L.; Zhang, C.F.; Bai, L.; Chen, W.; Liu, K.Y.; Zhou, Y.; Jiao, X.; et al. SIRT3 Transfection of Aged Human Bone Marrow-Derived Mesenchymal Stem Cells Improves Cell Therapy-Mediated Myocardial Repair. Rejuvenation Res. 2020, 23, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Sarikhani, M.; Garbern, J.C.; Ma, S.; Sereda, R.; Conde, J.; Krähenbühl, G.; Escalante, G.O.; Ahmed, A.; Buenrostro, J.D.; Lee, R.T. Sustained Activation of AMPK Enhances Differentiation of Human iPSC-Derived Cardiomyocytes via Sirtuin Activation. Stem. Cell Rep. 2020, 15, 498–514. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Song, C.; Young, N.L.; Sperling, A.S.; Xu, F.; Sridharan, R.; Conway, A.E.; Garcia, B.A.; Plath, K.; Clark, A.T.; et al. Histone h3 lysine 56 acetylation is linked to the core transcriptional network in human embryonic stem cells. Mol. Cell 2009, 33, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Zwaans, B.M.; Eckersdorff, M.; Lombard, D.B. The sirtuin SIRT6 deacetylates H3 K56Ac in vivo to promote genomic stability. Cell Cycle 2009, 8, 2662–2663. [Google Scholar] [CrossRef] [Green Version]
- Etchegaray, J.P.; Chavez, L.; Huang, Y.; Ross, K.N.; Choi, J.; Martinez-Pastor, B.; Walsh, R.M.; Sommer, C.A.; Lienhard, M.; Gladden, A.; et al. The histone deacetylase SIRT6 controls embryonic stem cell fate via TET-mediated production of 5-hydroxymethylcytosine. Nat. Cell Biol. 2015, 17, 545–557. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, L.D.; Ravi, V.; Jain, A.; Panicker, A.G.; Sundaresan, N.R.; Chatterjee, K. Sirtuin 6 mediated stem cell cardiomyogenesis on protein coated nanofibrous scaffolds. Nanomedicine 2019, 19, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Murray, K. The Occurrence of Epsilon-N-Methyl Lysine in Histones. Biochemistry 1964, 3, 10–15. [Google Scholar] [CrossRef]
- Turner, B.M. Reading signals on the nucleosome with a new nomenclature for modified histones. Nat. Struct. Mol. Biol. 2005, 12, 110–112. [Google Scholar] [CrossRef]
- Waterborg, J.H. Dynamic methylation of alfalfa histone H3. J. Biol. Chem. 1993, 268, 4918–4921. [Google Scholar] [CrossRef] [PubMed]
- Hon, G.; Wang, W.; Ren, B. Discovery and annotation of functional chromatin signatures in the human genome. PLoS Comput. Biol. 2009, 5, e1000566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, A.; Wu, R.; Peetz, M.; Gygi, S.P.; Moazed, D. Heterochromatic gene silencing by activator interference and a transcription elongation barrier. J. Biol. Chem. 2013, 288, 28771–28782. [Google Scholar] [CrossRef] [Green Version]
- Nimura, K.; Ura, K.; Shiratori, H.; Ikawa, M.; Okabe, M.; Schwartz, R.J.; Kaneda, Y. A histone H3 lysine 36 trimethyltransferase links Nkx2-5 to Wolf-Hirschhorn syndrome. Nature 2009, 460, 287–291. [Google Scholar] [CrossRef]
- Gottlieb, P.D.; Pierce, S.A.; Sims, R.J.; Yamagishi, H.; Weihe, E.K.; Harriss, J.V.; Maika, S.D.; Kuziel, W.A.; King, H.L.; Olson, E.N.; et al. Bop encodes a muscle-restricted protein containing MYND and SET domains and is essential for cardiac differentiation and morphogenesis. Nat. Genet. 2002, 31, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.Y.; Schones, D.E.; Wang, Z.; Wei, G.; Chepelev, I.; Zhao, K. High-resolution profiling of histone methylations in the human genome. Cell 2007, 129, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Fujii, T.; Tsunesumi, S.; Yamaguchi, K.; Watanabe, S.; Furukawa, Y. Smyd3 is required for the development of cardiac and skeletal muscle in zebrafish. PLoS ONE 2011, 6, e23491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Movassagh, M.; Choy, M.K.; Knowles, D.A.; Cordeddu, L.; Haider, S.; Down, T.; Siggens, L.; Vujic, A.; Simeoni, I.; Penkett, C.; et al. Distinct epigenomic features in end-stage failing human hearts. Circulation 2011, 124, 2411–2422. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Neppl, R.L.; Huang, Z.P.; Chen, J.; Tang, R.H.; Cao, R.; Zhang, Y.; Jin, S.W.; Wang, D.Z. The histone methyltransferase Set7/9 promotes myoblast differentiation and myofibril assembly. J. Cell Biol. 2011, 194, 551–565. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Lee, J.W.; Lee, S.K. UTX, a histone H3-lysine 27 demethylase, acts as a critical switch to activate the cardiac developmental program. Dev. Cell 2012, 22, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, S.; Choi, M.; Wakimoto, H.; Ma, L.; Jiang, J.; Overton, J.D.; Romano-Adesman, A.; Bjornson, R.D.; Breitbart, R.E.; Brown, K.K.; et al. De novo mutations in histone-modifying genes in congenital heart disease. Nature 2013, 498, 220–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, A.B.; Jones, T.A.; Herron, T.J.; Patel, S.R.; Day, S.M.; Noujaim, S.F.; Milstein, M.L.; Klos, M.; Furspan, P.B.; Jalife, J.; et al. Loss of H3K4 methylation destabilizes gene expression patterns and physiological functions in adult murine cardiomyocytes. J. Clin. Investig. 2011, 121, 2641–2650. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.B.; Goonewardena, S.N.; Jones, T.A.; Prusick, P.J.; Bazzi, A.A.; Belyavskaya, J.M.; McCoskey, M.M.; Dandar, R.A. The PTIP-Associated Histone Methyltransferase Complex Prevents Stress-Induced Maladaptive Cardiac Remodeling. PLoS ONE 2015, 10, e0127839. [Google Scholar] [CrossRef] [PubMed]
- Sims, R.J., III; Weihe, E.K.; Zhu, L.; O’Malley, S.; Harriss, J.V.; Gottlieb, P.D. m-Bop, a repressor protein essential for cardiogenesis, interacts with skNAC, a heart- and muscle-specific transcription factor. J. Biol. Chem. 2002, 277, 26524–26529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Rotllant, J.; Li, H.; De Deyne, P.; Du, S.J. SmyD1, a histone methyltransferase, is required for myofibril organization and muscle contraction in zebrafish embryos. Proc. Natl. Acad. Sci. USA 2006, 103, 2713–2718. [Google Scholar] [CrossRef] [Green Version]
- Sirinupong, N.; Brunzelle, J.; Ye, J.; Pirzada, A.; Nico, L.; Yang, Z. Crystal structure of cardiac-specific histone methyltransferase SmyD1 reveals unusual active site architecture. J. Biol. Chem. 2010, 285, 40635–40644. [Google Scholar] [CrossRef] [Green Version]
- Park, C.Y.; Pierce, S.A.; von Drehle, M.; Ivey, K.N.; Morgan, J.A.; Blau, H.M.; Srivastava, D. skNAC, a Smyd1-interacting transcription factor, is involved in cardiac development and skeletal muscle growth and regeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 20750–20755. [Google Scholar] [CrossRef] [Green Version]
- Just, S.; Meder, B.; Berger, I.M.; Etard, C.; Trano, N.; Patzel, E.; Hassel, D.; Marquart, S.; Dahme, T.; Vogel, B. The myosin-interacting protein SMYD1 is essential for sarcomere organization. J. Cell Sci. 2011, 124, 3127–3136. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhong, Y.; Wang, Z.; Gao, J.; Xu, J.; Chu, W.; Zhang, J.; Fang, S.; Du, S.J. Smyd1b is required for skeletal and cardiac muscle function in zebrafish. Mol. Biol. Cell. 2013, 24, 3511–3521. [Google Scholar] [CrossRef] [PubMed]
- Costantini, D.L.; Arruda, E.P.; Agarwal, P.; Kim, K.H.; Zhu, Y.; Zhu, W.; Lebel, M.; Cheng, C.W.; Park, C.Y.; Pierce, S.A.; et al. The homeodomain transcription factor Irx5 establishes the mouse cardiac ventricular repolarization gradient. Cell 2005, 123, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Diehl, F.; Brown, M.A.; van Amerongen, M.J.; Novoyatleva, T.; Wietelmann, A.; Harriss, J.; Ferrazzi, F.; Bottger, T.; Harvey, R.P.; Tucker, P.W.; et al. Cardiac deletion of Smyd2 is dispensable for mouse heart development. PLoS ONE 2010, 5, e9748. [Google Scholar] [CrossRef] [Green Version]
- Bergemann, A.D.; Cole, F.; Hirschhorn, K. The etiology of Wolf-Hirschhorn syndrome. Trends Genet. 2005, 21, 188–195. [Google Scholar] [CrossRef]
- Hojfeldt, J.W.; Laugesen, A.; Willumsen, B.M.; Damhofer, H.; Hedehus, L.; Tvardovskiy, A.; Mohammad, F.; Jensen, O.N.; Helin, K. Accurate H3K27 methylation can be established de novo by SUZ12-directed PRC2. Nat. Struct. Mol. Biol. 2018, 25, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Margueron, R.; Reinberg, D. The Polycomb complex PRC2 and its mark in life. Nature 2011, 469, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Liu, Y.; Hsu, Y.J.; Fujiwara, Y.; Kim, J.; Mao, X.; Yuan, G.C.; Orkin, S.H. EZH1 mediates methylation on histone H3 lysine 27 and complements EZH2 in maintaining stem cell identity and executing pluripotency. Mol. Cell 2008, 32, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasini, D.; Bracken, A.P.; Hansen, J.B.; Capillo, M.; Helin, K. The polycomb group protein Suz12 is required for embryonic stem cell differentiation. Mol. Cell. Biol. 2007, 27, 3769–3779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, N.D.; Yee, D.; Chen, A.; Kalantry, S.; Chamberlain, S.J.; Otte, A.P.; Magnuson, T. The murine polycomb group protein Eed is required for global histone H3 lysine-27 methylation. Curr. Biol. 2005, 15, 942–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Olguin, P.; Huang, Y.; Li, X.; Christodoulou, D.; Seidman, C.E.; Seidman, J.G.; Tarakhovsky, A.; Bruneau, B.G. Epigenetic repression of cardiac progenitor gene expression by Ezh2 is required for postnatal cardiac homeostasis. Nat. Genet. 2012, 44, 343–347. [Google Scholar] [CrossRef] [Green Version]
- He, A.; Ma, Q.; Cao, J.; von Gise, A.; Zhou, P.; Xie, H.; Zhang, B.; Hsing, M.; Christodoulou, D.C.; Cahan, P.; et al. Polycomb repressive complex 2 regulates normal development of the mouse heart. Circ. Res. 2012, 110, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Lachner, M.; O’Sullivan, R.J.; Jenuwein, T. An epigenetic road map for histone lysine methylation. J. Cell Sci. 2003, 116, 2117–2124. [Google Scholar] [CrossRef] [Green Version]
- Santos-Rosa, H.; Schneider, R.; Bannister, A.J.; Sherriff, J.; Bernstein, B.E.; Emre, N.C.; Schreiber, S.L.; Mellor, J.; Kouzarides, T. Active genes are tri-methylated at K4 of histone H3. Nature 2002, 419, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Sdek, P.; Zhao, P.; Wang, Y.; Huang, C.J.; Ko, C.Y.; Butler, P.C.; Weiss, J.N.; Maclellan, W.R. Rb and p130 control cell cycle gene silencing to maintain the postmitotic phenotype in cardiac myocytes. J. Cell Biol. 2011, 194, 407–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, A.; Shen, X.; Ma, Q.; Cao, J.; von Gise, A.; Zhou, P.; Wang, G.; Marquez, V.E.; Orkin, S.H.; Pu, W.T. PRC2 directly methylates GATA4 and represses its transcriptional activity. Genes Dev. 2012, 26, 37–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagawa, M.; Nakajima, K.; Makino, T.; Ogawa, S.; Kojima, M.; Ito, S.; Ikenishi, A.; Hayashi, T.; Schwartz, R.J.; Nakamura, K.; et al. Histone H3 lysine 9 methyltransferases, G9a and GLP are essential for cardiac morphogenesis. Mech. Dev. 2013, 130, 519–531. [Google Scholar] [CrossRef]
- Mezentseva, N.V.; Yang, J.; Kaur, K.; Iaffaldano, G.; Remond, M.C.; Eisenberg, C.A.; Eisenberg, L.M. The histone methyltransferase inhibitor BIX01294 enhances the cardiac potential of bone marrow cells. Stem Cells Dev. 2013, 22, 654–667. [Google Scholar] [CrossRef] [Green Version]
- Savickiene, J.; Treigyte, G.; Jazdauskaite, A.; Borutinskaite, V.V.; Navakauskiene, R. DNA methyltransferase inhibitor RG108 and histone deacetylase inhibitors cooperate to enhance NB4 cell differentiation and E-cadherin re-expression by chromatin remodelling. Cell Biol. Int. 2012, 36, 1067–1078. [Google Scholar] [CrossRef]
- Qian, Q.; Qian, H.; Zhang, X.; Zhu, W.; Yan, Y.; Ye, S.; Peng, X.; Li, W.; Xu, Z.; Sun, L.; et al. 5-Azacytidine induces cardiac differentiation of human umbilical cord-derived mesenchymal stem cells by activating extracellular regulated kinase. Stem Cells Dev. 2012, 21, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Minami, I.; Yamada, K.; Otsuji, T.G.; Yamamoto, T.; Shen, Y.; Otsuka, S.; Kadota, S.; Morone, N.; Barve, M.; Asai, Y.; et al. A small molecule that promotes cardiac differentiation of human pluripotent stem cells under defined, cytokine- and xeno-free conditions. Cell Rep. 2012, 2, 1448–1460. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Song, A.J.; Baker, R.; Micales, B.; Conway, S.J.; Lyons, G.E. Jumonji, a nuclear protein that is necessary for normal heart development. Circ. Res. 2000, 86, 932–938. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, T.; Watanabe, Y.; Takano-Shimizu, T.; Kondo, S. Roles of jumonji and jumonji family genes in chromatin regulation and development. Dev. Dyn. 2006, 235, 2449–2459. [Google Scholar] [CrossRef]
- Klose, R.J.; Kallin, E.M.; Zhang, Y. JmjC-domain-containing proteins and histone demethylation. Nat. Rev. Genet. 2006, 7, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.; Chen, Y.; Zhao, Y.; Bruick, R.K. JMJD6 is a histone arginine demethylase. Science 2007, 318, 444–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whetstine, J.R.; Nottke, A.; Lan, F.; Huarte, M.; Smolikov, S.; Chen, Z.; Spooner, E.; Li, E.; Zhang, G.; Colaiacovo, M.; et al. Reversal of histone lysine trimethylation by the JMJD2 family of histone demethylases. Cell 2006, 125, 467–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.J.; Chen, H.Z.; Wang, L.; Liu, D.P.; Hill, J.A.; Liu, Z.P. The histone trimethyllysine demethylase JMJD2A promotes cardiac hypertrophy in response to hypertrophic stimuli in mice. J. Clin. Investig. 2011, 121, 2447–2456. [Google Scholar] [CrossRef] [Green Version]
- Agger, K.; Cloos, P.A.; Christensen, J.; Pasini, D.; Rose, S.; Rappsilber, J.; Issaeva, I.; Canaani, E.; Salcini, A.E.; Helin, K. UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature 2007, 449, 731–734. [Google Scholar] [CrossRef]
- Lan, F.; Bayliss, P.E.; Rinn, J.L.; Whetstine, J.R.; Wang, J.K.; Chen, S.; Iwase, S.; Alpatov, R.; Issaeva, I.; Canaani, E.; et al. A histone H3 lysine 27 demethylase regulates animal posterior development. Nature 2007, 449, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Cho, Y.W.; Yu, L.R.; Yu, H.; Veenstra, T.D.; Ge, K. Identification of JmjC domain-containing UTX and JMJD3 as histone H3 lysine 27 demethylases. Proc. Natl. Acad. Sci. USA 2007, 104, 18439–18444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, M.; Krassowska, A.; Gilbert, N.; Chevassut, T.; Forrester, L.; Ansell, J.; Ramsahoye, B. Severe global DNA hypomethylation blocks differentiation and induces histone hyperacetylation in embryonic stem cells. Mol. Cell. Biol. 2004, 24, 8862–8871. [Google Scholar] [CrossRef] [Green Version]
- Klose, R.J.; Bird, A.P. Genomic DNA methylation: The mark and its mediators. Trends Biochem. Sci. 2006, 31, 89–97. [Google Scholar] [CrossRef]
- Trojer, P.; Reinberg, D. Histone lysine demethylases and their impact on epigenetics. Cell 2006, 125, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Dudley, D.D.; Chaudhuri, J.; Bassing, C.H.; Alt, F.W. Mechanism and control of V(D)J recombination versus class switch recombination: Similarities and differences. Adv. Immunol. 2005, 86, 43–112. [Google Scholar] [CrossRef] [PubMed]
- Iyer, L.M.; Tahiliani, M.; Rao, A.; Aravind, L. Prediction of novel families of enzymes involved in oxidative and other complex modifications of bases in nucleic acids. Cell Cycle 2009, 8, 1698–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahiliani, M.; Koh, K.P.; Shen, Y.; Pastor, W.A.; Bandukwala, H.; Brudno, Y.; Agarwal, S.; Iyer, L.M.; Liu, D.R.; Aravind, L.; et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 2009, 324, 930–935. [Google Scholar] [CrossRef] [Green Version]
- He, Y.F.; Li, B.Z.; Li, Z.; Liu, P.; Wang, Y.; Tang, Q.; Ding, J.; Jia, Y.; Chen, Z.; Li, L.; et al. Tet-mediated formation of 5-carboxylcytosine and its excision by TDG in mammalian DNA. Science 2011, 333, 1303–1307. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Shen, L.; Dai, Q.; Wu, S.C.; Collins, L.B.; Swenberg, J.A.; He, C.; Zhang, Y. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science 2011, 333, 1300–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, K.P.; Yabuuchi, A.; Rao, S.; Huang, Y.; Cunniff, K.; Nardone, J.; Laiho, A.; Tahiliani, M.; Sommer, C.A.; Mostoslavsky, G.; et al. Tet1 and Tet2 regulate 5-hydroxymethylcytosine production and cell lineage specification in mouse embryonic stem cells. Cell Stem Cell 2011, 8, 200–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szwagierczak, A.; Bultmann, S.; Schmidt, C.S.; Spada, F.; Leonhardt, H. Sensitive enzymatic quantification of 5-hydroxymethylcytosine in genomic DNA. Nucleic Acids Res. 2010, 38, e181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, H.; Zhao, Y.; He, J.; Zhang, Y.; Xi, H.; Liu, M.; Ma, J.; Wu, L. YTHDF2 destabilizes m(6)A-containing RNA through direct recruitment of the CCR4-NOT deadenylase complex. Nat. Commun. 2016, 7, 12626. [Google Scholar] [CrossRef]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Ping, X.L.; Sun, B.F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Weng, H.; Su, R.; Weng, X.; Zuo, Z.; Li, C.; Huang, H.; Nachtergaele, S.; Dong, L.; Hu, C.; et al. FTO Plays an Oncogenic Role in Acute Myeloid Leukemia as a N(6)-Methyladenosine RNA Demethylase. Cancer Cell 2017, 31, 127–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, K.D.; Jaffrey, S.R. Rethinking m(6)A Readers, Writers, and Erasers. Annu. Rev. Cell Dev. Biol. 2017, 33, 319–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaccara, S.; Ries, R.J.; Jaffrey, S.R. Reading, writing and erasing mRNA methylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 608–624. [Google Scholar] [CrossRef] [PubMed]
- Dorn, L.E.; Lasman, L.; Chen, J.; Xu, X.; Hund, T.J.; Medvedovic, M.; Hanna, J.H.; van Berlo, J.H.; Accornero, F. The N(6)-Methyladenosine mRNA Methylase METTL3 Controls Cardiac Homeostasis and Hypertrophy. Circulation 2019, 139, 533–545. [Google Scholar] [CrossRef]
- Li, W.; Xing, C.; Bao, L.; Han, S.; Luo, T.; Wang, Z.; Fan, H. Comprehensive analysis of RNA m6A methylation in pressure overload-induced cardiac hypertrophy. BMC Genomics 2022, 23, 576. [Google Scholar] [CrossRef]
- Gong, R.; Wang, X.; Li, H.; Liu, S.; Jiang, Z.; Zhao, Y.; Yu, Y.; Han, Z.; Yu, Y.; Dong, C.; et al. Loss of m(6)A methyltransferase METTL3 promotes heart regeneration and repair after myocardial injury. Pharmacol. Res. 2021, 174, 105845. [Google Scholar] [CrossRef]
- Han, Z.; Wang, X.; Xu, Z.; Cao, Y.; Gong, R.; Yu, Y.; Yu, Y.; Guo, X.; Liu, S.; Yu, M.; et al. ALKBH5 regulates cardiomyocyte proliferation and heart regeneration by demethylating the mRNA of YTHDF1. Theranostics 2021, 11, 3000–3016. [Google Scholar] [CrossRef]
- Barroso-delJesus, A.; Romero-Lopez, C.; Lucena-Aguilar, G.; Melen, G.J.; Sanchez, L.; Ligero, G.; Berzal-Herranz, A.; Menendez, P. Embryonic stem cell-specific miR302-367 cluster: Human gene structure and functional characterization of its core promoter. Mol. Cell. Biol. 2008, 28, 6609–6619. [Google Scholar] [CrossRef] [Green Version]
- Card, D.A.; Hebbar, P.B.; Li, L.; Trotter, K.W.; Komatsu, Y.; Mishina, Y.; Archer, T.K. Oct4/Sox2-regulated miR-302 targets cyclin D1 in human embryonic stem cells. Mol. Cell. Biol. 2008, 28, 6426–6438. [Google Scholar] [CrossRef] [Green Version]
- Czaja, W.; Mao, P.; Smerdon, M.J. The emerging roles of ATP-dependent chromatin remodeling enzymes in nucleotide excision repair. Int. J. Mol. Sci. 2012, 13, 11954–11973. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.P.; Bruneau, B.G. Epigenetics and cardiovascular development. Annu. Rev. Physiol. 2012, 74, 41–68. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, A.; Willis, M.S.; Bultman, S.J. SWI/SNF chromatin-remodeling complexes in cardiovascular development and disease. Cardiovasc. Pathol. 2014, 23, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hang, C.T.; Yang, J.; Han, P.; Cheng, H.L.; Shang, C.; Ashley, E.; Zhou, B.; Chang, C.P. Chromatin regulation by Brg1 underlies heart muscle development and disease. Nature 2010, 466, 62–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, J.K.; Bruneau, B.G. Directed transdifferentiation of mouse mesoderm to heart tissue by defined factors. Nature 2009, 459, 708–711. [Google Scholar] [CrossRef] [Green Version]
- Pisano, A.; Cerbelli, B.; Perli, E.; Pelullo, M.; Bargelli, V.; Preziuso, C.; Mancini, M.; He, L.; Bates, M.G.; Lucena, J.R.; et al. Impaired mitochondrial biogenesis is a common feature to myocardial hypertrophy and end-stage ischemic heart failure. Cardiovasc. Pathol. 2016, 25, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Romanello, V.; Sandri, M. Mitochondrial Quality Control and Muscle Mass Maintenance. Front. Physiol. 2015, 6, 422. [Google Scholar] [CrossRef]
- Ehrlich, K.C.; Deng, H.W.; Ehrlich, M. Epigenetics of Mitochondria-Associated Genes in Striated Muscle. Epigenomes 2021, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xu, W.W.; Hu, S.J. Heart failure: Advanced development in genetics and epigenetics. BioMed Res. Int. 2015, 2015, 352734. [Google Scholar] [CrossRef] [Green Version]
- Masliah-Planchon, J.; Bieche, I.; Guinebretiere, J.M.; Bourdeaut, F.; Delattre, O. SWI/SNF chromatin remodeling and human malignancies. Annu. Rev. Pathol. 2015, 10, 145–171. [Google Scholar] [CrossRef]
- Shi, J.; Whyte, W.A.; Zepeda-Mendoza, C.J.; Milazzo, J.P.; Shen, C.; Roe, J.S.; Minder, J.L.; Mercan, F.; Wang, E.; Eckersley-Maslin, M.A.; et al. Role of SWI/SNF in acute leukemia maintenance and enhancer-mediated Myc regulation. Genes Dev. 2013, 27, 2648–2662. [Google Scholar] [CrossRef] [Green Version]
- Bultman, S.J.; Holley, D.W.; de Ridder, G.G.; Pizzo, S.V.; Sidorova, T.N.; Murray, K.T.; Jensen, B.C.; Wang, Z.; Bevilacqua, A.; Chen, X.; et al. BRG1 and BRM SWI/SNF ATPases redundantly maintain cardiomyocyte homeostasis by regulating cardiomyocyte mitophagy and mitochondrial dynamics in vivo. Cardiovasc. Pathol. 2016, 25, 258–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koczor, C.A.; Jiao, Z.; Fields, E.; Russ, R.; Ludaway, T.; Lewis, W. AZT-induced mitochondrial toxicity: An epigenetic paradigm for dysregulation of gene expression through mitochondrial oxidative stress. Physiol. Genom. 2015, 47, 447–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koczor, C.A.; Ludlow, I.; Fields, E.; Jiao, Z.; Ludaway, T.; Russ, R.; Lewis, W. Mitochondrial polymerase gamma dysfunction and aging cause cardiac nuclear DNA methylation changes. Physiol. Genom. 2016, 48, 274–280. [Google Scholar] [CrossRef] [Green Version]
- Matsuhashi, T.; Hishiki, T.; Zhou, H.; Ono, T.; Kaneda, R.; Iso, T.; Yamaguchi, A.; Endo, J.; Katsumata, Y.; Atsushi, A.; et al. Activation of pyruvate dehydrogenase by dichloroacetate has the potential to induce epigenetic remodeling in the heart. J. Mol. Cell Cardiol. 2015, 82, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Partanen, A.; Motoyama, J.; Hui, C.C. Developmentally regulated expression of the transcriptional cofactors/histone acetyltransferases CBP and p300 during mouse embryogenesis. Int. J. Dev. Biol. 1999, 43, 487–494. [Google Scholar] [PubMed]
- Hou, Y.S.; Wang, J.Z.; Shi, S.; Han, Y.; Zhang, Y.; Zhi, J.X.; Xu, C.; Li, F.F.; Wang, G.Y.; Liu, S.L. Identification of epigenetic factor KAT2B gene variants for possible roles in congenital heart diseases. Biosci. Rep. 2020, 40, 4. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Li, M.; Li, X.; Li, H.; Lai, Y.; Huang, S.; He, X.; Si, X.; Zheng, H.; Liao, W.; et al. Sirt1-inducible deacetylation of p21 promotes cardiomyocyte proliferation. Aging (Albany NY) 2019, 11, 12546–12567. [Google Scholar] [CrossRef]
- Kidder, B.L.; Hu, G.; Cui, K.; Zhao, K. SMYD5 regulates H4K20me3-marked heterochromatin to safeguard ES cell self-renewal and prevent spurious differentiation. Epigenetics Chromatin. 2017, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Boyer, L.A.; Plath, K.; Zeitlinger, J.; Brambrink, T.; Medeiros, L.A.; Lee, T.I.; Levine, S.S.; Wernig, M.; Tajonar, A.; Ray, M.K.; et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature 2006, 441, 349–353. [Google Scholar] [CrossRef]
- Pursani, V.; Bhartiya, D.; Tanavde, V.; Bashir, M.; Sampath, P. Transcriptional activator DOT1L putatively regulates human embryonic stem cell differentiation into the cardiac lineage. Stem Cell Res. Ther. 2018, 9, 97. [Google Scholar] [CrossRef] [Green Version]
- El-Nachef, D.; Oyama, K.; Wu, Y.Y.; Freeman, M.; Zhang, Y.; MacLellan, W.R. Repressive histone methylation regulates cardiac myocyte cell cycle exit. J. Mol. Cell Cardiol. 2018, 121, 1–12. [Google Scholar] [CrossRef]
- Brien, G.L.; Gambero, G.; O’connell, D.J.; Jerman, E.; Turner, S.A.; Egan, C.M.; Dunne, E.J.; Jurgens, M.C.; Wynne, K.; Piao, L.; et al. Polycomb PHF19 binds H3K36me3 and recruits PRC2 and demethylase NO66 to embryonic stem cell genes during differentiation. Nat. Struct. Mol. Biol. 2012, 19, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Rosales, W.; Lizcano, F. The Histone Demethylase JMJD2A Modulates the Induction of Hypertrophy Markers in iPSC-Derived Cardiomyocytes. Front. Genet. 2018, 9, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, E.; Mysliwiec, M.R.; Carlson, C.D.; Ansari, A.; Schwartz, R.J.; Lee, Y. Cardiac-specific developmental and epigenetic functions of Jarid2 during embryonic development. J. Biol. Chem. 2018, 293, 11659–11673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, Y.; Guo, C.; Lu, Q.; Wang, W.; Jia, Z.; Chen, P.; Ma, K.; Reinberg, D.; Zhou, C. ISL1 and JMJD3 synergistically control cardiac differentiation of embryonic stem cells. Nucleic Acids Res. 2016, 44, 6741–6755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Lu, J.; Li, J.; Wang, P.; Li, Z.; Zhong, Y.; Guo, K.; Wang, J.; Ye, J.; Liu, P. JMJD3 inhibition protects against isoproterenol-induced cardiac hypertrophy by suppressing beta-MHC expression. Mol. Cell Endocrinol. 2018, 477, 1–14. [Google Scholar] [CrossRef]
- Tang, Y.; Hong, Y.Z.; Bai, H.J.; Wu, Q.; Chen, C.D.; Lang, J.Y.; Boheler, K.R.; Yang, H.T. Plant Homeo Domain Finger Protein 8 Regulates Mesodermal and Cardiac Differentiation of Embryonic Stem Cells Through Mediating the Histone Demethylation of pmaip1. Stem Cells. 2016, 34, 1527–1540. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.Q.; Papp, G.; Szodoray, P.; Zeher, M. The role of microRNAs in the pathogenesis of autoimmune diseases. Autoimmun. Rev. 2016, 15, 1171–1180. [Google Scholar] [CrossRef] [Green Version]
- Deb, B.; Uddin, A.; Chakraborty, S. miRNAs and ovarian cancer: An overview. J. Cell Physiol. 2018, 233, 3846–3854. [Google Scholar] [CrossRef]
- Li, X.; Wei, Y.; Wang, Z. microRNA-21 and hypertension. Hypertens. Res. 2018, 41, 649–661. [Google Scholar] [CrossRef]
- Cirillo, F.; Catellani, C.; Lazzeroni, P.; Sartori, C.; Street, M.E. The Role of MicroRNAs in Influencing Body Growth and Development. Horm. Res. Paediatr. 2020, 93, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Kanellopoulou, C.; Muljo, S.A.; Kung, A.L.; Ganesan, S.; Drapkin, R.; Jenuwein, T.; Livingston, D.M.; Rajewsky, K. Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev. 2005, 19, 489–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murchison, E.P.; Partridge, J.F.; Tam, O.H.; Cheloufi, S.; Hannon, G.J. Characterization of Dicer-deficient murine embryonic stem cells. Proc. Natl. Acad. Sci. USA 2005, 102, 12135–12140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Medvid, R.; Melton, C.; Jaenisch, R.; Blelloch, R. DGCR8 is essential for microRNA biogenesis and silencing of embryonic stem cell self-renewal. Nat. Genet. 2007, 39, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Mallanna, S.K.; Rizzino, A. Emerging roles of microRNAs in the control of embryonic stem cells and the generation of induced pluripotent stem cells. Dev. Biol. 2010, 344, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, E.M.; Olson, E.N. Pervasive roles of microRNAs in cardiovascular biology. Nature 2011, 469, 336–342. [Google Scholar] [CrossRef] [Green Version]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of tissue-specific microRNAs from mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Samal, E.; Srivastava, D. Serum response factor regulates a muscle-specific microRNA that targets Hand2 during cardiogenesis. Nature 2005, 436, 214–220. [Google Scholar] [CrossRef]
- Sayed, D.; Hong, C.; Chen, I.Y.; Lypowy, J.; Abdellatif, M. MicroRNAs play an essential role in the development of cardiac hypertrophy. Circ. Res. 2007, 100, 416–424. [Google Scholar] [CrossRef] [Green Version]
- Porrello, E.R. microRNAs in cardiac development and regeneration. Clin. Sci. 2013, 125, 151–166. [Google Scholar] [CrossRef] [Green Version]
- Izarra, A.; Moscoso, I.; Canon, S.; Carreiro, C.; Fondevila, D.; Martin-Caballero, J.; Blanca, V.; Valiente, I.; Diez-Juan, A.; Bernad, A. miRNA-1 and miRNA-133a are involved in early commitment of pluripotent stem cells and demonstrate antagonistic roles in the regulation of cardiac differentiation. J. Tissue Eng. Regen. Med. 2017, 11, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Dal-Pra, S.; Hodgkinson, C.P.; Mirotsou, M.; Kirste, I.; Dzau, V.J. Demethylation of H3K27 is essential for the induction of direct cardiac reprogramming by miR combo. Circ. Res. 2017, 120, 1403–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Sanchez, C.; Franco, D.; Bonet, F.; Garcia-Lopez, V.; Aranega, A.; Garcia-Martinez, V. Reciprocal repression between Fgf8 and miR-133 regulates cardiac induction through Bmp2 signaling. Data Brief 2015, 5, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoelscher, S.C.; Stich, T.; Diehm, A.; Lahm, H.; Dressen, M.; Zhang, Z.; Neb, I.; Aherrahrou, Z.; Erdmann, J.; Schunkert, H.; et al. miR-128a Acts as a Regulator in Cardiac Development by Modulating Differentiation of Cardiac Progenitor Cell Populations. Int. J. Mol. Sci. 2020, 21, 1158. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.H.; Zhao, H.; Zhou, L.P.; Zhao, C.X.; Wu, Y.F.; Zhen, L.X.; Li, J.; Ge, D.X.; Xu, L.; Lin, L.; et al. miR-134 Modulates the Proliferation of Human Cardiomyocyte Progenitor Cells by Targeting Meis2. Int. J. Mol. Sci. 2015, 16, 25199–25213. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Xiao, J.; Zhu, H.; Wei, X.; Platt, C.; Damilano, F.; Xiao, C.; Bezzerides, V.; Bostrom, P.; Che, L.; et al. miR-222 is necessary for exercise-induced cardiac growth and protects against pathological cardiac remodeling. Cell Metab. 2015, 21, 584–595. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Chen, J.H.; Lundy, D.J.; Liu, C.H.; Hwang, S.M.; Pabon, L.; Shieh, R.C.; Chen, C.C.; Wu, S.N.; Yan, Y.T.; et al. Defined MicroRNAs Induce Aspects of Maturation in Mouse and Human Embryonic-Stem-Cell-Derived Cardiomyocytes. Cell Rep. 2015, 12, 1960–1967. [Google Scholar] [CrossRef] [Green Version]
- Borden, A.; Kurian, J.; Nickoloff, E.; Yang, Y.; Troupes, C.D.; Ibetti, J.; Lucchese, A.M.; Gao, E.; Mohsin, S.; Koch, W.J.; et al. Transient Introduction of miR-294 in the Heart Promotes Cardiomyocyte Cell Cycle Reentry After Injury. Circ. Res. 2019, 125, 14–25. [Google Scholar] [CrossRef]
- Xiao, J.; Liu, H.; Cretoiu, D.; Toader, D.O.; Suciu, N.; Shi, J.; Shen, S.; Bei, Y.; Sluijter, J.P.; Das, S.; et al. miR-31a-5p promotes postnatal cardiomyocyte proliferation by targeting RhoBTB1. Exp. Mol. Med. 2017, 49, e386. [Google Scholar] [CrossRef] [Green Version]
- Poon, E.N.; Hao, B.; Guan, D.; Jun Li, M.; Lu, J.; Yang, Y.; Wu, B.; Wu, S.C.; Webb, S.E.; Liang, Y.; et al. Integrated transcriptomic and regulatory network analyses identify microRNA-200c as a novel repressor of human pluripotent stem cell-derived cardiomyocyte differentiation and maturation. Cardiovasc. Res. 2018, 114, 894–906. [Google Scholar] [CrossRef]
- Zhuang, S.; Fu, Y.; Li, J.; Li, M.; Hu, X.; Zhu, J.; Tong, M. MicroRNA-375 overexpression disrupts cardiac development of Zebrafish (Danio rerio) by targeting notch2. Protoplasma 2020, 257, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xu, M.; Li, M.; Wu, F.; Hu, S.; Chen, X.; Zhao, L.; Huang, Z.; Lan, F.; Liu, D.; et al. miR-25 Promotes Cardiomyocyte Proliferation by Targeting FBXW7. Mol. Ther. -Nucleic Acids 2020, 19, 1299–1308. [Google Scholar] [CrossRef]
- Yu, R.B.; Li, K.; Wang, G.; Gao, G.M.; Du, J.X. MiR-23 enhances cardiac fibroblast proliferation and suppresses fibroblast apoptosis via targeting TGF-beta1 in atrial fibrillation. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4419–4424. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.; Rajakaruna, C.; Caputo, M.; Emanueli, C. MicroRNAs in congenital heart disease. Ann. Transl. Med. 2015, 3, 333. [Google Scholar] [CrossRef]
- Zhao, Y.; Ransom, J.F.; Li, A.; Vedantham, V.; von Drehle, M.; Muth, A.N.; Tsuchihashi, T.; McManus, M.T.; Schwartz, R.J.; Srivastava, D. Dysregulation of cardiogenesis, cardiac conduction, and cell cycle in mice lacking miRNA-1-2. Cell 2007, 129, 303–317. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Yang, Q.; Mi, Y.; Wang, F.; Cai, K.; Zhang, Y.; Wang, Y.; Wang, X.; Gui, Y.; Li, Q. miR-29b-3p Inhibitor Alleviates Hypomethylation-Related Aberrations Through a Feedback Loop Between miR-29b-3p and DNA Methylation in Cardiomyocytes. Front. Cell Dev. Biol. 2022, 10, 788799. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, V.; Bhagavatheeswaran, S.; Shanmugam, S.; Vasudevan, M.; Ragunathan, M.; Cherian, K.M.; Munirajan, A.K.; Ravi, S.; Balakrishnan, A. Deep sequencing unveils altered cardiac miRNome in congenital heart disease. Mol. Genet. Genom. 2022, 297, 1123–1139. [Google Scholar] [CrossRef]
- Kan, Z.; Yan, W.; Wang, N.; Fang, Y.; Gao, H.; Song, Y. Identification of circRNA-miRNA-mRNA Regulatory Network and Crucial Signaling Pathway Axis Involved in Tetralogy of Fallot. Front. Genet. 2022, 13, 917454. [Google Scholar] [CrossRef]
- Yang, P.; Yang, Y.; He, X.; Sun, P.; Zhang, Y.; Song, X.; Tian, Y.; Zong, T.; Ma, J.; Chen, X.; et al. miR-153-3p Targets betaII Spectrin to Regulate Formaldehyde-Induced Cardiomyocyte Apoptosis. Front. Cardiovasc. Med. 2021, 8, 764831. [Google Scholar] [CrossRef]
- Meng, S.; Hu, Y.; Zhu, J.; Feng, T.; Quan, X. miR-30c-5p acts as a therapeutic target for ameliorating myocardial ischemia-reperfusion injury. Am. J. Transl. Res. 2021, 13, 2198–2212. [Google Scholar]
- Li, Y.; Li, X.; Wang, L.; Han, N.; Yin, G. miR-375-3p contributes to hypoxia-induced apoptosis by targeting forkhead box P1 (FOXP1) and Bcl2 like protein 2 (Bcl2l2) in rat cardiomyocyte h9c2 cells. Biotechnol. Lett. 2021, 43, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Ceci, M.; Carlantoni, C.; Missinato, M.A.; Bonvissuto, D.; Di Giacomo, B.; Contu, R.; Romano, N. Micro RNAs are involved in activation of epicardium during zebrafish heart regeneration. Cell Death Discov. 2018, 4, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, J.; Tang, F.M.K.; Su, X.; Lu, G.; Xu, J.; Lee, H.S.S.; Lee, K.K.H. microRNA-1 inhibits cardiomyocyte proliferation in mouse neonatal hearts by repressing CCND1 expression. Ann. Transl. Med. 2019, 7, 455. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Wang, Q.; Gao, N.; Wu, F.; Lan, F.; Zhang, F.; Jin, L.; Huang, Z.; Ge, J.; Wang, H.; et al. MircroRNA-10b Promotes Human Embryonic Stem Cell-Derived Cardiomyocyte Proliferation via Novel Target Gene LATS1. Mol. Ther. -Nucleic Acids 2020, 19, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.L.; Zhang, S.; Tellam, R.L.; Brooks, D.A.; McMillen, I.C.; Porrello, E.R.; Botting, K.J. Regulation of microRNA during cardiomyocyte maturation in sheep. BMC Genom. 2015, 16, 541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Ursin, R.; Mahapatra, S.; Gallicano, G.I. CRISPR/CAS9 ablation of individual miRNAs from a miRNA family reveals their individual efficacies for regulating cardiac differentiation. Mech. Dev. 2018, 150, 10–20. [Google Scholar] [CrossRef]
- Guzzolino, E.; Chiavacci, E.; Ahuja, N.; Mariani, L.; Evangelista, M.; Ippolito, C.; Rizzo, M.; Garrity, D.; Cremisi, F.; Pitto, L. Post-transcriptional Modulation of Sphingosine-1-Phosphate Receptor 1 by miR-19a Affects Cardiovascular Development in Zebrafish. Front. Cell Dev. Biol. 2018, 6, 58. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Kataoka, M.; Liu, N.; Liang, T.; Huang, Z.P.; Gu, F.; Ding, J.; Liu, J.; Zhang, F.; Ma, Q.; et al. Therapeutic role of miR-19a/19b in cardiac regeneration and protection from myocardial infarction. Nat. Commun. 2019, 10, 1802. [Google Scholar] [CrossRef] [Green Version]
- Ai, F.; Zhang, Y.; Peng, B. miR-20a regulates proliferation, differentiation and apoptosis in P19 cell model of cardiac differentiation by targeting Smoothened. Biol. Open 2016, 5, 1260–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Gao, S.; Yang, Y.; Wu, L.; Wang, L. microRNA-25 promotes cardiomyocytes proliferation and migration via targeting Bim. J. Cell. Physiol. 2019, 234, 22103–22115. [Google Scholar] [CrossRef]
- Wang, D.; Liu, C.; Wang, Y.; Wang, W.; Wang, K.; Wu, X.; Li, Z.; Zhao, C.; Li, L.; Peng, L. Impact of miR-26b on cardiomyocyte differentiation in P19 cells through regulating canonical/non-canonical Wnt signalling. Cell Prolif. 2017, 50, e12371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Lu, H.; Chen, R.; Zhu, L.; Song, G. MicroRNA-29c affects zebrafish cardiac development via targeting Wnt4. Mol. Med. Rep. 2020, 22, 4675–4684. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wu, F.; Mi, Y.; Wang, F.; Cai, K.; Yang, X.; Zhang, R.; Liu, L.; Zhang, Y.; Wang, Y.; et al. Aberrant expression of miR-29b-3p influences heart development and cardiomyocyte proliferation by targeting NOTCH2. Cell Prolif. 2020, 53, e12764. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, M.; Peng, Y.; Hu, X.; Xu, J.; Zhu, S.; Yu, Z.; Han, S. miR-30c regulates proliferation, apoptosis and differentiation via the Shh signaling pathway in P19 cells. Exp. Mol. Med. 2016, 48, e248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhang, Y.; Zhu, H.; Hu, J.; Xie, Z. MiR-34a/miR-93 target c-Ski to modulate the proliferaton of rat cardiac fibroblasts and extracellular matrix deposition in vivo and in vitro. Cell. Signal. 2018, 46, 145–153. [Google Scholar] [CrossRef]
- Wu, K.H.; Xiao, Q.R.; Yang, Y.; Xu, J.L.; Zhang, F.; Liu, C.M.; Zhang, Z.M.; Lu, Y.Q.; Huang, N.P. MicroRNA-34a modulates the Notch signaling pathway in mice with congenital heart disease and its role in heart development. J. Mol. Cell. Cardiol. 2018, 114, 300–308. [Google Scholar] [CrossRef]
- Beauchemin, M.; Smith, A.; Yin, V.P. Dynamic microRNA-101a and Fosab expression controls zebrafish heart regeneration. Development 2015, 142, 4026–4037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, R.; Zhang, X.; Fang, Y.; Nie, Y.; Cai, S.; Chen, Y.; Mo, D. mir-127-3p inhibits the proliferation of myocytes by targeting KMT5a. Biochem. Biophys. Res. Commun. 2018, 503, 970–976. [Google Scholar] [CrossRef]
- Huang, W.; Feng, Y.; Liang, J.; Yu, H.; Wang, C.; Wang, B.; Wang, M.; Jiang, L.; Meng, W.; Cai, W.; et al. Loss of microRNA-128 promotes cardiomyocyte proliferation and heart regeneration. Nat. Commun. 2018, 9, 700. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Jiang, B.; Xia, Y.; Wang, J.; Ji, C.; Tong, J.; Chen, T.; Jiang, Y. Downregulation of miR-133a contributes to the cardiac developmental toxicity of trichloroethylene in zebrafish. Chemosphere 2020, 251, 126610. [Google Scholar] [CrossRef]
- Wojciechowska, A.; Braniewska, A.; Kozar-Kaminska, K. MicroRNA in cardiovascular biology and disease. Adv. Clin. Exp. Med. 2017, 26, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Bostjancic, E.; Zidar, N.; Stajer, D.; Glavac, D. MicroRNAs miR-1, miR-133a, miR-133b and miR-208 are dysregulated in human myocardial infarction. Cardiology 2010, 115, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Shen, H.; Huang, Q.; Li, Q. The Circular RNA CDR1as Regulates the Proliferation and Apoptosis of Human Cardiomyocytes Through the miR-135a/HMOX1 and miR-135b/HMOX1 Axes. Genet. Test. Mol. Biomarkers 2020, 24, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.-L.; Zhu, B.-L.; Sun, Y.-Y.; Qiu, G.-R.; Fu, W.-N.; Jiang, H.-K. MicroRNA-144 regulates cardiomyocyte proliferation and apoptosis by targeting TBX1 through the JAK2/STAT1 pathway. Cytogenet. Genome Res. 2019, 159, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Ling, X.; Yao, D.; Kang, L.; Zhou, J.; Zhou, Y.; Dong, H.; Zhang, K.; Zhang, L.; Chen, H. Involment of RAS/ERK1/2 signaling and MEF2C in miR-155-3p inhibition-triggered cardiomyocyte differentiation of embryonic stem cell. Oncotarget 2017, 8, 84403–84416. [Google Scholar] [CrossRef] [Green Version]
- Guzzolino, E.; Pellegrino, M.; Ahuja, N.; Garrity, D.; D’Aurizio, R.; Groth, M.; Baumgart, M.; Hatcher, C.J.; Mercatanti, A.; Evangelista, M.; et al. miR-182-5p is an evolutionarily conserved Tbx5 effector that impacts cardiac development and electrical activity in zebrafish. Cell Mol. Life Sci. 2020, 77, 3215–3229. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Y.; Wang, X.; Guo, X.; Lu, C.; Kang, J.; Wang, G. MiR-184 directly targets Wnt3 in cardiac mesoderm differentiation of embryonic stem cells. Stem Cells 2020, 38, 1568–1577. [Google Scholar] [CrossRef]
- Tao, Y.; Zhang, H.; Huang, S.; Pei, L.; Feng, M.; Zhao, X.; Ouyang, Z.; Yao, S.; Jiang, R.; Wei, K. miR-199a-3p promotes cardiomyocyte proliferation by inhibiting Cd151 expression. Biochem. Biophys. Res. Commun. 2019, 516, 28–36. [Google Scholar] [CrossRef]
- Chen, H.P.; Wen, J.; Tan, S.R.; Kang, L.M.; Zhu, G.C. MiR-199a-3p inhibition facilitates cardiomyocyte differentiation of embryonic stem cell through promotion of MEF2C. J. Cell. Physiol. 2019, 234, 23315–23325. [Google Scholar] [CrossRef]
- Liang, D.; Li, J.; Wu, Y.; Zhen, L.; Li, C.; Qi, M.; Wang, L.; Deng, F.; Huang, J.; Lv, F.; et al. miRNA-204 drives cardiomyocyte proliferation via targeting Jarid2. Int. J. Cardiol. 2015, 201, 38–48. [Google Scholar] [CrossRef]
- Tian, Y.; Liu, Y.; Wang, T.; Zhou, N.; Kong, J.; Chen, L.; Snitow, M.; Morley, M.; Li, D.; Petrenko, N.; et al. A microRNA-Hippo pathway that promotes cardiomyocyte proliferation and cardiac regeneration in mice. Sci. Transl. Med. 2015, 7, 279ra238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, D.; Wang, X.; Wang, X.; Thomsen, B.; Yang, Y.; Lan, X.; Huang, Y.; Chen, H. MicroRNA bta-miR-365-3p inhibits proliferation but promotes differentiation of primary bovine myoblasts by targeting the activin A receptor type I. J. Anim. Sci. Biotechnol. 2021, 12, 16. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zhang, Y.; Tang, Y.; Dai, H.; OuYang, Y.; Li, C.; Yu, M. hsa-miR-4443 inhibits myocardial fibroblast proliferation by targeting THBS1 to regulate TGF-beta1/alpha-SMA/collagen signaling in atrial fibrillation. Braz. J. Med. Biol. Res. 2021, 54, e10692. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhang, P.; Zhang, L. Fetal Hypoxia Impacts on Proliferation and Differentiation of Sca-1+ Cardiac Progenitor Cells and Maturation of Cardiomyocytes: A Role of MicroRNA-210. Genes 2020, 11, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasanaro, P.; D’Alessandra, Y.; Di Stefano, V.; Melchionna, R.; Romani, S.; Pompilio, G.; Capogrossi, M.C.; Martelli, F. MicroRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand Ephrin-A3. J. Biol. Chem. 2008, 283, 15878–15883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, J.; Yang, J.; Liu, Q.; Gong, Y.; Zhang, Y.; Zhang, Z. Selenium deficiency inhibits myocardial development and differentiation by targeting the mir-215-5p/CTCF axis in chicken. Metallomics 2019, 11, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, J.; Cui, J.; Sun, M.; Du, W.; Chen, T.; Ming, X.; Zhang, L.; Tian, J.; Li, J.; et al. MiR218 Modulates Wnt Signaling in Mouse Cardiac Stem Cells by Promoting Proliferation and Inhibiting Differentiation through a Positive Feedback Loop. Sci. Rep. 2016, 6, 20968. [Google Scholar] [CrossRef] [Green Version]
- Cruz, F.M.; Tome, M.; Bernal, J.A.; Bernad, A. miR-300 mediates Bmi1 function and regulates differentiation in primitive cardiac progenitors. Cell Death Dis. 2015, 6, e1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Soibam, B.; Benham, A.; Xu, X.; Chopra, M.; Peng, X.; Yu, W.; Bao, W.; Liang, R.; Azares, A.; et al. miR-322/-503 cluster is expressed in the earliest cardiac progenitor cells and drives cardiomyocyte specification. Proc. Natl. Acad. Sci. USA 2016, 113, 9551–9556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Song, G.; Liu, M.; Chen, B.; Chen, Y.; Shen, Y.; Zhu, J.; Zhou, X. MicroRNA-375 overexpression influences P19 cell proliferation, apoptosis and differentiation through the Notch signaling pathway. Int. J. Mol. Med. 2016, 37, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.L.; Naya, F.J. MicroRNAs in the Myocyte Enhancer Factor 2 (MEF2)-regulated Gtl2-Dio3 Noncoding RNA Locus Promote Cardiomyocyte Proliferation by Targeting the Transcriptional Coactivator Cited2. J. Biol. Chem. 2015, 290, 23162–23172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Qi, J. miR-430a regulates the development of left-right asymmetry by targeting sqt in the teleost. Gene 2020, 745, 144628. [Google Scholar] [CrossRef]
- Pisano, F.; Altomare, C.; Cervio, E.; Barile, L.; Rocchetti, M.; Ciuffreda, M.C.; Malpasso, G.; Copes, F.; Mura, M.; Danieli, P.; et al. Combination of miRNA499 and miRNA133 exerts a synergic effect on cardiac differentiation. Stem Cells 2015, 33, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Neshati, V.; Mollazadeh, S.; Fazly Bazzaz, B.S.; de Vries, A.A.F.; Mojarrad, M.; Naderi-Meshkin, H.; Neshati, Z.; Mirahmadi, M.; Kerachian, M.A. MicroRNA-499a-5p Promotes Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells to Cardiomyocytes. Appl. Biochem. Biotechnol. 2018, 186, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, Z.; Yang, F.; Huang, J.; Hu, X.; Deng, S.; Tian, M.; Si, X. miR449a5p suppresses CDK6 expression to inhibit cardiomyocyte proliferation. Mol. Med. Rep. 2021, 23, 1. [Google Scholar] [CrossRef]
- Del Campo, C.V.; Liaw, N.Y.; Gunadasa-Rohling, M.; Matthaei, M.; Braga, L.; Kennedy, T.; Salinas, G.; Voigt, N.; Giacca, M.; Zimmermann, W.H.; et al. Regenerative potential of epicardium-derived extracellular vesicles mediated by conserved miRNA transfer. Cardiovasc. Res. 2022, 118, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, C.; Zhang, J.; Jiao, Z.; Dong, N.; Wang, G.; Wang, Z.; Wang, L. Localized injection of miRNA-21-enriched extracellular vesicles effectively restores cardiac function after myocardial infarction. Theranostics 2019, 9, 2346–2360. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Ma, R.; Cai, W.; Huang, W.; Paul, C.; Liang, J.; Wang, Y.; Zhao, T.; Kim, H.W.; Xu, M.; et al. Exosomes Secreted from CXCR4 Overexpressing Mesenchymal Stem Cells Promote Cardioprotection via Akt Signaling Pathway following Myocardial Infarction. Stem Cells Int. 2015, 2015, 659890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, R.C.; Arslan, F.; Lee, M.M.; Sze, N.S.; Choo, A.; Chen, T.S.; Salto-Tellez, M.; Timmers, L.; Lee, C.N.; El Oakley, R.M.; et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010, 4, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Shao, L.; Zhang, Y.; Lan, B.; Wang, J.; Zhang, Z.; Zhang, L.; Xiao, P.; Meng, Q.; Geng, Y.J.; Yu, X.Y.; et al. MiRNA-Sequence Indicates That Mesenchymal Stem Cells and Exosomes Have Similar Mechanism to Enhance Cardiac Repair. BioMed Res. Int. 2017, 2017, 4150705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Li, L.; Liu, T.; Jiang, W.; Hu, X. miR-19a/19b-loaded exosomes in combination with mesenchymal stem cell transplantation in a preclinical model of myocardial infarction. Regen. Med. 2020, 15, 1749–1759. [Google Scholar] [CrossRef] [PubMed]
- Huh, D.; Kim, H.J.; Fraser, J.P.; Shea, D.E.; Khan, M.; Bahinski, A.; Hamilton, G.A.; Ingber, D.E. Microfabrication of human organs-on-chips. Nat. Protoc. 2013, 8, 2135–2157. [Google Scholar] [CrossRef]
- Tandon, N.; Cannizzaro, C.; Chao, P.H.G.; Maidhof, R.; Marsano, A.; Au, H.T.H.; Radisic, M.; Vunjak-Novakovic, G. Electrical stimulation systems for cardiac tissue engineering. Nat. Protoc. 2009, 4, 155–173. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Jia, X.; Bai, K.; Gong, X.; Fan, Y. Effect of fluid shear stress on cardiomyogenic differentiation of rat bone marrow mesenchymal stem cells. Arch. Med. Res. 2010, 41, 497–505. [Google Scholar] [CrossRef]
- Toh, Y.C.; Voldman, J. Fluid shear stress primes mouse embryonic stem cells for differentiation in a self-renewing environment via heparan sulfate proteoglycans transduction. FASEB J. 2011, 25, 1208–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, F.; Ping, Z.; Handong, Y.; Zhengtong, Y.; Zhaodong, Z. Induced differentiation of human cord blood mesenchymal stem/progenitor cells into cardiomyocyte-like cells in vitro. J. Huazhong. Univ. Sci. Technolog. Med. Sci. 2003, 23, 154–157. [Google Scholar] [PubMed]
- Pavesi, A.; Adriani, G.; Rasponi, M.; Zervantonakis, I.K.; Fiore, G.B.; Kamm, R.D. Controlled electromechanical cell stimulation on-a-chip. Sci. Rep. 2015, 5, 11800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, S.M.; Jones, P.A. Multiple new phenotypes induced in 10T1/2 and 3T3 cells treated with 5-azacytidine. Cell 1979, 17, 771–779. [Google Scholar] [CrossRef]
- Sokolowska, P.; Zukowski, K.; Lasocka, I.; Szulc-Dabrowska, L.; Jastrzebska, E. Human mesenchymal stem cell (hMSC) differentiation towards cardiac cells using a new microbioanalytical method. Analyst 2020, 145, 3017–3028. [Google Scholar] [CrossRef]
- Kakkar, A.; Nandy, S.B.; Gupta, S.; Bharagava, B.; Airan, B.; Mohanty, S. Adipose tissue derived mesenchymal stem cells are better respondents to TGFbeta1 for in vitro generation of cardiomyocyte-like cells. Mol. Cell Biochem. 2019, 460, 53–66. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ahn, Y.; Kwon, J.S.; Cho, Y.K.; Jeong, M.H.; Cho, J.G.; Park, J.C.; Kang, J.C. Priming of mesenchymal stem cells with oxytocin enhances the cardiac repair in ischemia/reperfusion injury. Cells Tissues Organs. 2012, 195, 428–442. [Google Scholar] [CrossRef] [PubMed]
- Li, T.S.; Komota, T.; Ohshima, M.; Qin, S.L.; Kubo, M.; Ueda, K.; Hamano, K. TGF-beta induces the differentiation of bone marrow stem cells into immature cardiomyocytes. Biochem. Biophys. Res. Commun. 2008, 366, 1074–1080. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Li, F.; Zhao, X.; Ma, Y.; Li, Y.; Lin, M.; Jin, G.; Lu, T.J.; Genin, G.M.; Xu, F. Functional and Biomimetic Materials for Engineering of the Three-Dimensional Cell Microenvironment. Chem. Rev. 2017, 117, 12764–12850. [Google Scholar] [CrossRef] [PubMed]


| MicroRNAs | Effect(s) on Gene | Affected Gene | Gene Function | References |
|---|---|---|---|---|
| miR-1 | Activation | DELTA | Differentiation | [221,223,224,244,245] |
| Inhibition | CDK9 | Inhibition of differentiation | ||
| Inhibition | GATA4 | Proliferation | ||
| Inhibition | HAND2 | Proliferation | ||
| Inhibition | HSP70 | Proliferation | ||
| Inhibition | RALDH2 | Proliferation | ||
| Inhibition | IGF2B | Proliferation | ||
| Inhibition | IGF1 | Proliferation | ||
| Inhibition | FRS2 | Proliferation | ||
| Inhibition | FOXP1 | Proliferation | ||
| Inhibition | HAND2 | Inhibition of differentiation | ||
| Inhibition | HDAC4 | Inhibition of differentiation | ||
| Activation | NKX2.5 | Differentiation | ||
| Inhibition | CCND1 | Proliferation | ||
| Inhibition | NANOG | Inhibition of differentiation | ||
| Inhibition | OCT4 | Inhibition of differentiation | ||
| Activation | TROPT | Differentiation | ||
| Activation | A-ACTININ | Differentiation | ||
| Activation | KDM6A | Differentiation | ||
| Activation | KDM6B | Differentiation | ||
| Activation | TBX5 | Differentiation | ||
| Activation | MEF2C | Differentiation | ||
| Activation | GATA4 | Differentiation | ||
| miR-10b | Inhibition | LATS1 | Inhibition of proliferation | [246] |
| miR-15 | Inhibition | CHEK1 | Proliferation | [247] |
| Inhibition | CDC2A | Proliferation | ||
| Inhibition | BIRC5 | Proliferation | ||
| Inhibition | SPAG5 | Proliferation | ||
| miR-17 | Inhibition | FOG2 | Inhibition of differentiation | [248] |
| miR-19a | Inhibition | S1PR1 | Inhibition of development | [249,250] |
| Inhibition | BIM | Inhibition of proliferation | ||
| Inhibition | PTEN | Inhibition of proliferation | ||
| Activation | CCNB1 | Proliferation | ||
| Activation | CCND1 | Proliferation | ||
| Activation | CDK1 | Proliferation | ||
| miR-19b | Inhibition | WNT | Differentiation | [228,250] |
| Inhibition | CTNNB | Differentiation | ||
| Inhibition | PTEN | Inhibition of proliferation | ||
| Inhibition | BIM | Inhibition of proliferation | ||
| Activation | CCNB1 | Proliferation | ||
| Activation | CCND1 | Proliferation | ||
| Activation | CDK1 | Proliferation | ||
| miR-20a | Inhibition | SMO | Proliferation Differentiation | [251] |
| MiR-23 | Activation | TGF-Β1 | Proliferation | [235] |
| miR-25 | Inhibition | FBXW7 | Inhibition of proliferation | [234,252] |
| Inhibition | BIM | Inhibition of proliferation | ||
| miR-26b | Inhibition | WNT | Differentiation | [253] |
| miR-29c | Inhibition | WNT4 | Development | [254] |
| miR-29b-3p | Inhibition | NOTCH2 | Proliferation | [255] |
| Development | ||||
| miR-30c | Inhibition | GLI2 | Inhibition of proliferation | [256] |
| Differentiation | ||||
| miR-31a-5p | Inhibition | RHOBTB1 | Inhibition of proliferation | [231] |
| miR-34a | Inhibition | C-SKI | Inhibition of proliferation | [257,258] |
| Inhibition | NOTCH1 | Development | ||
| miR-93 | Inhibition | FOG2 | Inhibition of differentiation | [248,257] |
| Inhibition | C-SKI | Inhibition of proliferation | ||
| miR-101a | Activation | FOSAB | Proliferation | [259] |
| miR-106a | Inhibition | FOG2 | Inhibition of differentiation | [248] |
| miR-125b-5p | Inhibition | ERBB4 | Inhibition of development | [229] |
| mir-127-3p | Inhibition | KMT5A | Proliferation | [260] |
| miR-128 | Inhibition | SUZ12 | Proliferation | [261] |
| Regeneration | ||||
| miR-128a | Inhibition | ISL1 | Development | [226] |
| Inhibition | SFRP5 | Development | ||
| Inhibition | HCN4 | Development | ||
| Inhibition | NKX2.5 | Development | ||
| Inhibition | MEF2C | Development | ||
| Inhibition | MYL2 | Development | ||
| Activation | IRX4 | Inhibition of development | ||
| miR-133 | Activation | BMP2 | Differentiation | [224,225] |
| Activation | GATA4 | Differentiation | ||
| Activation | NKX2.5 | Differentiation | ||
| Inhibition | FGF8 | Inhibition of differentiation | ||
| Activation | KDM6A | Differentiation | ||
| Activation | KDM6B | Differentiation | ||
| Activation | TBX5 | Differentiation | ||
| Activation | MEF2C | Differentiation | ||
| Activation | CX43 | Differentiation | ||
| Activation | CTNT | Differentiation | ||
| miR-133a | Inhibition | NKX2.5 | Development | [223,244,247,262,263,264] |
| Inhibition | PGAM1 | Development | ||
| Inhibition | GJA1 | Development | ||
| Inhibition | SRF | Development | ||
| Inhibition | CCND2 | Development | ||
| Inhibition | IGF1R | Proliferation | ||
| Inhibition | HAND2 | Development | ||
| Inhibition | A-ACTININ | Development | ||
| Inhibition | CTNT | Development | ||
| Inhibition | VEGF | Proliferation | ||
| Inhibition | CTGF | Proliferation | ||
| Inhibition | FGFR1 | Proliferation | ||
| Inhibition | PP2AC | Proliferation | ||
| Inhibition | TBX18 | Proliferation | ||
| miR-134 | Inhibition | MEIS2 | Proliferation | [227] |
| Inhibition | CYCLIN A | Proliferation | ||
| Inhibition | CYCLIN B | Proliferation | ||
| Inhibition | CYCLIN E | Proliferation | ||
| Inhibition | CDC2 | Proliferation | ||
| Inhibition | CDK4 | Proliferation | ||
| Inhibition | PCNA | Proliferation | ||
| miR-135a | Inhibition | CDR1 | Proliferation | [265] |
| miR-135b | Inhibition | CDR1 | Proliferation | [265] |
| miR-144 | Activation | TBX1 | Proliferation | [266] |
| miR-155-3p | Inhibition | MEF2C | Differentiation | [267] |
| Inhibition | CTNT | Development | ||
| Inhibition | GATA4 | Development | ||
| Inhibition | NKX2.5 | Development | ||
| miR-182-5p | Inhibition | CACNB2.2 | Development | [268] |
| Inhibition | CACNB4B | Development | ||
| Inhibition | CACNA2D1 | Development | ||
| miR-184 | Inhibition | WNT3 | Differentiation | [269] |
| miR-199a | Activation | CLIC5 | Inhibition of proliferation | [247] |
| Activation | HOMER1 | Inhibition of proliferation | ||
| miR-199a-3p | Inhibition | CD151 | Inhibition of proliferation | [270] |
| Inhibition | MEF2C | Differentiation | [271] | |
| miR-199a-5p | Inhibition | ERBB4 | Inhibition of development | [229] |
| miR -200c | Inhibition | GATA4 | Differentiation | [232] |
| Development | ||||
| Inhibition | SRF | Differentiation | ||
| Development | ||||
| Inhibition | TBX5 | Differentiation | ||
| Development | ||||
| Inhibition | CACNA1C | Differentiation | ||
| Development | ||||
| Inhibition | KCNJ2 | Differentiation | ||
| Development | ||||
| Inhibition | SCN5A | Differentiation | ||
| Development | ||||
| miR-204 | Inhibition | JARID2 | Inhibition of proliferation | [272] |
| miR302-367 | Inhibition | MST1 | Inhibition of proliferation | [273] |
| Inhibition | LATS2 | Inhibition of proliferation | ||
| Inhibition | MOB1B | Inhibition of proliferation | ||
| Inhibition | CCND1 | Proliferation | ||
| Inhibition | GATA4 | Differentiation | ||
| Inhibition | NKX2.5 | Development | ||
| Inhibition | MYH6 | Differentiation | ||
| Inhibition | MYH7 | Differentiation | ||
| bta-miR-365-3p | Inhibition | ACVR1 | Inhibition of differentiation | [274] |
| Proliferation | ||||
| Inhibition | CCND1 | Proliferation | ||
| Inhibition | CDK2 | Proliferation | ||
| Inhibition | PCNA | Proliferation | ||
| Activation | MYOD1 | Differentiation | ||
| Activation | MYOG | Differentiation | ||
| hsa-miR-4443 | Inhibition | THBS1 | Proliferation | [275] |
| Inhibition | COLLAGEN I | Proliferation | ||
| Inhibition | COLLAGEN III | Proliferation | ||
| Inhibition | SMAD2 | Proliferation | ||
| Inhibition | SMAD3 | Proliferation | ||
| Inhibition | A-SMA | Proliferation | ||
| miR-208 | Activation | KDM6A | Differentiation | [224] |
| Activation | KDM6B | Differentiation | ||
| Activation | TBX5 | Differentiation | ||
| Activation | MEF2C | Differentiation | ||
| Activation | GATA4 | Differentiation | ||
| miR-210 | Inhibition | EPHRIN-A3 | Differentiation | [276,277] |
| miR-215-5p | Inhibition | CTCF | Development | [278] |
| Differentiation | ||||
| MiR218 | Inhibition | SFRP2 | Inhibition of proliferation | [279] |
| Differentiation | ||||
| Inhibition | A-MHC | Development | ||
| Differentiation | ||||
| Inhibition | Β-MHC | Development | ||
| Inhibition | ACTC1 | Development | ||
| Inhibition | TNI | Differentiation | ||
| miR-221 | Inhibition | ERBB4 | Inhibition of development | [229] |
| miR-222 | Activation | ANP | Proliferation | [228,229] |
| Growth | ||||
| Activation | BNP | Proliferation | ||
| Growth | ||||
| Activation | ACTA1 | Proliferation | ||
| Growth | ||||
| Activation | MHC-A | ProliferationGrowth | ||
| Activation | MHC- Β | Proliferation | ||
| Growth | ||||
| Activation | CX-43 | Development | ||
| Activation | KIR2.1 | Development | ||
| Inhibition | ERBB4 | Inhibition of development | ||
| Inhibition | HIPK1 | Remodeling | ||
| Inhibition | HIPK2 | Remodeling | ||
| Inhibition | HMBOX1 | Remodeling | ||
| Inhibition | P27 | Remodeling | ||
| miR-294 | Inhibition | WEE1 | Inhibition of proliferation | [230] |
| Activation | CCNB1 | Proliferation | ||
| Activation | CCND1 | Proliferation | ||
| Activation | CCNE1 | Proliferation | ||
| Activation | CCNA2 | Proliferation | ||
| Activation | CDK1 | Proliferation | ||
| Activation | E2F1 | Proliferation | ||
| Activation | E2F3 | Proliferation | ||
| miR-300 | Inhibition | BMI1 | Differentiation | [280] |
| miR302-367 | Inhibition | MST1 | Inhibition of proliferation | [273] |
| Inhibition | MOB1B | Inhibition of proliferation | ||
| Inhibition | LATS2 | Inhibition of proliferation | ||
| miR-322 | Inhibition | CELF1 | Inhibition of differentiation | [281] |
| Activation | TBX5 | Differentiation | ||
| Activation | MEF2C | Differentiation | ||
| Activation | NKX2.5 | Differentiation | ||
| Activation | A-MHC | Differentiation | ||
| Activation | A-ACTININ | Differentiation | ||
| Activation | CTNT | Differentiation | ||
| miR-375 | Inhibition | NOTCH2 | Differentiation | [233,282] |
| Proliferation | ||||
| Development | ||||
| Inhibition | DLL1 | Differentiation | ||
| Proliferation | ||||
| Inhibition | HES1 | Differentiation | ||
| Proliferation | ||||
| Inhibition | CTNT | Differentiation | ||
| Inhibition | NKX2.5 | Differentiation | ||
| Inhibition | GATA4 | Differentiation | ||
| miR-410 | Inhibition | CITED2 | Inhibition of proliferation | [283] |
| Activation | PCNA | Proliferation | ||
| Inhibition | CITED2 | Development | ||
| Inhibition | ERFI1 | Inhibition of proliferation | ||
| Inhibition | PPP1CB | Growth | ||
| Inhibition | RERE | Inhibition of proliferation | ||
| Inhibition | STAT3 | Differentiation | ||
| miR-430a | Inhibition | POSQT | Inhibition of development | [284] |
| miR-495 | Inhibition | CITED2 | Inhibition of proliferation | [283] |
| Activation | PCNA | Proliferation | ||
| Inhibition | CITED2 | Development | ||
| Inhibition | ERFI1 | Inhibition of proliferation | ||
| Inhibition | PPP1CB | Growth | ||
| Inhibition | RERE | Inhibition of proliferation | ||
| Inhibition | STAT3 | Differentiation | ||
| miR-499 | Inhibition | KDM6A | Reprogramming | [224] |
| Inhibition | KDM6B | Reprogramming | ||
| Activation | TBX5 | Differentiation | ||
| Activation | MEF2C | Differentiation | ||
| Activation | GATA4 | Differentiation | ||
| Activation | CX43 | Differentiation | [285] | |
| Activation | CTNT | Differentiation | ||
| Activation | NKX2.5 | Differentiation | ||
| Activation | GATA4 | Differentiation | ||
| miR -499a-5p | Activation | A-ACTININ | Differentiation | [286,287] |
| Activation | CTNI | Differentiation | ||
| Inhibition | CDK6 | Proliferation | ||
| miR-503 | Inhibition | CELF1 | Inhibition of differentiation | [281] |
| Activation | TBX5 | Differentiation | ||
| Activation | MEF2C | Differentiation | ||
| Activation | NKX2.5 | Differentiation | ||
| Activation | A-ACTININ | Differentiation | ||
| Activation | A-MHC | Differentiation | ||
| Activation | CTNT | Differentiation | ||
| miR-590 | Activation | CLIC5 | Inhibition of proliferation | [247] |
| Activation | HOMER1 | Inhibition of proliferation | ||
| miR-699a | Inhibition | MYOD | Inhibition of differentiation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zare, A.; Salehpour, A.; Khoradmehr, A.; Bakhshalizadeh, S.; Najafzadeh, V.; Almasi-Turk, S.; Mahdipour, M.; Shirazi, R.; Tamadon, A. Epigenetic Modification Factors and microRNAs Network Associated with Differentiation of Embryonic Stem Cells and Induced Pluripotent Stem Cells toward Cardiomyocytes: A Review. Life 2023, 13, 569. https://doi.org/10.3390/life13020569
Zare A, Salehpour A, Khoradmehr A, Bakhshalizadeh S, Najafzadeh V, Almasi-Turk S, Mahdipour M, Shirazi R, Tamadon A. Epigenetic Modification Factors and microRNAs Network Associated with Differentiation of Embryonic Stem Cells and Induced Pluripotent Stem Cells toward Cardiomyocytes: A Review. Life. 2023; 13(2):569. https://doi.org/10.3390/life13020569
Chicago/Turabian StyleZare, Afshin, Aria Salehpour, Arezoo Khoradmehr, Shabnam Bakhshalizadeh, Vahid Najafzadeh, Sahar Almasi-Turk, Mahdi Mahdipour, Reza Shirazi, and Amin Tamadon. 2023. "Epigenetic Modification Factors and microRNAs Network Associated with Differentiation of Embryonic Stem Cells and Induced Pluripotent Stem Cells toward Cardiomyocytes: A Review" Life 13, no. 2: 569. https://doi.org/10.3390/life13020569