
Comparative Proteomic Analysis of the Venoms from the Most Dangerous Scorpions in Morocco: Androctonus mauritanicus and Buthus occitanus

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Methods
2.1. Ethical Statements
2.2. Venom Preparation
2.3. Venom Lethality (LD50)
2.4. Venom Separation by SDS-PAGE
2.5. Venom Fractionation by RP-HPLC
2.6. Mass Spectrometry Analysis
2.7. Tryptic Digestion
2.8. LC/MS/MS Characterization
3. Results
3.1. Protein Quantification
3.2. Lethality of A. mauritanicus and B. occitanus Venoms
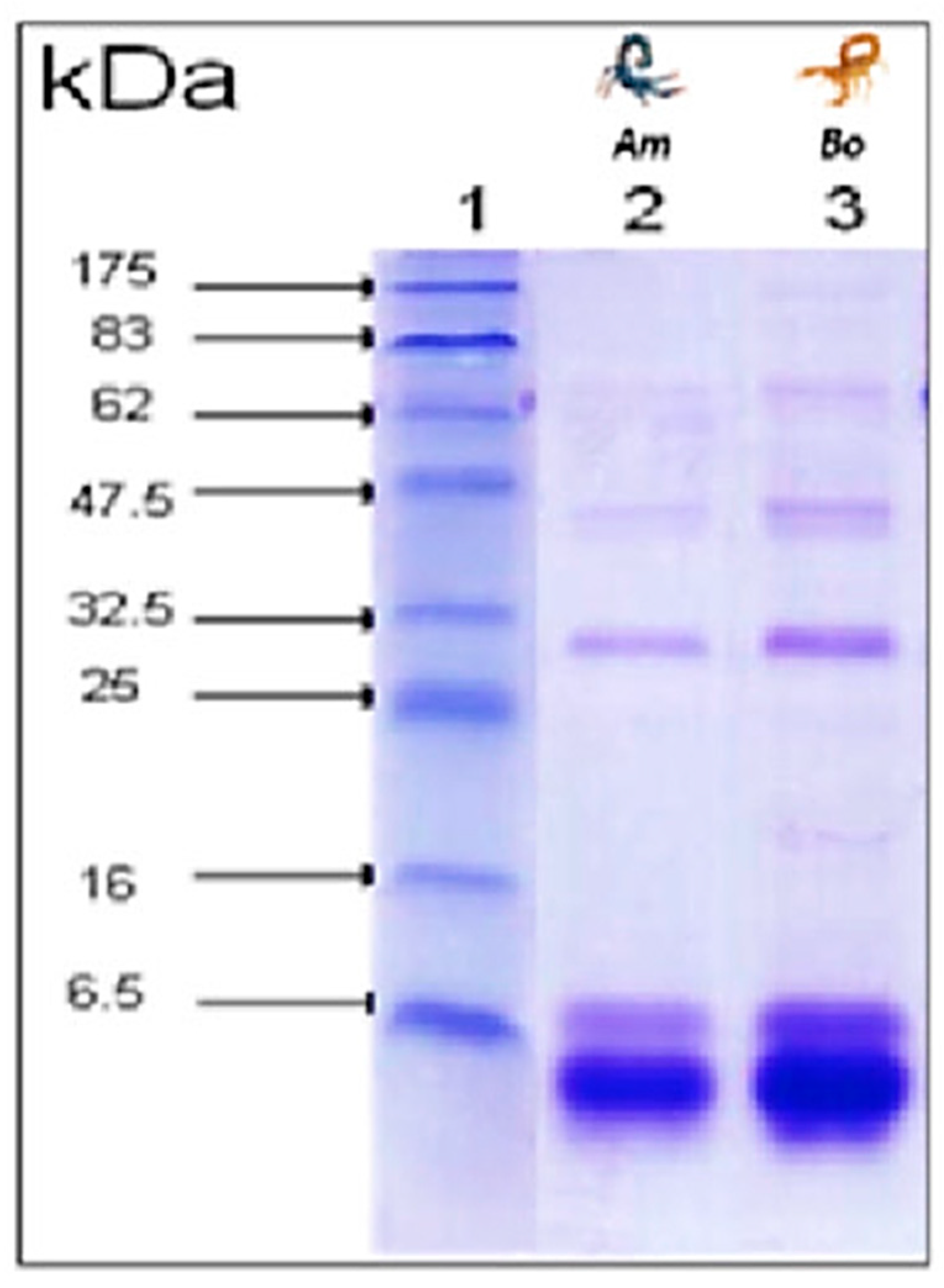
3.3. Electrophoretic Profile
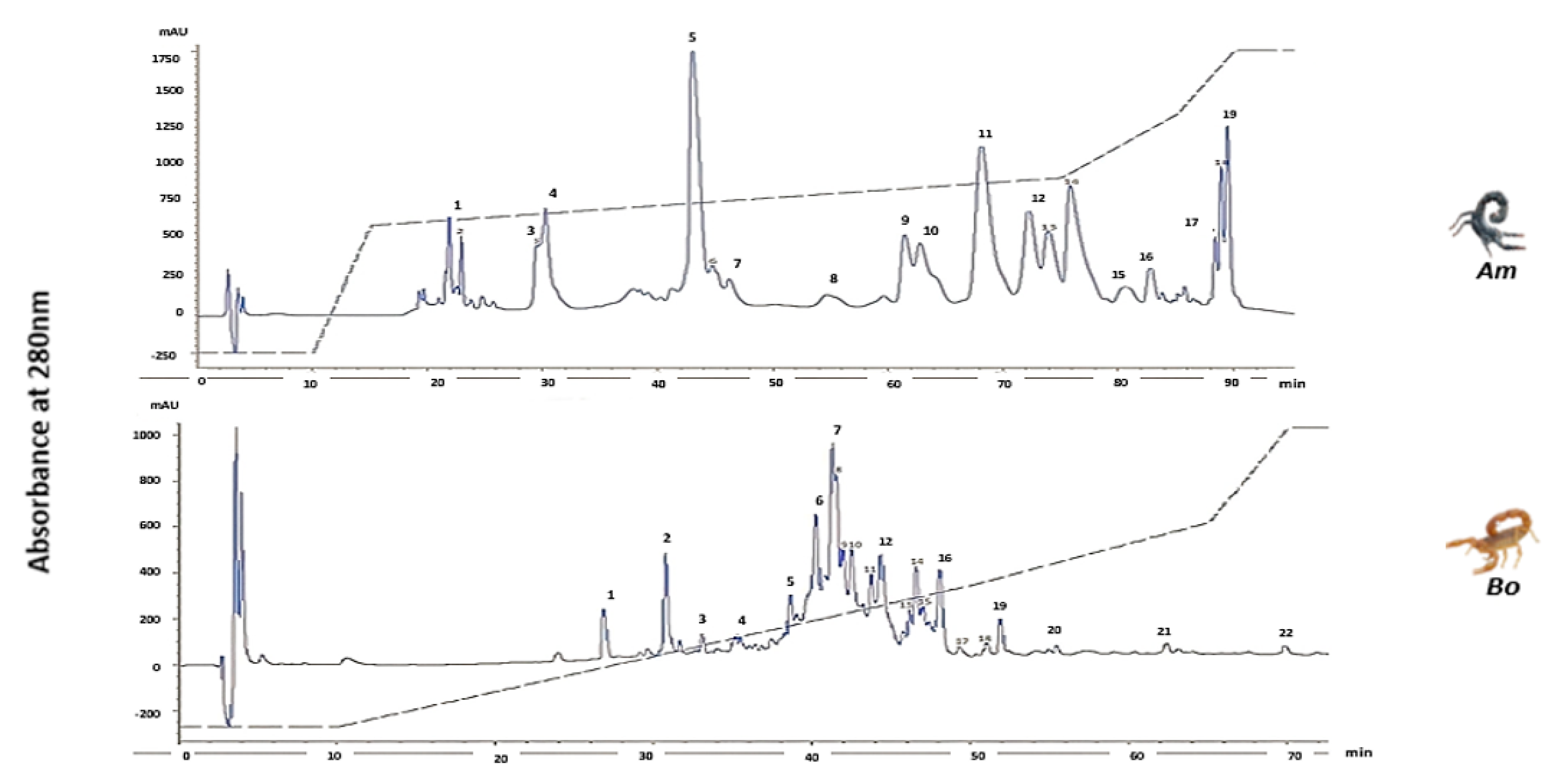
3.4. HPLC
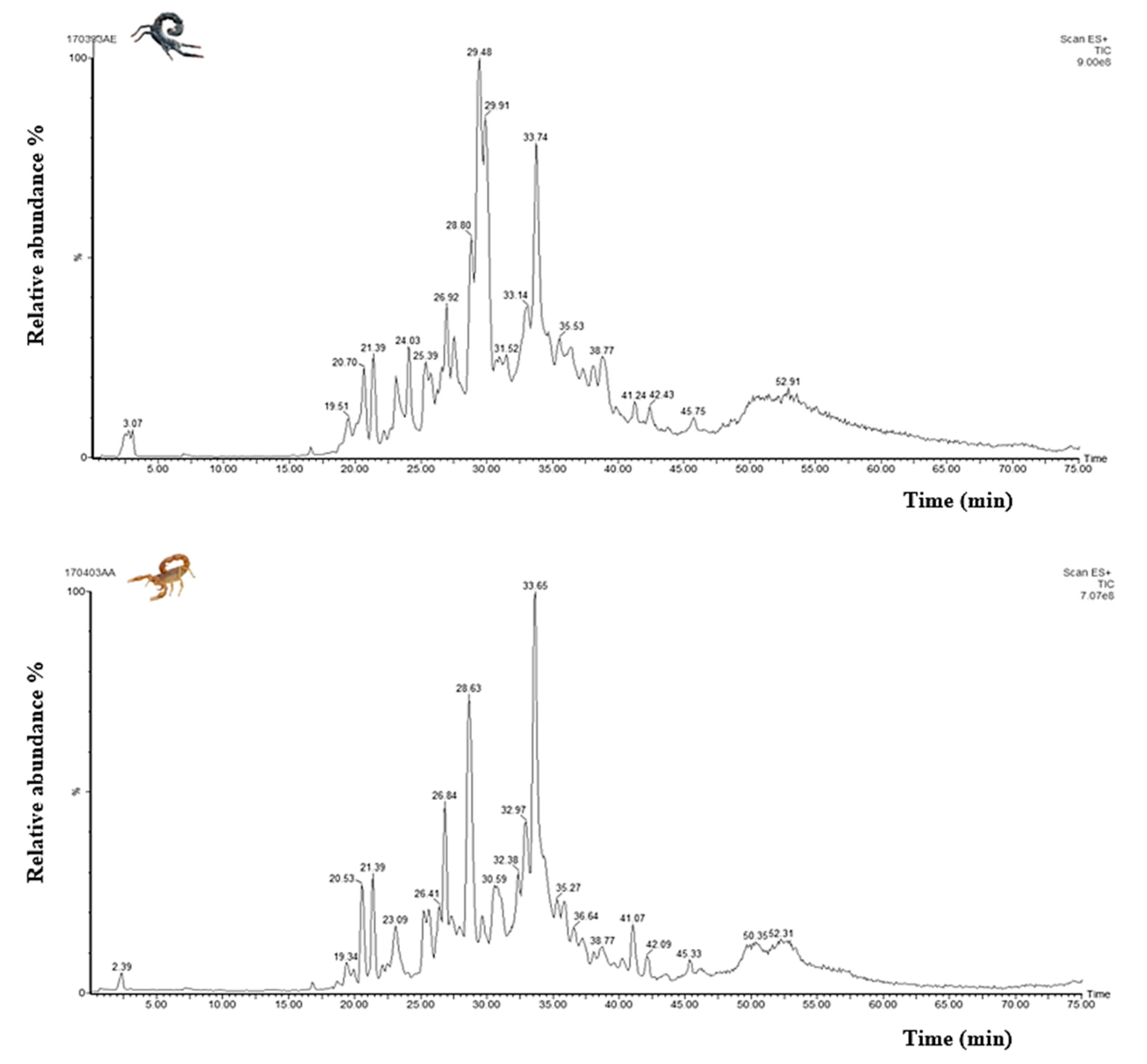
3.5. Mass Fingerprinting of Am and Bo Venoms
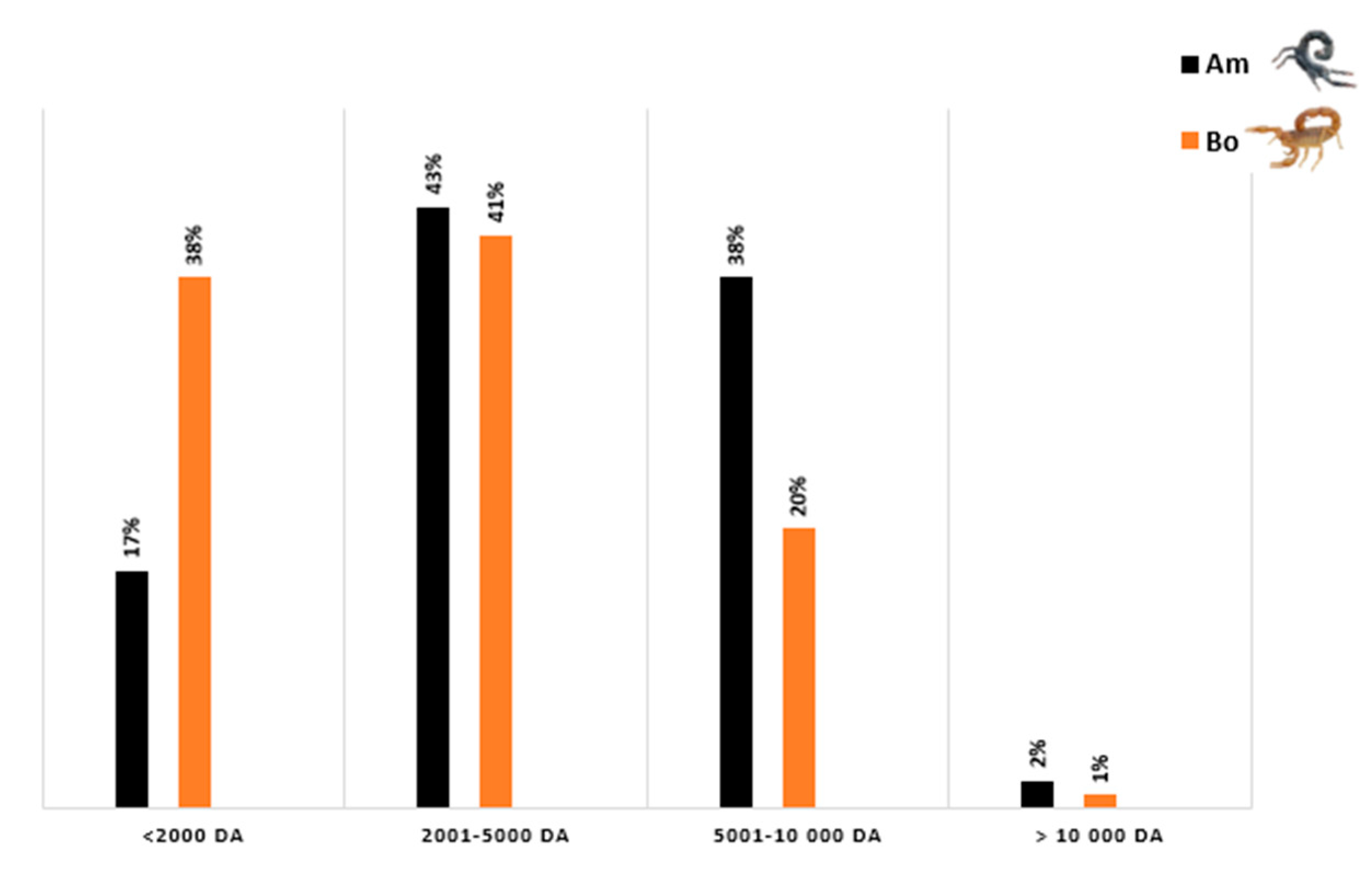
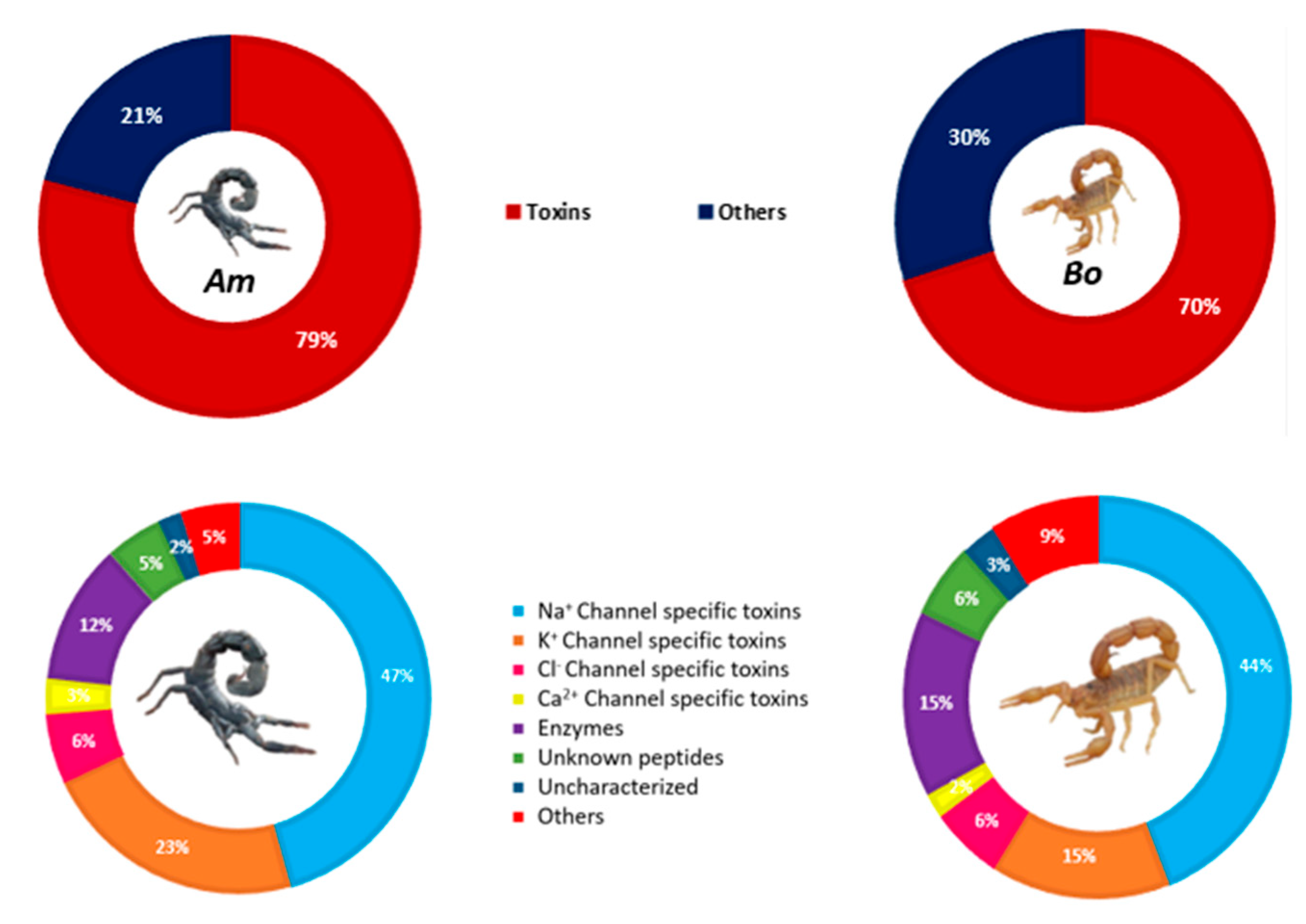
3.6. Composition of Am and Bo Venoms
3.7. Mass Spectrometry Identification
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lourenco, W.R. Diversity and endemism in tropical versus temperate scorpion communities. C. R. Séances Soc. Biogéogr. 1994, 70, 155–160. [Google Scholar]
- Ward, M.J.; Ellsworth, S.A.; Nystrom, G.S. A global accounting of medically significant scorpions: Epidemiology, major toxins, and comparative resources in harmless counterparts. Toxicon 2018, 151, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Briggs, D.E.G. Scorpions take to the water. Nature 1987, 326, 645–646. [Google Scholar] [CrossRef]
- Lourenço, W.R.; Cuellar, O. Scorpions, scorpionism, life history strategies and parthenogenesis. J. Venom. Anim. Toxins 1995, 1, 51–62. [Google Scholar] [CrossRef]
- Debont, T.; Swerts, A.; van der Walt, J.J.; Müller, G.J.; Verdonck, F.; Daenens, P.; Tytgat, J. Comparison and characterization of the venoms of three Parabuthus scorpion species occurring in southern Africa. Toxicon 1998, 36, 341–352. [Google Scholar] [CrossRef]
- Prendini, L.; Wheeler, W.C. Scorpion higher phylogeny and classification, taxonomic anarchy, and standards for peer review in online publishing. Cladistics 2005, 21, 446–494. [Google Scholar] [CrossRef]
- Dehesa-Dávila, M.; Possani, L.D. Scorpionism and serotherapy in Mexico. Toxicon 1994, 32, 1015–1018. [Google Scholar] [CrossRef]
- Graham, M.R. Scorpions of the World. J. Arachnol. 2011, 39, 166–167. [Google Scholar] [CrossRef]
- Shulov, A.; Levy, G.; Efrati, P.; Zlotkin, E.; Miranda, F.; Rochat, H. Venoms of Buthinae. In Arthropod Venoms; Handbook of Experimental Pharmacology/Handbuch der Experimentellen Pharmakologie; Bettini, S., Ed.; Springer: Berlin/Heidelberg, Germany, 1978; pp. 309–369. [Google Scholar] [CrossRef]
- Fontecilla-Camps, J.C.; Almassy, R.J.; Suddath, F.L.; Bugg, C.E. The three-dimensional structure of scorpion neurotoxins. Toxicon 1982, 20, 1–7. [Google Scholar] [CrossRef]
- Catterall, W.A. Neurotoxins that Act on Voltage-Sensitive Sodium Channels in Excitable Membranes. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 15–43. [Google Scholar] [CrossRef]
- Rochat, H.; Bernard, P.; Couraud, F. Scorpion toxins: Chemistry and mode of action. Adv. Cytopharmacol. 1979, 3, 325–334. [Google Scholar]
- Carbone, E.; Wanke, E.; Prestipino, G.; Possani, L.D.; Maelicke, A. Selective blockage of voltage-dependent K+ channels by a novel scorpion toxin. Nature 1982, 296, 90–91. [Google Scholar] [CrossRef]
- Domingos Possani, L.; Martin, B.M.; Svendsen, I. The primary structure of noxiustoxin: A K+ channel blocking peptide, purified from the venom of the scorpion Centruroides noxius Hoffmann. Carlsberg Res. Commun. 1982, 47, 285–289. [Google Scholar] [CrossRef]
- Miller, C.; Moczydlowski, E.; Latorre, R.; Phillips, M. Charybdotoxin, a protein inhibitor of single Ca2+-activated K+ channels from mammalian skeletal muscle. Nature 1985, 313, 316–318. [Google Scholar] [CrossRef]
- Valdivia, H.H.; Possani, L.D. Peptide Toxins as Probes of Ryanodine Receptor Structure and Function. Trends Cardiovasc. Med. 1998, 8, 111–118. [Google Scholar] [CrossRef]
- DeBin, J.A.; Maggio, J.E.; Strichartz, G.R. Purification and characterization of chlorotoxin, a chloride channel ligand from the venom of the scorpion. Am. J. Physiol. Physiol. 1993, 264, C361–C369. [Google Scholar] [CrossRef]
- Gwee, M.C.; Nirthanan, S.N.; Khoo, H.-E.; Gopalakrishnakone, P.; Kini, M.; Cheah, L.-S. Autonomic effects of some scorpion venoms and toxins. Clin. Exp. Pharmacol. Physiol. 2002, 29, 795–801. [Google Scholar] [CrossRef]
- Chippaux, J.-P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef]
- Santos, M.S.V.; Silva, C.G.L.; Neto, B.S.; Grangeiro, C.R.P.; Lopes, V.H.G.; Teixeira, A.G., Jr.; Bezerra, D.A.; Luna, J.; Cordeiro, J.B.; Goncalves, J.; et al. Clinical and Epidemiological Aspects of Scorpionism in the World: A Systematic Review. Wilderness Environ. Med. 2016, 27, 504–518. [Google Scholar] [CrossRef]
- Chippaux, J.-P. Emerging options for the management of scorpion stings. Drug Des. Dev. Ther. 2012, 6, 165–173. [Google Scholar] [CrossRef]
- El Hidan, M.A.; Touloun, O.; El Oufir, R.; Boumezzough, A. Epidemiological and spatial analysis of scorpion stings in two regions of Morocco: Marrakesh-Tensift-Al Haouz and Souss-Massa-Draa. J. Coast Life Med. 2016, 4, 299–304. [Google Scholar] [CrossRef]
- El Oufir, R. Piqures et Envenimations Scorpioniques (PES); Rapports general et specifiques. Revue toxicologie Maroc; Centre Anti Poison du Maroc (CAPM): Rabat, Morocco, 2019; Volume 43, p. 11. [Google Scholar]
- Ghalim, N.; Benslimane, A.; Sebti, F.; El-Hafny, B.; Lazar, N.; Moustanir, R.; Heikel, J. Scorpion envenomation and serotherapy in Morocco. Am. J. Trop. Med. Hyg. 2000, 62, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Oukkache, N.; Rosso, J.-P.; Alami, M.; Ghalim, N.; Saïle, R.; Hassar, M.; Bougis, P.E.; Martin-Eauclaire, M.-F. New analysis of the toxic compounds from the Androctonus mauretanicus mauretanicus scorpion venom. Toxicon 2008, 51, 835–852. [Google Scholar] [CrossRef] [PubMed]
- Oukkache, N.; Chgoury, F.; Lalaoui, M.; Cano, A.A.; Ghalim, N. Comparison between two methods of scorpion venom milking in Morocco. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 1–5. [Google Scholar] [CrossRef]
- Ladjel-Mendil, A.; Martin-Eauclaire, M.F.; Laraba-Djebari, F. Neuropathophysiological Effect and Immuno-Inflammatory Response Induced by Kaliotoxin of Androctonus Scorpion Venom. Neuroimmunomodulation 2013, 20, 99–106. [Google Scholar] [CrossRef]
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion venom components as potential candidates for drug development. Toxicon 2015, 93, 125–135. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Harrison, P.L.; Strong, P.N. Snapshots of scorpion venomics. J. Arid. Environ. 2015, 112, 170–176. [Google Scholar] [CrossRef]
- Smith, J.J.; Alewood, P.F. Modern Venom Profiling: Mining into Scorpion Venom Biodiversity. In Scorpion Venoms; Gopalakrishnakone, P., Possani, L.D., Schwartz, E.F., Rodríguez de la Vega, R.C., Eds.; Springer: Dordrecht, The Netherlands, 2015; p. 547. Available online: http://link.springer.com/10.1007/978-94-007-6404-0_26 (accessed on 5 March 2023).
- Compton, P.D.; Kelleher, N.L. Spinning up mass spectrometry for whole protein complexes. Nat. Methods 2012, 9, 1065–1066. [Google Scholar] [CrossRef]
- Pimenta, A.M.C.; Stöcklin, R.; Favreau, P.; Bougis, P.E.; Martin-Eauclaire, M.F. Moving pieces in a proteomic puzzle: Mass fingerprinting of toxic fractions from the venom of Tityus serrulatus (Scorpiones, Buthidae): Mass fingerprinting of Tityus serrulatus toxic fractions. Rapid Commun. Mass Spectrom. 2001, 15, 1562–1572. [Google Scholar] [CrossRef]
- Favreau, P.; Menin, L.; Michalet, S.; Perret, F.; Cheneval, O.; Stöcklin, M.; Bulet, P.; Stöcklin, R. Mass spectrometry strategies for venom mapping and peptide sequencing from crude venoms: Case applications with single arthropod specimen. Toxicon 2006, 47, 676–687. [Google Scholar] [CrossRef]
- Banerjee, S.; Mazumdar, S. Electrospray Ionization Mass Spectrometry: A technique to access the information beyond the molecular weight of the analyte. Int. J. Anal. Chem. 2012, 2012, 282574. [Google Scholar] [CrossRef]
- Aboumaâd, B.; Lahssaini, M.; Tiger, A.; Benhassain, S.M. Clinical comparison of scorpion envenomation by Androctonus mauritanicus and Buthus occitanus in children. Toxicon 2014, 90, 337–343. [Google Scholar] [CrossRef]
- Daoudi, K.; Chgoury, F.; Rezzak, M.; Bourouah, O.; Boussadda, L.; Soukri, A.; Sabatier, J.-M.; Oukkache, N. Consequences of Androctonus mauretanicus and Buthus occitanus scorpion venoms on electrolyte levels in rabbits. Heliyon 2017, 3, e00221. [Google Scholar] [CrossRef]
- Emerich, B.L.; De Lima, M.E.; Martin-Eauclaire, M.-F.; Bougis, P.E. Comparative analyses and implications for antivenom serotherapy of four Moroccan scorpion Buthus occitanus venoms: Subspecies tunetanus, paris, malhommei, and mardochei. Toxicon 2018, 149, 26–36. [Google Scholar] [CrossRef]
- Darkaoui, B.; Lafnoune, A.; Chgoury, F.; Daoudi, K.; Chakir, S.; Mounaji, K.; Karkouri, M.; Cadi, R.; Naoual, O. Induced pathophysiological alterations by the venoms of the most dangerous Moroccan scorpions Androctonus mauretanicus and Buthus occitanus: A comparative pathophysiological and toxic-symptoms study. Hum. Exp. Toxicol. 2022, 41, 2872. [Google Scholar] [CrossRef]
- Daoudi, K.; Malosse, C.; Lafnoune, A.; Darkaoui, B.; Chakir, S.; Sabatier, J.M.; Cadi, R.; Oukkache, N. Mass spectrometry-based top-down and bottom-up approaches for proteomic analysis of the Moroccan Buthus occitanus scorpion venom. FEBS Open Bio 2021, 11, 1867–1892. [Google Scholar] [CrossRef]
- Layne, E. Spectrophotometric and turbidimetric methods for measuring proteins. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1957; p. 447. Available online: https://www.sciencedirect.com/science/article/pii/S0076687957034138 (accessed on 5 March 2023).
- Stoscheck, C.M. Quantitation of protein. In Methods in Enzymology; Guide to Protein, Purification; Deutscher, M.P., Ed.; Academic Press: Cambridge, MA, USA, 1990; Volume 182, pp. 50–68. Available online: https://www.sciencedirect.com/science/article/pii/007668799082008P (accessed on 5 March 2023).
- World Health Organization. Progress in the Characterization of Venoms and Standardization of Antivenoms. 1981. Available online: https://apps.who.int/iris/handle/10665/37282 (accessed on 5 March 2023).
- Oukkache, N.; El Jaoudi, R.; Ghalim, N.; Chgoury, F.; Bouhaouala, B.; El Mdaghri, N.; Sabatier, J.-M. Evaluation of the Lethal Potency of Scorpion and Snake Venoms and Comparison between Intraperitoneal and Intravenous Injection Routes. Toxins 2014, 6, 1873–1881. [Google Scholar] [CrossRef]
- Trevors, J.T. A BASIC program for estimating LD50 values using the IBM-PC®. Bull. Environ. Contam. Toxicol. 1986, 37, 18–26. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- O’Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 1975, 250, 4007–4021. [Google Scholar] [CrossRef]
- Carrette, O.; Burkhard, P.R.; Sanchez, J.-C.; Hochstrasser, D.F. State-of-the-art two-dimensional gel electrophoresis: A key tool of proteomics research. Nat. Protoc. 2006, 1, 812–823. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, M.I.; Hearn, M.T.W. High-resolution reversed-phase high-performance liquid chromatography of peptides and proteins. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1996; p. 3. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0076687996700034 (accessed on 5 March 2023).
- Zhang, L.; Shi, W.; Zeng, X.-C.; Ge, F.; Yang, M.; Nie, Y.; Bao, A.; Wu, S.; Guoji, E. Unique diversity of the venom peptides from the scorpion Androctonus bicolor revealed by transcriptomic and proteomic analysis. J. Proteom. 2015, 128, 231–250. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Gu, J.; Yan, Z.; Wang, M.; Ma, C.; Zhang, J.; Jiang, G.; Ge, M.; Xu, S.; Xu, Z.; et al. De novo transcriptomic analysis of the venomous glands from the scorpion Heterometrus spinifer revealed unique and extremely high diversity of the venom peptides. Toxicon 2018, 143, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aziz, T.M.; Soares, A.G.; Stockand, J.D. Advances in venomics: Modern separation techniques and mass spectrometry. J. Chromatogr. B 2020, 1160, 122352. [Google Scholar] [CrossRef]
- Nadia, A.; Rachid, H.; Nadra, L.; Naima, R.; Azeddine, S. Méta-analyse des données des envenimations scorpioniques au Maroc. SMETox J. 2018, 1, 115–122. [Google Scholar]
- Borges, A.; Graham, M.R.; Gopalakrishnakone, P.; Calvete, J.J. Phylogenetics of Scorpions of Medical Importance. In Venom Genomics and Proteomics; Gopalakrishnakone, P., Calvete, J.J., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 81–104. [Google Scholar] [CrossRef]
- Zeng, X.C.; Peng, F.; Luo, F.; Zhu, S.Y.; Liu, H.; Li, W.X. Molecular cloning and characterization of four scorpion K+-toxin-like peptides: A new subfamily of venom peptides (α-KTx14) and genomic analysis of a member. Biochimie 2001, 83, 883–889. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Quintero-Hernandez, V.; Possani, L.D. Venom proteomic and venomous glands transcriptomic analysis of the Egyptian scorpion Scorpio maurus palmatus (Arachnida: Scorpionidae). Toxicon 2013, 74, 193–207. [Google Scholar] [CrossRef]
- Quintero-Hernández, V.; Ramírez-Carreto, S.; Romero-Gutiérrez, M.T.; Valdez-Velázquez, L.L.; Becerril, B.; Possani, L.D.; Ortiz, E. Transcriptome Analysis of Scorpion Species Belonging to the Vaejovis Genus. PLoS ONE 2015, 10, e0117188. [Google Scholar] [CrossRef]
- Almaaytah, A.; Albalas, Q. Scorpion venom peptides with no disulfide bridges: A review. Peptides 2014, 51, 35–45. [Google Scholar] [CrossRef]
- Couraud, F.; Jover, E.; Dubois, J.; Rochat, H. Two types of scorpion toxin receptor sites, one related to the activation, the other to the inactivation of the action potential sodium channel. Toxicon 1982, 20, 9–16. [Google Scholar] [CrossRef]
- Quintero-Hernández, V.; Jiménez-Vargas, J.; Gurrola, G.; Valdivia, H.; Possani, L. Scorpion venom components that affect ion-channels function. Toxicon 2013, 76, 328–342. [Google Scholar] [CrossRef]
- Cid-Uribe, J.I.; Veytia-Bucheli, J.I.; Romero-Gutierrez, T.; Ortiz, E.; Possani, L.D. Scorpion venomics: A 2019 overview. Expert Rev. Proteom. 2020, 17, 67–83. [Google Scholar] [CrossRef]
- Isbister, G.K.; Bawaskar, H.S. Scorpion Envenomation. N. Engl. J. Med. 2014, 371, 457–463. [Google Scholar] [CrossRef]
- Rosso, J.; Rochat, H. Characterization of ten proteins from the venom of the Moroccan scorpion Androctonus mauretanicus mauretanicus, six of which are toxic to the mouse. Toxicon 1985, 23, 113–125. [Google Scholar] [CrossRef]
- Zerrouk, H.; Bougis, P.; Céard, B.; Benslimane, A.; Martin-Eauclaire, M. Analysis by high-performance liquid chromatography of Androctonus mauretanicus mauretanicus (black scorpion) venom. Toxicon 1991, 29, 951–960. [Google Scholar] [CrossRef]
- Crest, M.; Jacquet, G.; Gola, M.; Zerrouk, H.; Benslimane, A.; Rochat, H.; Mansuelle, P.; Martin-Eauclaire, M.F. Kaliotoxin, a novel peptidyl inhibitor of neuronal BK-type Ca(2+)-activated K+ channels characterized from Androctonus mauretanicus mauretanicus venom. J. Biol. Chem. 1992, 267, 1640–1647. [Google Scholar] [CrossRef]
- Zerrouk, H.; Mansuelle, P.; Benslimane, A.; Rochat, H.; Martin-Eauclaire, M. Characterization of a new leiurotoxin I-like scorpion toxin. FEBS Lett. 1993, 320, 189–192. [Google Scholar] [CrossRef]
- Zerrouk, H.; Laraba-Djebari, F.; Fremont, V.; Meki, A.; Darbon, H.; Mansuelle, P.; Oughideni, R.; VAN Rietschoten, J.; Rochat, H.; Martin-Eauclaire, M. Characterization of PO1, a new peptide ligand of the apamin-sensitive Ca2+ activated K+ channel. Int. J. Pept. Protein Res. 1996, 48, 514–521. [Google Scholar] [CrossRef]
- Alami, M.; Ouafik, L.; Céard, B.; Legros, C.; Bougis, P.E.; Martin-Eauclaire, M.-F. Characterisation of the gene encoding the α-toxin Amm V from the scorpion Androctonus mauretanicus mauretanicus. Toxicon 2001, 39, 1579–1585. [Google Scholar] [CrossRef]
- Vacher, H.; Alami, M.; Crest, M.; Possani, L.D.; Bougis, P.E.; Martin-Eauclaire, M.-F. Expanding the scorpion toxin α-KTX 15 family with AmmTX3 from Androctonus mauretanicus. J. Biol. Inorg. Chem. 2002, 269, 6037–6041. [Google Scholar] [CrossRef]
- Almaaytah, A.; Tarazi, S.; Mhaidat, N.; Al-Balas, Q.; Mukattash, T.L. Mauriporin, a Novel Cationic α-Helical Peptide with Selective Cytotoxic Activity Against Prostate Cancer Cell Lines from the Venom of the Scorpion Androctonus mauritanicus. Int. J. Pept. Res. Ther. 2013, 19, 281–293. [Google Scholar] [CrossRef]
- Martin-Eauclaire, M.-F.; Céard, B.; Belghazi, M.; Lebrun, R.; Bougis, P.E. Characterization of the first K+ channel blockers from the venom of the Moroccan scorpion Buthus occitanus Paris. Toxicon 2013, 75, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Oukkache, N.; Eljaoudi, R.; Chgoury, F.; Rocha, M.T.; Sabatier, J.-M. Characterization of Am IT, an anti-insect β-toxin isolated from the venom of scorpion Androctonus mauretanicus. Acta Physiol. Sonica 2015, 25, 295–304. [Google Scholar]
- Valdez-Velázquez, L.L.; Quintero-Hernández, V.; Romero-Gutiérrez, M.T.; Coronas, F.I.V.; Possani, L.D. Mass Fingerprinting of the Venom and Transcriptome of Venom Gland of Scorpion Centruroides tecomanus. PLoS ONE 2013, 8, e66486. [Google Scholar] [CrossRef] [PubMed]
- Bringans, S.; Eriksen, S.; Kendrick, T.; Gopalakrishnakone, P.; Livk, A.; Lock, R.; Lipscombe, R. Proteomic analysis of the venom of Heterometrus longimanus (Asian black scorpion). Proteomics 2008, 8, 1081–1096. [Google Scholar] [CrossRef]
- Ma, Y.; Zhao, Y.; Zhao, R.; Zhang, W.; He, Y.; Wu, Y.; Guo, L. Molecular diversity of toxic components from the scorpion Heterometrus petersii venom revealed by proteomic and transcriptome analysis. Proteomics 2010, 10, 2471–2485. [Google Scholar] [CrossRef]
- Xu, X.; Duan, Z.; Di, Z.; He, Y.; Li, J.; Li, Z.; Xie, C.; Zeng, X.; Cao, Z.; Wu, Y.; et al. Proteomic analysis of the venom from the scorpion Mesobuthus martensii. J. Proteom. 2014, 106, 162–180. [Google Scholar] [CrossRef]
- Valdez-Velázquez, L.L.; Cid-Uribe, J.; Romero-Gutierrez, M.T.; Olamendi-Portugal, T.; Jimenez-Vargas, J.M.; Possani, L.D. Transcriptomic and proteomic analyses of the venom and venom glands of Centruroides hirsutipalpus, a dangerous scorpion from Mexico. Toxicon 2020, 179, 21–32. [Google Scholar] [CrossRef]
- Das, B.; Patra, A.; Mukherjee, A.K. Correlation of Venom Toxinome Composition of Indian Red Scorpion (Mesobuthus tamulus) with Clinical Manifestations of Scorpion Stings: Failure of Commercial Antivenom to Immune-Recognize the Abundance of Low Molecular Mass Toxins of This Venom. J. Proteome Res. 2020, 19, 1847–1856. [Google Scholar] [CrossRef]
- Santibáñez-López, C.E.; Cid-Uribe, J.I.; Zamudio, F.Z.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. Venom gland transcriptomic and venom proteomic analyses of the scorpion Megacormus gertschi Díaz-Najera, 1966 (Scorpiones: Euscorpiidae: Megacorminae). Toxicon 2017, 133, 95–109. [Google Scholar] [CrossRef]
- Romero-Gutierrez, T.; Peguero-Sanchez, E.; Cevallos, M.A.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. A Deeper Examination of Thorellius atrox Scorpion Venom Components with Omic Technologies. Toxins 2017, 9, 399. [Google Scholar] [CrossRef]
- Magalhães, A.C.M.; de Santana, C.J.C.; Melani, R.D.; Domont, G.B.; Castro, M.S.; Fontes, W.; Roepstorff, P.; Júnior, O.R.P. Exploring the biological activities and proteome of Brazilian scorpion Rhopalurus agamemnon venom. J. Proteom. 2021, 237, 104119. [Google Scholar] [CrossRef]
- Diego-García, E.; Batista, C.V.; García-Gómez, B.I.; Lucas, S.; Candido, D.M.; Gómez-Lagunas, F.; Possani, L.D. The Brazilian scorpion Tityus costatus Karsch: Genes, peptides and function. Toxicon 2005, 45, 273–283. [Google Scholar] [CrossRef]
- Barona, J.; Batista, C.V.F.; Zamudio, F.Z.; Gomez-Lagunas, F.; Wanke, E.; Otero, R.; Possani, L.D. Proteomic analysis of the venom and characterization of toxins specific for Na+- and K+-channels from the Colombian scorpion Tityus pachyurus. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2006, 1764, 76–84. [Google Scholar] [CrossRef]
- Batista, C.V.F.; D’Suze, G.; Gómez-Lagunas, F.; Zamudio, F.Z.; Encarnación, S.; Sevcik, C.; Possani, L.D. Proteomic analysis of Tityus discrepans scorpion venom and amino acid sequence of novel toxins. Proteomics 2006, 6, 3718–3727. [Google Scholar] [CrossRef]
- Nascimento, D.G.; Rates, B.; Santos, D.M.; Verano-Braga, T.; Barbosa-Silva, A.; Dutra, A.A.; Biondi, I.; Martin-Eauclaire, M.F.; De Lima, M.E.; Pimenta, A.M. Moving pieces in a taxonomic puzzle: Venom 2D-LC/MS and data clustering analyses to infer phylogenetic relationships in some scorpions from the Buthidae family (Scorpiones). Toxicon 2006, 47, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Batista, C.; Román-González, S.; Salas-Castillo, S.; Zamudio, F.; Gómez-Lagunas, F.; Possani, L. Proteomic analysis of the venom from the scorpion Tityus stigmurus: Biochemical and physiological comparison with other Tityus species. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 146, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Caliskan, F.; García, B.I.; Coronas, F.I.; Batista, C.V.; Zamudio, F.Z.; Possani, L.D. Characterization of venom components from the scorpion Androctonus crassicauda of Turkey: Peptides and genes. Toxicon 2006, 48, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Diego-García, E.; Peigneur, S.; Clynen, E.; Marien, T.; Czech, L.; Schoofs, L.; Tytgat, J. Molecular diversity of the telson and venom components from Pandinus cavimanus (Scorpionidae Latreille 1802): Transcriptome, venomics and function. Proteomics 2012, 12, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Coronas, F.I.; Diego-García, E.; Restano-Cassulini, R.; de Roodt, A.R.; Possani, L.D. Biochemical and physiological characterization of a new Na+-channel specific peptide from the venom of the Argentinean scorpion Tityus trivittatus. Peptides 2015, 68, 11–16. [Google Scholar] [CrossRef]
- Rodríguez-Ravelo, R.; Batista, C.V.; Coronas, F.I.; Zamudio, F.Z.; Hernández-Orihuela, L.; Espinosa-López, G.; Ruiz-Urquiola, A.; Possani, L.D. Comparative proteomic analysis of male and female venoms from the Cuban scorpion Rhopalurus junceus. Toxicon 2015, 107, 327–334. [Google Scholar] [CrossRef]
- Batista, C.; Martins, J.; Restano-Cassulini, R.; Coronas, F.; Zamudio, F.; Procópio, R.; Possani, L. Venom characterization of the Amazonian scorpion Tityus metuendus. Toxicon 2018, 143, 51–58. [Google Scholar] [CrossRef]
- Romero-Gutiérrez, M.T.; Santibáñez-López, C.E.; Jiménez-Vargas, J.M.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. Transcriptomic and Proteomic Analyses Reveal the Diversity of Venom Components from the Vaejovid Scorpion Serradigitus gertschi. Toxins 2018, 10, 359. [Google Scholar] [CrossRef]
- Cid-Uribe, J.I.; Santibáñez-López, C.E.; Meneses, E.P.; Batista, C.V.F.; Jiménez-Vargas, J.M.; Ortiz, E.; Possani, L.D. The diversity of venom components of the scorpion species Paravaejovis schwenkmeyeri (Scorpiones: Vaejovidae) revealed by transcriptome and proteome analyses. Toxicon 2018, 151, 47–62. [Google Scholar] [CrossRef]
- Verano-Braga, T.; Dutra, A.A.A.; León, I.R.; Melo-Braga, M.N.; Roepstorff, P.; Pimenta, A.M.C.; Kjeldsen, F. Moving Pieces in a Venomic Puzzle: Unveiling Post-translationally Modified Toxins from Tityus serrulatus. J. Proteome Res. 2013, 12, 3460–3470. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Venoms | LD50 | |
|---|---|---|
| Am Venom | Bo Venom | |
| IV (µg/mouse) | 4.7 (4.1–5.4) | 15.2 (14.8–15.6) |
| IP (µg/mouse) | 5.8 (5.3–6.4) | 17.1 (16.7–17.5) |
| Proteins Associated with Am | |||||
|---|---|---|---|---|---|
| Sample/Fraction | Protein Name | Molecular Weight (Da) | Sequence | Swiss-Prot ID | Coverage (%) |
| A5, A6, A7, A8, A9, A10 | Alpha-toxin Amm3 OS = Androctonus mauritanicus mauritanicus | 7009.1 | (K)PVINITWLR NSKSVTDG(L) | P0C910 | 92.9 |
| A7, A8, A9, A10, A11 | Alpha-toxin Amm5 OS = Androctonus mauritanicus mauritanicus | 7301.3 | (−)LKDGYIIDDLNCTFFCGR(N) | P01482 | 89.7 |
| A5, A6, A7 | Alpha-toxin Amm8 OS = Androctonus mauritanicus mauritanicus | 9654.2 | (K) LPDHVR (T) | Q7YXD3 | 94.4 |
| A1, A4, A5, A6 | Neurotoxin P2 OS = Androctonus mauritanicus mauritanicus | 3673.3 | (−) CGPCFTTDPYTESK (C) | P01498 | 93.8 |
| A4, A5 | Potassium channel toxin alpha-KTx 15.3 OS = Androctonus mauritanicus mauritanicus | 3845.6 | (K) VIGVAAGK (C) | P60208 | 96.3 |
| A2, A3 | Potassium channel toxin alpha-KTx 3.1 OS = Androctonus mauritanicus mauritanicus | 4156 | (K) CSGSPQCLKPCK (D) | P24662 | 88 |
| A3, A4 | Potassium channel toxin alpha-KTx 5.2 OS = Androctonus mauritanicus mauritanicus | 3421.2 | (R) SLGLLGKCIGVK (C) | P31719 | 95.8 |
| A2, A3, A4, A5, A7, A8, A11 | Potassium channel toxin alpha-KTx 8.1 OS = Androctonus mauritanicus mauritanicus | 3184.6 | (−) VSCEDCPEHCTQK (A) | P56215 | 94 |
| Proteins Associated with other Species | |||||
| Sample/Fraction | Protein Name | Molecular Weight (Da) | Sequence | Swiss-Prot ID | Coverage (%) |
| A9, A10, A16, A17 | Alpha-mammal toxin Lqq5 OS = Leirus quinquestriatus quinquestriatus | 7301.3 | (−)LKDGYIVDDKNCTFFCGR(N) | A7NGC5 | 77.3 |
| A8 | Alpha-toxin BeM10 OS = Buthus eupeus | 7373.4 | (R) NAYCDEECK (K) | P01490 | 70.6 |
| A6, A14 | Beta-bungarotoxin BF B1 chain OS = Bungarus fasciatus | 9636.2 | (R) AFYYLPSAK (R) | B2KTG2 | 71 |
| A8 | Beta-insect excitatory toxin 1 OS = Androctonus australis | 9852 | (K)KNGYAVDSSG(K) | P01497 | 99.5 |
| A7 | Beta-insect excitatory toxin 2 OS = Androctonus australis | 9862.7 | (R)YAVDSSGK APECLLSN(K) | P15147 | 82.5 |
| A8 | Beta-insect excitatory toxin LqhTTlc OS = Leirus quinquestriatus hebraeus | 9935.8 | (K) VMEISDTR (K) | P68723 | 77.3 |
| A6, A7, A8, A9, A10 | Beta-toxin KAaM1 OS = Androctonus australis | 9104.8 | (K) YGYCYAFQCWCEYLEDK (N) | Q4LCT0 | 97.3 |
| A6, A7, A12, A19 | Neurotoxin BmK-M10 OS = Mesobuthus martensii | 9346.0 | (K) YGNACWCIK (L) | Q61705 | 85.4 |
| A13, A15, A16 | Toxin Aah4 OS = Androctonus australis | 9156.7 | (K) NCVYHCYPPCGLCK (K) | P45658 | 82.3 |
| A17, A18 | Neurotoxin LmNaTx34.5 (Fragment) OS = Lychas mucronatus | 9472.2 | (K)GGSYGYCYFWK(L) | POC16 | 73.1 |
| A18 | Potassium channel toxin AaTXK-beta OS = Androctonus australis | 10,148.2 | (R)TILQTVVHK(V) | P69939 | 87.6 |
| A10, A11 | Neurotoxin LmNaTx34.5 (Fragment) OS = Lychas mucronatus | 9472.2 | (R)AGREKGCK VWCVIN(N) | P0CI60 | 88.2 |
| A4, A5 | Neurotoxin-1 OS = Androctonus australis | 9061 | (C)VYHCVPPCDGLCK(K) | P01479 | 99.6 |
| A7, A8, A9 | Neurotoxin-like protein STR1 OS = Androctonus australis | 7640.8 | (R) DGYIVHDGTNCK (Y) | P80950 | 83.8 |
| A11 | Potassium channel toxin AaTXK-beta OS = Androctonus australis | 10,148.2 | (R)TILQTVVHK(V) | P69939 | 87.6 |
| A2 | Potassium channel toxin alpha-KTx 3.4 OS = Leirus quinquestriatus hebraeus | 4020.9 | (K) CTGSPQCLKPCK (D) | P46110 | 85.5 |
| A2, A3 | Potassium channel toxin alpha-KTx 3.9 OS = Buthus occitanus tunetanus | 4028 | (C)KDAGMRFGKCMNRK(C) | P59290 | 98.7 |
| A5, A6, A7, A8, A9 | Potassium channel toxin BmTXK-beta OS = Mesobuthus martensii | 10,430.6 | (K) LTSMSEYACPVIEK (W) | Q9NJC6 | 85.6 |
| A4, A5, A6, A7 | Potassium channel toxin BmTXK-beta-2 OS = Mesobuthus martensii | 10,212.2 | (K) TQFGCPAYQGYCDDHCQDIK (K) | Q9N661 | 78.4 |
| A8, A9, A10, A11 | Toxin AaHIT4 OS = Androctonus australis | 7786.0 | (R) KSELWNYK (T) | P21150 | 97 |
| A4 | Toxin BmKaITI OS = Mesobuthus martensii | 9649.3 | (R) DAYIAQNYNCVYCAR (D) | Q9GQW3 | 88.2 |
| A9 | Toxin BmTxKS4 OS = Mesobuthus martensii | 8856 | (K) GHSSCTNGLEMTEEDF (C) | Q5F1N4 | 99.7 |
| Proteins Associated with Bo | |||||
|---|---|---|---|---|---|
| Sample/Fraction | Protein Name | MW | Sequence | Swiss-Prot ID | % Coverage |
| B8, B9, B10 | Alpha-like toxin Bom3 OS = Buthus occitanus mardochei | 6871.8 | (K)LPDKVPIKVPGK(C) | P13488 | 93.4 |
| B8, B12, B13 | Alpha-like toxin Bom4 OS = Buthus occitanus mardochei | 7296.3 | (K)YGNACWCEDLPDNVPIRIPGK(C) | P59354 | 91.3 |
| B8 | Alpha-mammal toxin Bot3 (Fragment) OS = Buthus occitanus tunetanus | 8059.1 | (K)LKGESGYCQWASPYGNACYCYKLPDHVR(T) | P01485 | 83.3 |
| B8, B9, B10, B11 | Alpha-toxin Bot1 OS = Buthus occitanus tunetanus | 7268.2 | (K)DLPDNVPIRIPGK(C) | P01488 | 88.2 |
| B8 | Alpha-toxin Bot11 OS = Buthus occitanus tunetanus | 7468.5 | (R)YGNACWCYKLPDHVR(T) | P01486 | 92.0 |
| B11, B12 | Beta-toxin BotIT2 OS = Buthus occitanus tunetanus | 6917.8 | (K)WGLACWCEDLPDEK(R) | P59863 | 82.1 |
| B13 | Lipolysis-activating peptide 1-beta chain OS = Buthus occitanus tunetanus | 10,386.3 | (R)ELGILYGCK(G) | P84809 | 82.0 |
| B8, B9, B10, B11, B12 | Neurotoxin Bot2 OS = Buthus occitanus tunetanus | 7354.4 | (−)GRDAYIAQPENCVYECAK(N) | P01483 | 100.0 |
| B3, B4, B5, B6, B7, B8, B9, B13 | Potassium channel toxin alpha-KTx 9.5 OS = Buthus occitanus tunetanus | 2949 | (K)GKHAVPTCD DGVCN(C) | P84744 | 85.7 |
| B6, B7, B8, B9, B10, B11, B12, B13, B14 | Potassium channel toxin BuTXK-beta OS = Buthus occitanus israelis | 10,192.2 | (K)YAVPESTLR(T) | B8XH40 | 89.8 |
| B8 | Toxin Boma6a OS = Buthus occitanus mardochei | 7477.5 | (R)DAYCNDLCTK(N) | P60255 | 85.6 |
| Proteins Associated with other Species | |||||
| Sample/Fraction | Protein Name | MW | Sequence | Swiss-Prot ID | % Coverage |
| B8, B9, B10, B11 | Alpha-insect toxin Lqq3 OS = Leirus quinquestriatus quinquestriatus | 7240.2 | (K)YGNACWCYALPDNVPIRVPGKCH(-) | P01487 | 97.7 |
| B10 | Alpha-like toxin BmK-M1 OS = Mesobuthus martensii | 3598.4 | (-)MCIPCFTTNPNMAAK(C) | P45697 | 81.0 |
| B11, B12 | Beta-insect excitatory toxin BmK IT-AP OS = Mesobuthus martensii | 10,225.1 | (K)VYYADK(G) | O77091 | 80.8 |
| B11 | Beta-insect excitatory toxin LqhITIa OS = Leirus quinquestriatus hebraeus | 9900.8 | (K)YCDFTIIN(-) | P68721 | 81.2 |
| B12 | Beta-insect excitatory toxin LqhITIb OS = Leirus quinquestriatus hebraeus | 9958.8 | (K)KYCDFTIIN(-) | P68722 | 85.4 |
| B10, B11, B12 | Beta-insect excitatory toxin LqhITIc OS = Leirus quinquestriatus hebraeus | 9935.8 | (K)VMEISDTR(K) | P68723 | 73.7 |
| B11, B12, B13 | Beta-insect excitatory toxin LqhITId OS = Leirus quinquestriatus hebraeus | 9992.8 | (K)VYYAEK(G) | P68724 | 71.7 |
| B10, B11, B12 | Beta-toxin Isom1 OS = Isometrus vittatus | 7908.1 | (R)KKYCDYTIIN(-) | P0C5H1 | 86.2 |
| B9, B10, B11, B12 | Chlorotoxin OS = Leirus quinquestriatus quinquestriatus | 4004.8 | (-)MCMPCFTTDHQMAR(K) | P45639 | 85.8 |
| B8, B9, B10 | Chlorotoxin-like peptide OS = Androctonus australis | 3598.4 | (-)MCIPCFTTNPNMAAK(C) | P86436 | 77.8 |
| B1 | L-amino-acid oxidase (Fragment) OS= Daboia russelli siamensis | 46,371.9 | (R)IFFAGEYTANAHGWIDSTIKSGLTAAR(D) | Q4F867 | 82.4 |
| B2, B16 | Neurotoxin-1″ OS = Androctonus australis | 9060.8 | (K)DLPDNVPIK(D) | PO1479 | 84.8 |
| B13, B18 | Lipolysis-activating peptide 1-alpha chain OS = Mesobuthus martensii GN = LVP1a | 11,329.2 | (K)YYCTILGENEYCR(K) | Q6WJF5 | 86.8 |
| B14 | Neurotoxin MeuNaTx-6 OS = Buthus eupeus | 7995 | (K)PHNCVYECFDAFSSYCNGV(C) | E7CZY9 | 89.4 |
| B17 | Toxin AaHIT4 OS = Androctonus australis | 7786.0 | (K)LACYCQGAR(K) | P21150 | 96.1 |
| B19 | Beta-insect excitatory toxin LqhIT1d OS = Leiurus quinquestriatus hebraeus | 9992.8 | (K)VYYAEK(G) | P68724 | 71.7 |
| B20, B21 | Beta-toxin BmKAs1 OS = Mesobuthus martensii | 9802.7 | (K)LACYCEGAPK(S) | Q9UAC8 | 87.3 |
| B22 | Beta-insect excitatory toxin LqhIT1c OS = Leiurus quinquestriatus hebraeus | 9935.8 | (K)VMEISDTR(K) | P68723 | 73.7 |
| B12, B13 | Neurotoxin-1 OS = Androctonus australis | 9060.8 | (K)DLPDNVPIK(D) | P01479 | 84.8 |
| B8 | Potassium channel toxin alpha-KTx 15.2 OS = Mesobuthus martensii | 6206.6 | (K)AIGVAAGK(C) | Q8I0L5 | 87.0 |
| B14, B15 | Potassium channel toxin alpha-KTx 3.4 OS = Leirus quinquestriatus hebraeus | 4020.9 | (-)GVPINVK(C) | P46110 | 95.2 |
| B3, B4, B5, B6, B8, B9, B13 | Potassium channel toxin alpha-KTx 8.5 OS = Odontobuthus doriae | 3188.6 | (-)VSCEDCPEHCSTQK(A) | P0CC12 | 82.2 |
| B8, B9 | Potassium channel toxin BmTXK-beta OS = Mesobuthus martensii | 10,430.6 | (K)AIGKCEDTECK(C) | Q9NJC6 | 90.6 |
| B11 | Potassium channel toxin MeuTXKbeta3 OS = Buthus eupeus | 10,338.2 | (K)YAVPESTLR(T) | A9XE60 | 86.1 |
| B12, B13 | Toxin AaHIT4 OS = Androctonus australis | 7786.0 | (K)LACYCQGAR(K) | P21150 | 96.1 |
| B10, B11 | Toxin Aam2 OS = Androctonus amoreuxi | 9283 | (K)NGAESGYCQWFGRYGNA(C) | Q86SE0 | 77.9 |
| B8 | Toxin BmKa3 OS = Mesobuthus martensii | 9425.9 | (K)LPDKVPIR(V) | Q9GUA7 | 85.7 |
| B13 | Toxin Isom2 OS = Isometrus vittatus | 7884.75 | (K)VHYADKGYCCLLSCY(C) | P0C5H2 | 98.7 |
| B8, B9 | Toxin Lqh4 OS = Leirus quinquestriatus hebraeus | 7220.3 | (K)YGNACWCIK(L) | P83644 | 89.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hilal, I.; Khourcha, S.; Safi, A.; Hmyene, A.; Asnawi, S.; Othman, I.; Stöcklin, R.; Oukkache, N. Comparative Proteomic Analysis of the Venoms from the Most Dangerous Scorpions in Morocco: Androctonus mauritanicus and Buthus occitanus. Life 2023, 13, 1133. https://doi.org/10.3390/life13051133
Hilal I, Khourcha S, Safi A, Hmyene A, Asnawi S, Othman I, Stöcklin R, Oukkache N. Comparative Proteomic Analysis of the Venoms from the Most Dangerous Scorpions in Morocco: Androctonus mauritanicus and Buthus occitanus. Life. 2023; 13(5):1133. https://doi.org/10.3390/life13051133
Chicago/Turabian StyleHilal, Ines, Soukaina Khourcha, Amal Safi, Abdelaziz Hmyene, Syafiq Asnawi, Iekhsan Othman, Reto Stöcklin, and Naoual Oukkache. 2023. "Comparative Proteomic Analysis of the Venoms from the Most Dangerous Scorpions in Morocco: Androctonus mauritanicus and Buthus occitanus" Life 13, no. 5: 1133. https://doi.org/10.3390/life13051133