
Mitochondrial DNA Corroborates the Genetic Variability of Clarias Catfishes (Siluriformes, Clariidae) from Cameroon

 , , , , and
, , , , and 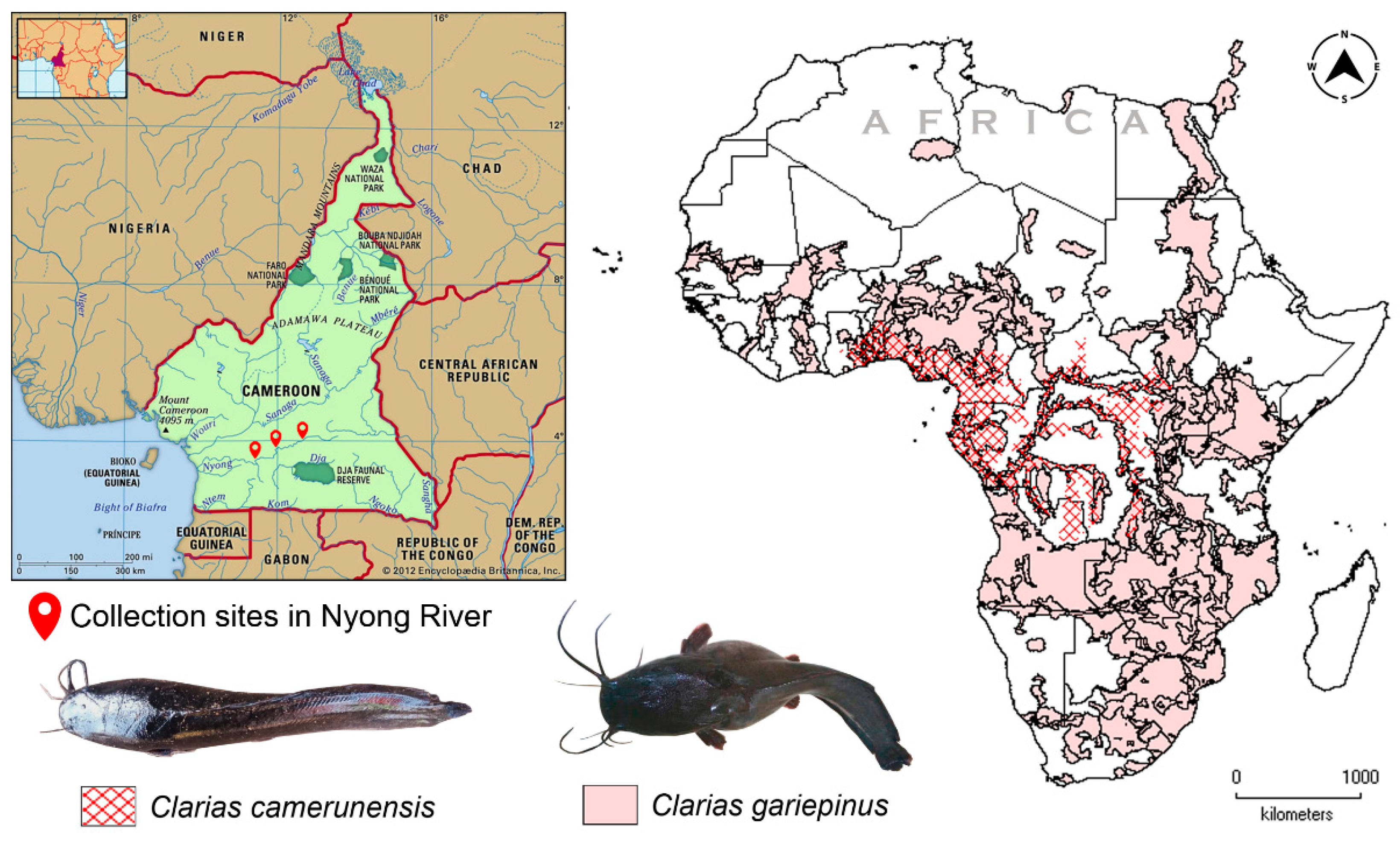{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling and Identification
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Sequence Quality Check
2.4. Dataset Construction and Analyses
3. Results
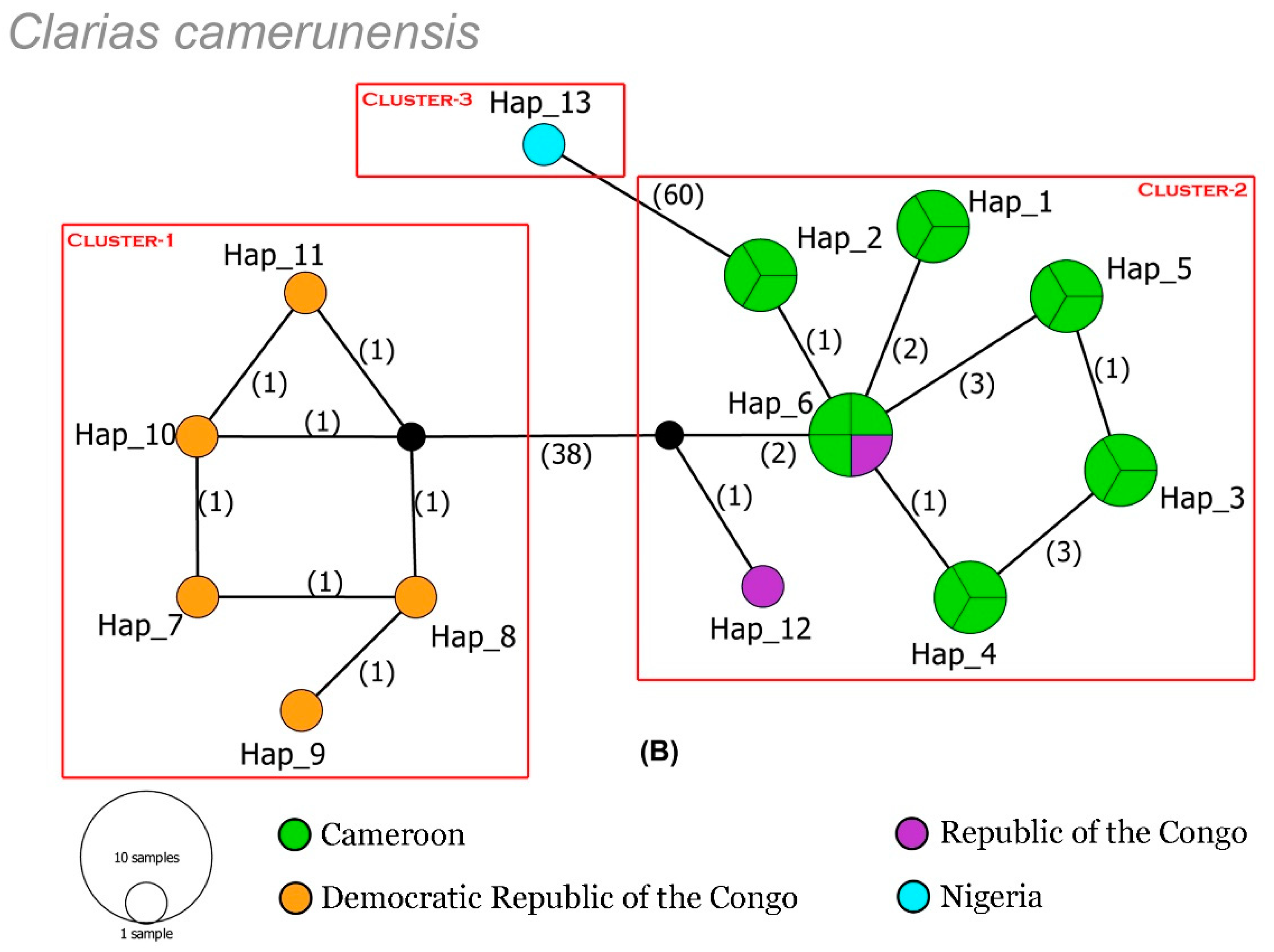
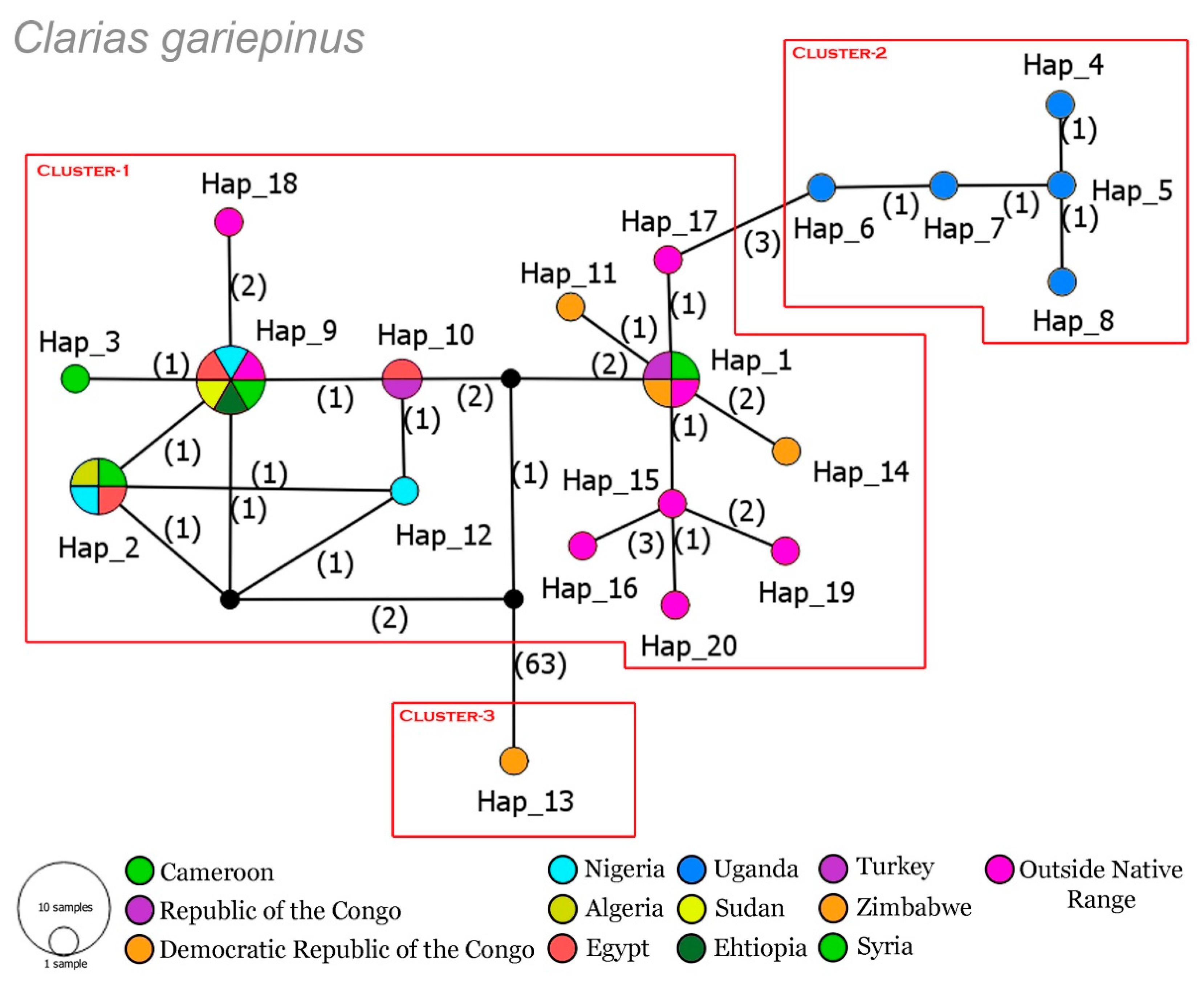
3.1. Genetic Divergence and Haplotype Distribution
3.2. MOTU Estimation and Phylogenetic Relationship
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fricke, R.; Eschmeyer, W.N.; van der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References. Electronic Version. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 4 March 2023).
- Froese, R.; Pauly, D. (Eds.) FishBase. World Wide Web Electronic Publication. Version (08/2022). Available online: www.fishbase.org (accessed on 4 March 2023).
- Low, B.W.; Ng, H.H.; Tan, H.H. Two new species of blackwater catfishes (Siluriformes: Siluridae and Clariidae) from the Natuna Archipelago, Indonesia. Raffles Bull. Zool. 2022, 70, 385–396. [Google Scholar]
- Bernt, M.J.; Stiassny, M.L.J. A New Species of Air-Breathing Catfish (Clariidae: Clarias) from Salonga National Park, Democratic Republic of the Congo. Am. Mus. Novit. 2022, 3990, 1–20. [Google Scholar] [CrossRef]
- Teugels, G.G. A systematic revision of the African species of the genus Clarias (Pisces; Clariidae). Annales—Musee Royal de l’Afrique Centrale. Sci. Zool. (Belg.) 1986, 247, 199. [Google Scholar]
- Skelton, P.H. A Complete Guide to the Freshwater Fishes of Southern Africa; RSA: Struik Publishers: Cape Town, South Africa, 2001. [Google Scholar]
- Seegers, L. The Catfishes of Africa; A Handbook for Identification and Maintenance; Aqualog Verlag A.C.S. GmbH: Rodgau, Germany, 2008; 604p. [Google Scholar]
- Hanssens, M. A review of the Clarias species (Pisces; Siluriformes) from the Lower Congo and the Pool Malebo. J. Afr. Zool. 2009, 5, 27–40. [Google Scholar]
- Vitule, J.R.; Umbria, S.C.; Aranha, J.M.R. Introduction of the African catfish Clarias gariepinus (Burchell, 1822) into Southern Brazil. Biol. Invasions 2006, 8, 677–681. [Google Scholar] [CrossRef]
- Weyl, O.L.; Dagam, V.S.; Ellender, B.R.; Vitule, J.R. A review of Clarias gariepinus invasions in Brazil and South Africa. J. Fish. Biol. 2016, 89, 386–402. [Google Scholar] [CrossRef] [PubMed]
- Merga, L.B.; Redondo-Hasselerharm, P.E.; Van den Brink, P.J.; Koelmans, A.A. Distribution of microplastic and small macroplastic particles across four fish species and sediment in an African lake. Sci. Total. Environ. 2020, 741, 140527. [Google Scholar] [CrossRef]
- Parvez, I.; Rumi, R.A.; Ray, P.R.; Hassan, M.M.; Sultana, S.; Pervin, R.; Suwanno, S.; Pradit, S. Invasion of African Clarias gariepinus Drives Genetic Erosion of the Indigenous C. batrachus in Bangladesh. Biology 2022, 11, 252. [Google Scholar] [CrossRef] [PubMed]
- Erasmus, J.H.; Zimmermann, S.; Smit, N.J.; Malherbe, W.; Nachev, M.; Sures, B.; Wepener, V. Human health risks associated with consumption of fish contaminated with trace elements from intensive mining activities in a peri-urban region. Sci. Total. Environ. 2022, 825, 154011. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-2. Available online: https://www.iucnredlist.org (accessed on 4 March 2023).
- Lind, C.E.; Ponzoni, R.W.; Nguyen, N.H.; Khaw, H.L. Selective Breeding in Fish and Conservation of Genetic Resources for Aquaculture. Reprod. Dom. Anim. 2012, 47, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, J.C.; Huber, C.D. The inflated significance of neutral genetic diversity in conservation genetics. Proc. Natl. Acad. Sci. USA 2021, 118, e2015096118. [Google Scholar] [CrossRef]
- Barasa, J.E.; Mdyogolo, S.; Abila, R.; Grobler, J.P.; Skilton, R.A.; Bindeman, H.; Njahira, M.N.; Chemoiwa, E.J.; Dangasuk, O.G.; Kaunda-Arara, B.; et al. Genetic diversity and population structure of the African catfish, Clarias gariepinus (Burchell, 1822) in Kenya: Implication for conservation and aquaculture. Belg. J. Zool. 2017, 147, 105–127. [Google Scholar]
- Awodiran, M.O.; Afolabi, O. Genetic Diversity in Cultured and Wild Population of Clarias gariepinus (Burchell, 1822) in Nigeria Using Random Amplified Polymorphic DNA (RAPD) and Microsatellite DNA. Fish. Aqua J. 2018, 9, 1000247. [Google Scholar] [CrossRef]
- Isa, S.I. Development of Genetic Improvement in the African Catfish (Clarias gariepinus, Burchell, 1822). Ph.D. Thesis, Institute of Aquaculture, Faculty of Natural Sciences, University of Stirling, Scotland, UK, 2019. [Google Scholar]
- Tsaparis, D.; Lecocq, T.; Kyriakis, D.; Oikonomaki, K.; Fontaine, P.; Tsigenopoulos, C.S. Assessing Genetic Variation in Wild and Domesticated Pikeperch Populations: Implications for Conservation and Fish Farming. Animals 2022, 12, 1178. [Google Scholar] [CrossRef]
- Chala, B.; Erko, B.; Animut, A.; Degarege, A.; Petros, B. Assessment of Clarias gariepinus as a biological control agent against mosquito larvae. BMC Ecol. 2016, 16, 27. [Google Scholar] [CrossRef]
- Swaleh, S.B.; Banday, U.Z.; Asadi, M.A.; Usmani, N. Biochemical profile and gene expression of Clarias gariepinus as a signature of heavy metal stress. Environ. Pollut. 2020, 264, 114693. [Google Scholar] [CrossRef]
- Van Steenberge, M.W.; Vanhove, M.P.M.; Manda, A.C.; Larmuseau, M.H.D.; Swart, B.L.; Khang’Mate, F.; Arndt, A.; Hellemans, B.; Houdt, J.V.; Micha, J.C.; et al. Unravelling the evolution of Africa’s drainage basins through a widespread freshwater fish, the African sharptooth catfish Clarias gariepinus. J. Biogeogr. 2020, 47, 1739–1754. [Google Scholar] [CrossRef]
- Lakra, K.C.; Banerjee, T.K.; Lal, B. Coal mine effluent-induced metal bioaccumulation, biochemical, oxidative stress, metallothionein, and histopathological alterations in vital tissues of the catfish, Clarias batrachus. Environ. Sci. Pollut. Res. 2021, 28, 25300–25315. [Google Scholar] [CrossRef]
- Okella, H.; Ikiriza, H.; Ochwo, S.; Ajayi, C.O.; Ndekezi, C.; Nkamwesiga, J.; Kaggwa, B.; Aber, J.; Mtewa, A.G.; Koffi, T.K.; et al. Identification of antimicrobial peptides isolated from the skin mucus of African Catfish, Clarias gariepinus (Burchell, 1822). Front. Microbiol. 2021, 12, 794631. [Google Scholar] [CrossRef] [PubMed]
- Agnese, J.F.; Teugels, G.G. Insight into the phylogeny of African Clariidae (Teleostei, Siluriformes): Implications for their body shape evolution, biogeography and taxonomy. Mol. Phylogenet. Evol. 2005, 36, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Pouyaud, L.; Sudarto, T.; Paradis, E. The phylogenetic structure of habitat shift and morphological convergence in Asian Clarias (Teleostei, Siluriformes: Clariidae). J. Zoolog. Syst. Evol. Res. 2009, 47, 344–356. [Google Scholar] [CrossRef]
- Nazia, A.K.; Siti Azizah, M.N. Isolation of microsatellites in the bighead catfish, Clarias macrocephalus and cross-amplification in selected Clarias species. Mol. Biol. Rep. 2014, 41, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Barasa, J.E.; Abila, R.; Grobler, J.P.; Agaba, M.; Chemoiwa, E.J.; Kaunda-Arara, B. High genetic diversity and population differentiation in Clarias gariepinus of Yala Swamp: Evidence from mitochondrial DNA sequences. J. Fish. Biol. 2016, 89, 2557–2570. [Google Scholar] [CrossRef] [PubMed]
- Tiknaik, A.; Khedkar, C.; Khedkar, G.; Prakash, B.; Mamatha, D.M.; Sangale, D.; Kalyankar, A. Microsatellite Genotyping Corroborated Loss of Genetic Diversity in Clarias batrachus as a Result of Lack of Regulatory Reforms in Aquaculture. Biochem. Genet. 2020, 58, 595–616. [Google Scholar] [CrossRef]
- Kushwaha, B.; Kumar, R.; Agarwal, S.; Pandey, M.; Nagpure, N.S.; Singh, M.; Srivastava, S.; Joshi, C.G.; Das, P.; Sahoo, L.; et al. Assembly and variation analyses of Clarias batrachus mitogenome retrieved from WGS data and its phylogenetic relationship with other catfishes. Meta Gene 2015, 5, 105–114. [Google Scholar] [CrossRef]
- Kappas, I.; Vittas, S.; Pantzartzi, C.N.; Drosopoulou, E.; Scouras, Z.G. A Time-Calibrated Mitogenome Phylogeny of Catfish (Teleostei: Siluriformes). PLoS ONE 2016, 11, e0166988. [Google Scholar]
- Li, N.; Bao, L.; Zhou, T.; Yuan, Z.; Liu, S.; Dunham, R.; Li, Y.; Wang, K.; Xu, X.; Jin, Y.; et al. Genome sequence of walking catfish (Clarias batrachus) provides insights into terrestrial adaptation. BMC Genom. 2018, 19, 952. [Google Scholar] [CrossRef]
- Elberri, A.I.; Galal-Khallaf, A.; Gibreel, S.E.; El-Sakhawy, S.F.; El-Garawani, I.; El-Sayed Hassab El Nabi, S.; Mohammed-Geba, K. DNA and eDNA-based tracking of the North African sharptooth catfish Clarias gariepinus. Mol. Cell. Probes 2020, 51, 101535. [Google Scholar] [CrossRef]
- Wong, L.L.; Peatman, E.; Lu, J.; Kucuktas, H.; He, S.; Zhou, C.; Na-nakorn, U.; Liu, Z. DNA barcoding of catfish: Species authentication and phylogenetic assessment. PLoS ONE 2011, 6, e17812. [Google Scholar] [CrossRef]
- Decru, E.; Moelants, T.; De Gelas, K.; Vreven, E.; Verheyen, E.; Snoeks, J. Taxonomic challenges in freshwater fishes: A mismatch between morphology and DNA barcoding in fish of the north-eastern part of the Congo basin. Mol. Ecol. Resour. 2016, 16, 342–352. [Google Scholar] [CrossRef]
- Iyiola, O.A.; Nneji, L.M.; Mustapha, M.K.; Nzeh, C.G.; Oladipo, S.O.; Nneji, I.C.; Okeyoyin, A.O.; Nwani, C.D.; Ugwumba, O.A.; Ugwumba, A.A.A.; et al. DNA barcoding of economically important freshwater fish species from north-central Nigeria uncovers cryptic diversity. Ecol. Evol. 2018, 8, 6932–6951. [Google Scholar] [CrossRef] [PubMed]
- Popoola, M.O.; Schedel, F.D.B.; Hebert, P.D.; Schliewen, U.K. First DNA barcode library for the ichthyofauna of the Jos Plateau (Nigeria) with comments on potential undescribed fish species. PeerJ 2022, 10, e13049. [Google Scholar] [CrossRef] [PubMed]
- De Alwis, P.S.; Kundu, S.; Gietbong, F.Z.; Amin, M.H.F.; Lee, S.-R.; Kim, H.-W.; Kim, A.R. Mitochondriomics of Clarias Fishes (Siluriformes: Clariidae) with a New Assembly of Clarias camerunensis: Insights into the Genetic Characterization and Diversification. Life 2023, 13, 482. [Google Scholar] [CrossRef]
- Baldwin, C.C.; Mounts, J.H.; Smith, D.G.; Weight, L.A. Genetic identification and color descriptions of early life-history stages of Belizean Phaeoptyx and Astrapogon (Teleostei: Apogonidae) with comments on identification of adult Phaeoptyx. Zootaxa 2009, 2008, 1–22. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Clement, M.; Snell, Q.; Walker, P.; Posada, D.; Crandall, K. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. Popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Simon, Y.; Ho, W.; Stephane, G.S. PartitionFinder: Combined Selection of Partitioning Schemes and Substitution Models for Phylogenetic Analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Jaa, N. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef]
- Vences, M.; Miralles, A.; Brouillet, S.; Ducasse, J.; Fedosov, A.; Kharchev, V.; Kostadinov, I.; Kumari, S.; Patmanidis, S.; Scherz, M.D. iTaxoTools 0.1: Kickstarting a specimen-based software toolkit for taxonomists. Megataxa 2021, 6, 77–92. [Google Scholar] [CrossRef]
- MINEP. Cameroon’s Fourth National Report on the Convention on Biological Diversity; MINEP: Yaounde, Cameroon, 2009. [Google Scholar]
- Ngala, G.N.; Gryzenhout, M. Biodiversity and conservation in Cameroon. Mycol. Balc. 2010, 7, 65–72. [Google Scholar]
- Tchouto, M.G.P.; Yemefack, M.; De Boer, W.F.; De Wilde, J.J.F.E.; Van Der Maesen, L.J.G.; Cleef, A.M. Biodiversity Hotspots and Conservation Priorities in the Campo-Ma’an Rain Forests, Cameroon. Biodivers. Conserv. 2006, 15, 1219–1252. [Google Scholar] [CrossRef]
- Takem, B.M.; Kaffo, C.; Fish, L. “Protected area” coverage in Cameroon on the eve of the Convention on Biological Diversity 2010 target. Int. For. Rev. 2010, 12, 231–239. [Google Scholar] [CrossRef]
- Burridge, C.P.; Craw, D.; Jack, D.C.; King, T.M.; Waters, J.M. Does fish ecology predict dispersal across a river drainage divide? Evolution 2008, 62, 1484–1499. [Google Scholar] [CrossRef]
- Mbi, B.M.T.; Lebga, A.K. ‘Protected Areas in Cameroon at the Mercy of the 2035 Emergent Project’. In Natural Resources Management and Biological Sciences; Rhodes, E.R., Naser, H., Eds.; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Barbarossa, V.; Bosmans, J.; Wanders, N.; King, H.; Bierkens, M.F.P.; Huijbregts, M.A.J.; Schipper, A.M. Threats of global warming to the world’s freshwater fishes. Nat. Commun. 2021, 12, 1701. [Google Scholar] [CrossRef]
- Goodier, S.A.M.; Cotterill, F.P.D.; O’Ryan, C.; Skelton, P.H.; de Wit, M.J. Cryptic diversity of African tigerfish (genus Hydrocynus) reveals palaeogeographic signatures of linked neogene geotectonic events. PLoS ONE 2011, 6, e28775. [Google Scholar] [CrossRef]
- Charlesworth, D.; Willis, J.H. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Day, J.J.; Peart, C.R.; Brown, K.J.; Friel, J.P.; Bills, R.; Moritz, T. Continental diversification of an African Catfish Radiation (Mochokidae: Synodontis). Syst. Biol. 2013, 62, 351–365. [Google Scholar] [CrossRef]
- Day, J.J.; Fages, A.; Brown, K.J.; Vreven, E.J.; Stiassny, M.L.J.; Bills, R.; Friel, J.P.; Rüber, L. Multiple independent colonizations into the Congo Basin during the continental radiation of African Mastacembelus spiny eels. J. Biogeogr. 2017, 44, 2308–2318. [Google Scholar] [CrossRef]
- Sholihah, A.; Delrieu-Trottin, E.; Condamine, F.L.; Wowor, D.; Rüber, L.; Pouyaud, L.; Agnèse, J.F.; Hubert, N. Impact of Pleistocene Eustatic Fluctuations on Evolutionary Dynamics in Southeast Asian Biodiversity Hotspots. Syst. Biol. 2021, 70, 940–960. [Google Scholar] [CrossRef] [PubMed]
- Säisä, M.; Salminen, M.; Koljonen, M.L.; Ruuhijärvi, J. Coastal and freshwater pikeperch (Sander lucioperca) populations differgenetically in the Baltic Sea basin. Hereditas 2010, 147, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Teletchea, F.; Fontaine, P. Levels of domestication in fish: Implications for the sustainable future of aquaculture. Fish. Fish. 2014, 15, 181–195. [Google Scholar] [CrossRef]
- Mignon-Grasteau, S.; Boissy, A.; Bouix, J.; Faure, J.-M.; Fisher, A.D.; Hinch, G.N.; Jensen, P.; Le Neindre, P.; Mormède, P.; Prunet, P.; et al. Genetics of adaptation and domestication in livestock. Livest. Prod. Sci. 2005, 93, 3–14. [Google Scholar] [CrossRef]
- Hoffman, J.I.; Simpson, F.; David, P.; Rijks, J.M.; Kuiken, T.; Thorne, M.A.; Lacy, R.C.; Dasmahapatra, K.K. High-throughput sequencing reveals inbreeding depression in a natural population. Proc. Natl. Acad. Sci. USA 2014, 111, 3775–3780. [Google Scholar] [CrossRef]
- Tine, M.; Ndiaye, F.; Bale, K.; Magblenou, L.D.; Sene, M.A. Effects of inbreeding depression on the success of artificial reproduction in the African catfish Clarias gariepinus (BURCHELL, 1822). Int. J. Aquac. Fish. Sci. 2022, 8, 045–053. [Google Scholar]
- Cross, T.F. Genetic implications of translocation and stocking of fish species, with particular reference to Western Australia. Aquac. Res. 2000, 31, 83–94. [Google Scholar] [CrossRef]
- Nousias, O.; Tzokas, K.; Papaharisis, L.; Ekonomaki, K.; Chatziplis, D.; Batargias, C.; Tsigenopoulos, C.S. Genetic Variability, Population Structure, and Relatedness Analysis of Meagre Stocks as an Informative Basis for New Breeding Schemes. Fishes 2021, 6, 78. [Google Scholar] [CrossRef]
- Senanan, W.; Kapunscinski, A.R.; Na-Nakorn, U.; Miller, L.M. Genetic impacts of hybrid catfish farming (Clarias macrocephalus × C. gariepinus) on native catfish populations in central Thailand. Aquaculture 2004, 235, 167–184. [Google Scholar] [CrossRef]
- Tan, M.T.C.; Jumawan, J.C.; Quilang, J.P. Low genetic diversity in Clarias macrocephalus Günther, 1864 (Siluriformes: Clariidae) populations in the Philippines and its implications for conservation and management. J. Threat. Taxa 2016, 8, 8849–8859. [Google Scholar] [CrossRef]
- D’Ambrosio, J.; Phocas, F.; Haffray, P.; Bestin, A.; Brard-Fudulea, S.; Poncet, C.; Quillet, E.; Dechamp, N.; Fraslin, C.; Charles, M.; et al. Genome-wide estimates of genetic diversity, inbreeding and effective size of experimental and commercial rainbow trout lines undergoing selective breeding. Genet. Sel. Evol. 2019, 51, 26. [Google Scholar] [CrossRef] [PubMed]
- Wachirachaikarn, A.; Na-Nakorn, U. Genetic diversity of the North African catfish, Clarias gariepinus (Burchell, 1822) hatchery stocks in Thailand. Sci. Asia 2019, 45, 301–308. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kundu, S.; De Alwis, P.S.; Binarao, J.D.; Lee, S.-R.; Kim, A.R.; Gietbong, F.Z.; Yi, M.; Kim, H.-W. Mitochondrial DNA Corroborates the Genetic Variability of Clarias Catfishes (Siluriformes, Clariidae) from Cameroon. Life 2023, 13, 1068. https://doi.org/10.3390/life13051068
Kundu S, De Alwis PS, Binarao JD, Lee S-R, Kim AR, Gietbong FZ, Yi M, Kim H-W. Mitochondrial DNA Corroborates the Genetic Variability of Clarias Catfishes (Siluriformes, Clariidae) from Cameroon. Life. 2023; 13(5):1068. https://doi.org/10.3390/life13051068
Chicago/Turabian StyleKundu, Shantanu, Piyumi S. De Alwis, Jerome D. Binarao, Soo-Rin Lee, Ah Ran Kim, Fantong Zealous Gietbong, Myunggi Yi, and Hyun-Woo Kim. 2023. "Mitochondrial DNA Corroborates the Genetic Variability of Clarias Catfishes (Siluriformes, Clariidae) from Cameroon" Life 13, no. 5: 1068. https://doi.org/10.3390/life13051068