1. Introduction
The set of microorganisms that usually live inside the human body, also called human microbiota, generates the microflora. Its genomic constitution and its by-products are called the human microbiome [
1]. This microbiome is unique for each individual and is never stable, due to environmental, nutritional and ecological changes. Furthermore, in everyone, due to environmental variations, each body site is home to a distinct microbial ecosystem [
2]. Microorganisms can adapt to different survival conditions.
Bacteroidetes and
Firmicutes, followed by
Proteobacteria,
Fusobacteria,
Tenericutes,
Actinobacteria and
Verrucomicrobia were reported to be the most dominant, constituting about 90% of the total microbial population in humans [
3]. A substantial amount of information is readily available regarding the human microbiome; much less is known regarding the human mycobiome, i.e., the type and number of fungal species that inhabit the human ecological niches. In fact, studies focusing on yeast infections are scarce compared with studies focusing on bacterial infections (
Figure 1).
Several human ecological niches are colonized by millions of different microorganisms, of which the gut is by far the most studied. The gut is recognized as a virtual organ closely associated with the health and longevity of the host. The gut microbiome has both beneficial and adverse impacts on gut tissue homeostasis [
4]. From a very early stage of life (after 3 years of age), the microbial composition of the intestinal microbiome varies over time, never managing to remain 100% stable [
4]. The intestinal mycobiome is constituted in abundance by fungi, such as
Candida spp.,
Aspergillus spp.,
Fusarium spp. and
Cryptococcus spp., which can have a pathogenic effect on the host [
5].
Although most oral microbiome studies have been performed on bacteria, there has been a recent surge in studies on oral fungal communities [
6]. Over the past few years, exploration of the composition of the mycobiome in oral samples has shown that they were colonized by
Candida spp.,
Cladosporium spp.,
Saccharomyces spp.,
Penicillium spp.,
Malassezia spp.,
Aspergillus spp.,
Cryptococcus spp.,
Rhodotorula spp. and
Trichosporon spp., among others [
6].
Characterization of fungal communities over time has revealed that mode of delivery influences colonization by
Candida species, where the relative abundance of
Candida albicans is highest on the skin of vaginally delivered infants [
7]. On the other hand, characterization of the fungal communities in the skin has revealed abundant colonization by
Malassezia spp.,
Candida spp.,
Cladosporium spp.,
Fusarium spp. and
Cryptococcus spp. [
8]. In the vagina, although much less abundant than bacteria (values less than 1%), fungi, namely
Candida albicans, have been identified as the most predominant microorganism affecting vaginal health. One study demonstrated the presence of
Candida,
Clavispora lusitanie,
Malassezia,
Rhodotorula,
Aspergillus and
Leptosphaerulina as some of the most prevalent fungi in the vaginal mucosa [
9].
With this review, we intend to obtain knowledge about the composition and role of commensal fungi in human health, with special focus on fungal microorganisms that can colonize the skin, vagina and mucous membranes.
2. Materials and Methods
This review was based on a literature search to obtain a blend of different types of systematic reviews, as well as to obtain information about yeasts in different niches (focusing mainly on the intestinal, vaginal and oral mucosae). The articles and dissertations mentioned were obtained through the following platforms: Scielo; PubMed and Web of Science. The titles of the subjects and the keywords chosen were mycobiome, intestinal microbiota, skin yeasts, fungi, vaginal microbiota, oral microbiota, vulvovaginal diseases and skin diseases. The inclusion criteria were language (Portuguese and English), text availability and preferred publication date (last 5 years).
3. Results
3.1. Frequent Colonizers of Human Ecological Niches
The different mycobiomes in the human body not only influence a particular niche, but rather, a range of niches in the human body, due to the interrelationships between the different microbial communities.
In recent years, there has been an increase in human invasive fungal infections, mostly caused by pathogens such as
Candida spp.; however, some less common ones, such as
Rhodotorula spp., have been recognized as emerging pathogens [
10]. Transmission of mycobioma of the skin and gastrointestinal tract is thought to be via the vaginal route during delivery, composed mostly of Candida albicans [
11].
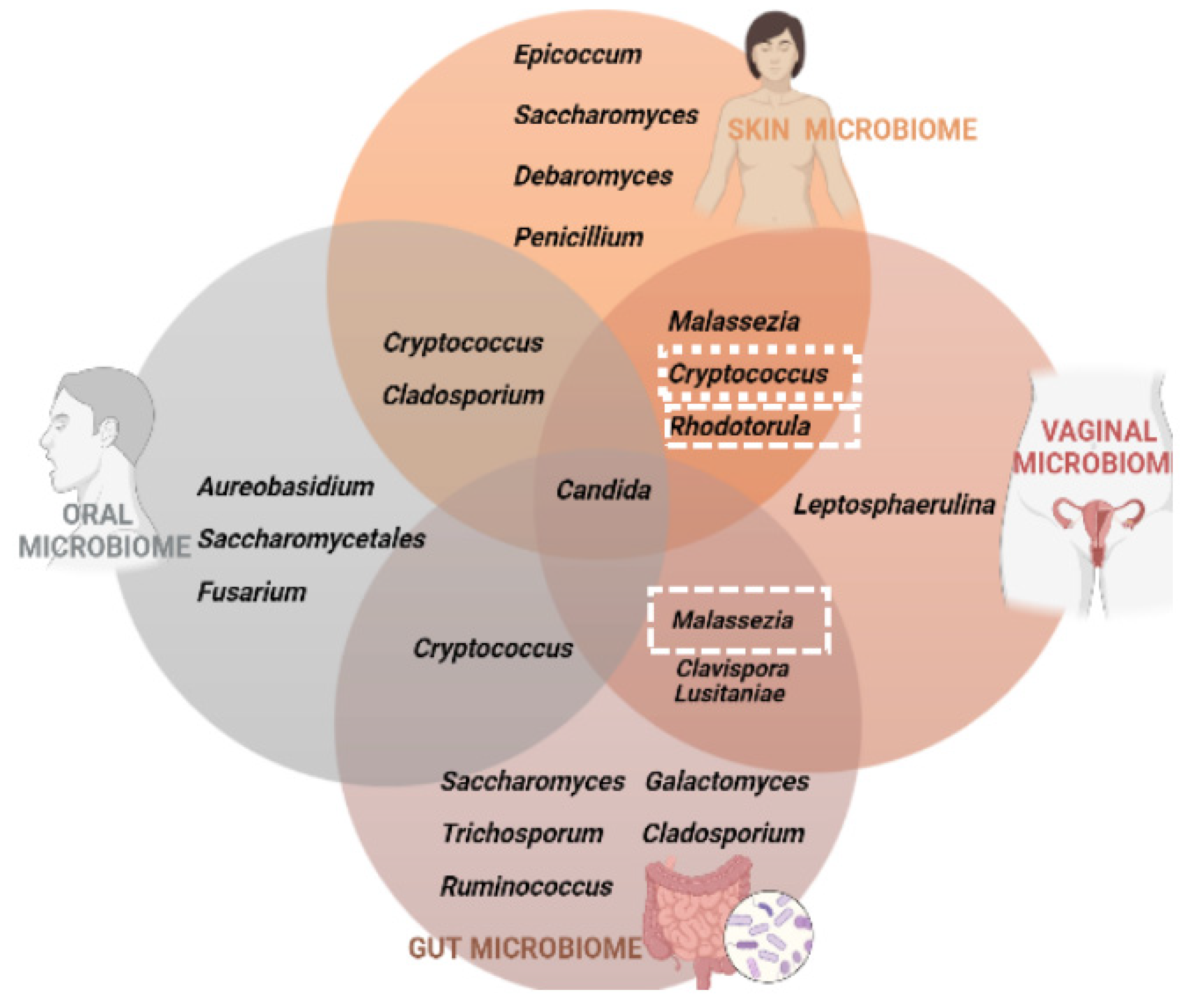
Although the skin is the niche where
Malassezia is most commonly found, in recent years it has been identified in other unrelated niches, including the human gut, breast milk and internal organs, including those of the central nervous system [
10]. Transmission of
Malassezia spp. by this way is suggested, although there is controversy as to whether transmission is initiated from the placenta [
12]. Therefore, the fungal diversity of the skin and gut of newborn babies is derived from the maternal mycobioma [
13].
Cryptococcus neoformans is a known human pathogen, particularly in the immunocompromised host. However, other species of this genus can be also found colonizing several human niches [
14]. In
Figure 2, we summarize the fungal communities that inhabit the four different niches under analysis. We were able to conclude that
Candida spp. is the common fungus present in the microbiota of the skin, vagina, intestine and oral cavity; on the other hand, the three other yeasts analyzed in this review, that correspond to the ones that are frequent colonizers of the human body but also are human pathogens, do not inhabit a niche only. Therefore, our research in this subject has revealed that these four genera possibly comprise opportunistic yeast species.
3.1.1. Candida spp.
This genus is one of the most commonly found in the normal human microbiota, managing to colonize niches such as oral mucous membranes, skin, gastrointestinal, genital and urinary tracts without causing infections. However, in immunocompromised people or people with chronic illnesses, this yeast can become pathogenic, thus causing infections called candidiasis [
15]. Within this genus, there are species that are one of the most frequent causes of opportunistic infections,
Candida albicans, the drug resistant,
C. glabrata, the new global threat to public health,
C. auris and other emerging species such as
C. tropicalis,
C. parapsilosis and
C. krusei [
16,
17].
C. albicans is a yeast that can reside in the human body while also living in certain environmental reservoirs [
18]. Disturbances induced either by antibiotics, immune system anomalies, alterations in the microbiome and/or alterations in the integrity of the mucocutaneous barrier allow
Candida spp. to become an opportunistic pathogen in the context of a series of virulence factors [
19,
20].
C. albicans colonization has been found in the vaginal niche by culture-dependent methods in approximately 20% to 30% of asymptomatic women and by molecular methods in about 65% [
21]. When the infection is symptomatic, it is defined as vulvovaginal candidosis (or candidosis), 80% to 92% of which are caused by
C. albicans [
22].
The success in the occurrence of infections comes from specific characteristics of this genus. One is the change from a yeast form to a branched hyphal form. As a rule, the yeast form is associated with commensalism, dissemination, or transmission of the infection [
23]. The balance between the two forms is maintained by the responses of the innate immune system [
24].
3.1.2. Cryptococcus spp.
The genus
Cryptococcus is a group of capsulated opportunistic fungi. Of the 37 species discovered,
C. neoformans and
C. gattii are the species that most cause pathogenesis in man [
14].
Cryptococcus spp. is an environmental yeast and is therefore very recurrently found in nature (soil, animal feces, among others).
C. neoformans essentially affects immunocompromised individuals (elderly people, patients with HIV/AIDS and recently transplanted organs) in addition to causing infections of the central nervous system [
25]. It is estimated that annually, 223,000 people with HIV/AIDS develop cryptococcal meningitis, of which 181,000 eventually die [
26].
C. gatti manages to infect both immunocompetent and immunosuppressed individuals, obtaining a mortality rate of almost 33% [
27]. That said, this genus has become a lethal pathogen that poses a serious threat to public health [
27].
3.1.3. Malassezia spp.
This yeast is included in the genus Basidiomycota, class
Malasseziomycetes and family
Malasseziaceae [
28].
Malassezia, a commensal yeast that is lipophilic and lipid dependent for its growth, except
M. pachydermatis, is the main component of the fungal skin microbiota of many mammals, corresponding to more than 90% of the total fungal population in the skin niche [
29].
The mechanisms by which these yeasts can trigger diseases are poorly studied and, therefore, are not yet clearly identified. However, the proposed hypothesis is that the diseases can be induced by direct invasion of fungal filament tissue or indirectly by immunological and metabolic mechanisms [
30,
31,
32,
33].
Studies already carried out on the composition of the fungal microbiota of breast milk revealed two new pieces of information [
32,
34]: in its composition, the proportion of fungi is much greater than that of bacteria (contrary to what is found in many other niches) and among the fungi found, 40% of the total of genera identified by pyrosequencing are represented by
Malassezia.
Different species of
Malassezia spp. can lead to different clinical manifestations, which can range from hypopigmentation without visible inflammation, to eczema with scaling and inflammation. Of all the skin diseases, although still controversial, malassezia is a pathogenic agent of pityriasis versicolor and malassezia folliculitis. The discovery of this yeast in seborrheic areas still needs further study since its commensal state is still difficult to distinguish from its pathogenic phase [
35]. Scientific evidence for the role of
Malassezia in psoriasis and atopic dermatitis is less robust than for other diseases [
36].
3.1.4. Rhodotorula spp.
The pathogenic role and its relevance in the intestinal microbiota have not been studied nor seen as important by researchers, although they are frequently found in human stool samples [
37,
38]. However, human fungal infections by
Rhodotorula spp. have been increasing in recent years [
39]. Until 2018, clinical cases have been reported, where of the 40 species,
R. mucilaginosa is the most common cause of human infection (72%), followed by
R. glutinis (21%) and
R. minuta (7%) [
40,
41,
42].
This yeast can be found as contaminants of the skin, nails, lungs, urine, feces, central nervous system and blood. The infection can be installed endogenously or exogenously, as well as by the hands of health professionals, contaminated materials and inanimate environmental sources [
43]. In addition to these transmission routes,
Rhodotorula spp., being an environmental yeast, can be found in nature by isolating it from environmental sources such as air, soil and plants [
44].
3.2. Mycobiomes in Various Body Niches and Their Associated Diseases
It is estimated that there are about three million species of fungi, of which about 300 cause infections in humans (
Table 1) [
45]. In studies previously performed, the authors identified about 66 different genres of fungi present, the most prevalent being
Candida,
Cladosporium and
Saccharomyces [
46].
The ecological relationships between bacteria, archaea, viruses and fungi maintain the host’s equilibrium and are part of several vital processes, such as nutrition and protection against pathogens. Being one of the largest eukaryotic kingdoms, fungi have a variety of life cycles with adaptations in metabolism and morphogenesis that allow them to adapt to environmental changes and give them survival throughout the human body, such as the intestine, skin, oral cavity, urogenital tract and digestive system [
47]. There is a correlation between the occurrence of changes in the fungal community and the diseases caused in humans.
Table 1.
Illnesses that are caused by colonizing fungi. The upward and downward arrows represent the increase and decrease, respectively, of fungi colonisation in the various diseases.
Table 1.
Illnesses that are caused by colonizing fungi. The upward and downward arrows represent the increase and decrease, respectively, of fungi colonisation in the various diseases.
| Human Ecological Niches | Diseases | Yeast Colonization | References |
|---|
| Gut | Inflammatory bowel disease (Crohn’s disease) | ![Life 13 00924 i001]() C. albicans, C. tropicalis, C. glabrata, Aspergillus clavatus, C. neoformans, Cyberlindnera jadinii, Clavispora lusitaniae, Debaryomyces hansenii, Kluyveromyces marxianus C. albicans, C. tropicalis, C. glabrata, Aspergillus clavatus, C. neoformans, Cyberlindnera jadinii, Clavispora lusitaniae, Debaryomyces hansenii, Kluyveromyces marxianus
![Life 13 00924 i002]() Saccharomyces cerevisiae Saccharomyces cerevisiae | [48,49,50,51] |
| Irritable bowel syndromes (IBS) | ![Life 13 00924 i001]() Candida spp., Malassezia spp., Cladosporium spp., Saccharomyces cerevisiae Candida spp., Malassezia spp., Cladosporium spp., Saccharomyces cerevisiae
![Life 13 00924 i002]() Mycosphaerella spp., Aspergillus spp., Sporidiobolus spp., Pandora spp. Mycosphaerella spp., Aspergillus spp., Sporidiobolus spp., Pandora spp. | [52,53] |
| Colorectal cancer | ![Life 13 00924 i001]() Trichosporon spp., Malassezia spp., Trichosporon spp., Malassezia spp., | [54] |
| Obesity | ![Life 13 00924 i001]() Candida, Nakaseomyces, Penicillium, Porphyromonas, Campylobacter, Bacteroides, Staphylococcus, Parabacteroides, Dialister and Ruminococcus Candida, Nakaseomyces, Penicillium, Porphyromonas, Campylobacter, Bacteroides, Staphylococcus, Parabacteroides, Dialister and Ruminococcus
![Life 13 00924 i002]() Mucor racemosus and M. fuscus Mucor racemosus and M. fuscus | [55,56] |
| Diabetes | ![Life 13 00924 i001]() C. albicans and Saccharomyces spp. In type 1 Diabetes C. albicans and Saccharomyces spp. In type 1 Diabetes
![Life 13 00924 i001]() C. albicans, Cladosporium spp., Meyerozyma spp., Mortierella spp. and Aspergillus spp. In type 2 diabetes C. albicans, Cladosporium spp., Meyerozyma spp., Mortierella spp. and Aspergillus spp. In type 2 diabetes | [55,57,58] |
| Skin | Pityriasis versicolor | ![Life 13 00924 i001]() M. globosa and M. sympodialis and M. furfur. M. globosa and M. sympodialis and M. furfur. | [59] |
| Seborrheic dermatitis | ![Life 13 00924 i001]() Malassezia spp. Malassezia spp. | [60] |
| Psoriasis | ![Life 13 00924 i001]() M.biota; M.sympodialis; Kocuria, Lactobacillus and Streptococcus with Saccharomyces, M.biota; M.sympodialis; Kocuria, Lactobacillus and Streptococcus with Saccharomyces, | [61,62] |
| Oral | Oropharyngeal candidosis | ![Life 13 00924 i001]() C.albicans C.albicans | [63] |
| Dental cavity | ![Life 13 00924 i001]() C.albicans, C. dubliniensis, Debaromyces spp., Rhodotorula spp., or malassezia spp. C.albicans, C. dubliniensis, Debaromyces spp., Rhodotorula spp., or malassezia spp. | [64,65,66] |
| Periodontitis | ![Life 13 00924 i001]() C. albicans C. albicans
![Life 13 00924 i002]() S. cerevisiae S. cerevisiae | [67,68] |
| Urogenital | Vulvovaginal candidosis (VVC) | ![Life 13 00924 i001]() C. albicans C. albicans | [8] |
| Candiduria | ![Life 13 00924 i001]() C. albicans, C. glabrata and C. tropicalis C. albicans, C. glabrata and C. tropicalis | |
3.2.1. Gut Mycobiome
The most studied fungal phyla present in the human intestine have been
Ascomycota as the most predominant phylum in the intestine, followed by
Zygomycota and
Basidiomycota [
69,
70,
71]. A newborn’s gut microbiome is highly dependent on its diet. Breastfed babies have higher levels of bacteria belonging to the Actinobacteria class [
72,
73] and the genera belonging to
Lactibacillus and
bifidobacterium [
72,
73,
74,
75]. High amounts of oligosaccharides and various fatty acids that make up breast milk positively influence the intestinal microbiome and their metabolites may help to act against hypersensitivity (allergy) and asthma counter reactions [
76].
Saccharomyces is a non-pathogenic yeast, thermotolerant and resistant to the action of gastric, enteric and pancreatic juices.
Saccharomyces has been used commercially in the production of probiotic foods. In recent decades,
S. cerevisiae and
S. boulardii have shown great promise as probiotic treatments [
77]. Several studies have shown
S. cerevisiae and
S. boulardii to be associated with an increase in the proportion of
Bacteroidetes in the gut microbiota composition and a decreased relative abundance of
Firmicutes and
Proteobacteria. Furthermore, this yeast can prevent inflammation by promoting pro-inflammatory immune function and increasing the production of short-chain fatty acids [
77,
78,
79,
80].
Other studies have also reported that
Malassezia,
Candida and
Saccharomyces constitute the intestinal microbiota, with
S. cerevisiae,
M. restricted and
C. albicans identified in 96.8%, 88.3% and 60.8% respectively [
81]. Similar to other microbial communities, fungi also have a high capacity to produce metabolites that can be applied for medicinal or therapeutic purposes. In addition to this functionality, they can also influence host homeostasis, causing biological effects on them as part of fungus–host interactions [
82].
Inflammatory Bowel Disease
Changes in the gut microbiome are associated with increased disease and dysbiosis of fungal communities which may contribute to susceptibility or increased disease severity. It is important to note that these changes, when caused by external influences, will facilitate invasion and that gastrointestinal (GI) infections affect the composition of the microbiota.
In CD (Crohn’s disease), there is an increase in the fecal fungal burden as well as an increase in the
Basidiomycota—
Ascomycota ratio. Regarding the opportunistic yeasts that cause this disease, an increase in the abundance of
C. albicans,
C. tropicalis,
C. glabrata,
Aspergillus clavatus,
Cryptococcus neoformans,
Cyberlindnera jadinii,
Clavispora lusitaniae,
Debaryomyces hansenii and
Kluyveromyces marxianus and a decrease in
Saccharomyces cerevisiae have been found [
48,
49,
50,
51].
Irritable Bowel Syndromes (IBS)
Although not considered a critical illness, it affects about 10–15% of individuals, reducing their quality of life [
30]. Studies have found an increase in the prevalence of
Candida spp. (notably
C. albicans),
Malassezia spp.,
Cladosporium spp. and
Saccharomyces cerevisiae. In contrast, yeasts such as
Mycosphaerella spp.,
Aspergillus spp.,
Sporidiobolus spp. and
Pandora spp. suffer a decrease in their abundance [
52,
53,
83,
84].
Cancer
The intestinal microbiome and its changes have been associated with the pathogenesis of diseases such as colorectal adenoma, which induces colorectal cancer (CRC), esophageal squamous cell carcinoma (ESCC), gastric cancer, hepatocellular carcinoma (HCC), cholangiocarcinoma and pancreatic ductal adenocarcinoma (PDAC) [
85].
Patients with these pathologies and with polyps have been observed with fungal dysbiosis, having a very high
Ascomycota/Basidiomycota ratio leading to the expansion of opportunistic fungi [
55].
Species such as
Trichosporon and
Malassezia were considered two of the populations capable of facilitating the progression and growth of colorectal cancer [
54]. Other studies have found that although no single type has been identified as the single leading cause, there is evidence to show an association between
Fusobacterium species (
F. mortiferum,
F. nucleatum, and
F. necrophorum) with colorectal cancer [
37]. Lev et al. also demonstrated that people with this pathology contain higher percentages of
Klebsiella,
E. coli,
Streptococcus and
Enterococcus as opposed to
Rothia which is in low amounts [
86].
Obesity
There are many causes that can lead to obesity, such as genetic, lifestyle and environmental factors. However, in addition to these causes, the intestinal microbiota plays a key role in the presence and development of obesity. The microbiota of overweight individuals has a greater capacity for fermentative processes and for capturing energy from the diet. It also has a high proportion of
Firmicutes to
bacteroides/
Prevotella leading to an increase in the microbiota gene involved in polysaccharide degradation and an increase in SCFAs [
87].
Candida,
Nakaseomyces,
Penicillium,
Porphyromonas,
Campylobacter,
Bacteroides,
Staphylococcus,
Parabacteroides,
Dialister and
Ruminococcus have been the most identified genera in overweight individuals while Mucor racemosus and M. fuscus have been found the most in non-obese patients [
55,
56]. It is now obvious that obesity is somehow associated with gut dysbiosis, low-grade inflammation and a host of metabolic disorders.
Diabetes
Autoimmune diseases, unlike IBD, are directly linked to an abnormal development of the intestinal microbiota throughout life. A study on the quantitative changes in
Candida species in patients with real-time PCR (qPCR) DM1 and DM2 was carried out, where
C. albicans was the most common strain found in the stool of these patients [
55]. However, no significant changes were observed between DM1 and DM2 patients in terms of
C. albicans colonization [
55].
Regarding genetics, nutrition and lifestyle choices, these also influence the prevalence of DM1, particularly if we are to assess the incidence rate in various countries around the world. Other studies have reported, in addition to an increase in
C. albicans, that in type 1 diabetes there is also an increase in the genus
Saccharomyces [
57]. In type 2 diabetes, in addition to an increase in
C. albicans, there an increase in
Cladosporium spp.,
Meyerozyma spp.,
Mortierella spp. and
Aspergillus spp. has also been found [
58].
In addition, it has been speculated that the high prevalence of
Saccharomyces may be due to the consumption of foods containing yeast (beer and bread), while the high percentage of
Candida is related to the consumption of carbohydrates [
46].
3.2.2. Oral Mycobiome
Despite the abundance of the mycobiome, the candida genus remains one of the few that unquestionably contributes to the emergence of the most common infections in the oral mucosa [
6]. However, although fungi that are present in a smaller percentage do not have a significant influence on metabolic activities, they can play a modulator role in immune responses or an opportunistic pathogenic role under surveillance conditions, harming the host [
6]. In some rare cases, involving severe immunosuppression,
Cryptococcus spp. and
Aspergillus spp. were described as causing lesions in the oral mucosa [
58].
Oropharyngeal Candidosis (OPC)
OPC can be classified into three main conditions: acute, chronic and chronic mucocutaneous candidosis [
63]. Some of the risk factors include nutritional deficiencies, local dysbiosis, salivary hypofunctions, smoking, use of dentures and a dysfunction in T-cell immunity [
63].
C. albicans is the fungus responsible for causing this disease, and life-threatening systemic infections can develop when this fungus enters the bloodstream [
63].
Dental Cavity
The role of the oral mycobiome in caries has been a recent focus. A study by Baraniya in 2020 found that advanced caries were associated with an abnormal increase in the prevalence and abundance of
C. albicans in adults and of
C. dubliniensis in children [
64,
65].
Furthermore, one study reported a regressive trend in mycobiome diversity as caries severity increased [
66]. Interestingly, in another study,
C. albicans was associated with severe disease, while
C. dubliniensis was shown to have a gradual and steady increase as the disease set in and grew [
66]. On the other hand, in caries-free children, one of the most common fungi was found to be
Malassezia globosa [
64].
In summary, it is thought that
C. albicans will be involved in more advanced lesions while
C. dubliniensis plays a pivotal role earlier in the disease process. Some investigations suggest that some fungi are found in caries-free children, namely
Debaromyces spp.,
Rhodotorula spp. or
malassezia spp. [
64,
66].
Periodontitis
Although fungal communities have already been detected in the subgingival plaque, their role is still unclear. Research has reported an increase in yeast detection, namely
Candida spp., in subjects with periodontitis [
67]. However, this study was carried out through cultures, because when tests were carried out at the molecular level, this increase was not verified [
68]. This study saw that this pathogenesis was associated with a decrease in microbiome diversity and changes in the relative richness of two genera (decrease in
S. cerevisiae and an increase in
Filobasidiales species) when compared to individuals with or without mild disease [
68].
3.2.3. Skin Mycobiome
Most studies on the microbiome focus on understanding the bacterial composition, but the microorganisms present on the skin are not limited to bacteria, but also include viruses, fungi and arthropods. Notably, the skin being the largest organ in the human body, it serves as a defensive obstruction against possible injury and microbial attack [
36].
Pityriasis Versicolor
Pityriasis versicolor is a chronic skin infection characterized by discrete or confluent, scaly, dark or depigmented patches, mainly on the upper torso, but which can extend to the neck, abdomen and other locations. The
Malassezia species identified mainly in pityriasis versicolor lesions are
M. globosa and
M. sympodialis and
M. furfur. In addition to the findings of the action of this yeast in pityriasis versicolor, there are two facts that further emphasize its role in this disease: (i) a positive culture is found more in samples cultured from skin lesions than from healthy skin [
59], and (ii) in its hyphal form, it is usually found in samples obtained from pityriasis versicolor lesions, regardless of the species of malassezia present [
88].
Seborrheic Dermatitis
The relationship between malassezia colonization and seborrheic dermatitis was first described by Louis-Charles Malassez in 1874. Seborrheic dermatitis is an inflammatory dermatosis with a predilection for anatomical areas with a high concentration of sebaceous glands, such as the middle third of the face, chest, back and scalp. The prevalence of seborrheic dermatitis also increases with age (2.0% in <35 years; 3.6% in 35–64 years; 4.4% ≥65 years) and there is an association with other fungal diseases such as tinea pedis, onychomycosis and pityriasis versicolor. The age dependence of seborrheic dermatitis is probably responsible for the increased prevalence (14.3%) reported in the Rotterdam study [
89].
Fungi such as
Malassezia is found in the sebaceous gland where lipids are the main source of energy. As this yeast is usually commensal, the mechanism that triggers this disease has been increasingly investigated [
60]. Its DNA has been detected in about 90% of skin lesions; the skin lesions of atopic dermatitis and colonization increase with the severity of the disease [
90].
Psoriasis
Psoriasis is an inflammatory skin disease that affects approximately 2% of the world’s population. Little is known about the role of the skin microbiome in psoriasis. However, some studies have already been carried out on this disease. In one study, an increase in
Brevibacterium,
Kocuria palustris and
Gordonia was found to be associated with back and elbow injuries [
61]. In that same study, it was also found that there was a high abundance of the
Malassezia fungus. Strains such as
M. biota were detected on the coast, while
M. sympodialis dominated elbow mycosis [
61].
In psoriatic elbow skin, there was found to be a significant correlation between the occurrence of
Kocuria,
Lactobacillus and
Streptococcus with
Saccharomyces, which was not observed in healthy skin [
62].
3.2.4. Urogenital Mycobiome
The composition of the female microbiota varies according to age, hormone production, menstrual cycle, drug use and sexual activity. Evidence has shown that the composition of the microbiota is important for reproductive and genital tract health.
Vulvovaginal Candidosis (VVC)
Mucosal infections, characterized by fungal colonization associated with an inflammatory host response, are extremely common and cause a negative impact on the quality of life of patients.
VVC is a multifactorial condition with an abnormal immune system response. Undeniably, colonization is carried out by both bacteria and fungi, with communication between them through physical interactions, production of metabolites and chemical interactions, among others. Most in vivo studies have revealed an inhibitory capacity on the part of
Lactobacillus on the growth, dimorphic transition, virulence factors and biofilm formation of
C. albicans [
8]. For example, about 75% of women of childbearing age have at least one episode of VVC and up to 9% of them suffer from recurrent VVC (more than four episodes per year) [
63,
91,
92]. Some of the risk factors associated with this pathogenesis include a very high level of estrogen, the use of oral contraceptives and uncontrolled diabetes [
63].
Candiduria
Most clinicians accept that Candiduria is defined as colonization or contamination, but it may be the only sign of invasive candidosis. The consensus is that Candiduria is quite common in hospitalized patients [
93,
94]. The spread of the disease usually happens through contamination. It is usually diagnosed in elderly patients, with
Candida being the most frequently isolated microorganism in nosocomial urinary tract infections (UTIs). Overall,
C. albicans accounts for 50 to 70% of cases, followed by
C. glabrata and
C. tropicalis, which is the third most common species. There are studies that claim that the use of antibiotics is associated with an increase in their incidence; however, these estimated values can be disregarded, as the standard urine culture is not very sensitive [
95].
From all the studies carried out in patients with Candiduria, a set of risk factors that are associated with an increase in its incidence was gathered. In diabetes mellitus, frequent or prolonged use of antimicrobials, broad-spectrum antibiotics (which suppress the gastrointestinal and genital flora), can lead to ICU stay and use of indwelling catheters, among others.
3.3. Interconnection between Different Niches
Studies indicate that alterations in intestinal microbial diversity (dysbiosis) can lead to an increase in host susceptibility and an interruption of mucosal immune tolerance, which will influence skin health in the future [
96]. Other studies have associated a bidirectional interaction between gastrointestinal health and skin homeostasis through the metabolic activity and immune impact of the gut microbiome [
97,
98]. For example, commensal intestinal microorganisms can promote skin allostasis by controlling T-cell differentiation [
97].
Although a healthy gut microbiota is critical to host health, overgrowth of the host and changes in diversity can also result in disease processes, for example, patients with Crohn’s disease are also found primarily with psoriasis as a comorbidity [
99,
100].
Studies carried out deepening the knowledge of the possible relationship between the intestinal and oral microbiome reported that patients with intestinal diseases exhibited a considerable abnormal increase in oral microorganisms in the lumen and in the tissues of the intestinal mucosa [
101,
102]. Thus, it is plausible to believe that the cavity serves as a reservoir; however, we have still not obtained comprehensive information on which microorganisms act as pathogens.
Some researchers have already found evidence that suggests the existence of a gut–vagina axis. There are already some data that the intestinal and vaginal microbiota can be completely linked, for example, in the onset and progression of endometriosis [
103]. Yet, another study about this connection between endometriosis and the intestine showed that after 42 days of persistence of endometrial injury, a distinct intestinal microbiota develops [
104], that is, not only does the intestinal microbiota change the vaginal one, but vice versa.
These hypotheses open many doors to new preventive, diagnostic and therapeutic possibilities, and are therefore an emerging area for investigation.
4. Conclusions
With this review, we intended to synthesize the existing literature about the microbiota and mycobiota. In addition, we highlighted the communities that live in the different niches of the human body, namely the oral, vaginal, intestinal and skin cavities, as well as the diseases that occur when there are changes in the composition of the microbiota.
It should be noted that the importance and significant role that fungal communities play in human health are becoming increasingly more evident. However, more studies are needed to understand the effect of geography on the human mycobiome. Other aspects such as dietary habits or age should also be studied in detail to provide a deeper knowledge on the dynamics of the human mycobiome.
Furthermore, future research involving the study of the mycobiome should have the evolution and improvement in the health of humans, animals and also plants as an objective, as well as the improvement in the ecosystem as a whole. The increase in the amount of data available on mycobiomes allows for a greater knowledge to be attained on their use in increasing productivity and possible therapies. One of the major problems for health is the acquisition of resistance of microorganisms to antifungal drugs.
To contribute to possible alternatives for solving this problem, systems based on the mycobiome for monitoring and sanitization are being tested and considered good solutions, especially in the hospital environment. When the imbalance caused at a more specific level is known, it is possible to know in which human niches there are greater chances of proliferation of these pathogens. In addition, sanitizing products containing probiotics are already being developed to modulate these mycobiomes, making it difficult for pathogens to appear in these environments.
Therefore, for future work, it is crucial to start including these yeasts in studies so that we begin to have a more realistic perception of the action of fungal communities in the homeostasis of the microbiota and, consequently, of human health, and to further deepen the knowledge of clinical mycology.
Author Contributions
Conceptualization, J.M.-d.-O. and J.R.; methodology, C.F.C. and C.G.; investigation, C.F.C.; resources, A.P.-d.-O.; writing—original draft preparation, C.F.C.; writing—review and editing; J.R., J.M.-d.-O. and A.P.-d.-O.; supervision, C.G. and J.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financed by Fundação para a Ciência e a Tecnologia (FCT; UIDP/00709/2020 and UIDB/00709/2020) through funding attributed to Centro de Investigação em Ciências da Saúde (CICS-UBI). J.R. was financed by fellowship SFRH/BPD/115145/2016 funded by FCT.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jethwani, P.; Grover, K. Gut Microbiota in Health and Diseases—A Review. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 1586–1599. [Google Scholar] [CrossRef]
- Ellis, S.R.; Nguyen, M.; Vaughn, A.R.; Notay, M.; Burney, W.A.; Sandhu, S.; Sivamani, R.K. The Skin and Gut Microbiome and Its Role in Common Dermatologic Conditions. Microorganisms 2019, 7, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillman, E.T.; Lu, H.; Yao, T.; Nakatsu, C.H. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ. 2017, 32, 300–313. [Google Scholar] [CrossRef] [Green Version]
- Diaz, P.; Dongari-Bagtzoglou, A. Critically Appraising the Significance of the Oral Mycobiome. J. Dent. Res. 2021, 100, 133–140. [Google Scholar] [CrossRef]
- Ward, T.L.; Dominguez-Bello, M.G.; Heisel, T.; Al-Ghalith, G.; Knights, D.; Gale, C.A. Development of the Human Mycobiome over the First Month of Life and across Body Sites. Msystems 2018, 3, e00140-17. Available online: http://msystems.asm.org (accessed on 1 February 2023). [CrossRef] [PubMed] [Green Version]
- Paul, A.A.; Hoffman, K.L.; Hagan, J.L.; Sampath, V.; Petrosino, J.F.; Pammi, M. Fungal cutaneous microbiome and host determinants in preterm and term neonates. Pediatr. Res. 2020, 88, 225–233. [Google Scholar] [CrossRef]
- Lehtoranta, L.; Hibberd, A.A.; Yeung, N.; Laitila, A.; Maukonen, J.; Ouwehand, A.C. Characterization of vaginal fungal communities in healthy women and women with bacterial vaginosis (BV); a pilot study. Microb. Pathog. 2021, 161, 105055. [Google Scholar] [CrossRef]
- Spatz, M.; Richard, M.L. Overview of the potential role of Malassezia in gut health and disease. Front. Cell. Infect. Micro-biol. 2020, 10, 201. [Google Scholar] [CrossRef]
- Ward, T.L.; Knights, D.; Gale, C.A. Infant fungal communities: Current knowledge and research opportunities. BMC Med. 2017, 15, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Rautava, S.; Aakko, J.; Isolauri, E.; Salminen, S. Human gut colonisation may be initiated in utero by distinct mi-crobial communities in the placenta and amniotic fluid. Sci. Rep. 2016, 6, 23129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schei, K.; Avershina, E.; Øien, T.; Rudi, K.; Follestad, T.; Salamati, S.; Ødegård, R.A. Early gut mycobiota and mother-offspring transfer. Microbiome 2017, 5, 107. [Google Scholar] [CrossRef] [Green Version]
- Franco-Paredes, C.; Womack, T.; Bohlmeyer, T.; Sellers, B.; Hays, A.; Patel, K.; Lizarazo, J.; Lockhart, S.R.; Siddiqui, W.; Marr, K.A. Management of Cryptococcus gattii meningoencephalitis. Lancet Infect. Dis. 2015, 15, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.D.A.G.D.; Leite-Andrade, M.C.; Brandão, I.D.S.; Alves, A.I.D.S.; Buonafina, M.D.S.; Nunes, M.; de Araújo-Neto, L.N.; de Freitas, M.A.; Brayner, F.A.; Alves, L.C.; et al. Anti-biofilm effect by the combined action of fluconazole and acetylsalicylic acid against species of Candida parapsilosis complex. Infect. Genet. Evol. 2020, 84, 104378. [Google Scholar] [CrossRef]
- Lamoth, F.; Lockhart, S.R.; Berkow, E.L.; Calandra, T. Changes in the epidemiological landscape of invasive candidiasis. J. Antimicrob. Chemother. 2018, 73, i4–i13. [Google Scholar] [CrossRef] [Green Version]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive Candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef]
- Opulente, D.A.; Langdon, Q.K.; Buh, K.V.; Haase, M.A.B.; Sylvester, K.; Moriarty, R.V.; Jarzyna, M.; Considine, S.L.; Schneider, R.M.; Hittinger, C.T. Pathogenic budding yeasts isolated outside of clinical settings. FEMS Yeast Res. 2019, 19, foz032. [Google Scholar] [CrossRef] [PubMed]
- Zhai, B.; Ola, M.; Rolling, T.; Tosini, N.L.; Joshowitz, S.; Littmann, E.R.; Amoretti, L.A.; Fontana, E.; Wright, R.J.; Miranda, E.; et al. High-resolution mycobiota analysis reveals dynamic intestinal translocation preceding invasive candidiasis. Nat. Med. 2020, 26, 59–64. [Google Scholar] [CrossRef]
- McCarty, T.P.; Pappas, P.G. Invasive Candidiasis. Infect. Dis. Clin. North Am. 2016, 30, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Drell, T.; Lillsaar, T.; Tummeleht, L.; Simm, J.; Aaspõllu, A.; Väin, E.; Saarma, I.; Salumets, A.; Donders, G.; Metsis, M. Characterization of the Vaginal Micro- and Mycobiome in Asymptomatic Reproductive-Age Estonian Women. PLoS ONE 2013, 8, e54379. [Google Scholar] [CrossRef]
- Ceccarani, C.; Foschi, C.; Parolin, C.; D’Antuono, A.; Gaspari, V.; Consolandi, C.; Laghi, L.; Camboni, T.; Vitali, B.; Severgnini, M.; et al. Diversity of vaginal microbiome and metabolome during genital infections. Sci. Rep. 2019, 9, 14095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.A.; Noverr, M. Fungal interactions with the human host: Exploring the spectrum of symbiosis. Curr. Opin. Microbiol. 2017, 40, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Bojang, E.; Ghuman, H.; Kumwenda, P.; Hall, R. Immune Sensing of Candida albicans. J. Fungi 2021, 7, 119. [Google Scholar] [CrossRef]
- Yu, C.-H.; Sephton-Clark, P.; Tenor, J.L.; Toffaletti, D.L.; Giamberardino, C.; Haverkamp, M.; Cuomo, C.A.; Perfect, J.R. Gene Expression of Diverse Cryptococcus Isolates during Infection of the Human Central Nervous System. Mbio 2021, 12, e02313-21. [Google Scholar] [CrossRef] [PubMed]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Jamil, K.; Polyak, M.J.; Feehan, D.D.; Surmanowicz, P.; Stack, D.; Li, S.S.; Ogbomo, H.; Olszewski, M.; Ganguly, A.; Mody, C.H. Phagosomal F-Actin Retention by Cryptococcus gattii Induces Dendritic Cell Immunoparalysis. Mbio 2020, 11, e01821-20. [Google Scholar] [CrossRef]
- Rhimi, W.; Theelen, B.; Boekhout, T.; Otranto, D.; Cafarchia, C. Malassezia spp. Yeasts of Emerging Concern in Fungemia. Front. Cell. Infect. Microbiol. 2020, 10, 370. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.L. Malassezia: The Forbidden Kingdom Opens. Cell Host Microbe 2019, 25, 345–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suhr, M.J.; Banjara, N.; Hallen-Adams, H.E. Sequence-based methods for detecting and evaluating the human gut mycobiome. Lett. Appl. Microbiol. 2016, 62, 209–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallen-Adams, H.E.; Suhr, M.J. Fungi in the healthy human gastrointestinal tract. Virulence 2017, 8, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Boix-Amorós, A.; Martinez-Costa, C.; Querol, A.; Collado, M.C.; Mira, A. Multiple Approaches Detect the Presence of Fungi in Human Breastmilk Samples from Healthy Mothers. Sci. Rep. 2017, 7, 13016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, R.; Fernández-Fernández, A.M.; Pisa, D.; Carrasco, L. Multiple sclerosis and mixed microbial infections. Direct identification of fungi and bacteria in nervous tissue. Neurobiol. Dis. 2018, 117, 42–61. [Google Scholar] [CrossRef]
- Boix-Amorós, A.; Puente-Sánchez, F.; Du Toit, E.; Linderborg, K.M.; Zhang, Y.; Yang, B.; Salminen, S.; Isolauri, E.; Tamames, J.; Mira, A.; et al. Mycobiome Profiles in Breast Milk from Healthy Women Depend on Mode of Delivery, Geographic Location, and Interaction with Bacteria. Appl. Environ. Microbiol. 2019, 85, e02994-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, C.W.; Scheynius, A.; Heitman, J. Malassezia fungi are specialized to live on skin and associated with dandruff, eczema, and other skin diseases. PLoS Pathog. 2012, 8, e1002701. [Google Scholar] [CrossRef]
- Hsu, D.K.; Fung, M.A.; Chen, H.-L. Role of skin and gut microbiota in the pathogenesis of psoriasis, an inflammatory skin disease. Med. Microecol. 2020, 4, 100016. [Google Scholar] [CrossRef]
- Hof, H. Pilze im Darm—Das Mykobiom des Darms. Z. Gastroenterol. 2017, 55, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.M.; de Paula, T.O.; Sarmiento, M.R.A.; de Oliveira, M.G.; Pereira, M.L.M.; Toledo, I.V.; Nascimento, T.C.; Ferreira-Machado, A.B.; Silva, V.L.; Diniz, C.G. Fungal Diversity of Human Gut Microbiota Among Eutrophic, Overweight, and Obese Individuals Based on Aerobic Culture-Dependent Approach. Curr. Microbiol. 2018, 75, 726–735. [Google Scholar] [CrossRef]
- Ioannou, P.; Vamvoukaki, R.; Samonis, G. Rhodotorula species infections in humans: A systematic review. Mycoses 2019, 62, 90–100. [Google Scholar] [CrossRef]
- Wirth, F.; Goldani, L.Z. Epidemiology of Rhodotorula: An Emerging Pathogen. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 465717. [Google Scholar] [CrossRef] [Green Version]
- Meletiadis, J.; Roilides, E. Rare Invasive Fungal Infections: Epidemiology, Diagnosis and Management. Curr. Fungal Infect. Rep. 2013, 7, 351–360. [Google Scholar] [CrossRef]
- Falces-Romero, I.; Cendejas-Bueno, E.; Romero-Gómez, M.P.; García-Rodríguez, J. Isolation of Rhodotorula mucilaginosa from blood cultures in a tertiary care hospital. Mycoses 2018, 61, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Setiawan, W.; Wiyono, S.; Tondok, E.T.; Kanti, A.; Sudiana, I.M. In Vitro Study of Action Mode of Rhodotorula minuta Dmg 16 BEP as Biocontrol Agents on Alternaria solani. J. Perlindungan Tanam. Indones. 2020, 24, 28–33. [Google Scholar] [CrossRef]
- Chaud, L.C.S.; Lario, L.D.; Bonugli-Santos, R.C.; Sette, L.D.; Junior, A.P.; Felipe, M.d.G.d.A. Improvement in extracellular protease production by the marine antarctic yeast Rhodotorula mucilaginosa L7. New Biotechnol. 2016, 33, 807–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. Fungal Infections—Protect Your Health. Fungal Diseases. 2019. Available online: https://www.cdc.gov/fungal/features/fungal-infections.html (accessed on 24 January 2023).
- Chin, V.K.; Yong, V.C.; Chong, P.P.; Nordin, S.A.; Basir, R.; Abdullah, M. Mycobiome in the Gut: A Multiperspective Review. Mediat. Inflamm. 2020, 2020, 9560684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Kim, S.-H. A genome Tree of Life for the Fungi kingdom. Proc. Natl. Acad. Sci. USA 2017, 114, 9391–9396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, E.R.; Zisman, T.L.; Suskind, D.L. The microbiota in inflammatory bowel disease: Current and therapeutic insights. J. Inflamm. Res. 2017, ume 10, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal microbiota dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, A.; Stewart, C.J.; A Kennedy, N.; Lodge, J.K.; Tremelling, M.; Probert, C.S.; Parkes, M.; Mansfield, J.C.; Smith, D.L.; Hold, G.L.; et al. The Impact of NOD2 Genetic Variants on the Gut Mycobiota in Crohn’s Disease Patients in Remission and in Individuals Without Gastrointestinal Inflammation. J. Crohn’s Colitis 2021, 15, 800–812. [Google Scholar] [CrossRef]
- Hoarau, G.; Mukherjee, P.K.; Gower, C.; Hager, C.; Chandra, J.; Retuerto, M.A.; Neut, C.; Vermeire, S.; Clemente, J.; Colombel, J.F.; et al. Bacteriome and Mycobiome Interactions Underscore Microbial Dysbiosis in Familial Crohn’s Disease. Mbio 2016, 7, e01250-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botschuijver, S.; Roeselers, G.; Levin, E.; Jonkers, D.M.; Welting, O.; Heinsbroek, S.E.; de Weerd, H.H.; Boekhout, T.; Fornai, M.; Masclee, A.A.; et al. Intestinal Fungal Dysbiosis Is Associated With Visceral Hypersensitivity in Patients With Irritable Bowel Syndrome and Rats. Gastroenterology 2017, 153, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; O’Herlihy, E.; Shanahan, F.; O’Toole, P.W.; Jeffery, I.B. The fecal mycobiome in patients with Irritable Bowel Syndrome. Sci. Rep. 2021, 11, 124. [Google Scholar] [CrossRef] [PubMed]
- Lv, G.; Cheng, N.; Wang, H. The Gut Microbiota, Tumorigenesis, and Liver Diseases. Engineering 2017, 3, 110–114. [Google Scholar] [CrossRef]
- Richard, M.L.; Liguori, G.; Lamas, B.; Brandi, G.; DA Costa, G.; Hoffmann, T.W.; Di Simone, M.P.; Calabrese, C.; Poggioli, G.; Langella, P.; et al. Mucosa-associated microbiota dysbiosis in colitis associated cancer. Gut Microbes 2018, 9, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Zorena, K.; Kowalewska, B.; Szmigiero-Kawko, M.; Wąż, P.; Myśliwiec, M. Higher diversity in fungal species discriminates children with type 1 diabetes mellitus from healthy control. Patient Preference Adherence 2016, 10, 591–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayasudha, R.; Das, T.; Kalyana Chakravarthy, S.; Sai Prashanthi, G.; Bhargava, A.; Tyagi, M.; Rani, P.K.; Pappuru, R.R.; Shivaji, S. Gut mycobiomes are altered in people with type 2 Diabetes Mellitus and Diabetic Retinopathy. PLoS ONE 2020, 15, e0243077. [Google Scholar] [CrossRef]
- Iatta, R.; Napoli, C.; Borghi, E.; Montagna, M.T. Rare mycoses of the oral cavity: A literature epidemiologic review. Oral Surgery, Oral Med. Oral Pathol. Oral Radiol. Endodontology 2009, 108, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Prohic, A.; Ozegovic, L. Malassezia species isolated from lesional and non-lesional skin in patients with pityriasis versicolor. Mycoses 2007, 50, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Wakelin, S. Seborrheic Dermatitis. In Rook’s Textbook of Dermatology; Griffiths, C., Barker, J., Bleiker, T., Chalmers, R., Creamer, D., Eds.; John Wiley & Sons: Chichester, UK, 2016; p. 40.2. [Google Scholar]
- Chang, H.-W.; Yan, D.; Singh, R.; Liu, J.; Lu, X.; Ucmak, D.; Lee, K.; Afifi, L.; Fadrosh, D.; Leech, J.M.; et al. Alteration of the cutaneous microbiome in psoriasis and potential role in Th17 polarization. Microbiome 2018, 6, 154. [Google Scholar] [CrossRef]
- Stehlikova, Z.; Kostovcik, M.; Kostovcikova, K.; Kverka, M.; Juzlova, K.; Rob, F.; Hercogova, J.; Bohac, P.; Pinto, Y.; Uzan, A.; et al. Dysbiosis of Skin Microbiota in Psoriatic Patients: Co-occurrence of Fungal and Bacterial Communities. Front. Microbiol. 2019, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- Rosati, D.; Bruno, M.; Jaeger, M.; Ten Oever, J.; Netea, M.G. Recurrent Vulvovaginal Candidiasis: An Immunological Perspective. Microorganisms 2020, 8, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baraniya, D.; Chen, T.; Nahar, A.; Alakwaa, F.; Hill, J.; Tellez, M.; Ismail, A.; Puri, S.; Al-Hebshi, N.N. Supragingival mycobiome and inter-kingdom interactions in dental caries. J. Oral Microbiol. 2020, 12, 1729305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jesus, V.C.; Shikder, R.; Oryniak, D.; Mann, K.; Alamri, A.; Mittermuller, B.; Duan, K.; Hu, P.; Schroth, R.J.; Chelikani, P. Sex-based diverse plaque microbiota in children with severe caries. J. Dent. Res. 2020, 99, 703–712. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, L.M.; Santos, R.; Springer, G.; Burne, R.A.; Nascimento, M.M.; Richards, V.P. Site-Specific Profiling of the Dental Mycobiome Reveals Strong Taxonomic Shifts during Progression of Early-Childhood Caries. Appl. Environ. Microbiol. 2020, 86, e02825-19. [Google Scholar] [CrossRef] [PubMed]
- Canabarro, A.; Valle, C.; Farias, M.R.; Santos, F.B.; Lazera, M.; Wanke, B. Association of subgingival colonization of Candida albicans and other yeasts with severity of chronic periodontitis. J. Periodontal Res. 2013, 48, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Annavajhala, M.K.; Khan, S.D.; Sullivan, S.B.; Shah, J.; Pass, L.; Kister, K.; Kunen, H.; Chiang, V.; Monnot, G.C.; Ricupero, C.L.; et al. Oral and Gut Microbial Diversity and Immune Regulation in Patients with HIV on Antiretroviral Therapy. Msphere 2020, 5, e00798-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lücking, R.; et al. The high-level phylogenetic classification of fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Dollive, S.; Grunberg, S.; Chen, J.; Li, H.; Wu, G.D.; Lewis, J.D.; Bushman, F.D. Archaea and Fungi of the Human Gut Microbiome: Correlations with Diet and Bacterial Residents. PLoS ONE 2013, 8, e66019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sam, Q.H.; Chang, M.W.; Chai, L.Y.A. The Fungal Mycobiome and Its Interaction with Gut Bacteria in the Host. Int. J. Mol. Sci. 2017, 18, 330. [Google Scholar] [CrossRef] [Green Version]
- Kordy, K.; Gaufin, T.; Mwangi, M.; Li, F.; Cerini, C.; Lee, D.J.; Adisetiyo, H.; Woodward, C.; Pannaraj, P.S.; Tobin, N.H.; et al. Contributions to human breast milk microbiome and enteromammary transfer of Bifidobacterium breve. PLoS ONE 2020, 15, e0219633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamash, M.; Maurice, C.F. Phages in the infant gut: A framework for virome development during early life. ISME J. 2022, 16, 323–330. [Google Scholar] [CrossRef]
- Duranti, S.; Lugli, G.A.; Mancabelli, L.; Armanini, F.; Turroni, F.; James, K.; Ferretti, P.; Gorfer, V.; Ferrario, C.; Milani, C.; et al. Maternal inheritance of bifidobacterial communities and bifidophages in infants through vertical transmission. Microbiome 2017, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 2017, 66, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Thijs, C.; Vink, C.; Stelma, F.F.; Snijders, B.; Kummeling, I.; Van den Brandt, P.A.; Stobberingh, E.E. Factors Influencing the Composition of the Intestinal Microbiota in Early Infancy. Pediatrics 2006, 118, 511–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Zhao, X.-K.; Cheng, M.-L.; Yang, G.-Z.; Wang, B.; Liu, H.-J.; Hu, Y.-X.; Zhu, L.-L.; Zhang, S.; Xiao, Z.-W.; et al. Saccharomyces boulardii Administration Changes Gut Microbiota and Attenuates D-Galactosamine-Induced Liver Injury. Sci. Rep. 2017, 7, 1359. [Google Scholar] [CrossRef] [Green Version]
- Everard, A.; Matamoros, S.; Geurts, L.; Delzenne, N.M.; Cani, P.D. Saccharomyces boulardii Administration Changes Gut Microbiota and Reduces Hepatic Steatosis, Low-Grade Inflammation, and Fat Mass in Obese and Type 2 Diabetic db/db Mice. mBio 2014, 5, e01011-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zhu, L.; Xie, A.; Yuan, J. Oral Administration of Saccharomyces boulardii Ameliorates Carbon Tetrachloride-Induced Liver Fibrosis in Rats via Reducing Intestinal Permeability and Modulating Gut Microbial Composition. Inflammation 2014, 38, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Brun, P.; Scarpa, M.; Marchiori, C.; Sarasin, G.; Caputi, V.; Porzionato, A.; Giron, M.C.; Palù, G.; Castagliuolo, I. Saccharomyces boulardii CNCM I-745 supplementation reduces gastrointestinal dysfunction in an animal model of IBS. PLoS ONE 2017, 12, e0181863. [Google Scholar] [CrossRef] [Green Version]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The intestinal mycobioma of the healthy cohort of the Human Microbiome Project. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef]
- Mogilnicka, I.; Ufnal, M. Gut mycobiota and fungal metabolites in human homeostasis. Curr. Drug Targets 2018, 20, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Hong, G.; Li, Y.; Yang, M.; Li, G.; Qian, W.; Xiong, H.; Bai, T.; Song, J.; Zhnag, L.; Hou, X.; et al. Gut fungal dysbiosis and altered bacterial-fungal interaction in patients with diar-rhea-predominant irritable bowel syndrome: An explorative study. Neurogastroenterol. Motil. 2020, 32, e13891. [Google Scholar] [CrossRef]
- Sciavilla, P.; Strati, F.; Di Paola, M.; Modesto, M.; Vitali, F.; Cavalieri, D.; Prati, G.M.; Di Vito, M.; Aragona, G.; De Filippo, C.; et al. Gut microbiota profiles and characterization of cultivable fungal isolates in IBS patients. Appl. Microbiol. Biotechnol. 2021, 105, 3277–3288. [Google Scholar] [CrossRef]
- Liu, Y.; Baba, Y.; Ishimoto, T.; Gu, X.; Zhang, J.; Nomoto, D.; Okadome, K.; Baba, H.; Qiu, P. Gut microbiome in gastrointestinal cancer: A friend or foe? Int. J. Biol. Sci. 2022, 18, 4101–4117. [Google Scholar] [CrossRef] [PubMed]
- Andoh, A.; Nishida, A.; Takahashi, K.; Inatomi, O.; Imaeda, H.; Bamba, S.; Kito, K.; Sugimoto, M.; Kobayashi, T. Comparison of the gut microbial community between obese and lean peoples using 16S gene sequencing in a Japanese population. J. Clin. Biochem. Nutr. 2016, 59, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Gaitanis, G.; Magiatis, P.; Hantschke, M.; Bassukas, I.D.; Velegraki, A. The Malassezia Genus in Skin and Systemic Diseases. Clin. Microbiol. Rev. 2012, 25, 106–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, M.; Pardo, L.; Franco, O.; Ginger, R.; Nijsten, T. Prevalence and determinants of seborrhoeic dermatitis in a middle-aged and elderly population: The Rotterdam Study. Br. J. Dermatol. 2018, 178, 148–153. [Google Scholar] [CrossRef] [PubMed]
- VSCT-HM, R. Atopisches Ekzem und Mikrobiom. Z. Fur Dermatol. Venerol. Und Verwandte Geb. 2019, 70, 407–415. [Google Scholar]
- Willems, H.M.E.; Ahmed, S.S.; Liu, J.; Xu, Z.; Peters, B.M. Vulvovaginal Candidiasis: A Current Understanding and Burning Questions. J. Fungi 2020, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Foxman, B.; Muraglia, R.; Dietz, J.-P.; Sobel, J.D.; Wagner, J. Prevalence of Recurrent Vulvovaginal Candidiasis in 5 European Countries and the United States: Results from an Internet Panel Survey. 2013. Available online: www.ipsos.com (accessed on 1 February 2023).
- Yano, J.; Sobel, J.D.; Nyirjesy, P.; Sobel, R.; Williams, V.L.; Yu, Q.; Noverr, M.C.; Fidel, P.L., Jr. Current patient perspectives of vulvovaginal candidiasis: Incidence, symptoms, management and post-treatment outcomes. BMC Women’s Health 2019, 19, 48. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Lerma, F.; Nolla-Salas, J.; León, C.; Palomar, M.; Jordá, R.; Carrasco, N.; Bobillo, F. Candiduria in critically ill patients admitted to intensive care medical units. Intensiv. Care Med. 2003, 29, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, C.A.; Vazquez, J.A.; Sobel, J.D.; Gallis, H.A.; McKinsey, D.S.; Karchmer, A.W.; Sugar, A.M.; Sharkey, P.K.; Wise, G.J.; Mangi, R.; et al. Prospective Multicenter Surveillance Study of Funguria in Hospitalized Patients. Clin. Infect. Dis. 2000, 30, 14–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberger, M.; Sweet, S.; Leibovici, L.; Pitlik, S.; Samra, Z. Correlation between candiduria and departmental antibiotic use. J. Hosp. Infect. 2003, 53, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Pascal, M.; Perez-Gordo, M.; Caballero, T.; Escribese, M.M.; Longo, M.N.L.; Luengo, O.; Manso, L.; Matheu, V.; Seoane, E.; Zamorano, M.; et al. Microbiome and Allergic Diseases. Front. Immunol. 2018, 9, 1584. [Google Scholar] [CrossRef]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The Gut Microbiome as a Major Regulator of the Gut-Skin Axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, C.A.; Monteleone, G.; McLaughlin, J.T.; Paus, R. The gut-skin axis in health and disease: A paradigm with therapeutic implications. Bioessays 2016, 38, 1167–1176. [Google Scholar] [CrossRef]
- Vaughn, A.R.; Notay, M.; Clark, A.K.; Sivamani, R.K. 52 Skin-Gut Axis: The Relationship between Intestinal Bacteria and Skin Health CASE REPORT 59 Pleomorphic Cutaneous Xanthomas Disclosing Homozygous Familial Hypercholesterolemia. World J. Derm. 2017, 6, 52–58. Available online: http://www.f6publishing.com (accessed on 28 February 2023). [CrossRef]
- Alzahrani, Y.A.; Alesa, D.I.; Alshamrani, H.M.; Alzahrani, Y.A.; Alamssi, D.N.; Alzahrani, N.S.; Almohammadi, M.E. The role of gut microbiome in the pathogenesis of psoriasis and the therapeutic effects of probiotics. J. Fam. Med. Prim. Care 2019, 8, 3496–3503. [Google Scholar] [CrossRef]
- Gevers, D.; Kugathasan, S.; Denson, L.A.; Vázquez-Baeza, Y.; Van Treuren, W.; Ren, B.; Schwager, E.; Knights, D.; Song, S.J.; Yassour, M.; et al. The Treatment-Naive Microbiome in New-Onset Crohn’s Disease. Cell Host Microbe 2014, 15, 382–392. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T. Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer. BIO Web Conf. 2021, 41, 01001. [Google Scholar] [CrossRef]
- Jiang, I.; Yong, P.; Allaire, C.; Bedaiwy, M. Intricate Connections between the Microbiota and Endometriosis. Int. J. Mol. Sci. 2021, 22, 5644. [Google Scholar] [CrossRef] [PubMed]
| Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).

 ,
, 
{kind=link}
{kind=link}
 C. albicans, C. tropicalis, C. glabrata, Aspergillus clavatus, C. neoformans, Cyberlindnera jadinii, Clavispora lusitaniae, Debaryomyces hansenii, Kluyveromyces marxianus
C. albicans, C. tropicalis, C. glabrata, Aspergillus clavatus, C. neoformans, Cyberlindnera jadinii, Clavispora lusitaniae, Debaryomyces hansenii, Kluyveromyces marxianus Saccharomyces cerevisiae
Saccharomyces cerevisiae
