
Araeoanasillus leptosomus gen. et sp. nov., (Hemiptera, Cercopoidea, Sinoalidae?), a New Froghopper from Mid-Cretaceous Burmese Amber with Evidence of Its Possible Host Plant

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
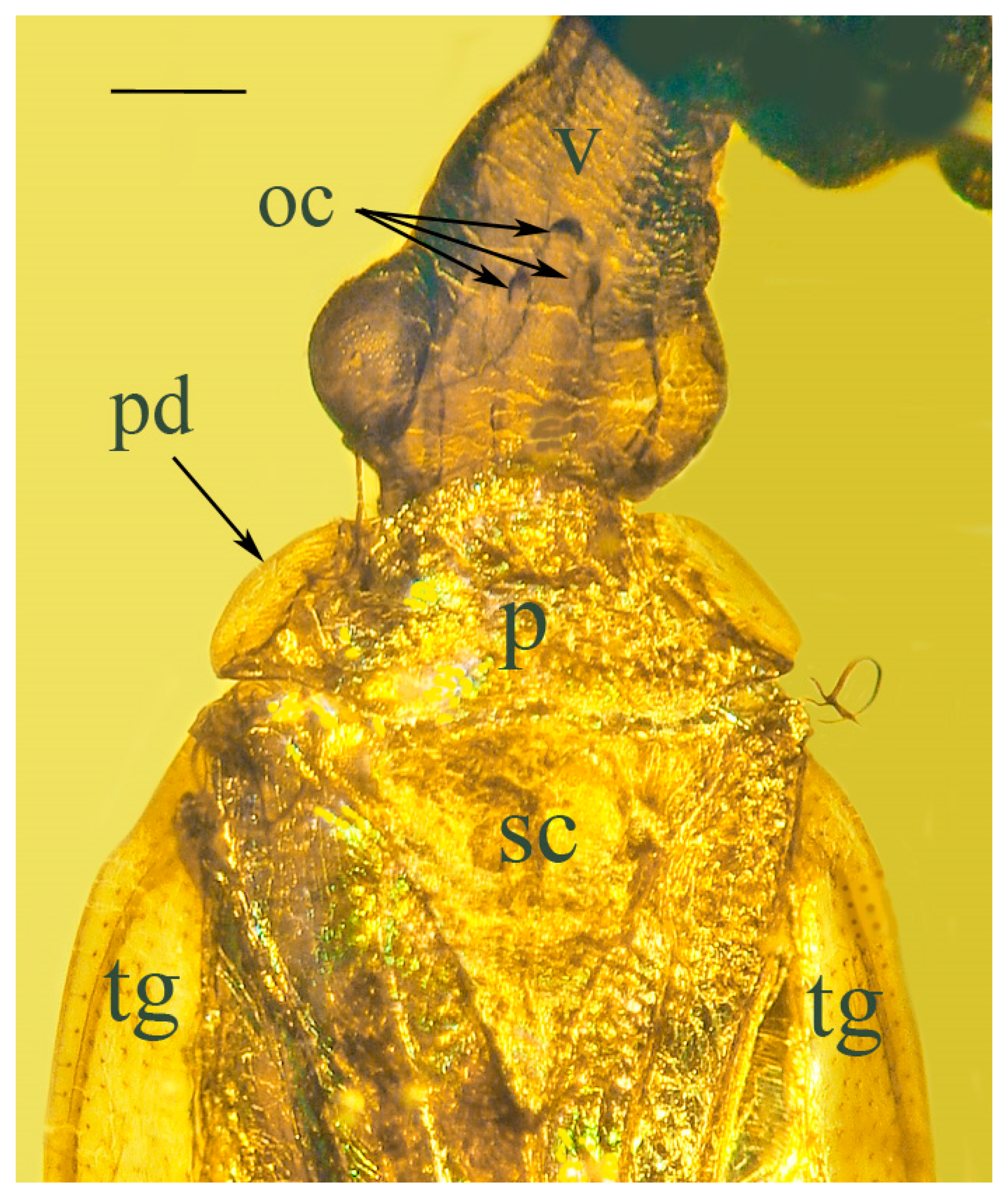
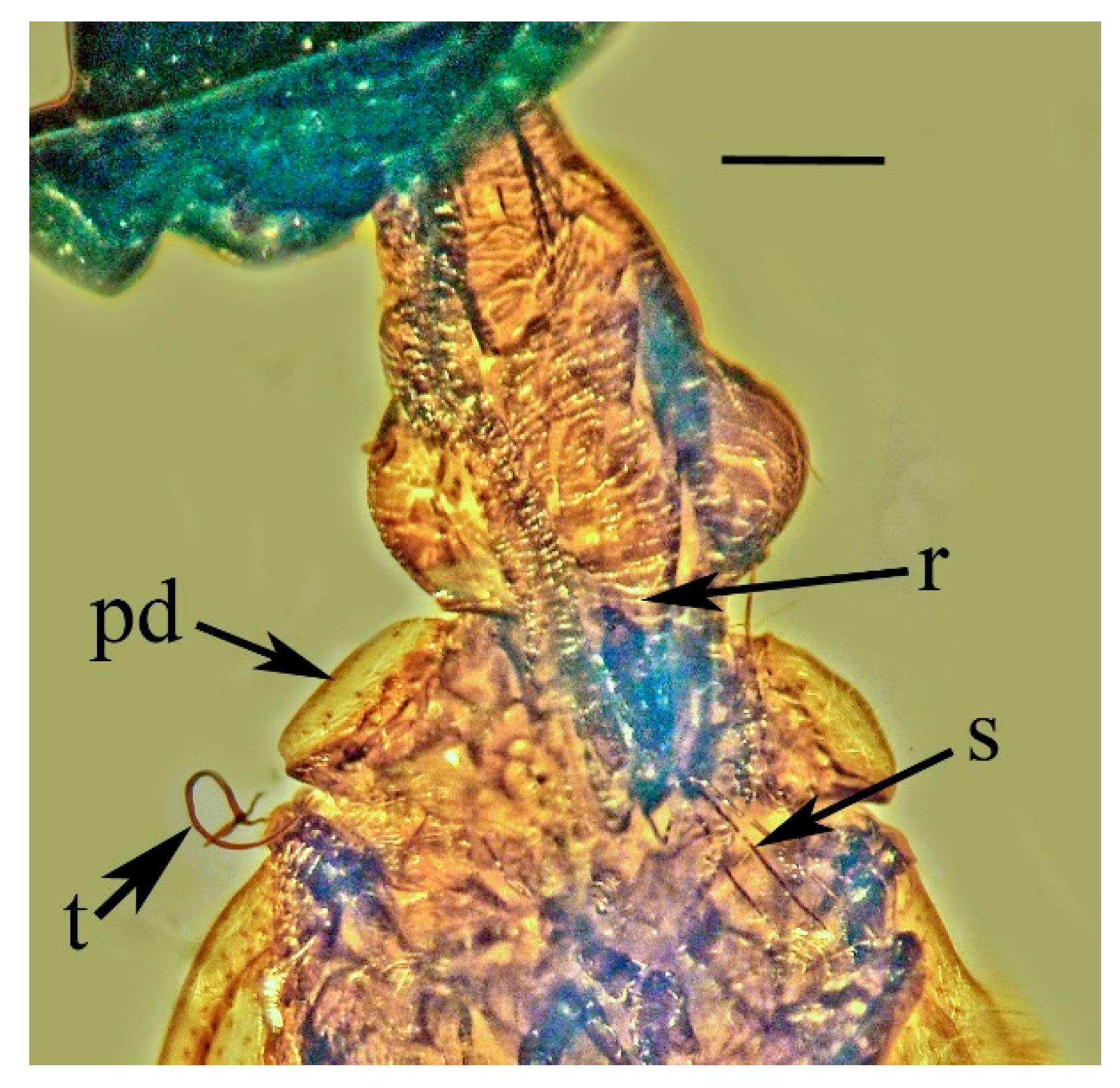
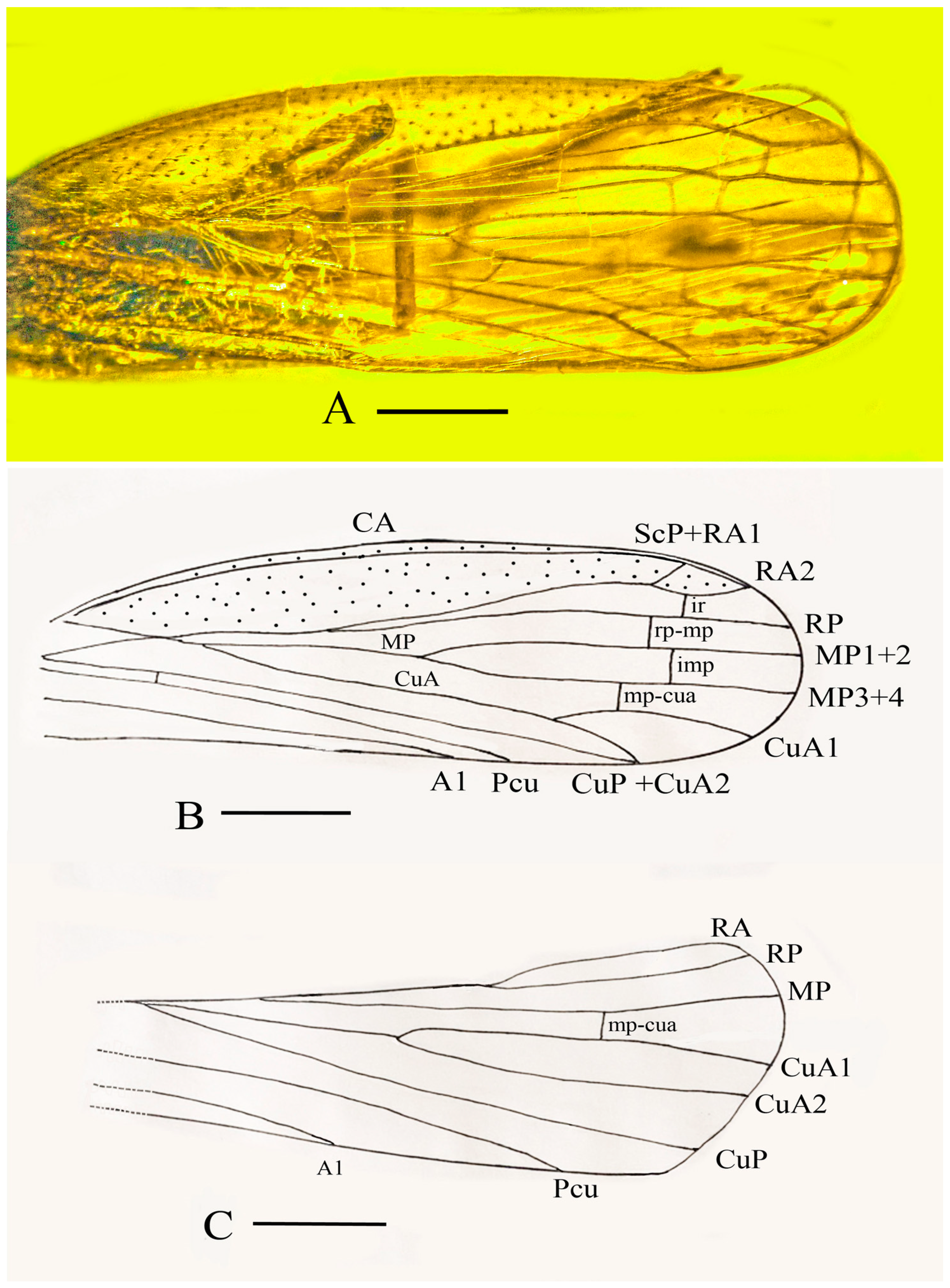
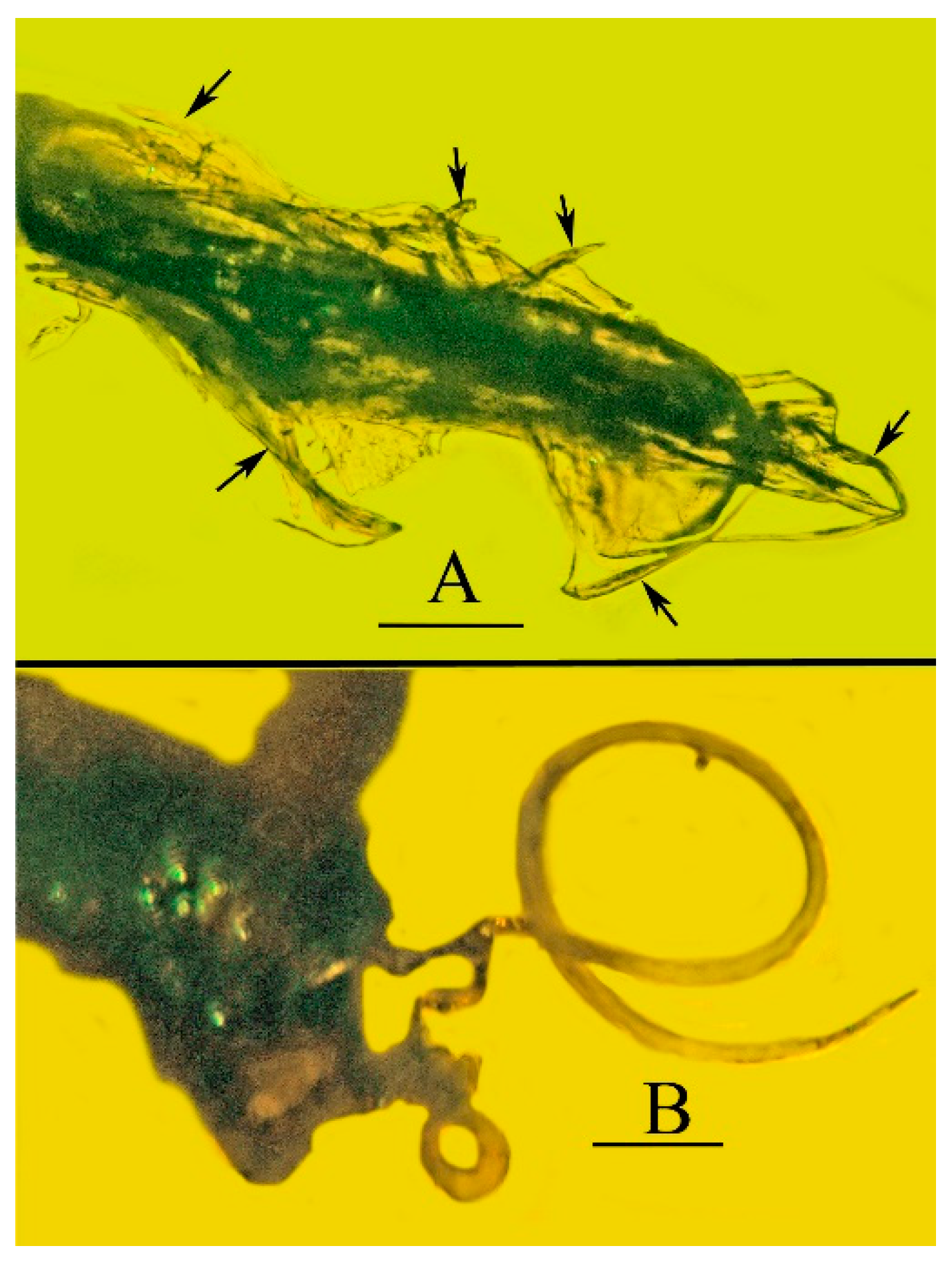
Systematic Paleontology
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.; Wang, B.; Zheng, Y.; Jiang, H.; Jiang, T.; Bozdoğan, H.; Zhang, J.; An, B.; Wang, X.; Zhang, H. Taxonomic Review and Phylogenetic Inference Elucidate the Evolutionary History of Mesozoic Procercopidae, with New Data from the Cretaceous Jehol Biota of NE China (Hemiptera: Cicadomorpha). J. Zool. Syst. Evol. Res. 2020, 58, 174–193. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, H.; Li, Y.; Zhang, Q.; Zheng, Y.; Yu, S.; Zhang, H. Overview of Sinoalidae in Cenomanian Kachin Amber from Northern Myanmar (Cicadomorpha, Cercopoidea). Cretac. Res. 2023, 142, 105399. [Google Scholar] [CrossRef]
- Cruickshank, R.D.; Ko, K. Geology of an Amber Locality in the Hukawng Valley, Northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–455. [Google Scholar] [CrossRef]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yand, M.; Lei, W.; Li, Q.; Li, X. Age Constraint on Burmese Amber Based on U-Pb Dating of Zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Poinar, G.O., Jr.; Lambert, G.J.B.; Wu, Y. Araucarian Source of Fossiliferous Burmese Amber: Spectroscopic and Anatomical Evidence. J. Bot. Res. Inst. Texas 2007, 1, 449–455. [Google Scholar]
- Poinar, G.O., Jr. Eggs, Oviposition and Maternal Care in Amber. In Fossil Behavior Compendium; Boucot, A.J., Poinar, G.O., Jr., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 152–158. [Google Scholar]
- Barber, G.W.; Ellis, W.O. Eggs of Three Cercopidae. Psyche 1922, 29, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Wigglesworth, V.B. The Principles of Insect Physiology; Methuen & Co. Ltd.: London, UK, 1965; p. 741. [Google Scholar]
- Wheeler, D.E. Egg coverings. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 312–313. [Google Scholar]
- Chen, J.; Wang, B.; Zhang, H.; Wang, X.; Zheng, X. New Fossil Procercopidae (Hemiptera: Cicadomorpha) From the Middle Jurassic of Daohugou, Inner Mongolia, China. Eur. J. Entomol. 2015, 112, 373–380. [Google Scholar] [CrossRef]
- Berendse, F.; Scheffer, M. The Angiosperm Radiation Revisited, an Ecological Explanation for Darwin’s ‘Abominable Mystery’. Ecol. Lett. 2009, 12, 865–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friis, E.M.; Crane, P.R.; Pedersen, K.R. Early Flowers and Angiosperm Evolution; Cambridge University Press: Cambridge, UK, 2011; p. 585. [Google Scholar]
- Poinar, G. Flowers in Amber; Springer Nature: Cham, Switzerland, 2022; p. 215. [Google Scholar]
- Bateman, R.M. Hunting the Snark: The Flawed Search for Mythical Jurassic Angiosperms. J. Exp. Bot. 2020, 71, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Pérez-de la Fuente, R.; Delclòs, X.; Peñalver, E.; Speranza, M.; Wierzchos, J.; Ascaso, C.; Engel, M.S. Early Evolution and Ecology of Camouflage in Insects. Proc. Natl. Acad. Sci. USA 2012, 109, 21414–21419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poinar, G., Jr. A New Fern, Cladarastega burmanica gen. et sp. nov. (Dennstaedtiaceae: Polypodiales) in Mid-Cretaceous Burmese Amber. Palaeodiversity 2021, 14, 153–160. [Google Scholar] [CrossRef]
- Thompson, J.A.; Martin, A.B. Gnarled Trichomes: An Understudied Character in Pteridium. Am. Fern J. 1996, 86, 36–51. [Google Scholar] [CrossRef]
- Wagner, G.J. Secreting Glandular Trichomes: More than Just Hairs. Plant Physiol. 1991, 96, 675–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, R.E. A Field Guide to The Beetles of North America; Houghton Mifflin Company: Boston, MA, USA, 1983; p. 368. [Google Scholar]
- Poinar, G., Jr. Insect Herbivores of Ferns Along the Pacific Northwest Coast of North America. Biosis. Biol. Syst. 2021, 2, 329–336. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P. History of Insects; Quicke, D.L.J., Ed.; Kleuver Academic Publishers: Dordrecht, The Netherlands, 2002; p. 517. [Google Scholar]
- Hamilton, K.G.A. Aphrophorinae of Polynesia (Rhynchota:Homoptera: Cercopidae). Pac. Insects 1980, 22, 347–360. [Google Scholar]
- Angelova, R.; Gjonov, L. Spittlebugs (Hemiptera:Cercopoidea) of Sarnena Sredna Gora Mts (Bulgaria). In Fauna of Sarnena Sredna Gora Mts. Part 3; Georgiev, D., Bechev, D., Yancheva, V., Eds.; Suppl. 11; Plovdiv University: Plovdiv, Bulgaria, 2022; pp. 1–9. [Google Scholar]
- Fu, Y.; Huang, D.; Engel, M.S. A New Species of the Extinct Family Procercopidae (Hemiptera: Cercopoidea) from the Jurassic of Northeastern China. Palaeoentomology 2018, 1, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Szwedo, J.; Wang, B.; Zheng, Y.; Wang, Y.; Wang, X.; Zhang, H. The First Mesozoic Froghopper in Amber from Northern Myanmar (Hemiptera, Cercopoidea, Sinoalidae). Cretac. Res. 2018, 85, 243–249. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poinar, G., Jr.; Brown, A.E. Araeoanasillus leptosomus gen. et sp. nov., (Hemiptera, Cercopoidea, Sinoalidae?), a New Froghopper from Mid-Cretaceous Burmese Amber with Evidence of Its Possible Host Plant. Life 2023, 13, 922. https://doi.org/10.3390/life13040922
Poinar G Jr., Brown AE. Araeoanasillus leptosomus gen. et sp. nov., (Hemiptera, Cercopoidea, Sinoalidae?), a New Froghopper from Mid-Cretaceous Burmese Amber with Evidence of Its Possible Host Plant. Life. 2023; 13(4):922. https://doi.org/10.3390/life13040922
Chicago/Turabian StylePoinar, George, Jr., and Alex E. Brown. 2023. "Araeoanasillus leptosomus gen. et sp. nov., (Hemiptera, Cercopoidea, Sinoalidae?), a New Froghopper from Mid-Cretaceous Burmese Amber with Evidence of Its Possible Host Plant" Life 13, no. 4: 922. https://doi.org/10.3390/life13040922