
Bioactive Compounds and Signaling Pathways of Wolfiporia extensa in Suppressing Inflammatory Response by Network Pharmacology


 ,
, 
Abstract
:1. Introduction
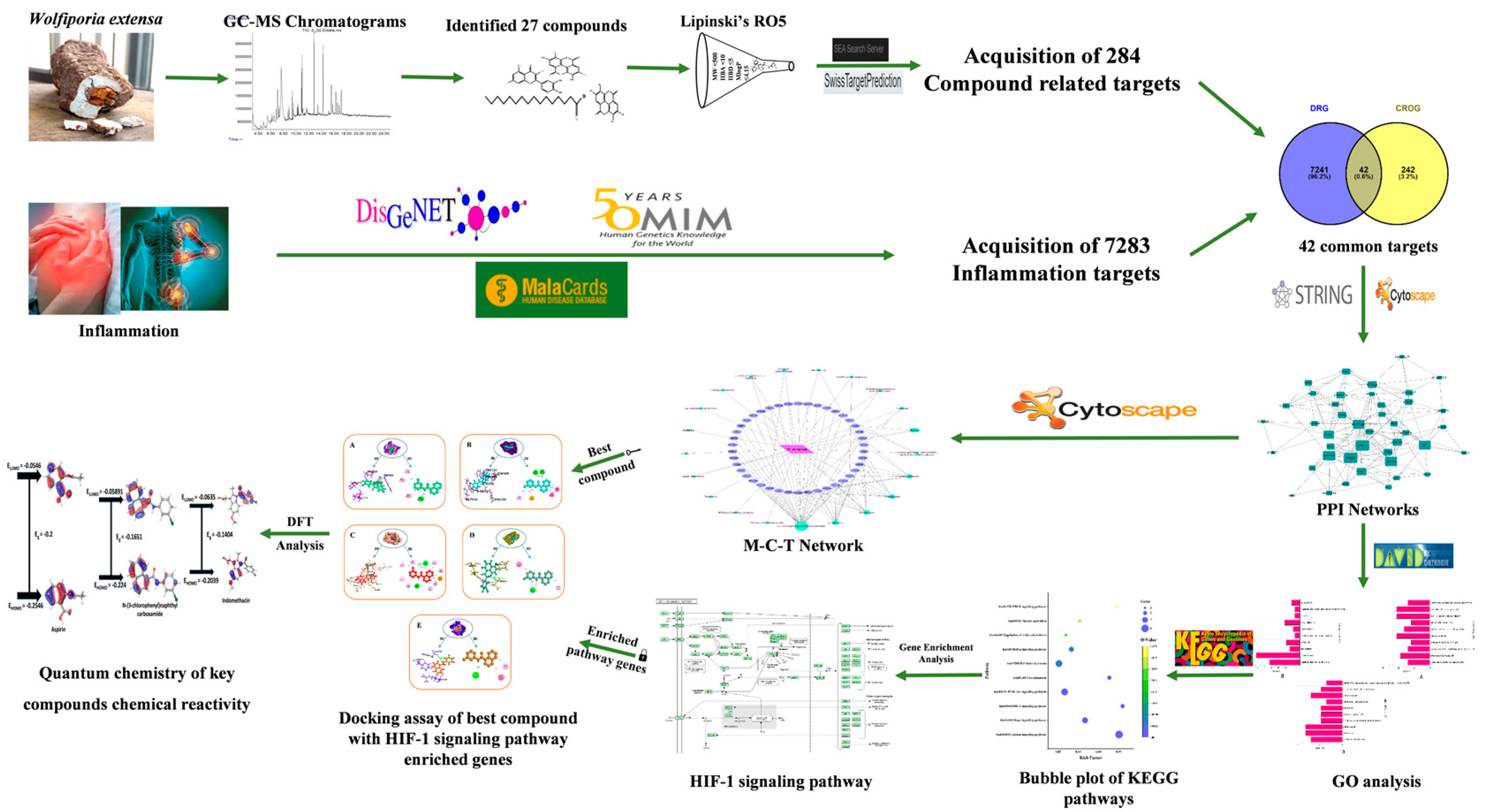
2. Materials and Methods
2.1. Mushroom Collection, Identification, and Extraction
2.2. GC-MS Analysis
2.3. Bioactive Compounds Filtration
2.4. Extraction of Compound Associated Targets and Inflammatory Targets
2.5. Common Targets Network Construction
2.6. Mushroom-Bioactives-Targets Network Construction
2.7. Analysis of GO and KEGG Pathway Involvement within Common Targets
2.8. Preparation of Ligand and Receptor Protein
2.9. Molecular Docking Simulation Using Glide
2.10. Quantum Chemistry of Frontier Molecular Orbitals
3. Results
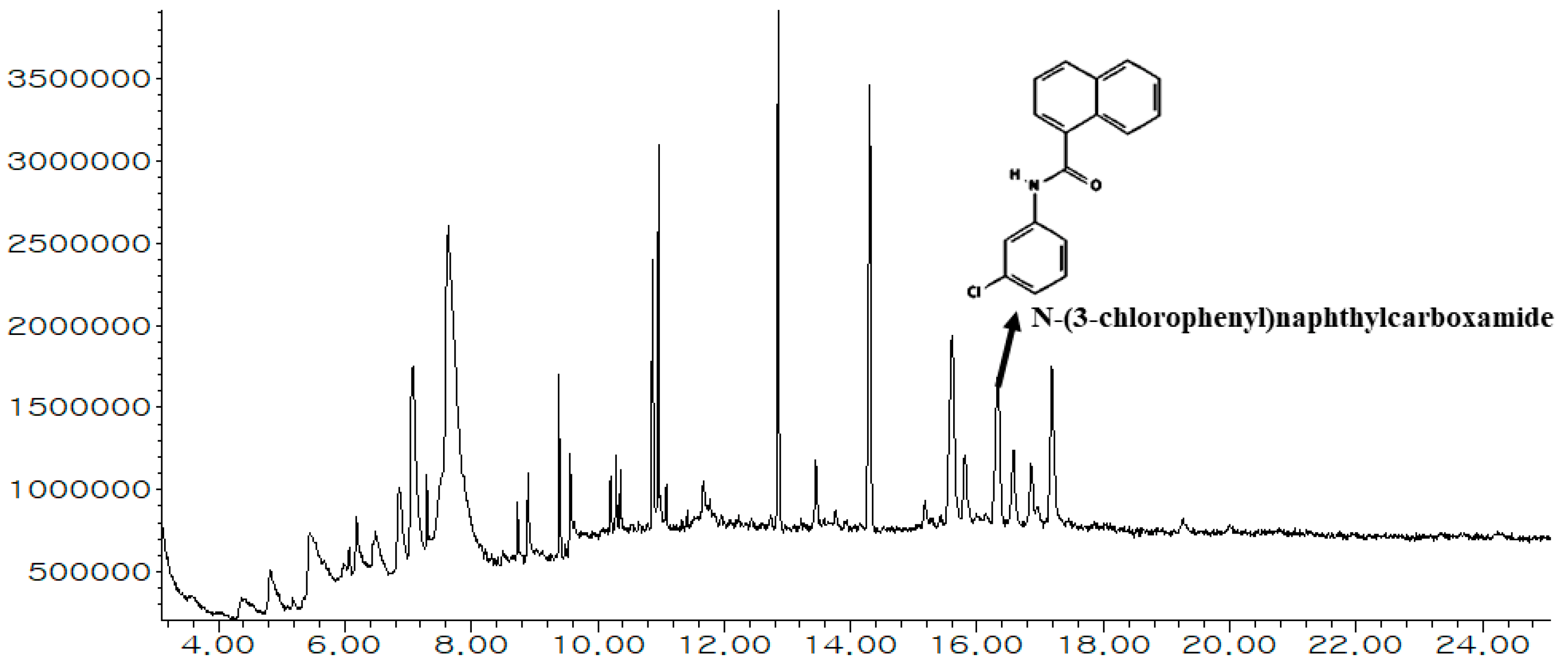
3.1. Chemical Composition of MEWE
3.2. Bioactive Compounds Filtration
3.3. Common Targets of Bioactives Intersected from SEA and STP Databases
3.4. Identification of Inflammation Targets within Disease Targets and 284 Common Targets
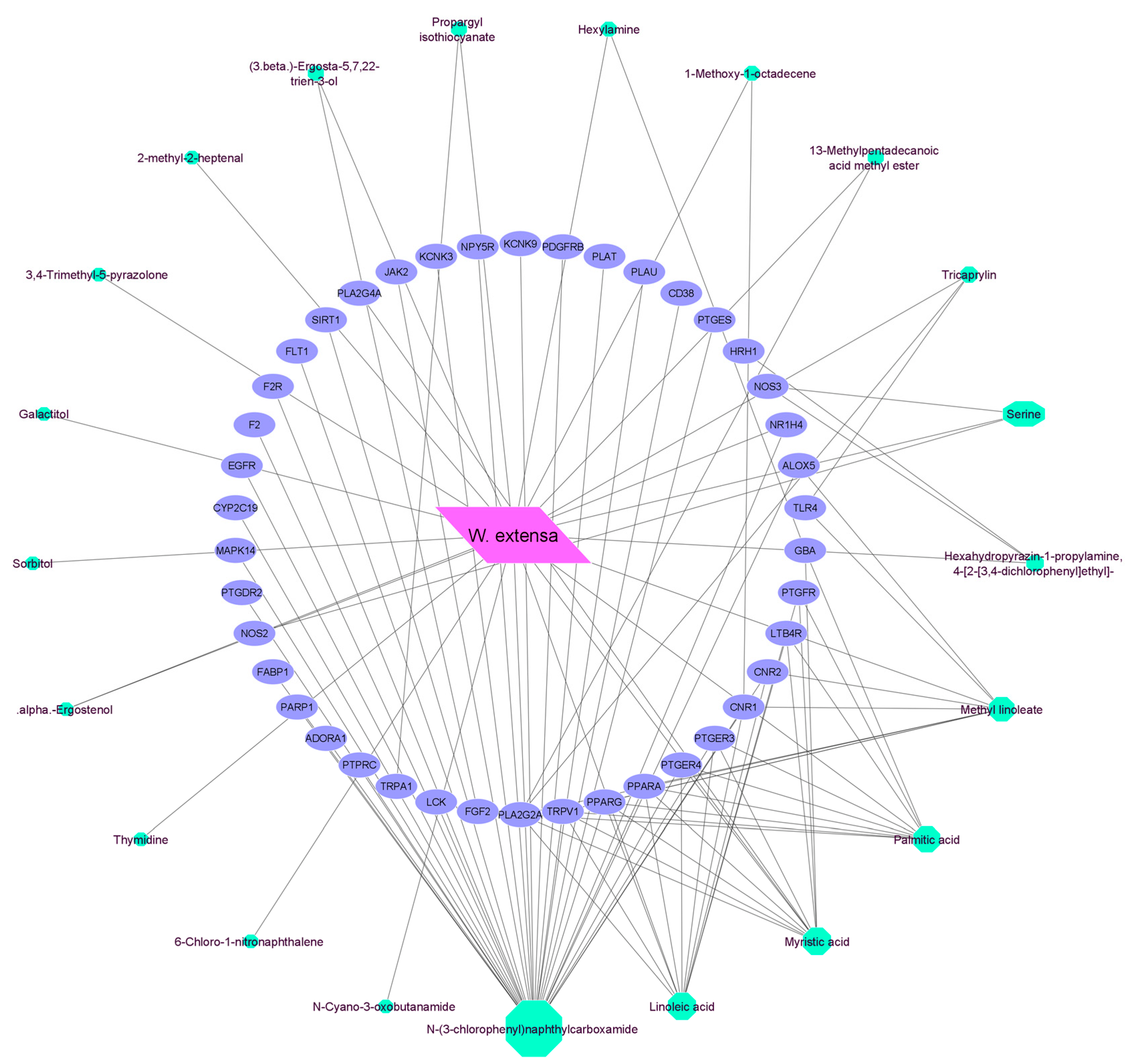
3.5. Network Construction of 42 Common Targets
3.6. Analysis of Mushroom-Compound-Targets Network
3.7. Gene Ontology (GO) Analysis of 42 Common Targets
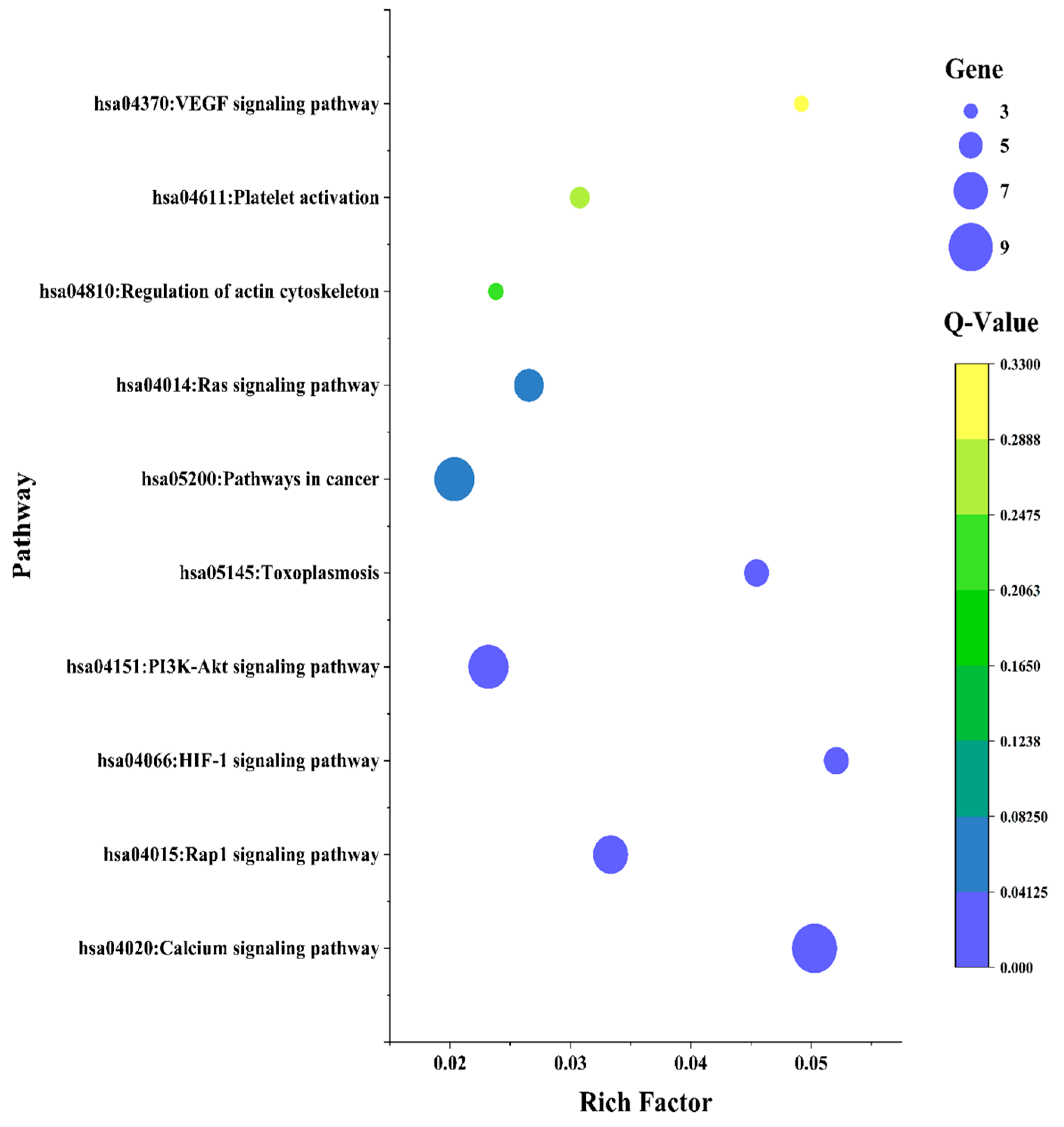
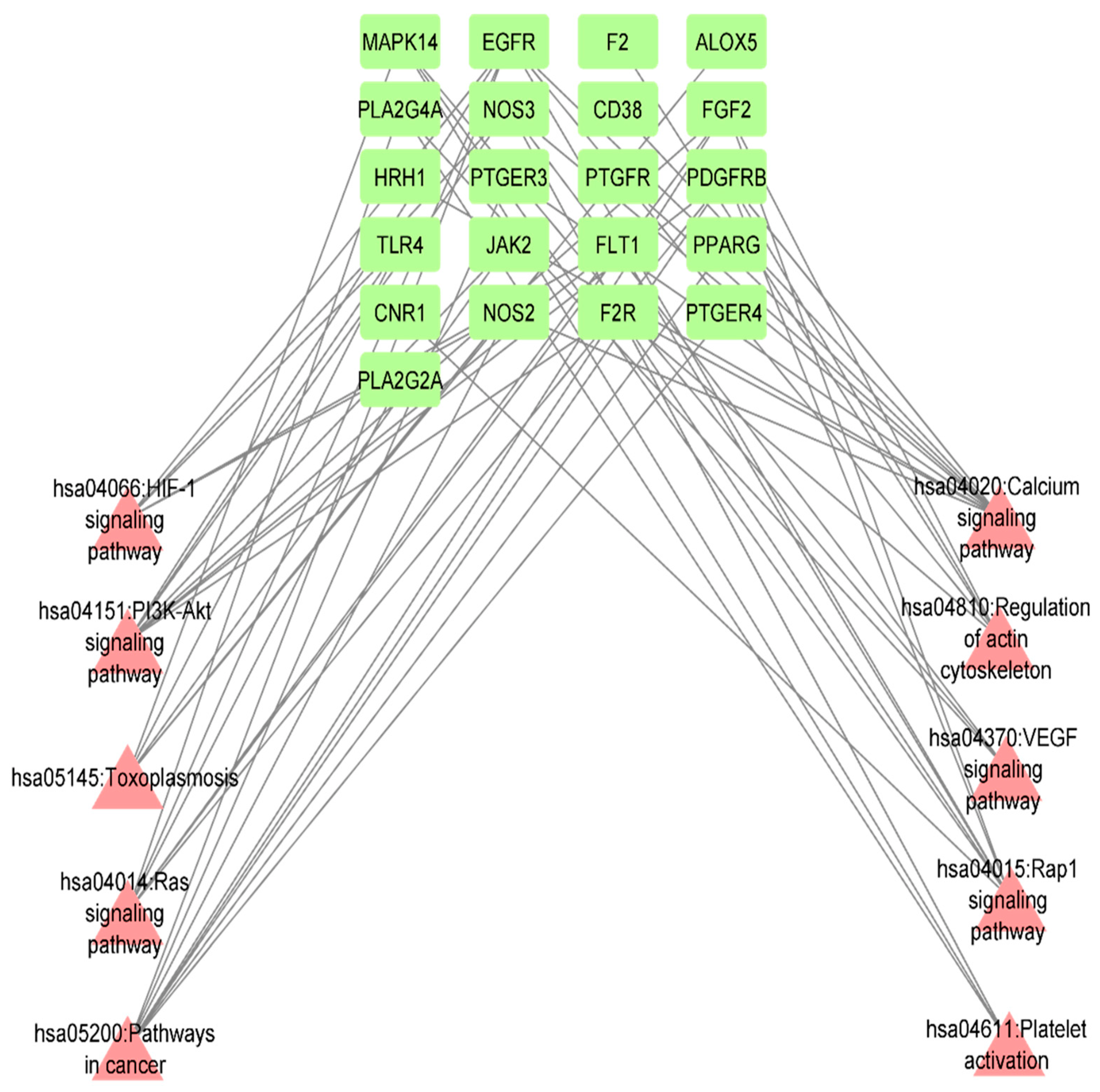
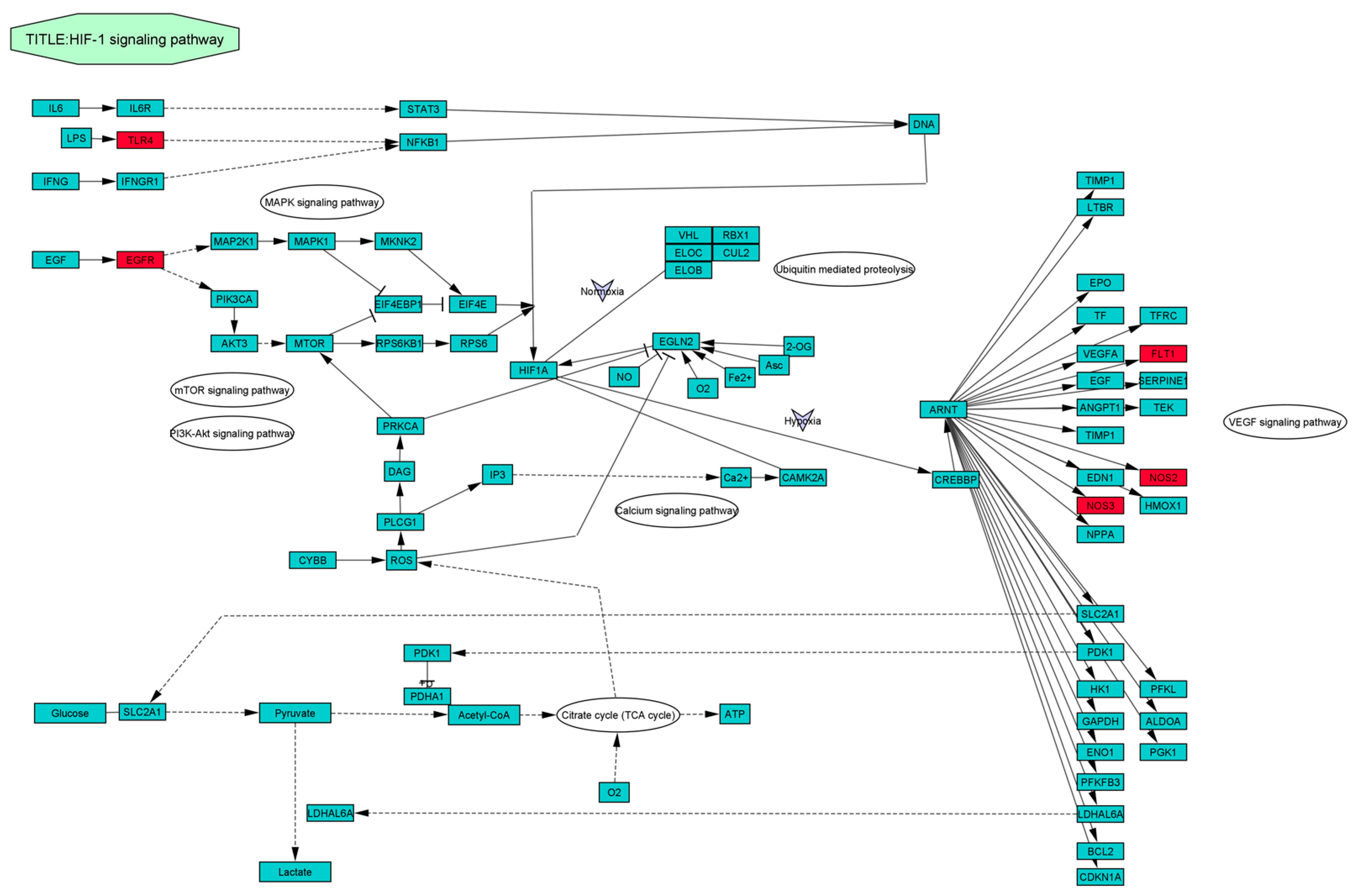
3.8. KEGG Pathway Enrichment Analysis of Identified 42 Common Targets
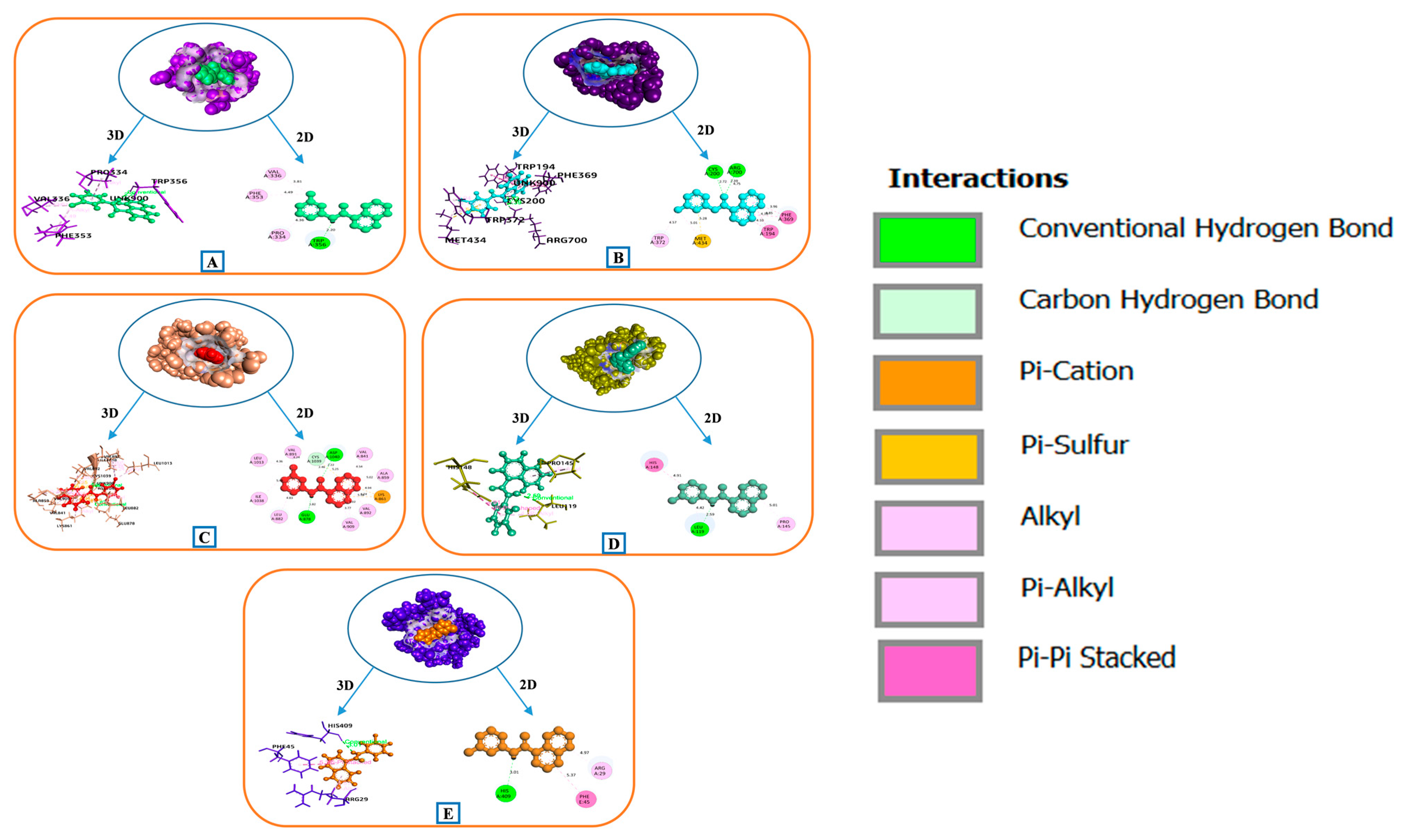
3.9. Docking Score Assessment of Key Bioactives and 5 HIF-1 Signaling Pathway Targets
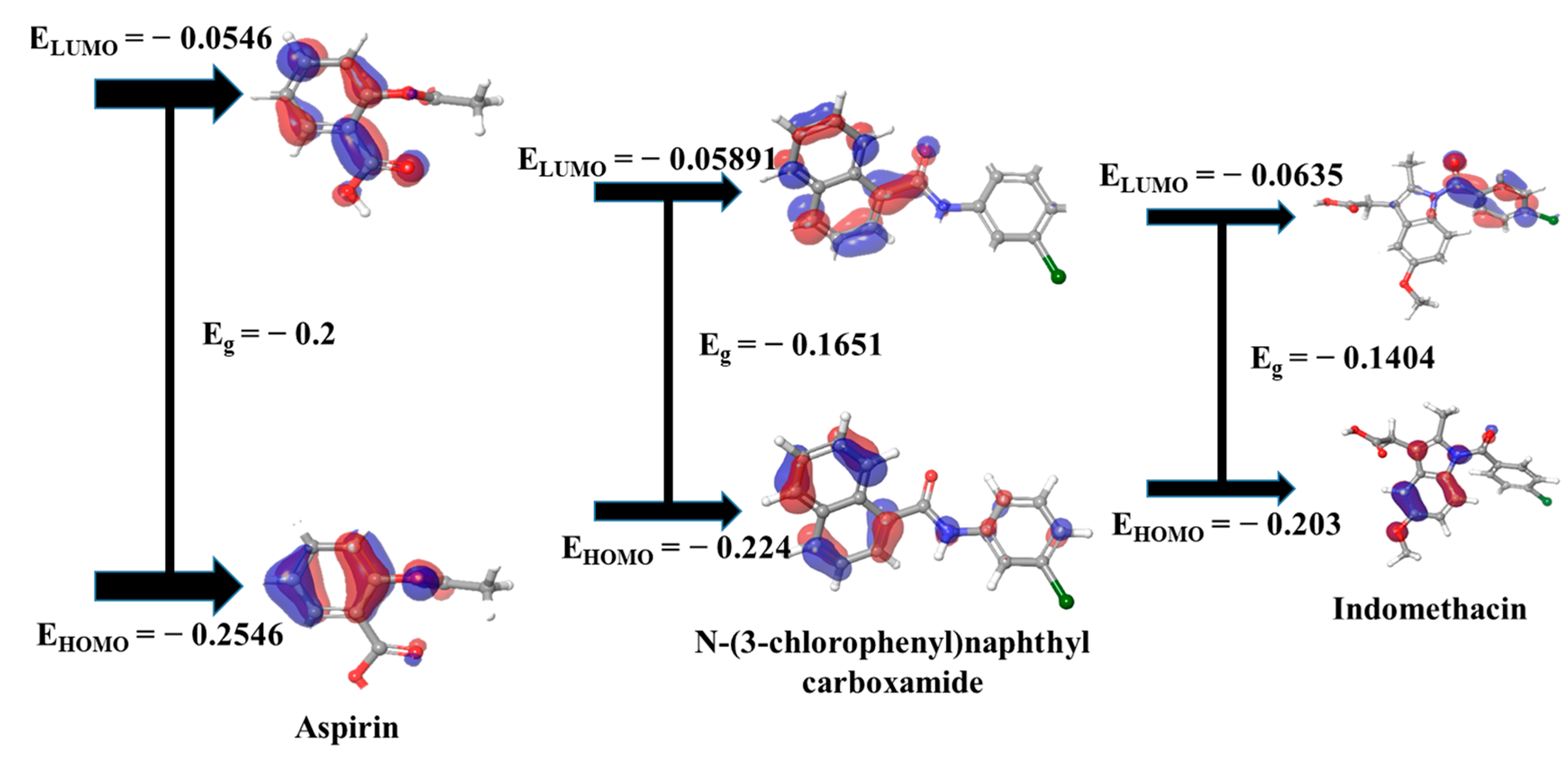
3.10. DFT Evaluation of Key Compound and Standard Drugs
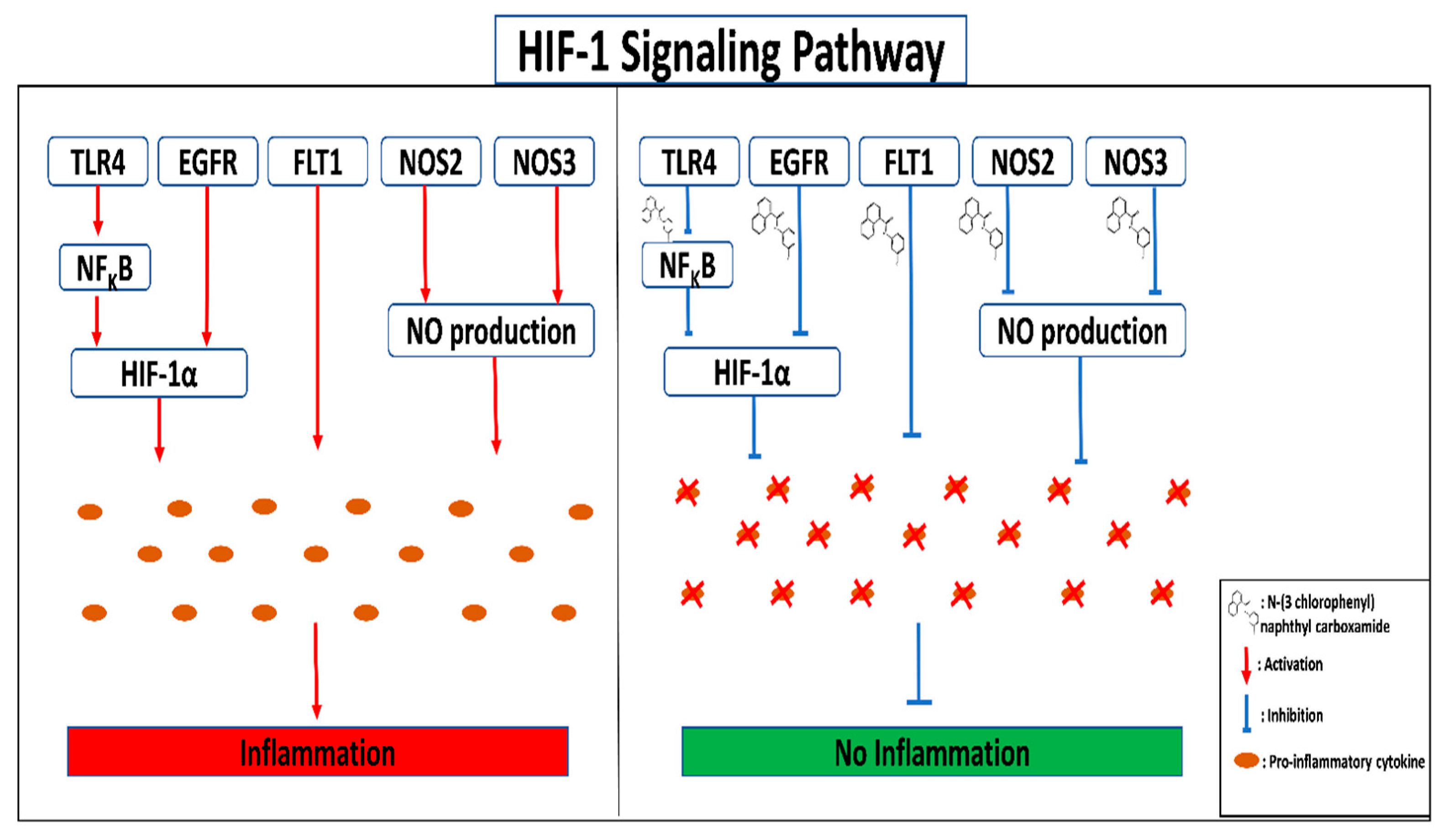
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SEA | Similarity Ensemble Approach |
| STP | Swiss Target Prediction |
| OMIM | Online Mendelian Inheritance in Man |
| MEWE | Methanolic Extract of Wolfiporia extensa |
| HIF | Hypoxia Inducible Factor |
| MNC | Maximum Neighborhood Component |
| MCC | Maximal Clique Centrality |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| FDR | False discovery rate |
| PPI | Protein-protein interaction |
| SMILES | Simplified molecular input line entry system |
| NFkB | Nuclear factor kappa B |
| EGFR | Epidermal Growth Factor Receptor |
| TLR4 | Toll Like Receptor 4 |
| FLT1 | fms related tyrosine kinase 1 |
| NOS2 | Nitric Oxide Synthase 2 |
| NOS3 | Nitric Oxide Synthase 23 |
References
- Ilic, N.M.; Dey, M.; Poulev, A.A.; Logendra, S.; Kuhn, P.E.; Raskin, I. Anti-inflammatory activity of grains of paradise (Aframomum Melegueta Schum) extract. J. Agric. Food Chem. 2014, 62, 10452–10457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Qiao, Y.; Zhang, Z.; Deng, Y.; Chen, T.; Tao, L.; Xu, Q.; Liu, J.; Sun, W.; Ye, Y.; et al. New Polyketides with Anti-Inflammatory Activity from the Fungus Aspergillus rugulosa. Front. Pharmacol. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Inflammation 2010: New Adventures of an Old Flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, A.S.; Diwan, P.V.; Jain, N.K.; Tomalia, D.A. Unexpected in vivo anti-inflammatory activity observed for simple, surface functionalized poly(amidoamine) dendrimers. Biomacromolecules 2009, 10, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.C.; Skovbakke, S.L.; Masoudi, H.; Hancock, R.E.W.; Franzyk, H. In Vivo Anti-inflammatory Activity of Lipidated Peptidomimetics Pam-(Lys-βNspe)6-NH2 and Lau-(Lys-βNspe)6-NH2 against PMA-Induced Acute Inflammation. Front. Immunol. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L.; Chen, L.; et al. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Taofiq, O.; Calhelha, R.C.; Heleno, S.; Barros, L.; Martins, A.; Santos-Buelga, C.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. The contribution of phenolic acids to the anti-inflammatory activity of mushrooms: Screening in phenolic extracts, individual parent molecules and synthesized glucuronated and methylated derivatives. Food Res. Int. 2015, 76, 821–827. [Google Scholar] [CrossRef] [Green Version]
- Kanwar, J.R.; Kanwar, R.K.; Burrow, H.; Baratchi, S. Recent advances on the roles of NO in cancer and chronic inflammatory disorders. Curr. Med. Chem. 2009, 16, 2373–2394. [Google Scholar] [CrossRef]
- Moro, C.; Palacios, I.; Lozano, M.; D’Arrigo, M.; Guillamón, E.; Villares, A.; Martínez, J.A.; García-Lafuente, A. Anti-inflammatory activity of methanolic extracts from edible mushrooms in LPS activated RAW 264.7 macrophages. Food Chem. 2012, 130, 350–355. [Google Scholar] [CrossRef]
- Taofiq, O.; Martins, A.; Barreiro, M.F.; Ferreira, I.C.F.R. Anti-inflammatory potential of mushroom extracts and isolated metabolites. Trends Food Sci. Technol. 2016, 50, 193–210. [Google Scholar] [CrossRef] [Green Version]
- Gunawardena, D.; Bennett, L.; Shanmugam, K.; King, K.; Williams, R.; Zabaras, D.; Head, R.; Ooi, L.; Gyengesi, E.; Münch, G. Anti-inflammatory effects of five commercially available mushroom species determined in lipopolysaccharide and interferon-γ activated murine macrophages. Food Chem. 2014, 148, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Jayasuriya, W.J.A.B.N.; Handunnetti, S.M.; Wanigatunge, C.A.; Fernando, G.H.; Abeytunga, D.T.U.; Suresh, T.S. Anti-Inflammatory Activity of Pleurotus ostreatus, a Culinary Medicinal Mushroom, in Wistar Rats. Evid. Based Complement. Altern. Med. 2020, 2020, 6845383. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.S.; Tan, N.H.; Fung, S.Y.; Sim, S.M.; Tan, C.S.; Ng, S.T. Anti-inflammatory effect of the sclerotium of Lignosus rhinocerotis (Cooke) Ryvarden, the Tiger Milk mushroom. BMC Complement. Altern. Med. 2014, 14, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kalač, P. A review of chemical composition and nutritional value of wild-growing and cultivated mushrooms. J. Sci. Food Agric. 2013, 93, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Dore, C.M.P.G.; Alves, M.G.d.C.F.; Santos, M.D.G.L.; Souza, L.A.R.D.; Baseia, I.G.; Leite, E.L. Antioxidant and Anti-Inflammatory Properties of an Extract Rich in Polysaccharides of the Mushroom Polyporus dermoporus. Antioxidants 2014, 3, 730–744. [Google Scholar] [CrossRef]
- Yuan, B.; Zhao, L.; Rakariyatham, K.; Han, Y.; Gao, Z.; Kimatu, B.M.; Hu, Q.; Xiao, H. Isolation of a novel bioactive protein from an edible mushroom Pleurotus eryngii and its anti-inflammatory potential. Food Funct. 2017, 8, 2175–2183. [Google Scholar] [CrossRef] [PubMed]
- Souilem, F.; Fernandes, Â.; Calhelha, R.C.; Barreira, J.C.M.; Barros, L.; Skhiri, F.; Martins, A.; Ferreira, I.C.F.R. Wild mushrooms and their mycelia as sources of bioactive compounds: Antioxidant, anti-inflammatory and cytotoxic properties. Food Chem. 2017, 230, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, D.; Tabernero, M.; Largo, C.; Polo, G.; Piris, A.J.; Soler-Rivas, C. Effect of traditional and modern culinary processing, bioaccessibility, biosafety and bioavailability of eritadenine, a hypocholesterolemic compound from edible mushrooms. Food Funct. 2018, 9, 6360–6368. [Google Scholar] [CrossRef] [Green Version]
- Haque, M.A.; Reza, A.A.; Nasrin, M.S.; Rahman, M.A. Pleurotus highking mushrooms potentiate antiproliferative and antimigratory activity against triple-negative breast cancer cells by suppressing Akt signaling. Integr. Cancer Ther. 2020, 19, 1534735420969809. [Google Scholar] [CrossRef]
- Park, W.; Joo, S.; Park, K.; Chang, Y.; Kim, C. Effects of the Geiji-Bokryung-Hwan on carrageenan-induced inflammation in mice and cyclooxygenase-2 in hepatoma cells of HepG2 and Hep3B. Immunopharmacol. Immunotoxicol. 2004, 26, 103–112. [Google Scholar] [CrossRef]
- Lu, M.K.; Cheng, J.J.; Lin, C.Y.; Chang, C.C. Purification, structural elucidation, and anti-inflammatory effect of a water-soluble 1,6-branched 1,3-α-d-galactan from cultured mycelia of Poria cocos. Food Chem. 2010, 118, 349–356. [Google Scholar] [CrossRef]
- Liu, X.; Yu, X.; Xu, X.; Zhang, X.; Zhang, X. The protective effects of Poria cocos-derived polysaccharide CMP33 against IBD in mice and its molecular mechanism. Food Funct. 2018, 9, 5936–5949. [Google Scholar] [CrossRef]
- Park, W.-H.; Lee, S.-K.; Oh, H.-K.; Bae, J.-Y.; Kim, C.-H. Tumor initiation inhibition through inhibition COX-1 activity of a traditional Korean herbal prescription, Geiji-Bokryung-Hwan, in human hepatocarcinoma cells. Immunopharmacol. Immunotoxicol. 2005, 27, 473–483. [Google Scholar] [CrossRef]
- Nie, A.; Chao, Y.; Zhang, X.; Jia, W.; Zhou, Z.; Zhu, C. Phytochemistry and Pharmacological Activities of Wolfiporia cocos (F.A. Wolf) Ryvarden & Gilb. Front. Pharmacol. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Lee, S.R.; Lee, S.; Moon, E.; Park, H.J.; Park, H.B.; Kim, K.H. Bioactivity-guided isolation of anti-inflammatory triterpenoids from the sclerotia of Poria cocos using LPS-stimulated Raw264.7 cells. Bioorg. Chem. 2017, 70, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, A.L. Network pharmacology. Nat. Biotechnol. 2007, 25, 1110–1111. [Google Scholar] [CrossRef]
- Chen, W.; Li, J.; Li, C.; Fan, H.N.; Zhang, J.; Zhu, J.S. Network pharmacology-based identification of the antitumor effects of taraxasterol in gastric cancer. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420933107. [Google Scholar] [CrossRef]
- Jiang, L.; Lu, J.; Qin, Y.; Jiang, W.; Wang, Y. Antitumor effect of guava leaves on lung cancer: A network pharmacology study. Arab. J. Chem. 2020, 13, 7773–7797. [Google Scholar] [CrossRef]
- Chen, Y.; Kern, T.S.; Kiser, P.D.; Palczewski, K. Eyes on systems pharmacology. Pharmacol. Res. 2016, 114, 39–41. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Huang, J.; Wang, N.; Tan, H.Y.; Cheung, F.; Chen, F.; Feng, Y. Integrating network pharmacology and pharmacological evaluation for deciphering the action mechanism of herbal formula Zuojin Pill in suppressing hepatocellular carcinoma. Front. Pharmacol. 2019, 10, 1–21. [Google Scholar] [CrossRef]
- Oh, K.; Adnan, M.; Cho, D. Uncovering Mechanisms of Zanthoxylum piperitum Fruits for the Alleviation of Rheumatoid Arthritis Based on Network Pharmacology. Biology 2021, 10, 703. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, W.; Huang, C.; Li, Y.; Yu, H.; Wang, Y.; Duan, J.; Ling, Y. A Novel Chemometric Method for the Prediction of Human Oral Bioavailability. Int. J. Mol. Sci. 2012, 13, 6964–6982. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappaport, N.; Twik, M.; Plaschkes, I.; Nudel, R.; Iny Stein, T.; Levitt, J.; Gershoni, M.; Morrey, C.P.; Safran, M.; Lancet, D. MalaCards: An amalgamated human disease compendium with diverse clinical and genetic annotation and structured search. Nucleic Acids Res. 2017, 45, D877–D887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamosh, A.; Scott, A.F.; Amberger, J.S.; Bocchini, C.A.; McKusick, V.A. Online Mendelian Inheritance in Man (OMIM), a knowledgebase of human genes and genetic disorders. Nucleic Acids Res. 2005, 33, D514. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software Environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Lu, C.; Hu, X.; Wang, G.; Leach, L.J.; Yang, S.; Kearsey, M.J.; Luo, Z.W. Why do essential proteins tend to be clustered in the yeast interactome network? Mol. Biosyst. 2010, 6, 871–877. [Google Scholar] [CrossRef]
- Chin, C.; Chen, S.; Wu, H.; Ho, C.; Ko, M.; Lin, C. cytoHubba: Identifying hub objects and sub- networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Shi, X.; Li, X.; Zhang, L.; Wu, P.; Mao, J.; Xing, R.; Zhang, N.; Wang, P. Network Pharmacology Approach to Uncover the Mechanism Governing the Effect of Simiao Powder on Knee Osteoarthritis. Biomed Res. Int. 2020, 2020, 6971503. [Google Scholar] [CrossRef]
- Xiong, Y.; Yang, Y.; Xiong, W.; Yao, Y.; Wu, H.; Zhang, M. Network pharmacology-based research on the active component and mechanism of the antihepatoma effect of Rubia cordifolia L. J. Cell. Biochem. 2019, 120, 12461–12472. [Google Scholar] [CrossRef] [PubMed]
- Rudra, S.; Tahamina, A.; Emon, N.U.; Adnan, M.; Shakil, M.; Chowdhury, M.H.U.; Barlow, J.W.; Alwahibi, M.S.; Soliman Elshikh, M.; Faruque, M.O.; et al. Evaluation of Various Solvent Extracts of Tetrastigma leucostaphylum (Dennst.) Alston Leaves, a Bangladeshi Traditional Medicine Used for the Treatment of Diarrhea. Molecules 2020, 25, 4994. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Nazim Uddin Chy, M.; Mostafa Kamal, A.T.M.; Azad, M.O.K.; Paul, A.; Uddin, S.B.; Barlow, J.W.; Faruque, M.O.; Park, C.H.; Cho, D.H. Investigation of the biological activities and characterization of bioactive constituents of ophiorrhiza rugosa var. prostrata (D.Don) & Mondal leaves through in vivo, in vitro, and in silico approaches. Molecules 2019, 24, 1367. [Google Scholar] [CrossRef] [Green Version]
- Shovo, M.A.R.B.; Tona, M.R.; Mouah, J.; Islam, F.; Chowdhury, M.H.U.; Das, T.; Paul, A.; Ağagündüz, D.; Rahman, M.M.; Emran, T.B.; et al. Computational and Pharmacological Studies on the Antioxidant, Thrombolytic, Anti-Inflammatory, and Analgesic Activity of Molineria capitulata. Curr. Issues Mol. Biol. 2021, 43, 434–456. [Google Scholar] [CrossRef]
- Bochevarov, A.D.; Harder, E.; Hughes, T.F.; Greenwood, J.R.; Braden, D.A.; Philipp, D.M.; Rinaldo, D.; Halls, M.D.; Zhang, J.; Friesner, R.A. Jaguar: A high-performance quantum chemistry software program with strengths in life and materials sciences. Int. J. Quantum Chem. 2013, 113, 2110–2142. [Google Scholar] [CrossRef]
- Ko, S.G.; Yin, C.S.; Du, B.; Kim, K. Herbal medicines for inflammatory diseases. Mediators Inflamm. 2014, 2014, 982635. [Google Scholar] [CrossRef]
- Qi, Q.; Li, R.; Li, H.Y.; Cao, Y.B.; Bai, M.; Fan, X.J.; Wang, S.Y.; Zhang, B.; Li, S. Identification of the anti-tumor activity and mechanisms of nuciferine through a network pharmacology approach. Acta Pharmacol. Sin. 2016, 37, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, S.; Anand, V.; Roy, S. Vascular Endothelial growth factor signaling in hypoxia and Inflammation. J. Neuroimmune Pharmacol. 2014, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.-A.; Bae, D.-G.; Ryoo, J.-W.; Kim, H.-R.; Park, G.-S.; Cho, C.-S.; Chae, C.-B.; Kim, W.-U. Arginine-Rich Anti-Vascular Endothelial Growth Factor (Anti-VEGF) Hexapeptide Inhibits Collagen-Induced Arthritis and VEGF-Stimulated Productions of TNF-α and IL-6 by Human Monocytes. J. Immunol. 2005, 174, 5846–5855. [Google Scholar] [CrossRef] [Green Version]
- Arman, M.; Payne, H.; Ponomaryov, T.; Brill, A. Role of Platelets in Inflammation. In The Non-Thrombotic Role of Platelets in Health and Disease; IntechOpen: London, UK, 2015. [Google Scholar] [CrossRef]
- Kopecki, Z.; Ludwig, R.J.; Cowin, A.J. Cytoskeletal Regulation of Inflammation and Its Impact on Skin Blistering Disease Epidermolysis Bullosa Acquisita. Int. J. Mol. Sci. 2016, 17, 1116. [Google Scholar] [CrossRef] [Green Version]
- Catanzaro, J.M.; Sheshadri, N.; Pan, J.-A.; Sun, Y.; Shi, C.; Li, J.; Powers, R.S.; Crawford, H.C.; Zong, W.-X. Oncogenic Ras induces inflammatory cytokine production by upregulating the squamous cell carcinoma antigens SerpinB3/B4. Nat. Commun. 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Multhoff, G.; Molls, M.; Radons, J. Chronic Inflammation in Cancer Development. Front. Immunol. 2012, 2, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, M.B.; Jensen, K.D.C.; Saeij, J.P.J. Toxoplasma gondii effectors are master regulators of the inflammatory response. Trends Parasitol. 2011, 27, 487–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nidai Ozes, O.; Mayo, L.D.; Gustin, J.A.; Pfeffer, S.R.; Pfeffer, L.M.; Donner, D.B. NF-κB activation by tumour necrosis factor requires the Akt serine–threonine kinase. Nature 1999, 401, 82–85. [Google Scholar] [CrossRef]
- Taylor, C.T. Interdependent roles for hypoxia inducible factor and nuclear factor-κB in hypoxic inflammation. J. Physiol. 2008, 586, 4055–4059. [Google Scholar] [CrossRef]
- Cai, Y.; Sukhova, G.K.; Wong, H.K.; Xu, A.; Tergaonkar, V.; Vanhoutte, P.M.; Tang, E.H.C. Rap1 induces cytokine production in pro-inflammatory macrophages through NFκB signaling and is highly expressed in human atherosclerotic lesions. Cell Cycle 2015, 14, 3580. [Google Scholar] [CrossRef] [Green Version]
- Dalal, P.J.; Muller, W.A.; Sullivan, D.P. Endothelial Cell Calcium Signaling during Barrier Function and Inflammation. Am. J. Pathol. 2020, 190, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; He, G.; Zhang, W.; Xu, T.; Qi, H.; Li, J.; Zhang, Y.; Gao, M.Q. Stromal fibroblasts derived from mammary gland of bovine with mastitis display inflammation-specific changes. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Konisti, S.; Kiriakidis, S.; Paleolog, E.M. Hypoxia-a key regulator of angiogenesis and inflammation in rheumatoid arthritis. Nat. Rev. Rheumatol. 2012, 8, 153–162. [Google Scholar] [CrossRef]
- Wilson, K.T.; Fu, S.; Ramanujam, K.S.; Meltzer, S.J. Increased Expression of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 in Barrett’s Esophagus and Associated Adenocarcinomas. Cancer Res. 1998, 58, 2929–2934. [Google Scholar]
- Ying, L.; Hofseth, L.J. An Emerging Role for Endothelial Nitric Oxide Synthase in Chronic Inflammation and Cancer. Cancer Res. 2007, 67, 1407–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.-A.; Yoon, H.-J.; Kim, H.-S.; Chae, C.-B.; De Falco, S.; Cho, C.-S.; Kim, W.-U. Role of placenta growth factor and its receptor flt-1 in rheumatoid inflammation: A link between angiogenesis and inflammation. Arthritis Rheum. 2009, 60, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Olson, N.; Vliet, A. van der Interactions between Nitric Oxide and Hypoxia-Inducible Factor Signaling Pathways in Inflammatory Disease. Nitric Oxide 2011, 25, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.M.; Shawon, J.; Halim, M.A. Multiple receptor conformers based molecular docking study of fluorine enhanced ethionamide with mycobacterium enoyl ACP reductase (InhA). J. Mol. Graph. Model. 2017, 77, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Shahinozzaman, M.; Taira, N.; Ishii, T.; Halim, M.; Hossain, M.; Tawata, S. Anti-Inflammatory, Anti-Diabetic, and Anti-Alzheimer’s Effects of Prenylated Flavonoids from Okinawa Propolis: An Investigation by Experimental and Computational Studies. Molecules 2018, 23, 2479. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R.G. Absolute electronegativity and hardness correlated with molecular orbital theory. Proc. Natl. Acad. Sci. USA 1986, 83, 8440–8441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucido, M.J.; Orlando, B.J.; Vecchio, A.J.; Malkowski, M.G. Crystal Structure of Aspirin-Acetylated Human Cyclooxygenase-2: Insight into the Formation of Products with Reversed Stereochemistry. Biochemistry 2016, 55, 1226–1238. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.N. | R.T. (min) | Area (%) | PubChem CID | Chemical Formula | Bioactives Class | Bioactives Name |
|---|---|---|---|---|---|---|
| 1 | 4.38 | 0.43 | 54544338 | C5H6N2O2 | Organooxygen compounds | N-Cyano-3-oxobutanamide |
| 2 | 4.43 | 0.8 | 580975 | C7H10O2 | Organooxygen compounds | 1,3-Cyclopentanedione, 2,4-dimethyl- or 2,4-Dimethyl-1,3-cyclopentanedione |
| 3 | 4.83 | 1.76 | 119838 | C6H8O4 | Pyrans | 2,3-Dihydro-3,5-dihydroxy-6-methyl-4h-pyran-4-one or 3-Hydroxy-2,3-dihydromaltol |
| 4 | 5.45 | 4.58 | 12991 | C4H6O | Organooxygen compounds | 2-Butyn-1-ol or, 2-Butynol |
| 5 | 5.68 | 0.74 | 643131 | C4H5ClO | Acyl halides | (2E)-2-Butenoyl chloride or 2-Butenoyl chloride |
| 6 | 5.75 | 0.82 | 123411 | C4H3NS | Isothiocyanates | Thiocyanic acid, 2-propynyl ester or Propargyl isothiocyanate |
| 7 | 5.99 | 0.99 | 76665 | C6H10N2O | Azolines | 3H-Pyrazol-3-one, 2,4-dihydro-4,4,5-trimethyl- or 3,4-Trimethyl-5-pyrazolone |
| 8 | 6.08 | 0.68 | 5362763 | C8H14O | Organooxygen compounds | 2-Heptenal, 2-methyl- or 2-methyl-2-heptenal |
| 9 | 6.2 | 1.96 | 8102 | C6H15N | Organooxygen compounds | 1-Hexanamine or Hexylamine |
| 10 | 6.49 | 2.41 | 538272 | C8H15BO3 | Boronic acid derivatives | Lactic acid, 2-methyl-, monoanhydride with 1-butaneboronic acid, cyclic ester or alpha-Hydroxyisobutyric acid cyclic butaneboronate |
| 11 | 6.87 | 2.88 | 11850 | C6H14O6 | Organooxygen compounds | Galactitol |
| 12 | 7.08 | 7.14 | 5951 | C3H7NO3 | Carboxylic acids and derivatives | L-Serine or Serine |
| 13 | 7.31 | 0.8 | 5366263 | C19H38O | Organooxygen compounds | Ether, methyl 1-octadecenyl or 1-Methoxy-1-octadecene |
| 14 | 7.35 | 0.41 | 11005 | C14H28O2 | Fatty acyls | Tetradecanoic acid or Myristic acid |
| 15 | 7.64 | 24.3 | 5780 | C6H14O6 | Organooxygen compounds | Sorbitol |
| 16 | 8.74 | 0.41 | 554151 | C17H34O2 | Fatty acyls | Pentadecanoic acid, 13-methyl-, methyl ester or 13-Methylpentadecanoic acid methyl ester |
| 17 | 8.9 | 1.21 | 985 | C16H32O2 | Fatty acyls | Hexadecanoic acid or Palmitic acid |
| 18 | 9.39 | 1.27 | 5284421 | C19H34O2 | Fatty acyls | 9,12-Octadecadienoic acid (Z,Z)-, methyl ester or Methyl linoleate |
| 19 | 9.57 | 1.33 | 5280450 | C18H32O2 | Fatty acyls | 9,12-Octadecadienoic acid (Z,Z)- or, Linoleic acid |
| 20 | 10.2 | 1.12 | 610075 | C10H6ClNO2 | Naphthalenes | Naphthalene, 6-chloro-1-nitro- or 6-Chloro-1-nitronaphthalene |
| 21 | 10.3 | 1.07 | 581332 | C15H23Cl2N3 | Benzene and substituted derivatives | Hexahydropyrazin-1-propylamine, 4-[2-[3,4-dichlorophenyl]ethyl]- |
| 22 | 11 | 3.29 | 5789 | C10H14N2O5 | Pyrimidine nucleosides | Thymidine |
| 23 | 12.9 | 4.96 | 10850 | C27H50O6 | Glycerolipids | Glycerol tricaprylate or Tricaprylin |
| 24 | 14.3 | 6.43 | 159892 | C4H5N3O2 | Organonitrogen compounds | 2-Hydroxy-4-hydroxylaminopyrimidine or, N4-hydroxycytosine |
| 25 | 15.6 | 5.75 | 6432563 | C28H44O | Steroids and steroid derivatives | Ergosta-5,7,22-trien-3-ol, (3.beta.,22E)- or (3.beta.)-Ergosta-5,7,22- trien-3-ol |
| 26 | 16.3 | 4.24 | 532200 | C17H12ClNO | Naphthalenes | 1-Naphthalenecarboxamide, N-(3-chlorophenyl)- or N-(3-chlorophenyl)naphthylcarboxamide |
| 27 | 16.6 | 1.8 | 312796 | C28H48O | Steroids and steroid derivatives | .alpha.-Ergostenol |
| MCC | MNC | Degree | ||||||
|---|---|---|---|---|---|---|---|---|
| Rank | Genes | Score | Rank | Genes | Score | Rank | Genes | Degree |
| 1 | TLR4 | 1298 | 1 | TLR4 | 18 | 1 | TLR4 | 18 |
| 2 | EGFR | 1232 | 1 | EGFR | 18 | 2 | EGFR | 18 |
| 3 | NOS3 | 1180 | 3 | NOS3 | 16 | 3 | NOS3 | 16 |
| 4 | PTPRC | 1158 | 3 | PTPRC | 16 | 4 | PTPRC | 16 |
| 5 | MAPK14 | 1085 | 5 | F2 | 15 | 5 | MAPK14 | 15 |
| 6 | PPARG | 969 | 6 | MAPK14 | 14 | 6 | F2 | 15 |
| 7 | FGF2 | 920 | 7 | TRPV1 | 13 | 7 | TRPV1 | 13 |
| 8 | JAK2 | 217 | 8 | FGF2 | 12 | 8 | FGF2 | 12 |
| 9 | NOS2 | 148 | 9 | ALOX5 | 11 | 9 | ALOX5 | 11 |
| 10 | CNR1 | 146 | 10 | CNR1 | 10 | 10 | PPARG | 11 |
| 11 | SIRT1 | 142 | 10 | PPARG | 10 | 11 | CNR1 | 10 |
| 12 | CNR2 | 134 | 10 | PTGER4 | 10 | 12 | PTGER4 | 10 |
| 13 | ADORA1 | 132 | 10 | SIRT1 | 10 | 13 | SIRT1 | 10 |
| 14 | PTGER3 | 130 | 14 | PTGER3 | 9 | 14 | PTGER3 | 9 |
| 15 | PTGDR2 | 122 | 14 | PTGES | 9 | 15 | PTGES | 9 |
| 16 | NPY5R | 121 | 16 | CNR2 | 8 | 16 | JAK2 | 9 |
| 17 | LCK | 96 | 16 | NOS2 | 8 | 17 | CNR2 | 8 |
| 18 | PTGER4 | 84 | 16 | JAK2 | 8 | 18 | PLA2G4A | 8 |
| 19 | TRPV1 | 82 | 16 | PLA2G4A | 8 | 19 | LTB4R | 8 |
| 20 | F2 | 72 | 20 | ADORA1 | 7 | 20 | F2R | 8 |
| 21 | PDGFRB | 54 | 20 | HRH1 | 7 | 21 | NOS2 | 8 |
| 21 | ALOX5 | 54 | 20 | LCK | 7 | 22 | ADORA1 | 7 |
| 23 | PTGES | 38 | 20 | TRPA1 | 7 | 23 | PTGDR2 | 7 |
| 24 | FLT1 | 36 | 20 | F2R | 7 | 24 | LCK | 7 |
| 25 | F2R | 33 | 25 | PDGFRB | 6 | 25 | TRPA1 | 7 |
| 26 | PLA2G4A | 32 | 25 | FLT1 | 6 | 26 | PPARA | 7 |
| 26 | HRH1 | 32 | 25 | PLAU | 6 | 27 | HRH1 | 7 |
| 26 | CD38 | 32 | 25 | CD38 | 6 | 28 | NPY5R | 6 |
| 29 | LTB4R | 30 | 29 | PTGDR2 | 5 | 29 | CD38 | 6 |
| 30 | PTGFR | 26 | 29 | NPY5R | 5 | 30 | PDGFRB | 6 |
| 31 | PLAU | 24 | 29 | PARP1 | 5 | 31 | FLT1 | 6 |
| 32 | TRPA1 | 16 | 29 | PLAT | 5 | 32 | PLAU | 6 |
| 33 | PLAT | 14 | 29 | PTGFR | 5 | 33 | NR1H4 | 5 |
| 34 | PLA2G2A | 12 | 29 | PPARA | 5 | 34 | PARP1 | 5 |
| 34 | PPARA | 12 | 35 | NR1H4 | 4 | 35 | PLAT | 5 |
| 36 | PARP1 | 10 | 35 | LTB4R | 4 | 36 | PTGFR | 5 |
| 37 | NR1H4 | 7 | 35 | PLA2G2A | 4 | 37 | CYP2C19 | 4 |
| 37 | CYP2C19 | 7 | 38 | CYP2C19 | 3 | 38 | PLA2G2A | 4 |
| 39 | KCNK3 | 6 | 38 | KCNK3 | 3 | 39 | FABP1 | 4 |
| 39 | KCNK9 | 6 | 38 | KCNK9 | 3 | 40 | KCNK3 | 3 |
| 41 | FABP1 | 5 | 38 | FABP1 | 3 | 41 | KCNK9 | 3 |
| 42 | GBA | 2 | 42 | GBA | 2 | 42 | GBA | 2 |
| Gene | PDB ID | Bioactives | Docking Score Kcal/mol |
|---|---|---|---|
| TLR4 | 3UL7 | N-(3-chlorophenyl)naphthylcarboxamide | −4.027 |
| Aspirin * | −4.178 | ||
| Indomethacin * | −4.168 | ||
| 2-acetamido−2-deoxy-beta-d-glucopyranose | −4.797 | ||
| EGFR | 5WB7 | N-(3-chlorophenyl)naphthylcarboxamide | −4.123 |
| Aspirin * | −4.182 | ||
| Indomethacin * | −4.707 | ||
| 2-acetamido-2-deoxy-beta-d-glucopyranose | −4.205 | ||
| FLT1 | 3HNG | N-(3-chlorophenyl)naphthylcarboxamide | −8.88 |
| Aspirin * | −6.794 | ||
| Indomethacin * | −5.538 | ||
| N-(4-chlorophenyl)-2-[(pyridin-4-ylmethyl)amino]benzamide | −11.044 | ||
| NOS3 | 1M9J | N-(3-chlorophenyl)naphthylcarboxamide | −7.236 |
| Aspirin * | −5.262 | ||
| Indomethacin * | −7.451 | ||
| Chlorzoxazone | −6.118 | ||
| NOS2 | 1NSI | N-(3-chlorophenyl)naphthylcarboxamide | −8.85 |
| Aspirin * | −6.467 | ||
| Indomethacin * | −7.491 | ||
| Protoporphyrin Ix Containing Fe | −14.755 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, J.; Chowdhury, M.H.U.; Hafizur Rahman, M.; Choi, K.-Y.; Adnan, M. Bioactive Compounds and Signaling Pathways of Wolfiporia extensa in Suppressing Inflammatory Response by Network Pharmacology. Life 2023, 13, 893. https://doi.org/10.3390/life13040893
Jin J, Chowdhury MHU, Hafizur Rahman M, Choi K-Y, Adnan M. Bioactive Compounds and Signaling Pathways of Wolfiporia extensa in Suppressing Inflammatory Response by Network Pharmacology. Life. 2023; 13(4):893. https://doi.org/10.3390/life13040893
Chicago/Turabian StyleJin, Juri, Md. Helal Uddin Chowdhury, Md. Hafizur Rahman, Ki-Young Choi, and Md. Adnan. 2023. "Bioactive Compounds and Signaling Pathways of Wolfiporia extensa in Suppressing Inflammatory Response by Network Pharmacology" Life 13, no. 4: 893. https://doi.org/10.3390/life13040893