



Food Toxicity of Mycotoxin Citrinin and Molecular Mechanisms of Its Potential Toxicity Effects through the Implicated Targets Predicted by Computer-Aided Multidimensional Data Analysis

Abstract
:1. Introduction
2. Material and Methods
2.1. Fungal Culture and Analysis of Citrinin Production
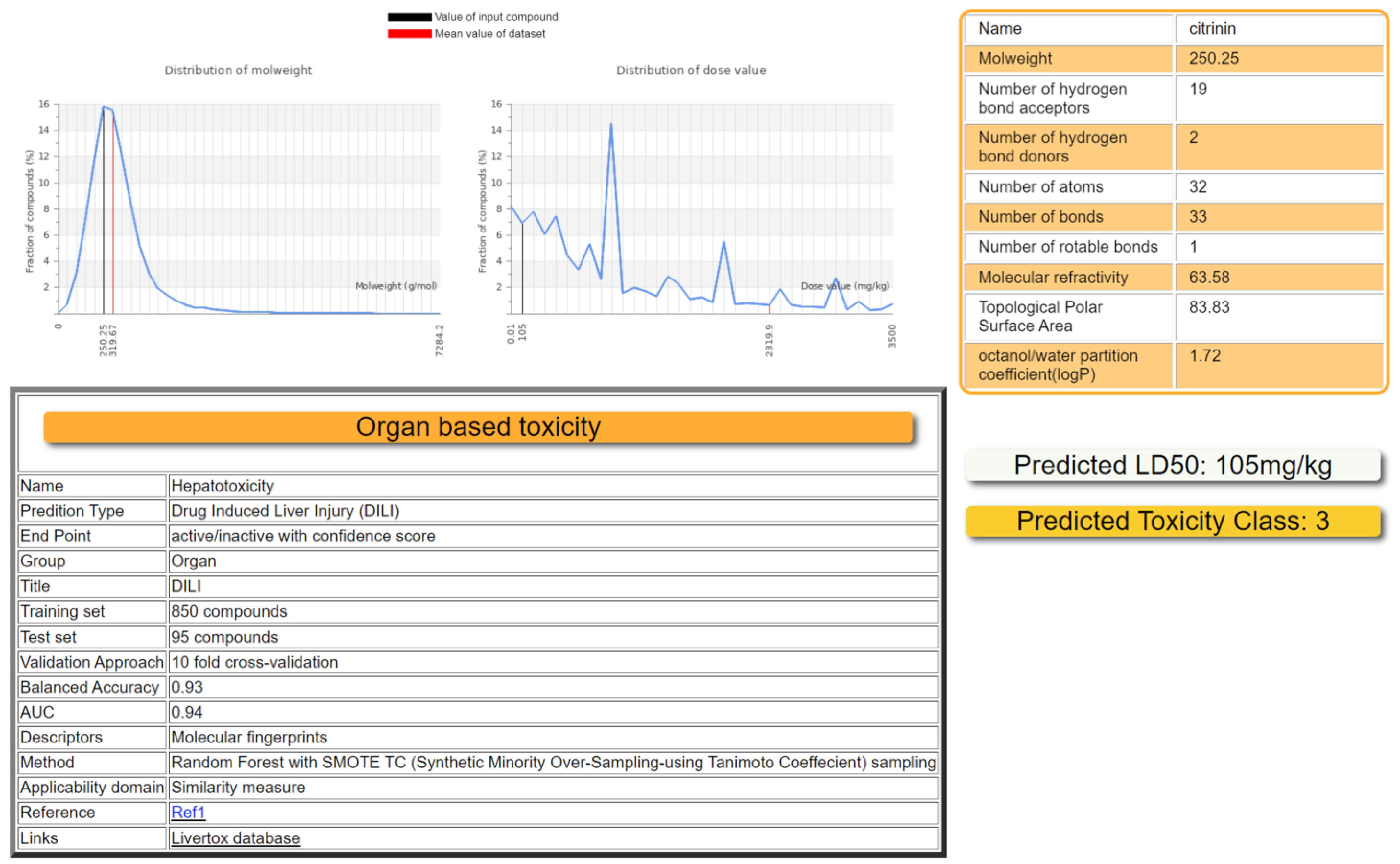
2.2. Toxicity Prediction by ProTox-II
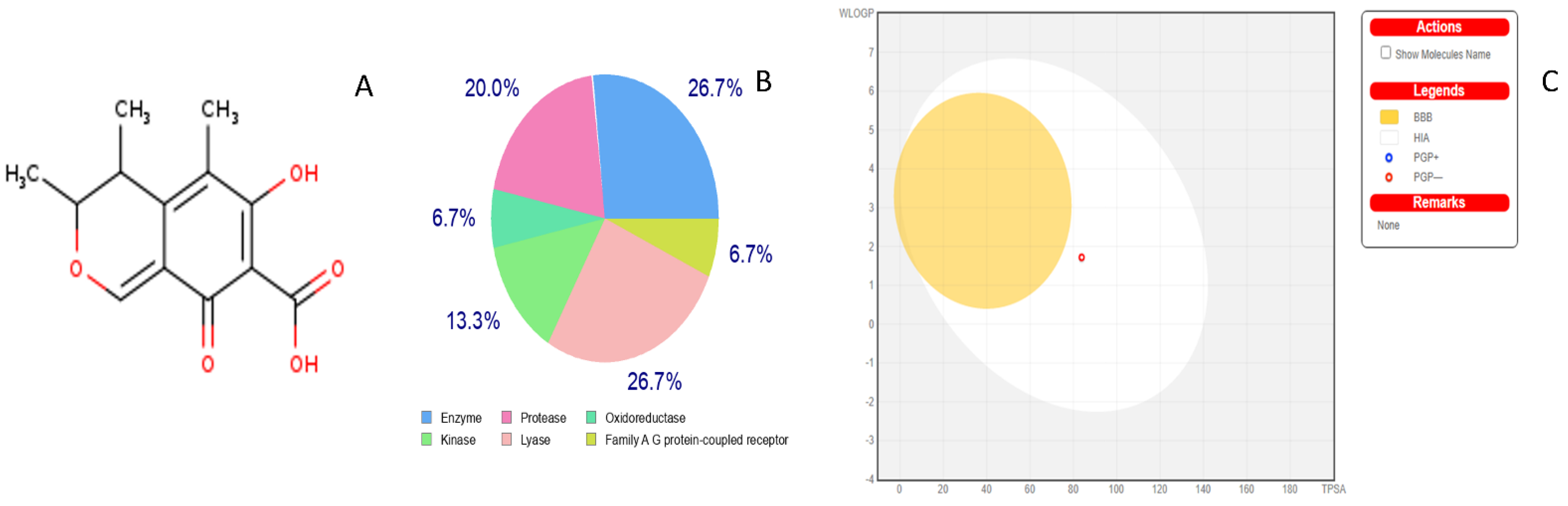
2.3. Toxicity Radar Targets of Citrinin
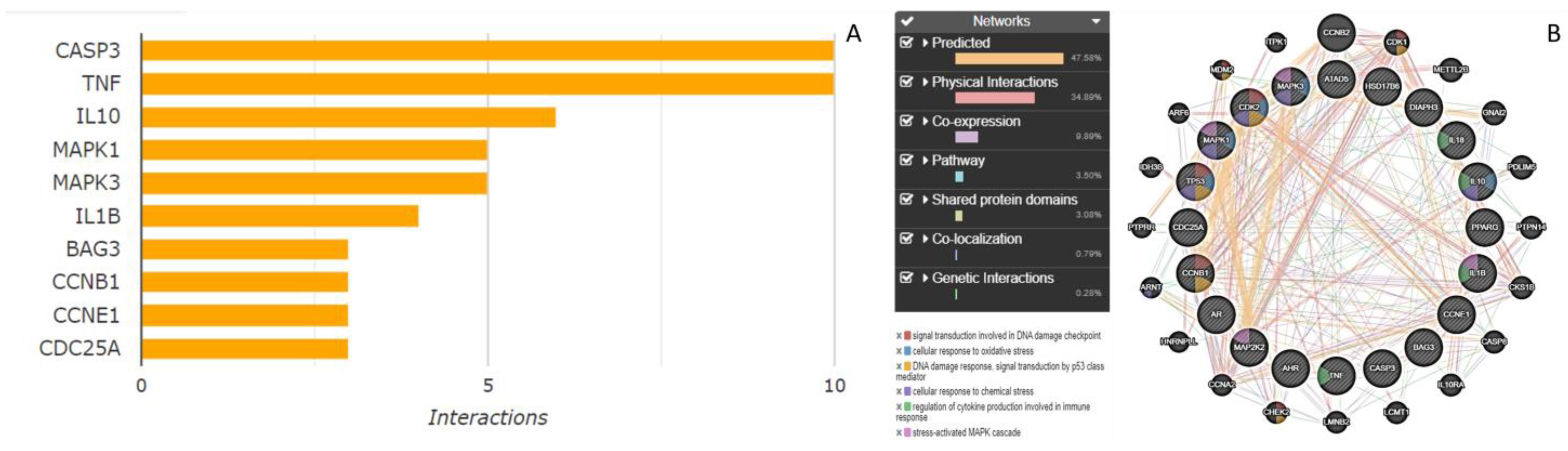
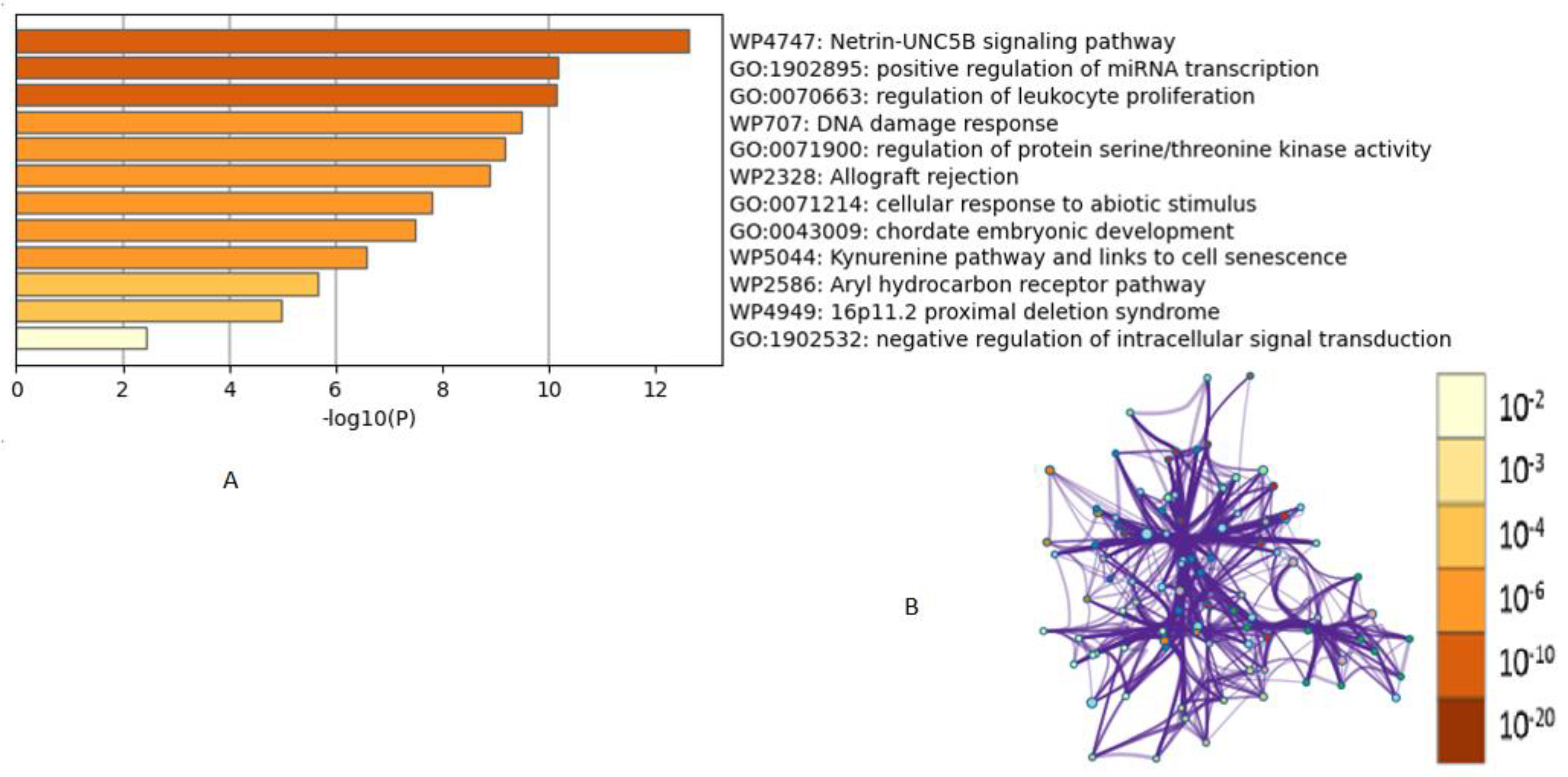
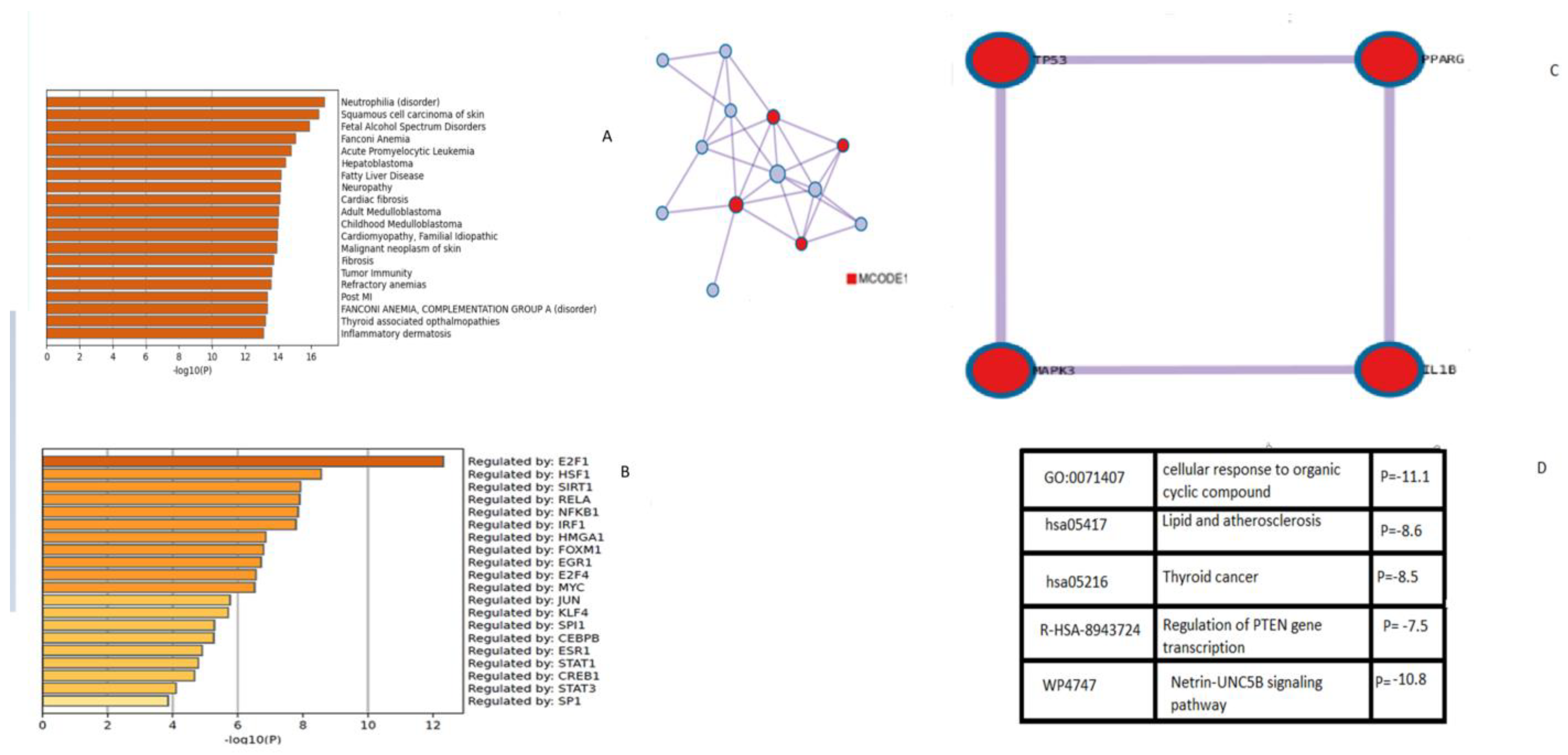
2.4. Gene Interaction Networks and Pathway Enrichment Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zargar, S. Effect of Aqueous Extract of Rhazyastricta Decne on Citrinin Production and Fungal Biomass by Pencillium notatum and Optimization of Experimental Design Using Response Surface Methodology. Biosci. Biotechnol. Res. Asia 2016, 13, 887–893. [Google Scholar] [CrossRef] [Green Version]
- Dalefield, R. Mycotoxins and mushrooms. In Veterinary Toxicology for Australia and New Zealand; Elsevier: Amsterdam, The Netherlands, 2017; pp. 373–419. [Google Scholar]
- Scott, P.M. Other mycotoxins. In Mycotoxins in Food: Detection and Control; Magan, N., Olsen, M., Eds.; Woodhead Publishing Ltd.: Cambridge, UK; CRC Press LLC.: Cambridge, UK, 2004; pp. 1–471. [Google Scholar]
- Silva, L.J.G.; Pereira, A.M.P.T.; Pena, A.; Lino, C.M. Citrinin in Foods and Supplements: A Review of Occurrence and Analytical Methodologies. Foods 2020, 10, 14. [Google Scholar] [CrossRef]
- Ojuri, O.T.; Ezekiel, C.N.; Sulyok, M.; Ezeokoli, O.T.; Oyedele, O.A.; Ayeni, K.I.; Eskola, M.K.; Šarkanj, B.; Hajslova, J.; Adeleke, R.; et al. Assessing the mycotoxicological risk from consumption of complementary foods by infants and young children in Nigeria. Food Chem. Toxicol. 2018, 121, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Sáncheza, P.L.; De Nijsa, M.; Spanjerb, M.; Pietric, A.; Bertuzzic, T.; Starski, A.; Postupolski, J.; Castellari, M.; Hortós, M. Generation of occurrence data on citrinin in food. EFSA Support. Publ. 2017, 14, 1177E. [Google Scholar] [CrossRef] [Green Version]
- Ouhibi, S.; Vidal, A.; Martins, C.; Gali, R.; Hedhili, A.; De Saeger, S.; De Boevre, M. LC-MS/MS methodology for simultaneous determination of patulin and citrinin in urine and plasma applied to a pilot study in colorectal cancer patients. Food Chem. Toxicol. 2020, 136, 110994. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Blaszkewicz, M.; Mohanto, N.C.; Rahman, M.; Alim, A.; Hossain, K.; Degen, G.H. First results on citrinin biomarkers in urines from rural and urban cohorts in Bangladesh. Mycotoxin Res. 2015, 31, 9–16. [Google Scholar] [CrossRef]
- Liu, B.-H.; Wu, T.-S.; Su, M.-C.; Chung, C.P.; Yu, F.-Y. Evaluation of Citrinin Occurrence and Cytotoxicity in Monascus Fermentation Products. J. Agric. Food Chem. 2005, 53, 170–175. [Google Scholar] [CrossRef]
- Flajs, D.; Peraica, M. Toxicological Properties of Citrinin. Arch. Ind. Hyg. Toxicol. 2009, 60, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Sabater-Vilar, M.; Maas, R.F.M.; Fink-Gremmels, J. Mutagenicity of commercial Monascus fermentation products and the role of citrinin contamination. Mutat. Res. Toxicol. Environ. Mutagen. 1999, 444, 7–16. [Google Scholar] [CrossRef]
- Adam, M.A.A.; Tabana, Y.M.; Musa, K.B.; Sandai, D.A. Effects of different mycotoxins on humans, cell genome and their involvement in cancer. Oncol. Rep. 2017, 37, 1321–1336. [Google Scholar] [CrossRef] [Green Version]
- Mally, A.; Dekant, W. DNA adduct formation by ochratoxin A: Review of the available evidence. Food Addit. Contam. 2005, 22, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, S.M.; Chagas, G.M.; Campello, A.P.; Klüppel, M.L. Mechanism of citrinin-induced dysfunction of mitochondria. V. Effect on the homeostasis of the reactive oxygen species. Cell Biochem. Funct. 1997, 15, 203–209. [Google Scholar] [CrossRef]
- Pascual-Ahuir, A.; Vanacloig-Pedros, E.; Proft, M. Toxicity Mechanisms of the Food Contaminant Citrinin: Application of a Quantitative Yeast Model. Nutrients 2014, 6, 2077–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degen, G.H.; Ali, N.; Gundert-Remy, U. Preliminary data on citrinin kinetics in humans and their use to estimate citrinin exposure based on biomarkers. Toxicol. Lett. 2018, 282, 43–48. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Filho, J.W.G.; Islam, M.T.; Ali, E.S.; Uddin, S.J.; de Oliveira Santos, J.V.; de Alencar, M.V.O.B.; Júnior, A.L.G.; Paz, M.F.C.J.; de Brito, M.D.R.M.; de Castro e Sousa, J.M.; et al. A compre-hensive review on biological properties of citrinin. Food Chem. Toxicol. 2017, 110, 130–141. [Google Scholar] [CrossRef]
- Li, M.; He, M.; Xu, F.; Guan, Y.; Tian, J.; Wan, Z.; Zhou, H.; Gao, M.; Chong, T. Abnormal expression and the significant prognostic value of aquaporins in clear cell renal cell carcinoma. PLoS ONE 2022, 17, e0264553. [Google Scholar] [CrossRef]
- Hackbart, H.; Prietto, L.; Primel, E.G.; Garda-Buffon, J.; Badiale-Furlong, E. Simultaneous extraction and detection of ochra-toxin A and citrinin in rice. J. Braz. Chem. Soc. 2012, 23, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-P.; Xu, Y.; Huang, Z.-B. Isolation and characterization of the citrinin biosynthetic gene cluster from Monascus aurantiacus. Biotechnol. Lett. 2012, 34, 131–136. [Google Scholar] [CrossRef]
- Shimizu, T.; Kinoshita, H.; Ishihara, S.; Sakai, K.; Nagai, S.; Nihira, T. Polyketide synthase gene responsible for citrinin bio-synthesis in Monascus purpureus. Appl. Environ. Microbiol. 2005, 71, 3453–3457. [Google Scholar] [CrossRef] [Green Version]
- Chai, X.; Ai, Z.; Liu, J.; Guo, T.; Wu, J.; Bai, J.; Lin, Q. Effects of pigment and citrinin biosynthesis on the metabolism and morphology of Monascus purpureus in submerged fermentation. Food Sci. Biotechnol. 2020, 29, 927–937. [Google Scholar] [CrossRef]
- Lurá, M.C.; Fuentes, M.B.; Cabagna, M.; González, A.M.; Nepote, A.F.; Giugni, M.C.; Rico, M.; Latorre, M.G. Actividad de metabolitos de Penicillium citrinum sobre ratones Mus musculus. Rev. Iberoam. Micol. 2001, 18, 183–186. [Google Scholar]
- Chan, W.-H. Effects of citrinin on maturation of mouse oocytes, fertilization, and fetal development in vitro and in vivo. Toxicol. Lett. 2008, 180, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.; O Eckert, A.; Schrey, A.K.; Preissner, R. ProTox-II: A webserver for the prediction of toxicity of chemicals. Nucleic Acids Res. 2018, 46, W257–W263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, C.-Y.; Li, J.-Y.; Hao, G.-F.; Yang, G.-F. A drug-likeness toolbox facilitates ADMET study in drug discovery. Drug Discov. Today 2020, 25, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Zoete, V. Application of the SwissDrugDesign Online Resources in Virtual Screening. Int. J. Mol. Sci. 2019, 20, 4612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. iLOGP: A Simple, Robust, and Efficient Description of n-Octanol/Water Partition Coeffi-cient for Drug Design Using the GB/SA Approach. J. Chem. Inf. Model. 2014, 54, 3284–3301. [Google Scholar] [CrossRef]
- Davis, A.P.; Wiegers, T.C.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Mattingly, C.J. Comparative Toxicogenomics Database (CTD): Update 2023. Nucleic Acids Res. 2023, 51, D1257–D1262. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Jo, W.H.; Hoang, N.H.M.; Kim, M.-S. Curcumin-attenuated TREM-1/DAP12/NLRP3/Caspase-1/IL1B, TLR4/NF-κB pathways, and tau hyperphosphorylation induced by 1, 2-diacetyl benzene: An in vitro and in silico study. Neurotox. Res. 2022, 40, 1272–1291. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.D.; Oh, H.; Kim, M.-S. The effects of chemical mixtures on lipid profiles in the Korean adult population: Threshold and molecular mechanisms for dyslipidemia involved. Environ. Sci. Pollut. Res. 2022, 29, 39182–39208. [Google Scholar] [CrossRef]
- Groestlinger, J.; Spindler, V.; Pahlke, G.; Rychlik, M.; Del Favero, G.; Marko, D. Alternaria alternata Mycotoxins Activate the Aryl Hydrocarbon Receptor and Nrf2-ARE Pathway to Alter the Structure and Immune Response of Colon Epithelial Cells. Chem. Res. Toxicol. 2022, 35, 731–749. [Google Scholar] [CrossRef] [PubMed]
- Miki, Y.; Hata, S.; Ono, K.; Suzuki, T.; Ito, K.; Kumamoto, H.; Sasano, H. Roles of Aryl Hydrocarbon Receptor in Aromatase-Dependent Cell Proliferation in Human Osteoblasts. Int. J. Mol. Sci. 2017, 18, 2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrou, A.; Geronikaki, A.; Kartsev, V.; Kousaxidis, A.; Papadimitriou-Tsantarliotou, A.; Kostic, M.; Ivanov, M.; Sokovic, M.; Nicolaou, I.; Vizirianakis, I.S. N-Derivatives of (Z)-Methyl 3-(4-Oxo-2-thioxothiazolidin-5-ylidene)methyl)-1H-indole-2-carboxylates as Antimicrobial Agents—In Silico and In Vitro Evaluation. Pharmaceuticals 2023, 16, 131. [Google Scholar] [CrossRef]
- Kamle, M.; Mahato, D.K.; Gupta, A.; Pandhi, S.; Sharma, N.; Sharma, B.; Mishra, S.; Arora, S.; Selvakumar, R.; Saurabh, V.; et al. Citrinin Mycotoxin Contamination in Food and Feed: Impact on Agriculture, Human Health, and Detection and Management Strategies. Toxins 2022, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- Montanari, F.; Ecker, G.F. Prediction of drug-ABC-transporter interaction–Recent advances and future challenges. Adv. Drug Deliv. Rev. 2015, 86, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Zargar, S.; Wani, T.A.; Ahamad, S.R. An Insight into Wheat Germ Oil Nutrition, Identification of Its Bioactive Constituents and Computer-Aided Multidimensional Data Analysis of Its Potential Anti-Inflammatory Effect via Molecular Connections. Life 2023, 13, 526. [Google Scholar] [CrossRef]
- Behrens, M.; Hüwel, S.; Galla, H.-J.; Humpf, H.-U. Efflux at the Blood-Brain Barrier Reduces the Cerebral Exposure to Ochratoxin A, Ochratoxin α, Citrinin and Dihydrocitrinone. Toxins 2021, 13, 327. [Google Scholar] [CrossRef]
- He, J.; Ji, X.; Li, Y.; Xue, X.; Feng, G.; Zhang, H.; Wang, H.; Gao, M. Subchronic exposure of benzo(a)pyrene interferes with the expression of Bcl-2, Ki-67, C-myc and p53, Bax, Caspase-3 in sub-regions of cerebral cortex and hippocampus. Exp. Toxicol. Pathol. 2016, 68, 149–156. [Google Scholar] [CrossRef]
- Yu, F.-Y.; Liao, Y.-C.; Chang, C.-H.; Liu, B.-H. Citrinin induces apoptosis in HL-60 cells via activation of the mitochondrial pathway. Toxicol. Lett. 2006, 161, 143–151. [Google Scholar] [CrossRef]
- Johannessen, L.; Nilsen, A.; Løvik, M. The mycotoxins citrinin and gliotoxin differentially affect production of the pro-inflammatory cytokines tumour necrosis factor-α and interleukin-6, and the anti-inflammatory cytokine interleukin-10. Clin. Exp. Allergy 2005, 35, 782–789. [Google Scholar] [CrossRef]
- Adnan, M.; Siddiqui, A.J.; Noumi, E.; Hannachi, S.; Ashraf, S.A.; Awadelkareem, A.M.; Snoussi, M.; Badraoui, R.; Bardakci, F.; Sachidanandan, M.; et al. Integrating Network Pharmacology Approaches to Decipher the Multi-Target Pharmacological Mechanism of Microbial Biosurfactants as Novel Green Antimicrobials against Listeriosis. Antibiotics 2022, 12, 5. [Google Scholar] [CrossRef]
- Li, Y.; Arita, Y.; Koo, H.-C.; Davis, J.M.; Kazzaz, J.A. Inhibition of c-Jun N-Terminal Kinase Pathway Improves Cell Viability in Response to Oxidant Injury. Am. J. Respir. Cell Mol. Biol. 2003, 29, 779–783. [Google Scholar] [CrossRef]
- Pradella, D.; Deflorian, G.; Pezzotta, A.; Di Matteo, A.; Belloni, E.; Campolungo, D.; Paradisi, A.; Bugatti, M.; Vermi, W.; Campioni, M.; et al. A ligand-insensitive UNC5B splicing isoform regulates angiogenesis by promoting apoptosis. Nat. Commun. 2021, 12, 4872. [Google Scholar] [CrossRef] [PubMed]
- Stambolic, V.; MacPherson, D.; Sas, D.; Lin, Y.; Snow, B.; Jang, Y.; Benchimol, S.; Mak, T. Regulation of PTEN Transcription by p53. Mol. Cell 2001, 8, 317–325. [Google Scholar] [CrossRef]
- Tibbles, L.A.; Woodgett, J.R. The stress-activated protein kinase pathways. Cell. Mol. Life Sci. 1999, 55, 1230–1254. [Google Scholar] [CrossRef]
- Zargar, S.; Wani, T.A. Protective Role of Quercetin in Carbon Tetrachloride Induced Toxicity in Rat Brain: Biochemical, Spectrophotometric Assays and Computational Approach. Molecules 2021, 26, 7526. [Google Scholar] [CrossRef]
- Wang, S.; Zhen, L.; Li, X.; Fu, X.; Li, P.; Zhang, D. Search for Key Genes and Functional Pathways of Ulcerative Colitis to Colon Cancer Based on Bioinformatics. Front. Oncol. 2022, 12, 857148. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Zhao, Z.; Wei, L.; Zhou, D.; Xue, Z.; Ge, S. HSF1/HSP pathway in the hippocampus is involved in SIRT1-mediated caloric restriction-induced neuroprotection after surgery in aged mice. Exp. Gerontol. 2019, 119, 184–192. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Target | Probability |
|---|---|---|
| Organ toxicity | Hepatotoxicity | 0.7 |
| Toxicity endpoints | Carcinogenicity | 0.56 |
| Toxicity endpoints | Immunogenicity | 0.69 |
| Toxicity endpoints | Mutagenicity | 0.85 |
| Toxicity endpoints | Cytotoxicity | 0.77 |
| Tox21-nuclear receptor signaling pathways | Aryl hydrocarbon receptor (AhR) | 0.85 |
| Tox21-nuclear receptor signaling pathways | Androgen receptor (AR) | 0.93 |
| Tox21-nuclear receptor signaling pathways | Androgen receptor ligand-binding domain (AR-LBD) | 0.96 |
| Tox21-nuclear receptor signaling pathways | Aromatase | 0.75 |
| Tox21-nuclear receptor signaling pathways | Estrogen receptor alpha (ER) | 0.75 |
| Tox21-nuclear receptor signaling pathways | Estrogen receptor ligand-binding domain (ER-LBD) | 0.91 |
| Tox21-nuclear receptor signaling pathways | Peroxisome proliferator-activated receptor gamma (PPAR-Gamma) | 0.94 |
| Tox21-stress response pathways | Nuclear factor (erythroid-derived 2)-like 2/antioxidant response element (nrf2/ARE) | 0.85 |
| Tox21-stress response pathways | Heat shock factor response element (HSE) | 0.85 |
| Tox21-stress response pathways | Mitochondrial membrane potential (MMP) | 0.61 |
| Tox21-stress response pathways | Phosphoprotein (tumor suppressor) p53 | 0.83 |
| Tox21-stress response pathways | ATPase family AAA domain-containing protein 5 (ATAD5) | 0.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zargar, S.; Wani, T.A. Food Toxicity of Mycotoxin Citrinin and Molecular Mechanisms of Its Potential Toxicity Effects through the Implicated Targets Predicted by Computer-Aided Multidimensional Data Analysis. Life 2023, 13, 880. https://doi.org/10.3390/life13040880
Zargar S, Wani TA. Food Toxicity of Mycotoxin Citrinin and Molecular Mechanisms of Its Potential Toxicity Effects through the Implicated Targets Predicted by Computer-Aided Multidimensional Data Analysis. Life. 2023; 13(4):880. https://doi.org/10.3390/life13040880
Chicago/Turabian StyleZargar, Seema, and Tanveer A. Wani. 2023. "Food Toxicity of Mycotoxin Citrinin and Molecular Mechanisms of Its Potential Toxicity Effects through the Implicated Targets Predicted by Computer-Aided Multidimensional Data Analysis" Life 13, no. 4: 880. https://doi.org/10.3390/life13040880