

Antifouling Potential of Ethyl Acetate Extract of Marine Bacteria Pseudomonas aeruginosa Strain RLimb

 , , , ,
, , , ,  and
and
Abstract
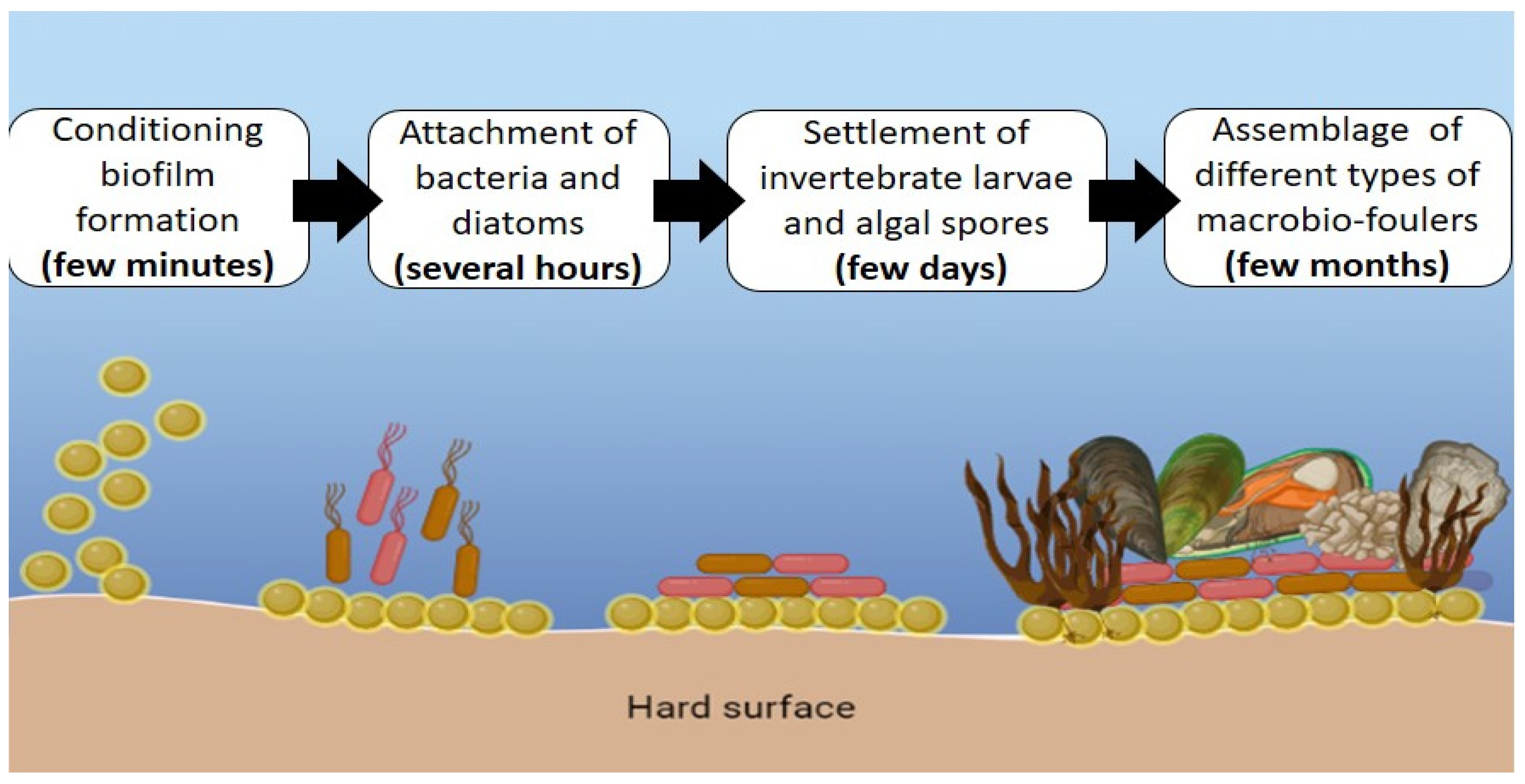
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Bacteria Isolate
2.2. Biosynthesis of Bioactive Compounds from Bacterial Isolate
2.3. Extraction of Bioactive Compounds
2.4. Antibacterial Assay
2.5. Antifouling Test
2.5.1. Crystal Violet Assay
2.5.2. Aquarium Test
2.6. Bacterial Isolation and Characterization
Morphological Characterization (Gram-Staining Test)
2.7. Biochemical Identification
2.7.1. Hydrolysis of Enzymes
2.7.2. Catalase Test
2.7.3. Oxidase Test
2.7.4. Susceptibility Test
2.7.5. Determination of Marine Isolate
2.8. Molecular Identification of Bacteria Isolate
2.9. Statistical Analysis
3. Results and Discussion
3.1. Antibacterial Assay
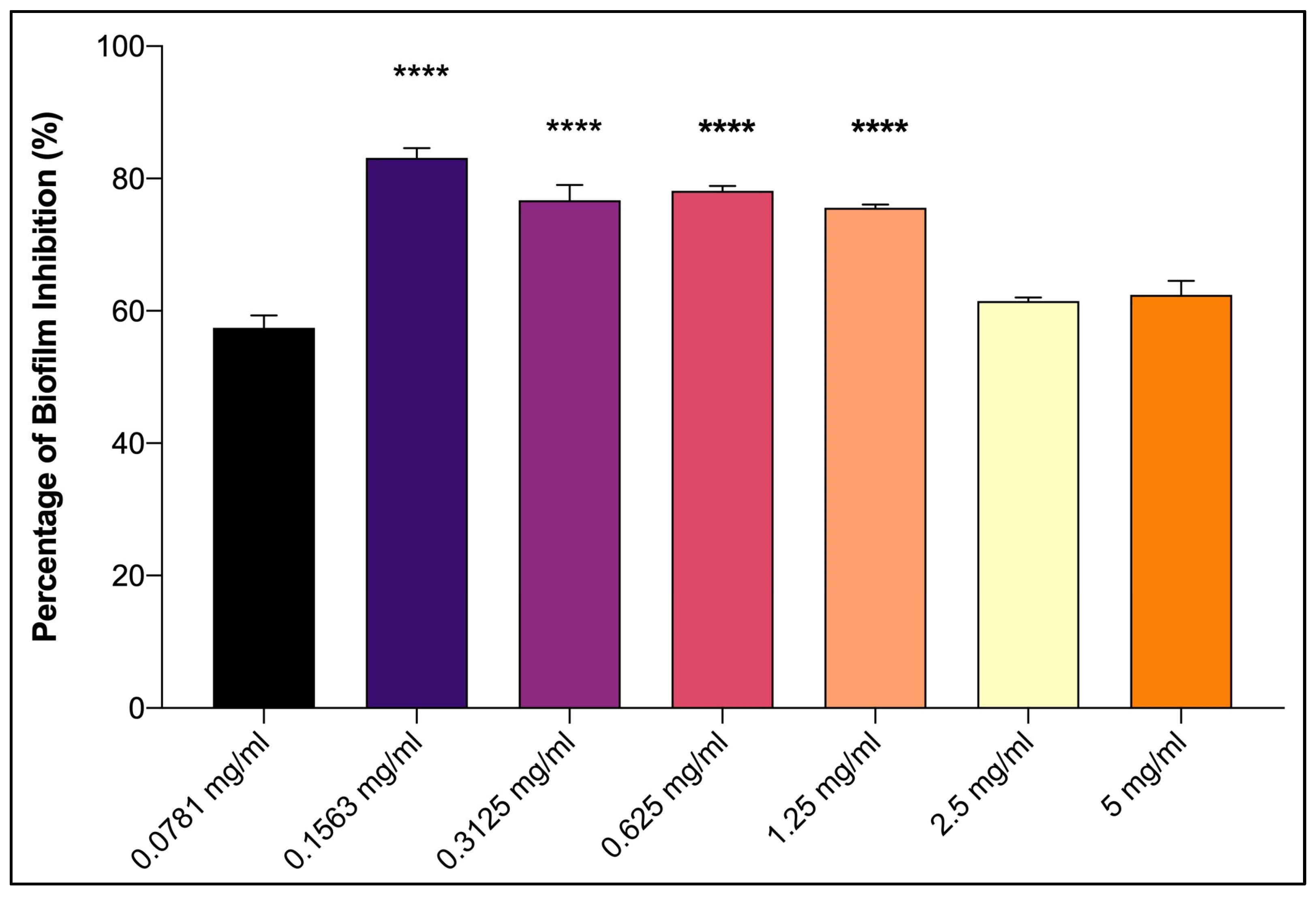
3.2. Antifouling Activity
3.3. Characterization of Bacteria Isolate
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heidarian, S.; Mohammadipanah, F.; Maghsoudlou, A.; Dashti, Y.; Challis, G.L. Anti-microfouling activity of Glycomyces Sediminimaris UTMC 2460 on dominant fouling bacteria of Iran marine habitats. Front. Microbiol. 2019, 10, 3148. [Google Scholar] [CrossRef] [PubMed]
- Soliman, Y.A.; Mohamed, A.S.; NaserGomaa, M. Antifouling activity of crude extracts isolated from two red sea puffer fishes. Egypt J. Aquat. Res. 2014, 40, 24. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Pan, J.; Ma, C.; Zhang, G. Dynamic surface antifouling: Mechanism and systems. Soft. Matter. 2019, 15, 1087–1107. [Google Scholar] [CrossRef]
- Bengen, D.G.; Khoeri, M.M.; Marhaeni, B.; Radjasa, O.K.; Sabdono, A.; Sudoyo, H. Antifouling activity of bacterial symbionts of seagrasses against marine biofilm-forming bacteria. J. Environ. Prot. 2011, 2, 1245–1249. [Google Scholar] [CrossRef] [Green Version]
- Rampadarath, S.; Bandhoa, K.; Puchooa, D.; Jeewon, R.; Bal, S. Early bacterial biofilm colonizers in the coastal waters of mauritius. Electron. J. Biotechnol. 2017, 29, 13–31. [Google Scholar] [CrossRef]
- Cho, J.Y.; Kang, J.Y.; Hong, Y.K.; Baek, H.H.; Shin, H.W.; Kim, M.S. Isolation and structural determination of the antifouling diketopiperazines from marine-derived streptomyces praecox 291-11. Biosci. Biotechnol. Biochem. 2012, 76, 1116–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Duan, Y.; Cui, M.; Huang, R.; Su, R.; Qi, W.; He, Z. Biomimetic surface coatings for marine antifouling: Natural antifoulants, synthetic polymers and surface microtopography. Sci. Total Environ. 2021, 766, 144469. [Google Scholar] [CrossRef]
- Ramasubburayan, R.; Titus, S.; Verma, P.K.; Immanuel, G.; Palavesam, A. Isolation, screening and optimization of culture conditions for enhanced antibacterial activity by a marine epibiotic bacterium bacillus flexus APGI against fouling bacterial strains. J. Pure Appl. Microbiol. 2014, 8, 2909–2920. [Google Scholar]
- Jorge, P.; Lourenço, A.; Pereira, M.O. New trends in peptide-based anti-biofilm strategies: A review of recent achievements and bioinformatic approaches. Biofouling 2012, 28, 1033–1061. [Google Scholar] [CrossRef] [Green Version]
- Thiyagarajan, S.; Bavya, M.; Jamal, A. Isolation of marine fungi sp. and its antifouling activity against marine bacteria. J. Environ. Biol. 2016, 37, 895–903. [Google Scholar]
- Pérez, M.; Sánchez, M.; García, M.; Patiño C., L.P.; Blustein, G.; Palermo, J.A. Antifouling activity of peracetylated cholic acid, a natural bile acid derivative. Steroids 2019, 149, 108414. [Google Scholar] [CrossRef] [PubMed]
- Suresh, M.; Iyapparaj, P.; Anantharaman, P. Antifouling activity of lipidic metabolites derived from Padina tetrastromatica. Appl. Biochem. Biotechnol. 2016, 179, 805–818. [Google Scholar] [CrossRef]
- Zulkifli, M.F.R.; Radzi, N.M.; Jusoh, S.M.; Saidin, J.; Nik, W.M.N.W. Potential of cabbage extract (Brassica oleracea) as anti-fouling agent in alkyd undercoat for mild steel in seawater. Malaysian J. Anal. Sci. 2019, 23. [Google Scholar] [CrossRef]
- Pinteus, S.; Lemos, M.F.L.; Alves, C.; Silva, J.; Pedrosa, R. The marine invasive seaweeds Asparagopsis armata and Sargassum muticum as targets for greener antifouling solutions. Sci. Total Environ. 2021, 750, 141372. [Google Scholar] [CrossRef]
- Park, M.S.; Kim, Y.D.; Kim, B.M.; Kim, Y.J.; Kim, J.K.; Rhee, J.S. Effects of antifouling biocides on molecular and biochemical defense system in the gill of the pacific oyster Crassostrea gigas. PLoS ONE 2016, 11, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Tian, L.; Bing, W.; Zhao, J.; Ren, L. Bioinspired marine antifouling coatings: Status, prospects, and future. Prog. Mater. Sci. 2022, 124, 152. [Google Scholar] [CrossRef]
- Pawlik, J.R. Antipredatory defensive roles of natural products from marine invertebrates. In Handbook of Marine Natural Products; Springer: Dordrecht, The Netherlands, 2012; pp. 677–710. [Google Scholar] [CrossRef]
- Wahl, M.; Jensen, P.R.; Fenical, W. Chemical control of bacterial epibiosis on ascidians. Mar. Ecol. Prog. Ser. 1994, 110, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Clare, A.S. Marine natural product antifoulants: Status and potential. Biofouling 1996, 9, 211–229. [Google Scholar] [CrossRef]
- Palanichamy, S.; Subramanian, G. Antifouling properties of marine bacteriocin incorporated epoxy based paint. Prog. Org. Coat. 2017, 103, 33–39. [Google Scholar] [CrossRef]
- Liu, L.L.; Wu, C.H.; Qian, P.Y. Marine natural products as antifouling molecules—A mini-review (2014–2020). Biofouling 2020, 36, 1210–1226. [Google Scholar] [CrossRef] [PubMed]
- Sayem, S.M.A.; Manzo, E.; Ciavatta, L.; Tramice, A.; Cordone, A.; Zanfardino, A.; de Felice, M.; Varcamonti, M. Anti-biofilm activity of an exopolysaccharide from a sponge-associated strain of Bacillus licheniformis. Microb. Cell. Fact. 2011, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Muras, A.; Larroze, S.; Mayer, C.; Teixeira, T.; Wengier, R.; Benayahu, Y.; Otero, A. Evaluation of the anti-fouling efficacy of Bacillus licheniformis extracts under environmental and natural conditions. Front. Mar. Sci. 2021, 8, 711108. [Google Scholar] [CrossRef]
- Viju, N.; Satheesh, S.; Punitha, S.M.J. Antibiofilm and antifouling activities of extracellular polymeric substances isolated from the bacteria associated with marine gastropod Turbo sp. Oceanol. Hydrobiol. Stud. 2016, 45, 11–19. [Google Scholar] [CrossRef]
- Azemi, M.A.F.M.; Rashid, N.F.M.; Saidin, J.; Effendy, A.W.M.; Bhubalan, K. Application of sweetwater as potential carbon source for rhamnolipid production by marine Pseudomonas aeruginosa UMTKB-5. Int. J. Biosci. Biochem. Bioinforma. 2016, 6, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Tamothran, A.M.; Sevakumaran, V.; Bhubalan, K. Production and toxicity evaluation of rhamnolipids produced by pseudomonas strains on L6 and HepG2 cells. Malaysian Appl. Biol. 2019, 48, 149–156. [Google Scholar]
- Mohd Yatim, A.F.; Syafiq, I.M.; Huong, K.H.; Abdullah Amirul, A.A.; Mohd Effendy, A.W.; Bhubalan, K. Bioconversion of novel and renewable agro-industry by-products into a biodegradable Poly(3-Hydroxybutyrate) by marine Bacillus megaterium UMTKB-1 strain. Biotechnologia 2017, 98, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Qiang, J.; Jia, Y.; Ye, J.; Peng, H.; Qin, H.; Zhang, N.; He, B. Characteristics of biosurfactant produced by Pseudomonas aeruginosa S6 isolated from oil-containing wastewater. Proc. Biochem. 2009, 44, 556–564. [Google Scholar] [CrossRef]
- Kanagasabhapathy, M.; Sasaki, H.; Haldar, S.; Yamasaki, S.; Nagata, S. Antibacterial activities of marine epibiotic bacteria isolated from brown algae of Japan. Ann. Microbiol. 2006, 56, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.S.; Ismail, N.; Eldeen, I.M.S. Disruption of biofilm formation by fractions from book gills and carapace of horseshoe crab Tachypleus gigas. Mar. Biol. Res. 2020, 16, 556–564. [Google Scholar] [CrossRef]
- Leroy, C.; Delbarre-Ladrat, C.; Ghillebaert, F.; Rochet, M.J.; Compère, C.; Combes, D. A marine bacterial adhesion microplate test using the DAPI fluorescent dye: A new method to screen antifouling agents. Lett. Appl. Microbiol. 2007, 44, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Salama, A.J.; Satheesh, S.; Balqadi, A.A. Antifouling activities of methanolic extracts of three macroalgal species from the Red Sea. J. Appl. Phycol. 2018, 30, 1943–1953. [Google Scholar] [CrossRef]
- Chen, L.; Xia, C.; Qian, P.Y. Optimization of antifouling coatings incorporating butenolide, a potent antifouling agent via field and laboratory tests. Prog. Org. Coat. 2017, 109, 22–29. [Google Scholar] [CrossRef]
- Viju, N.; Satheesh, S.; Punitha, S.M.J. Antifouling activities of antagonistic marine Bacterium pseudomonas putida associated with an octopus. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 1113–1124. [Google Scholar] [CrossRef]
- Ligo, A.; Diki Juliandi, M.; Agustien, A.; Djamaan, A. Isolation and characterization of polyethylene-degrading marine bacteria. IOSR J. Pharm. Biol. Sci. 2020, 15, 22–25. [Google Scholar]
- Tripathi, N.; Sapra, A. Gram Staining. Available online: https://www.ncbi.nlm.nih.gov/books/NBK562156/#!po=75.0000 (accessed on 8 December 2021).
- Cheng, T.H.; Ismail, N.; Kamaruding, N.; Saidin, J.; Danish-Daniel, M. Industrial enzymes-producing marine bacteria from marine resources. Biotechnol. Rep. 2020, 27, e00482. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, S.; Islam, S.; Ahmed, M.M.; Belal Hossain, M.; Hossain, M.A. Protease producing bacteria and activity in gut of tiger shrimp (Penaeus monodon). J. Fish Aquat. Sci. 2015, 10, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Puja, G.; Jyoti, V. Culturable bacterial diversity and hydrolytic enzymes from Drass, a cold desert in India. Afr. J. Microbiol. Res. 2015, 9, 1866–1876. [Google Scholar] [CrossRef] [Green Version]
- Dutta, D.; Ghosh, K. Screening of extracellular enzyme-producing and pathogen inhibitory gut bacteria as putative probiotics in Mrigal, Cirrhinus Mrigala (Hamilton, 1822). Int. J. Fish Aquat. Stud. 2015, 2, 310–318. [Google Scholar]
- Yang, Y.; Gao, Y.; Liu, Y.; Liu, B.; Wang, D.; Xu, Y.; Wei, Y. Pseudomonas marianensis sp. Nov., a marine bacterium isolated from deep-sea sediments of the mariana trench. Arch. Microbiol. 2022, 204, 638. [Google Scholar] [CrossRef]
- Ismail, M.I.; Azizan, N.H.; Ashaari, M.M. Isolation and screening of biosurfactant-producing marine bacteria from Kuantan Port, Pahang, Malaysia. Sci. Heritage J. 2018, 2, 21–26. [Google Scholar] [CrossRef]
- Al-Dhabaan, F.A. Morphological, biochemical and molecular identification of petroleum hydrocarbons biodegradation bacteria isolated from oil polluted soil in Dhahran, Saud Arabia. Saudi J. Biol. Sci. 2019, 26, 1247–1252. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.E.; Smith, H.R. Benson’s Microbiological Applications Laboratory Manual in General Microbiology; McGraw-Hill Education: New York, NY, USA, 2017. [Google Scholar]
- Topic Popovic, N.; Benussi Skukan, A.; Strunjak-Perovic, I.; Coz-Rakovac, R.; Hacmanjek, M.; Hunjak, B. Comparison of the API 20E and BBL Crystal E/NF identification systems for differentiating bacterial isolates from apparently healthy reared sea bass (Dicentrarchus labrax). Vet. Res. Commun. 2004, 28, 93–101. [Google Scholar] [CrossRef]
- Das, P.; Chatterjee, S.; Behera, B.K.; Dangar, T.K.; Das, B.K.; Mohapatra, T. Isolation and characterization of marine bacteria from east coast of India: Functional screening for salt stress tolerance. Heliyon 2019, 5, e01869. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.I.; Rahman, M.M.; Salam, M.A.; Khan, M.A.R.; Islam, M.T. Identification of marine sponge-associated bacteria of the Saint Martin’s Island of the bay of bengal emphasizing on the prevention of motile Aeromonas septicemia in Labeo Rohita. Aquaculture 2021, 545, 7156. [Google Scholar] [CrossRef]
- Jeganathan, P.; Rajasekaran, K.M.; Asha Devi, N.K.; Karuppusamy, S. Antimicrobial activity and characterization of marine bacteria. Indian J. Pharm. Biol. Res. 2013, 1, 38–44. [Google Scholar] [CrossRef]
- Petersen, L.-E.; Kellermann, M.Y.; Schupp, P.J. Secondary metabolites of marine microbes: From natural products chemistry to chemical ecology. Ocean. Res. Future 2020, 9, 159–180. [Google Scholar] [CrossRef] [Green Version]
- Gopikrishnan, V.; Radhakrishnan, M.; Shanmugasundaram, T.; Pazhanimurugan, R.; Balagurunathan, R. Antibiofouling potential of quercetin compound from marine-derived actinobacterium, Streptomyces fradiae PE7 and its characterization. Environ. Sci. Pollut. Res. 2016, 23, 13832–13842. [Google Scholar] [CrossRef] [PubMed]
- Srilekha, V.; Krishna, G.; Sesha Srinivas, G. Singara Charya MA Isolation and screening of marine pigmented bacteria from Nellore Coast for antimicrobial studies. J. Cell Tissue Res. 2016, 16, 5413–5419. [Google Scholar]
- Rosaline, X.D.; Sakthivelkumar, S.; Rajendran, K.; Janarthanan, S. Screening of selected marine algae from the Coastal Tamil Nadu, South India for antibacterial activity. Asian Pac. J. Trop. Biomed. 2012, 2, 140–146. [Google Scholar] [CrossRef]
- Sujatha, R.; Siva, D.; Mohideen, P.; Nawas, A. Screening of phytochemical profile and antibacterial activity of various solvent extracts of marine algae Sargassum swartzii. World Sci. News 2019, 115, 27–40. [Google Scholar]
- Jayatilake, G.S.; Thornton, M.P.; Leonard, A.C.; Grimwade, J.E.; Baker, B.J. Metabolites from an antarctic sponge-associated bacterium, Pseudomonas aeruginosa. J. Nat. Prod. 1996, 59, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Paul, D.; Sinha, S.N. Isolation and characterization of phosphate solubilizing Bacterium pseudomonas aeruginosa KUPSB12 with antibacterial potential from River Ganga, India. Ann. Agrar. Sci. 2017, 15, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Wilson, G.S.; Raftos, D.A.; Nair, S.V. Antimicrobial activity of surface attached marine bacteria in biofilms. Microbiol. Res. 2011, 166, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, S.H.; Nagle, V.L.; Jagtap, T.G. Antifouling Potential of Some Marine Organisms from India against Species of Bacillus and Pseudomonas. Mar. Biotechnol. 2002, 4, 111–118. [Google Scholar] [CrossRef]
- Satheesh, S.; Ba-Akdah, M.A.; Al-Sofyani, A.A. Natural antifouling compound production by microbes associated with marine macroorganisms—A review. Electron. J. Biotechnol. 2016, 21, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.I.; Gray, K.A. Chemical composition and Cu complexation characteristics of the extracellular polymeric substances from Pseudomonas aeruginosa biofilms. Metalorg. Interact. Environ. Syst. 2003, 43, 529–530. [Google Scholar]
- Sarala, V.; Radhakrishnan, M.; Balagurunathan, R. Inhibitory activity of terpenoid from the medicinal plant Andrographis paniculata against biofouling bacteria. Int. J. Chemtech. Res. 2011, 3, 1225–1231. [Google Scholar]
- Chebbi, A.; Elshikh, M.; Haque, F.; Ahmed, S.; Dobbin, S.; Marchant, R.; Sayadi, S.; Chamkha, M.; Banat, I.M. Rhamnolipids from Pseudomonas aeruginosa strain W10; as antibiofilm/antibiofouling products for metal protection. J. Basic Microbiol. 2017, 57, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of Staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Pratt, L.A.; Kolter, R. Genetic analysis of escherichia coli biofilm formation: Roles of flagella, motility, chemotaxis and type I pili. Mol. Microbiol. 1998, 30, 285–293. [Google Scholar] [CrossRef]
- Nurioglu, A.G.; Esteves, A.C.C.; de With, G. Non-toxic, non-biocide-release antifouling coatings based on molecular structure design for marine applications. J. Mater. Chem. B 2015, 3, 6547–6570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archana, S.; Sundaramoorthy, B. Review on biofouling prevention using nanotechnology. J. Entomol. Zool. Stud. 2019, 7, 640–648. [Google Scholar]
- Agostini, V.O.; Macedo, A.J.; Muxagata, E.; da Silva, M.V.; Pinho, G.L.L. Non-toxic antifouling potential of caatinga plant extracts: Effective inhibition of marine initial biofouling. Hydrobiologia 2020, 847, 45–60. [Google Scholar] [CrossRef]
- Trentin, D.D.S.; Giordani, R.B.; Zimmer, K.R.; da Silva, A.G.; da Silva, M.V.; Correia, M.T.D.S.; Baumvol, I.J.R.; MacEdo, A.J. Potential of medicinal plants from the Brazilian Semi-Arid Region (Caatinga) against Staphylococcus epidermidis Planktonic and Biofilm Lifestyles. J. Ethnopharmacol. 2011, 137, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Malafaia, C.B.; Jardelino, A.C.S.; da Silva, A.G.; de Souza, E.B.; Macedo, A.J.; dos Correia, M.T.S.; Silva, M.V. Effects of Caatinga plant extracts in planktonic growth and biofilm formation in Ralstonia solanacearum. Microb. Ecol. 2018, 75, 555–561. [Google Scholar] [CrossRef]
- Silva, L.N.; Trentin, D.D.S.; Zimmer, K.R.; Treter, J.; Brandelli, C.L.C.; Frasson, A.P.; Tasca, T.; da Silva, A.G.; da Silva, M.V.; Macedo, A.J. Anti-infective effects of Brazilian Caatinga plants against pathogenic bacterial biofilm formation. Pharm. Biol. 2015, 53, 464–468. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, K.; Hashemian, A.M.; Bolvardi, E.; Hosseini, P.K. Vancomycin-resistant Pseudomonas aeroginosa in the cases of trauma. Med. Arch. 2016, 70, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Maimaitiming, M.; Zhou, Y.; Li, H.; Wang, P.; Liu, Y.; Schäberle, T.F.; Liu, Z.; Wang, C.Y. Discovery of marine natural products as promising antibiotics against Pseudomonas aeruginosa. Mar. Drugs 2022, 20, 192. [Google Scholar] [CrossRef]
- Kim, J.; Kim, K.H.; Chun, B.H.; Khan, S.A.; Jeon, C.O. Flagellimonas algicola Sp. Nov., isolated from a marine red alga, Asparagopsis taxiformis. Curr. Microbiol. 2020, 77, 294–299. [Google Scholar] [CrossRef]
- Kumar, R.S.; Prabu, D.; Shankar, T.; Sankaralingam, S.; Anandapandian, K.T.K. Optimization of alkalophilic protease production by Pseudomonas aeruginosa isolated from the gut of Penaus monodon. World J. Fish Mar. Sci. 2011, 3, 371–375. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Strains Samples | B. cereus | S. uberis | P. aeruginosa | V. parahaemolyticus |
|---|---|---|---|---|
| Cell-free supernatant of bacteria isolates | − | − | − | − |
| Ethyl acetate extract of bacteria isolates | ++ | +++ | +++ | +++ |
| Antibiotic | ||||
| Kanamycin (30 μg/disc) | ++++ | ++++ | NA | NA |
| Gentamicin (10 μg/disc) | NA | NA | ++++ | ++++ |
| Methanol-soaked disk | − | − | − | − |
| Isolate | Rlimb |
|---|---|
| Morphological characteristics | |
| Gram reaction | Gram-negative |
| Size and shape | Long rod-shape bacteria |
| Colonial characteristics | |
| Form | Circular |
| Colony size | Intermediate |
| Margin | Entire |
| Pigmentation | No pigmentation |
| Elevation | Flat |
| Opacity | Opaque |
| Antibiotic Used | Results |
|---|---|
| Vancomycin (5 μg/disc) | − |
| Penicillin G (10 U/disc) | − |
| Amoxicillin (25 μg/disc) | − |
| Streptomycin (10 μg/disc) | + |
| Ampicillin (10 μg/disc) | − |
| Kanamycin (30 μg/disc) | − |
| Gentamicin (10 μg/disc) | ++ |
| Carbon Source | Results |
|---|---|
| Arabinose | − |
| Mannose | − |
| Sucrose | − |
| Melibiose | − |
| Rhamnose | − |
| Sorbitol | − |
| Mannitol | − |
| Adonitol | − |
| Galactose | − |
| Inositol | − |
| Tests | Results |
|---|---|
| Catalase | + |
| Oxidase | + |
| Protease | + |
| Lipase | + |
| Amylase | − |
| p-nitrophenyl phosphate | + |
| p-nitrophenyl α-β-glucoside | − |
| p-nitrophenyl β-galactoside | − |
| Proline nitroanilide | + |
| p-nitrophenyl bis-phosphate | − |
| p-nitrophenyl xyloside | − |
| p-nitrophenyl α-arabinoside | − |
| p-nitrophenyl phosphorylcholine | + |
| p-nitrophenyl β-glucuronide | − |
| p-nitrophenyl-N-acetyl glucosaminide | − |
| γ-L-glutamyl p-nitroanilide | + |
| Esculin hydrolysis | − |
| Phenylalanine deamination | + |
| Urea hydrolysis | + |
| Glycine degradation | + |
| Citrate utilization | + |
| Malonate utilization | + |
| Tetrazolium reduction | − |
| Arginine catabolism | + |
| Lysine catabolism | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rawi, N.N.; Ramzi, M.M.; Rahman, N.I.A.; Ariffin, F.; Saidin, J.; Bhubalan, K.; Mazlan, N.W.; Zin, N.A.M.; Siong, J.Y.F.; Bakar, K.; et al. Antifouling Potential of Ethyl Acetate Extract of Marine Bacteria Pseudomonas aeruginosa Strain RLimb. Life 2023, 13, 802. https://doi.org/10.3390/life13030802
Rawi NN, Ramzi MM, Rahman NIA, Ariffin F, Saidin J, Bhubalan K, Mazlan NW, Zin NAM, Siong JYF, Bakar K, et al. Antifouling Potential of Ethyl Acetate Extract of Marine Bacteria Pseudomonas aeruginosa Strain RLimb. Life. 2023; 13(3):802. https://doi.org/10.3390/life13030802
Chicago/Turabian StyleRawi, Nurul Najihah, Mujahidah Mohd Ramzi, Nor Izzati Abd Rahman, Fazilah Ariffin, Jasnizat Saidin, Kesaven Bhubalan, Noor Wini Mazlan, Nor Atikah Mohd Zin, Julius Yong Fu Siong, Kamariah Bakar, and et al. 2023. "Antifouling Potential of Ethyl Acetate Extract of Marine Bacteria Pseudomonas aeruginosa Strain RLimb" Life 13, no. 3: 802. https://doi.org/10.3390/life13030802