

Prospective Use of Probiotics to Maintain Astronaut Health during Spaceflight
 , , and
, , and
Abstract
:1. Introduction
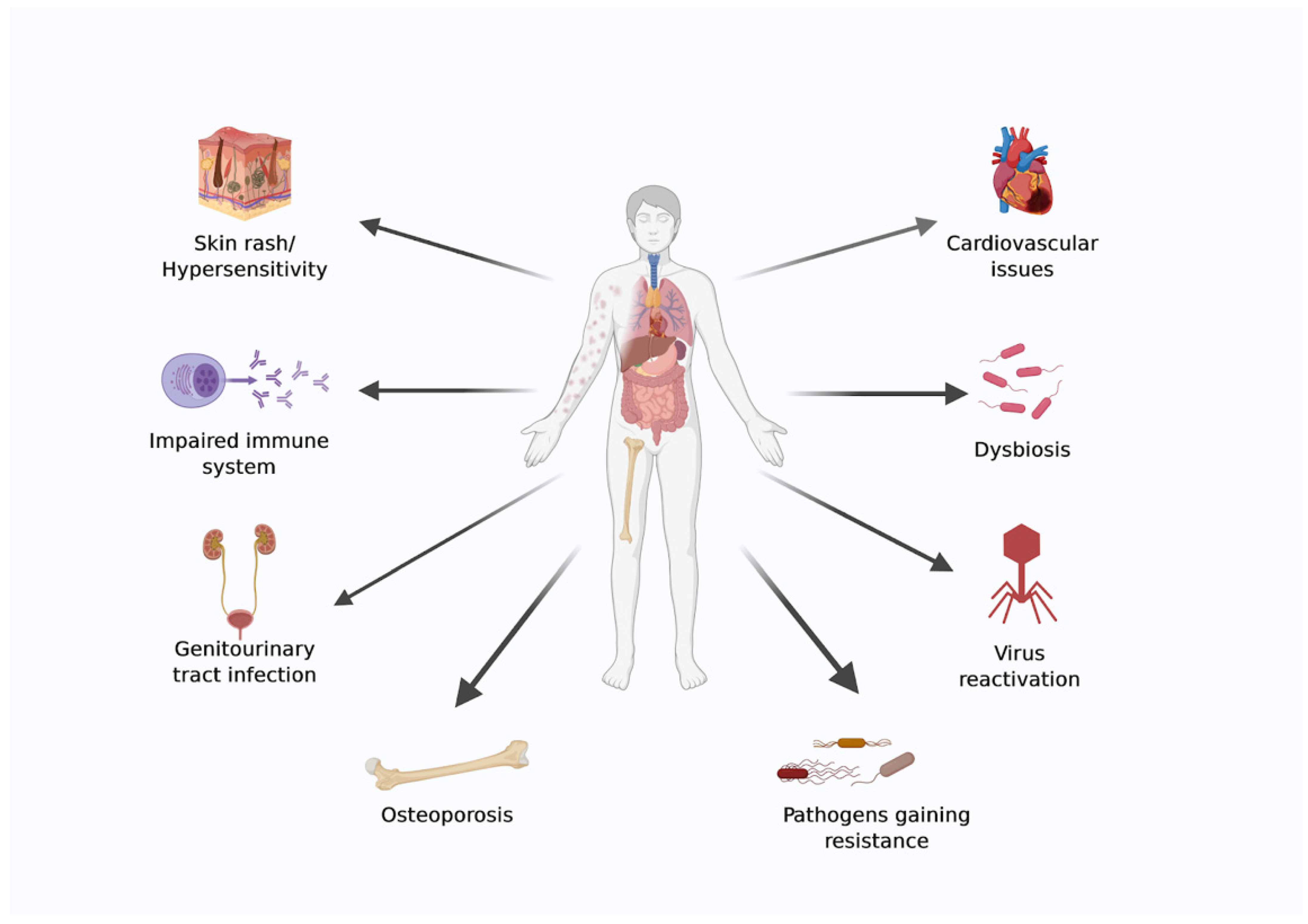
2. Health Issues during Spaceflight
2.1. Changes in the Microbiome
2.1.1. The Microbiota–Gut–Brain Axis and Its Relation to the Mental Health of Astronauts
2.1.2. Mental Health Challenges Faced by Astronauts during Spaceflight
2.2. Genitourinary Tract Infection
2.3. Virus Reactivation in Spaceflight
2.4. Resistance of Bacteria and Changes in Bacterial Virulence
2.5. Epithelial Barrier Disruption and Inflammatory Bowel Disease (IBD)
2.6. Immunological Alteration during Spaceflight
2.7. Changes in Cardiovascular Functions
2.8. Effect of Cosmic Radiation on Astronauts
3. Probiotics and Their Role in Space Biology
3.1. General Mechanism of Action of Probiotics
3.1.1. Inhibition of Pathogen Binding
3.1.2. Use of Probiotics for Intestinal Disorders
3.1.3. Immune System Maintenance
3.1.4. Antimicrobial Activity of Probiotics
3.1.5. Probiotics Used for Antibiotic-Associated Diarrhoea
3.1.6. Probiotics as Prophylaxis for Cancer
3.1.7. Probiotics for Stress/Anxiety
3.1.8. Probiotics for Urinary Tract Infection

{kind=link}
| Sr. no | Probiotics | Mechanism of Action | Primary Outcome of the Probiotic | Reference |
|---|---|---|---|---|
| 1 | Lactobacillus rhamnosus GG and L. plantarum | Inhibition of pathogen binding | Has been shown to inhibit the attachment of pathogenic E. coli to the epithelium | (Wilson and Perini et al., 1988), [74] |
| 2 | Escherichia coli strain Nissle 1917 (EcN) and Lactobacillus plantarum MB452 | Intestinal disorders | Escherichia coli strain Nissle 1917 (EcN) restoration of disrupted epithelial cells Lactobacillus plantarum MB452 enhances intestinal barrier integrity | (Zyrek et al., 2007) and (Ulluwishewa et al., 2011), [41,67] |
| 3 | Lactobacillus rhamnosus GG | Immune system maintenance | Induction of IL-6 production | (Yan and Polk, 2011), [80] |
| 4 | Lactic acid bacteria and Bifidobacteria | Antimicrobial activity | By synthesis of organic acids, toxic substances, and bacteriocins | (Bermudez-Brito et al., 2012) and (Dunne at al., 2001), [85,86] |
| 5 | Kefir | Prevention of cancer | Production of bioactive compounds which can inhibit proliferation and apoptosis induction in tumour cells | (Sharifi et al., 2017), [96] |
| 6 | Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri RC-14 | Prevention of urinary tract infections | Anti-infective properties | (Urbaniak and Reid, 2016), [3] |
3.1.9. Short-Chain Fatty Acids and Their Role in Gut Microbiota Maintenance
3.2. Microgravity/Simulated Microgravity Studies on Probiotics
3.3. Commercial Probiotics’ Shelf Life and Survival in a Simulated Gastrointestinal Tract
- Long-term storage at ambient conditions;
- Simulated galactic cosmic radiation and solar particle event radiation;
- Exposure to simulated gastric fluid;
- Exposure to simulated intestinal fluid.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Voorhies, A.A.; Mark Ott, C.; Mehta, S.; Pierson, D.L.; Crucian, B.E.; Feiveson, A.; Oubre, C.M.; Torralba, M.; Moncera, K.; Zhang, Y.; et al. Study of the impact of long-duration space missions at the International Space Station on the astronaut microbiome. Sci. Rep. 2019, 9, 9911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, J.; Cho, S.W.; Kim, B.; Park, S.; Han, Y.H.; Seo, S.W. Transcriptional profiling of the probiotic Escherichia coli nissle 1917 strain under simulated microgravity. Int. J. Mol. Sci. 2020, 21, 2666. [Google Scholar] [CrossRef]
- Urbaniak, C.; Reid, G. The potential influence of the microbiota and probiotics on women during long spaceflights. Womens Health 2016, 12, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Takada, T.; Shimizu, K.; Kado, Y.; Kawakami, K.; Makino, I.; Yamaoka, Y.; Hirano, K.; Nishimura, A.; Kajimoto, O.; et al. The effects of a probiotic milk product containing Lactobacillus casei strain shirota on the defecation frequency and the intestinal microflora of sub-optimal health state volunteers: A randomized placebo-controlled cross-over study. Biosci. Microflora 2006, 25, 39–48. [Google Scholar] [CrossRef]
- Sakai, T.; Moteki, Y.; Takahashi, T.; Shida, K.; Kiwaki, M.; Shimakawa, Y.; Matsui, A.; Chonan, O.; Morikawa, K.; Ohta, T.; et al. Probiotics into outer space: Feasibility assessments of encapsulated freeze-dried probiotics during 1 month’s storage on the International Space Station. Sci. Rep. 2018, 8, 10687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turroni, S.; Rampelli, S.; Biagi, E.; Consolandi, C.; Severgnini, M.; Peano, C.; Quercia, S.; Soverini, M.; Carbonero, F.G.; Bianconi, G.; et al. Temporal dynamics of the gut microbiota in people sharing a confined environment, a 520-day ground-based space simulation, MARS500. Microbiome 2017, 5, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia muciniphila is a promising probiotic. Microb. Biotechnol. 2019, 12, 1109–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [Green Version]
- Gotova, I.; Dimitrov, Z.; Najdenski, H. Selected Lactobacillus bulgaricus and Streptococcus thermophilus strains from bulgarian yogurt demonstrate significant anti-inflammatory potential. Acta Microbiol. Bulg 2017, 33, 137–142. [Google Scholar]
- Day, R.L.; Harper, A.J.; Woods, R.M.; Davies, O.G.; Heaney, L.M. Probiotics: Current landscape and future horizons. Future Sci. OA 2019, 5, FSO391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraf, K.; Shashikanth, M.C.; Priy, T.; Sultana, N.; Chaitanya, N.C. Probiotics-do they have a role in medicine and dentistry. J. Assoc. Physicians India 2010, 58, 488–490. [Google Scholar]
- Douglas, G.; Voorhies, A. Evidence based selection of probiotic strains to promote astronaut health or alleviate symptoms of illness on long duration spaceflight missions. Benef. Microbes 2017, 8, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.; Stowe, R.P.; Mehta, S.; Quiriarte, H.; Pierson, D.; Sams, C. Alterations in adaptive immunity persist during long-duration spaceflight. npj Microgravity 2015, 1, 15013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Luo, G.; Du, R.; Sun, W.; Li, J.; Lan, H.; Chen, P.; Yuan, X.; Cao, D.; Li, Y.; et al. Effects of spaceflight on the composition and function of the human gut microbiota. Gut Microbes 2020, 11, 807–819. [Google Scholar] [CrossRef]
- O’Flaherty, S.; Klaenhammer, T.R. The role and potential of probiotic bacteria in the gut, and the communication between gut microflora and gut/host. Int. Dairy J. 2010, 20, 262–268. [Google Scholar] [CrossRef]
- Turroni, F.; Ventura, M.; Butto, L.F.; Duranti, S.; O’Toole, P.W.; Motherway, M.O.; Van Sinderen, D. Molecular dialogue between the human gut microbiota and the host: A Lactobacillus and Bifidobacterium perspective. Cell. Mol. Life Sci. 2014, 71, 183–203. [Google Scholar] [CrossRef]
- Wall, R.; Cryan, J.F.; Ross, R.P.; Fitzgerald, G.F.; Dinan, T.G.; Stanton, C. Bacterial neuroactive compounds produced by psychobiotics. Adv. Exp. Med. Biol. 2014, 817, 221–239. [Google Scholar]
- Blaber, E.; Marçal, H.; Burns, B.P. Bioastronautics: The influence of microgravity on astronaut health. Astrobiology 2010, 10, 463–473. [Google Scholar] [CrossRef]
- Crucian, B.; Babiak-Vazquez, A.; Johnston, S.; Pierson, D.L.; Ott, C.M.; Sams, C. Incidence of clinical symptoms during long-duration orbital spaceflight. Int. J. Gen. Med. 2016, 2016, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA twins study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364, 6436. [Google Scholar] [CrossRef]
- Siddiqui, R.; Qaisar, R.; Khan, N.A.; Alharbi, A.M.; Alfahemi, H.; Elmoselhi, A. Effect of Microgravity on the Gut Microbiota Bacterial Composition in a Hindlimb Unloading Model. Life 2022, 12, 1865. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Green, S.J.; Chlipala, G.E.; Turek, F.W.; Vitaterna, M.H. Reproducible changes in the gut microbiome suggest a shift in microbial and host metabolism during spaceflight. Microbiome 2019, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar] [PubMed]
- Sajdel-Sulkowska, E.M. Disruption of the Microbiota-Gut-Brain (MGB) Axis and Mental Health of Astronauts During Long-Term Space Travel. In Handbook of the Cerebellum and Cerebellar Disorders; Manto, M., Gruol, D., Schmahmann, J., Koibuchi, N., Sillitoe, R., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Palinkas, L.A. Psychosocial issues in long-term space flight: Overview. Gravit. Space Biol. Bull. 2001, 14, 25–33. [Google Scholar]
- Mallis, M.M.; DeRoshia, C.W. Circadian rhythms, sleep, and performance in space. Aviat. Space Environ. Med. 2005, 76, B94–B107. [Google Scholar] [PubMed]
- Oluwafemi, F.A.; Abdelbaki, R.; Lai, J.C.Y.; Mora-Almanza, J.G.; Afolayan, E.M. A review of astronaut mental health in manned missions: Potential interventions for cognitive and mental health challenges. Life Sci. Space Res. 2021, 28, 26–31. [Google Scholar] [CrossRef]
- Arone, A.; Ivaldi, T.; Loganovsky, K.; Palermo, S.; Parra, E.; Flamini, W.; Marazziti, D. The burden of space exploration on the mental health of astronauts: A narrative review. Clin. Neuropsychiatry 2021, 18, 237. [Google Scholar] [PubMed]
- Zivi, P.; De Gennaro, L.; Ferlazzo, F. Sleep in isolated, confined, and extreme (ICE): A review on the different factors affecting human sleep in ICE. Front. Neurosci. 2020, 14, 851. [Google Scholar] [CrossRef]
- Ma, T.; Jin, H.; Kwok, L.Y.; Sun, Z.; Liong, M.T.; Zhang, H. Probiotic consumption relieved human stress and anxiety symptoms possibly via modulating the neuroactive potential of the gut microbiota. Neurobiol. Stress 2021, 14, 100294. [Google Scholar] [CrossRef]
- Hao, Z.; Li, L.; Fu, Y.; Liu, H. The influence of bioregenerative life-support system dietary structure and lifestyle on the gut microbiota: A 105-day ground-based space simulation in Lunar Palace 1. Environ. Microbiol. 2018, 20, 3643–3656. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Jennings, R.; Pietryzk, R.; Ciftcioglu, N.; Stepaniak, P. Genitourinary issues during spaceflight: A review. Int. J. Impot. Res. 2005, 17 (Suppl. 1), S64–S67. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Luderer, U. Reproductive hazards of space travel in women and men. Nat Rev Endocrinol. 2019, 15, 713–730, Erratum in Nat. Rev. Endocrinol. 2019, 15, 713–730. [Google Scholar] [CrossRef] [PubMed]
- Sonnenfeld, G.; Shearer, W.T. Immune function during space flight. Nutrition 2002, 18, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Rooney, B.V.; Crucian, B.E.; Pierson, D.L.; Laudenslager, M.L.; Mehta, S.K. Herpes virus reactivation in astronauts during spaceflight and its application on earth. Front. Microbiol. 2019, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Bai, P.; Zhao, X.; Yu, Y.; Zhang, X.; Li, D.; Liu, C. Increased growth rate and amikacin resistance of Salmonella enteritidis after one-month spaceflight on China’s Shenzhou-11 spacecraft. MicrobiologyOpen 2019, 8, e00833. [Google Scholar] [CrossRef] [Green Version]
- Tirumalai, M.R.; Karouia, F.; Tran, Q.; Stepanov, V.G.; Bruce, R.J.; Ott, C.M.; Pierson, D.L.; Fox, G.E. Evaluation of acquired antibiotic resistance in Escherichia coli exposed to long-term low-shear modeled microgravity and background antibiotic exposure. MBio 2019, 10, e02637-18. [Google Scholar] [CrossRef] [Green Version]
- Morrison, M.D.; Thissen, J.B.; Karouia, F.; Mehta, S.; Urbaniak, C.; Venkateswaran, K.; Smith, D.J.; Jaing, C. Investigation of Spaceflight Induced Changes to Astronaut Microbiomes. Front. Microbiol. 2021, 12, 659179. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Zyrek, A.A.; Cichon, C.; Helms, S.; Enders, C.; Sonnenborn, U.; Schmidt, M.A. Molecular mechanisms underlying the probiotic effects of Escherichia coli Nissle 1917 involve ZO-2 and PKCζ redistribution resulting in tight junction and epithelial barrier repair. Cell. Microbiol. 2007, 9, 804–816. [Google Scholar] [CrossRef]
- Johnson-Henry, K.C.; Donato, K.A.; Shen-Tu, G.; Gordanpour, M.; Sherman, P.M. Lactobacillus rhamnosus strain GG prevents enterohemorrhagic Escherichia coli O157:H7-induced changes in epithelial barrier function. Infect. Immun. 2008, 76, 1340–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, R.; Stork, C.A.; Sayoc-Becerra, A.; Marchelletta, R.R.; Prisk, G.K.; McCole, D.F. A Simulated Microgravity Environment Causes a Sustained Defect in Epithelial Barrier Function. Sci. Rep. 2019, 9, 17531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrieta, M.C.; Bistritz, L.; Meddings, J.B. Alterations in intestinal permeability. Gut 2006, 55, 1512–1520. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H. Intestinal Permeability Regulation by Tight Junction: Implication on Inflammatory Bowel Diseases. Intest. Res. 2015, 13, 11. [Google Scholar] [CrossRef] [Green Version]
- Blair, S.A.; Kane, S.V.; Clayburgh, D.R.; Turner, J.R. Epithelial myosin light chain kinase expression and activity are upregulated in inflammatory bowel disease. Lab. Investig. 2006, 86, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Wang, L.; Xiong, Y.; Wang, Z.; Qiu, Y.; Wen, X.; Jiang, Z.; Yang, X.; Ma, X. Lactobacillus reuteri LR1 Improved Expression of Genes of Tight Junction Proteins via the MLCK Pathway in IPEC-1 Cells during Infection with Enterotoxigenic Escherichia coli K88. Mediat. Inflamm. 2018, 2018, 6434910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, I.; Simons, E.R.; Castro, V.A.; Mark Ott, C.; Pierson, D.L. Changes in neutrophil functions in astronauts. Brain Behav. Immun. 2004, 18, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Stowe, R.P.; Sams, C.F.; Pierson, D.L. Effects of Mission Duration on Neuroimmune Responses in Astronauts. Aviat. Space Environ. Med. 2003, 74, 1281–1284. [Google Scholar]
- Makedonas, G.; Mehta, S.; Choukèr, A.; Simpson, R.J.; Marshall, G.; Orange, J.S.; Aunon-Chancellor, S.; Smith, S.M.; Zwart, S.R.; Stowe, R.P.; et al. Specific Immunologic Countermeasure Protocol for Deep-Space Exploration Missions. Front. Immunol. 2019, 10, 2407. [Google Scholar] [CrossRef] [Green Version]
- Buchheim, J.I.; Matzel, S.; Rykova, M.; Vassilieva, G.; Ponomarev, S.; Nichiporuk, I.; Hörl, M.; Moser, D.; Biere, K.; Feuerecker, M.; et al. Stress related shift toward inflammaging in cosmonauts after long-duration space flight. Front. Physiol. 2019, 10, 85. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, T.; Horie, K.; Hinoi, E.; Hiraiwa, M.; Kato, A.; Maekawa, Y.; Takahashi, A.; Furukawa, S. How does spaceflight affect the acquired immune system? npj Microgravity 2020, 6, 14. [Google Scholar] [CrossRef]
- Kaur, I.; Simons, E.R.; Castro, V.A.; Ott, C.M.; Pierson, D.L. Changes in monocyte functions of astronauts. Brain Behav. Immun. 2005, 19, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Voss, E.W. Prolonged weightlessness and humoral immunity. Science 1984, 225, 214–215. [Google Scholar] [CrossRef] [PubMed]
- Mills, P.J.; Meck, J.V.; Waters, W.W.; D’Aunno, D.; Ziegler, M.G. Peripheral leukocyte subpopulations and catecholamine levels in astronauts as a function of mission duration. Psychosom. Med. 2001, 63, 886–890. [Google Scholar] [CrossRef] [PubMed]
- Stowe, R.P.; Sams, C.F.; Mehta, S.K.; Kaur, I.; Jones, M.L.; Feeback, D.L.; Pierson, D.L. Leukocyte subsets and neutrophil function after short-term spaceflight. J. Leukoc. Biol. 1999, 65, 179–186. [Google Scholar] [CrossRef]
- Guell, A.; Braak, L. Cardiovascular deconditioning syndrome during space flights. Ann. Cardiol. D’angéiol. Paris 1989, 38, 499–502. [Google Scholar]
- Tang, H.; Rising, H.H.; Majji, M.; Brown, R.D. Long-Term Space Nutrition: A Scoping Review. Nutrients 2021, 14, 194. [Google Scholar] [CrossRef]
- Caiani, E.G.; Martin-Yebra, A.; Landreani, F.; Bolea, J.; Laguna, P.; Vaïda, P. Weightlessness and Cardiac Rhythm Disorders: Current Knowledge from Space Flight and Bed-Rest Studies. Front. Astron. Space Sci. 2016, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Jian, Y.; Zhang, D.; Liu, M.; Wang, Y.; Xu, Z.-X. The Impact of Gut Microbiota on Radiation-Induced Enteritis. Front. Cell. Infect. Microbiol. 2021, 11, 586392. [Google Scholar] [CrossRef]
- Tesei, D.; Jewczynko, A.; Lynch, A.; Urbaniak, C. Understanding the Complexities and Changes of the Astronaut Microbiome for Successful Long-Duration Space Missions. Life 2022, 12, 495. [Google Scholar] [CrossRef]
- Durante, M.; Cucinotta, F.A. Heavy ion carcinogenesis and human space exploration. Nat. Rev. Cancer 2008, 8, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Stahn, A.C.; Werner, A.; Opatz, O.; Maggioni, M.A.; Steinach, M.; von Ahlefeld, V.W.; Moore, A.; Crucian, B.E.; Smith, S.M.; Zwart, S.R.; et al. Increased core body temperature in astronauts during long-duration space missions. Sci. Rep. 2017, 7, 16180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kligler, B.; Cohrssen, A. Probiotics. 2008. Available online: www.aafp.org/afp (accessed on 25 January 2022).
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the future of probiotics and prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef]
- Shi, L.H.; Balakrishnan, K.; Thiagarajah, K.; Mohd Ismail, N.I.; Yin, O.S. Beneficial properties of probiotics. Trop. Life Sci. Res. 2016, 27, 73–90. [Google Scholar] [CrossRef]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mack, D.R.; Ahrne, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extracellular MUC3 mucin secretion follows adherence of Lactobacillus strains to intestinal epithelial cells in vitro. Gut 2003, 52, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Caballero-Franco, C.; Keller, K.; De Simone, C.; Chadee, K. The VSL#3 probiotic formula induces mucin gene expression and secretion in colonic epithelial cells. Am. J. Physiol.—Gastrointest. Liver Physiol. 2007, 292, G315–G322. [Google Scholar] [CrossRef]
- Otte, J.M.; Podolsky, D.K. Functional modulation of enterocytes by gram-positive and gram-negative microorganisms. Am. J. Physiol.—Gastrointest. Liver Physiol. 2004, 286, G613–G626. [Google Scholar] [CrossRef] [Green Version]
- Kankainen, M.; Paulin, L.; Tynkkynen, S.; Von Ossowski, I.; Reunanen, J.; Partanen, P.; Satokari, R.; Vesterlund, S.; Hendrickx, A.P.A.; Lebeer, S.; et al. Comparative genomic analysis of Lactobacillus rhamnosus GG reveals pili containing a human-mucus binding protein. Proc. Natl. Acad. Sci. USA 2009, 106, 17193–17198. [Google Scholar] [CrossRef] [Green Version]
- Mukai, T.; Kaneko, S.; Matsumoto, M.; Ohori, H. Binding of Bifidobacterium bifidum and Lactobacillus reuteri to the carbohydrate moieties of intestinal glycolipids recognized by peanut agglutinin. Int. J. Food Microbiol. 2004, 90, 357–362. [Google Scholar] [CrossRef]
- Tallon, R.; Arias, S.; Bressollier, P.; Urdaci, M.C. Strain- and matrix-dependent adhesion of Lactobacillus plantarum is mediated by proteinaceous bacterial compounds. J. Appl. Microbiol. 2007, 102, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.H.; Perini, F. Role of competition for nutrients in suppression of Clostridium difficile by the colonic microflora. Infect. Immun. 1988, 56, 2610–2614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, C.; Zheng, C.Q.; Jiang, M.; Ma, X.Y.; Jiang, L.J. Probiotics and irritable bowel syndrome. World J. Gastroenterol. 2013, 19, 5973–5980. [Google Scholar] [CrossRef] [PubMed]
- Toi, M.; Hirota, S.; Tomotaki, A.; Sato, N.; Hozumi, Y.; Anan, K.; Nagashima, T.; Tokuda, Y.; Masuda, N.; Ohsumi, S.; et al. Probiotic Beverage with Soy Isoflavone Consumption for Breast Cancer Prevention: A Case-control Study. Curr. Nutr. Food Sci. 2013, 9, 194–200. [Google Scholar] [CrossRef]
- Buckley, N.D.; Champagne, C.P.; Masotti, A.I.; Wagar, L.E.; Tompkins, T.A.; Green-Johnson, J.M. Harnessing functional food strategies for the health challenges of space travel—Fermented soy for astronaut nutrition. Acta Astronaut. 2011, 68, 731–738. [Google Scholar] [CrossRef]
- Rao, R.K.; Samak, G. Protection and Restitution of Gut Barrier by Probiotics: Nutritional and Clinical Implications. Curr. Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galdeano, C.M.; Perdigón, G. Role of viability of probiotic strains in their persistence in the gut and inmucosal immune stimulation. J. Appl. Microbiol. 2004, 97, 673–681. [Google Scholar] [CrossRef]
- Yan, F.; Polk, D.B. Probiotics and immune health. Curr. Opin. Gastroenterol. 2011, 27, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, P.A.; Hoffmann, M.; Szcesny, S.; Blaut, M.; Haller, D. Innate mechanisms for Bifidobacterium lactis to activate transient pro-inflammatory host responses in intestinal epithelial cells after the colonization of germ-free rats. Immunology 2005, 115, 441–450. [Google Scholar] [CrossRef]
- Van Hemert, S.; Meijerink, M.; Molenaar, D.; Bron, P.A.; De Vos, P.; Kleerebezem, M.; Wells, J.M.; Marco, M.L. Identification of Lactobacillus plantarum genes modulating the cytokine response of human peripheral blood mononuclear cells. BMC Microbiol. 2010, 10, 293. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Polk, D.B. Probiotic bacterium prevents cytokine-induced apoptosis in intestinal epithelial cells. J. Biol. Chem. 2002, 277, 50959–50965. [Google Scholar] [CrossRef] [Green Version]
- Turroni, S.; Magnani, M.; Kc, P.; Lesnik, P.; Vidal, H.; Heer, M. Gut Microbiome and Space Travelers’ Health: State of the Art and Possible Pro/Prebiotic Strategies for Long-Term Space Missions. Front. Physiol. 2020, 11, 553929. [Google Scholar] [CrossRef]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Dunne, C.; O’Mahony, L.; Murphy, L.; Thornton, G.; Morrissey, D.; O’Halloran, S.; Feeney, M.; Flynn, S.; Fitzgerald, G.; Daly, C.; et al. In vitro selection criteria for probiotic bacteria of human origin: Correlation with in vivo findings. Am. J. Clin. Nutr. 2001, 73, S386–S392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplice, E.; Fitzgerald, G.F. Food fermentations: Role of microorganisms in food production and preservation. Int. J. Food Microbiol. 1999, 50, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Solberg, M. Mechanisms of action of food preservation procedures. Trends Food Sci. Technol. 1991, 2, 155. [Google Scholar] [CrossRef]
- Zacharof, M.P.; Lovitt, R.W. Bacteriocins Produced by Lactic Acid Bacteria a Review Article. APCBEE Procedia 2012, 2, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Parada, J.L.; Caron, C.R.; Medeiros, A.B.P.; Soccol, C.R. Bacteriocins from lactic acid bacteria: Purification, properties and use as biopreservatives. Braz. Arch. Biol. Technol. 2007, 50, 512–542. [Google Scholar] [CrossRef] [Green Version]
- Preidis, G.A.; Versalovic, J. Targeting the Human Microbiome with Antibiotics, Probiotics, and Prebiotics: Gastroenterology Enters the Metagenomics Era. Gastroenterology 2014, 136, 2015–2031. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; Park, Z.; McCann, M.J.; Kelly, W.J.; Roy, N.C. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 2010, 10, 316. [Google Scholar] [CrossRef] [Green Version]
- Pattani, R.; Pattani, R.; Palda, V.A.; Hwang, S.W. Probiotics for the prevention of antibiotic-associated diarrhea and Clostridium difficile infection among hospitalized patients: Systematic review and meta-analysis. Open Med. 2013, 7, e56. [Google Scholar]
- Singh, B.P.; Yadav, D.; Vij, S. Soybean Bioactive Molecules: Current Trend and Future Prospective; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–29. [Google Scholar] [CrossRef]
- Ambalam, P.; Raman, M.; Purama, R.K.; Doble, M. Probiotics, prebiotics and colorectal cancer prevention. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, M.; Moridnia, A.; Mortazavi, D.; Salehi, M.; Bagheri, M.; Sheikhi, A. Kefir: A powerful probiotics with anticancer properties. Med. Oncol. 2017, 34, 183. [Google Scholar] [CrossRef] [PubMed]
- Geuna, S.; Brunelli, F.; Perino, M.A. Stressors, stress and stress consequences during long-duration manned space missions: A descriptive model. Acta Astronaut. 1995, 36, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Jeong, C.; Cheng, W.; Choi, Y.; Shin, D.; Lee, S.; Han, S. Quality Characteristics of Yogurts Fermented with Short-Chain Fatty Acid-Producing Probiotics and Their Effects on Mucin Production and Probiotic Adhesion onto Human Colon Epithelial Cells. J. Dairy Sci. 2021, 104, 7415–7425. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; O’Riordan, M.X.D. Regulation of bacterial pathogenesis by intestinal short-chain fatty acids. Adv. Appl. Microbiol. 2013, 85, 93–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. In Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Kolodziejczyk, A.A.; Zheng, D.; Elinav, E. Diet–microbiota interactions and personalized nutrition. Nat. Rev. Microbiol. 2019, 17, 742–753. [Google Scholar] [CrossRef]
- Parladore, S.Y.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Li, Z.R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef] [Green Version]
- Shao, D.; Yao, L.; Riaz, M.S.; Zhu, J.; Shi, J.; Jin, M.; Huang, Q.; Yang, H. Simulated microgravity affects some biological characteristics of Lactobacillus acidophilus. Appl. Microbiol. Biotechnol. 2017, 101, 3439–3449. [Google Scholar] [CrossRef] [PubMed]
- Senatore, G.; Mastroleo, F.; Leys, N.; Mauriello, G. Growth of Lactobacillus reuteri DSM17938 under two simulated microgravity systems: Changes in reuterin production, gastrointestinal passage resistance, and stress genes expression response. Astrobiology 2020, 20, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Fajardo-Cavazos, P.; Nicholson, W.L. Shelf Life and Simulated Gastrointestinal Tract Survival of Selected Commercial Probiotics during a Simulated Round-Trip Journey to Mars. Front. Microbiol. 2021, 12, 748950. [Google Scholar] [CrossRef] [PubMed]

| S. No | Study | Changes Observed during Spaceflight | Reference |
|---|---|---|---|
| 1 | The NASA Twin Study: one comparative study on the impact of spaceflight on one twin the twin served as genetically match ground control |
| (Garrett-Bakelman et al., 2019), [21] |
| 2 | The impact of long-term spaceflight on the microbiome of nine astronauts who spent six to twelve months on the International Space Station (ISS) was studied. |
| (Voorhies et al., 2019), [1] |
| 3 | Effect of short-term spaceflight on human microbiota (fecal sample study) |
| (Liu et al., 2020), [15] |
| 4 | STARMAPs Test (similarity in two space research database findings) |
| (Jiang et al., 2019), [23] |
| 5 | Lunar Palace 1 (Bliss Study), study of gut microbiota of astronauts and humans on Earth. |
| (Hao et al., 2018), [32] |
| Sr. no. | Duration of Space-Flight | No. of Individuals | Changes in Immune Cells | Reference |
|---|---|---|---|---|
| 1. | 5–11 days | 25 | An 85% increase in the granulocytes was observed as compared to pre-flight values with a significant reduction in phagocytosis and oxidative burst capacities. | Kaur et al., 2004, [48] |
| 2. | 5–11 days | 25 | A reduction in ability to engulf E. coli, oxidative burst and degranulation was elucidated by monocytes following spaceflight. A reduction in phagocytosis was observed with changes observed in the expression of surface markers. | Kaur et al., 2005, [53] |
| 3. | 10 days | 4 | The continuous production of immunoglobulins was prolonged in weightlessness and the process of lymphocyte activation may be impaired and hence altering the responses to new antigenic stimuli in microgravity conditions. | Voss, 1984, [54] |
| 4. | 4–16 days | 11 | A significant increase in the number of circulating WBCs, neutrophils, monocytes, T-helper cells and B cells. The number of NK cells decreased. | Mills et al., 2001, [55] |
| 5. | 8–15 days | 16 | A slight decrease in lymphocytes and a 1.5-fold increase in the neutrophil number with increased adhesion to the endothelial cells after spaceflight. | Stowe et al., 1999, [56] |
| 6. | 9–16 days | 28 | An increase in CD4+ cells, polymorphonuclear leukocytes and monocytes whereas there was a decrease in NK cells and monocytes after 9-day and 16-day spaceflight, respectively. | Stowe et al., 2003, [49] |
| 7. | 6 months | 23 | A reduction in the function of T-cells was observed with alterations in CD8+ cells. An increase in the number and redistribution of WBCs was observed. | Crucian et al., 2015, [14] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bharindwal, S.; Goswami, N.; Jha, P.; Pandey, S.; Jobby, R. Prospective Use of Probiotics to Maintain Astronaut Health during Spaceflight. Life 2023, 13, 727. https://doi.org/10.3390/life13030727
Bharindwal S, Goswami N, Jha P, Pandey S, Jobby R. Prospective Use of Probiotics to Maintain Astronaut Health during Spaceflight. Life. 2023; 13(3):727. https://doi.org/10.3390/life13030727
Chicago/Turabian StyleBharindwal, Sahaj, Nidhi Goswami, Pamela Jha, Siddharth Pandey, and Renitta Jobby. 2023. "Prospective Use of Probiotics to Maintain Astronaut Health during Spaceflight" Life 13, no. 3: 727. https://doi.org/10.3390/life13030727