

Exploring the Potentiality of Native Actinobacteria to Combat the Chilli Fruit Rot Pathogens under Post-Harvest Pathosystem
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit Rot Pathogens
2.2. Antagonistic Actinobacteria
2.3. In Vitro Antifungal Bioassay
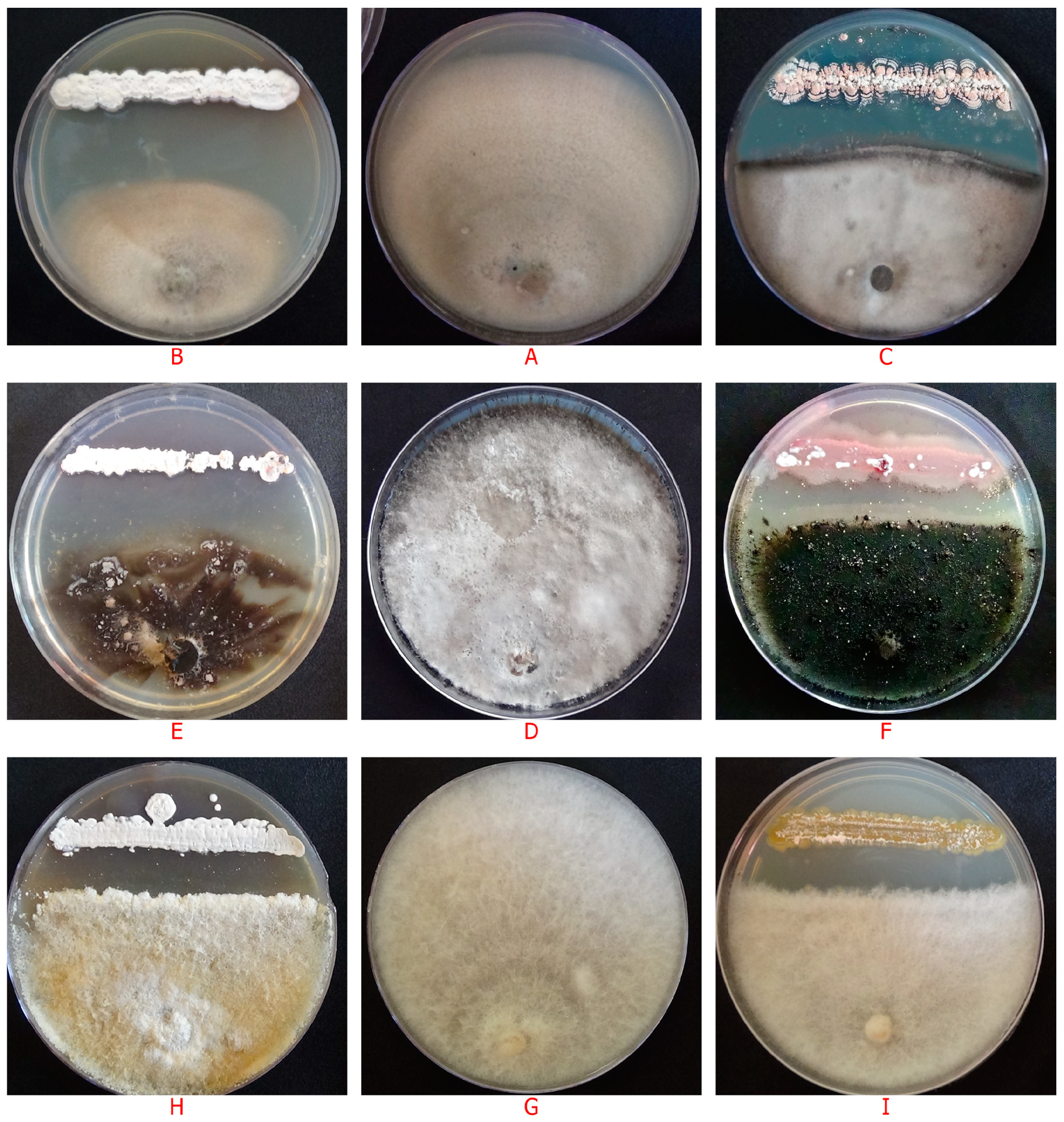
2.3.1. Primary Screening of Actinobacterial Isolates for the Antifungal Activity against Chilli Fruit Rot Pathogens
2.3.2. Secondary Screening for the Antifungal Activity of Actinobacterial Isolates
2.3.3. In Vitro Screening of Actinobacterial Isolates for Production of Extracellular Lytic Enzymes and Siderophore
2.3.4. Antifungal Activity of Volatile Organic Compounds
2.3.5. Antifungal Activity of Non-Volatile Metabolites
2.3.6. Antifungal Activity of Thermostable Compounds
2.3.7. Assessment of In Vitro Antifungal Traits
2.4. Scanning Electron Microscopy (SEM)
2.5. Molecular Characterization of Actinobacterial Isolates
2.6. Antifungal Bioassay of Liquid Formulation of Actinobacterial Isolate on Chilli Fruits
2.7. Antifungal Bioassay of Soluble Metabolites of Actinobacterial Isolate on Chilli Fruits
2.8. Statistical Analysis
3. Results
3.1. Primary Screening for Antifungal Activity of Actinobacterial Isolates
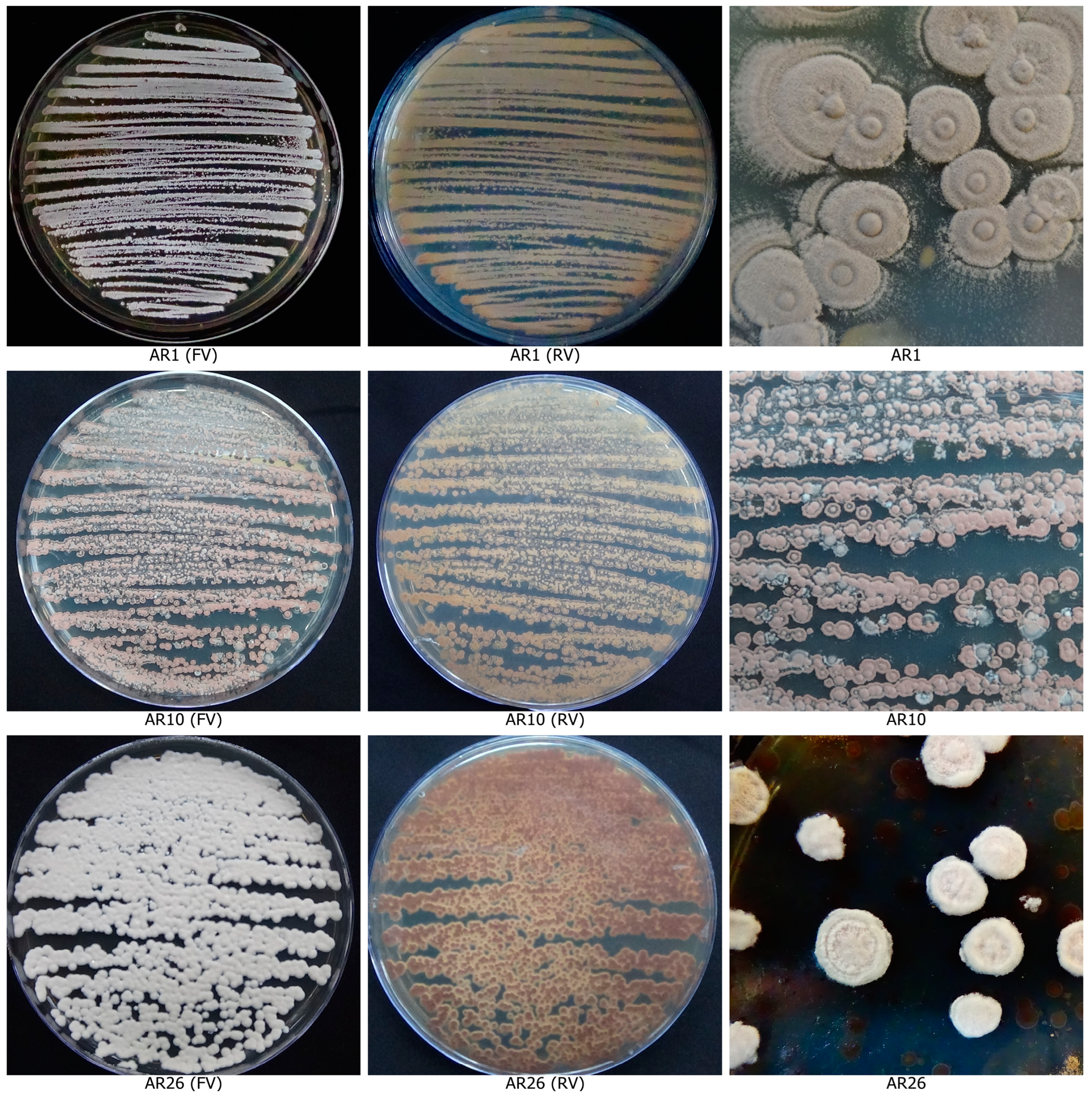
3.2. Secondary Screening for the Antifungal Activity of Actinobacterial Isolates and Scanning Electron Microscopic Assay
3.3. Screening for the Production of Extracellular Lytic Enzymes and Siderophore by the Antagonists
3.4. Antifungal Activity of Volatile, Non-Volatile and Thermostable Compounds
3.5. Assessment of In Vitro Antifungal Traits
3.6. Molecular Confirmation of Actinobacterial Isolates
3.7. Biocontrol Potential of Liquid Formulation and Methanol Extract of S. tuirus AR26
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Osuna, G.; Jorge, A.; Wall, M.M.; Waddell, C.A. Endogenous levels of tocopherols and ascorbic acid during fruit ripening of New Mexican-type chile (Capsicum annuum L.) cultivars. J. Agric. Food Chem. 1998, 46, 5093–5096. [Google Scholar] [CrossRef]
- Than, P.; Prihastuti, H.; Phoulivong, S.; Taylor, P.W.; Hyde, K.D. Chilli anthracnose disease caused by Colletotrichum species. J. Zhejiang Univ. Sci. B 2008, 9, 764–778. [Google Scholar] [CrossRef]
- Dwivedi, N.; Tirkey, D.S.; Katoch, S.; Prasad, L. Evaluation of resistance against anthracnose (Colletotrichum capsici and C. gloeosporioides) in chilli landraces collected from the northeastern region of India. Plant Genet. Resour. 2021, 19, 538–544. [Google Scholar] [CrossRef]
- Machenahalli, S.; Nargund, V.; Patil, S. Quick detection and diagnosis of chilli fruit rot pathogens. Vegetos 2014, 27, 188–191. [Google Scholar] [CrossRef]
- Parey, M.A.; Razdan, V.; Sofi, T. Comparative study of different fungi associated with fruit rot of chilli and screening of chilli germplasm against Colletotrichum capsici. Int. J. Agric. Crop Sci. 2013, 5, 723. [Google Scholar]
- Liu, F.; Tang, G.; Zheng, X.; Li, Y.; Sun, X.; Qi, X.; Chang, X. Molecular and phenotypic characterization of Colletotrichum species associated with anthracnose disease in peppers from Sichuan Province, China. Sci. Rep. 2016, 6, 32761. [Google Scholar] [CrossRef]
- Machenahalli, S.; Nargund, V.; Byadgi, A.; Hegde, Y. Integrated management of die-back and fruit rot of Chilli (Capsicum annuum L.). Vegetos 2016, 29, 1000122. [Google Scholar] [CrossRef]
- Garg, R.; Loganathan, M.; Saha, S.; Roy, B. Chilli Anthracnose: A review of causal organism, resistance source and mapping of gene. In Microbial Diversity and Biotechnology in Food Security; Springer: New Delhi, India, 2014; pp. 589–610. [Google Scholar]
- Maharaj, A.; Rampersad, S.N. Genetic differentiation of Colletotrichum gloeosporioides and C. truncatum associated with anthracnose disease of papaya (Carica papaya L.) and bell pepper (Capsium annuum L.) based on ITS PCR-RFLP fingerprinting. Mol. Biotechnol. 2012, 50, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Raghuwanshi, R.; Gupta, V.K.; Singh, H.B. Chilli anthracnose: The epidemiology and management. Front. Microbiol. 2016, 7, 1527. [Google Scholar] [CrossRef]
- Chanchaichaovivat, A.; Panijpan, B.; Ruenwongsa, P. Putative modes of action of Pichia guilliermondii strain R13 in controlling chilli anthracnose after harvest. Biol. Control 2008, 47, 207–215. [Google Scholar] [CrossRef]
- Chinthagunta, L.; Zachariah, S. Evaluation of biological, chemical and IDM modules for the management of anthracnose and die-back of chilli. J. Mycol. Plant Pathol. 2012, 42, 435–438. [Google Scholar]
- Unnikrishnan, V.; Nath, B. Hazardous chemicals in foods. Indian J. Dairy Biosci. 2000, 11, 155–158. [Google Scholar]
- Boukaew, S.; Petlamul, W.; Bunkrongcheap, R.; Chookaew, T.; Kabbua, T.; Thippated, A.; Prasertsan, P. Fumigant activity of volatile compounds of Streptomyces philanthi RM-1-138 and pure chemicals (acetophenone and phenylethyl alcohol) against anthracnose pathogen in postharvest chili fruit. Crop Prot. 2018, 103, 1–8. [Google Scholar] [CrossRef]
- Ruangwong, O.-U.; Pornsuriya, C.; Pitija, K.; Sunpapao, A. Biocontrol mechanisms of Trichoderma koningiopsis PSU3-2 against postharvest anthracnose of chili pepper. J. Fungi 2021, 7, 276. [Google Scholar] [CrossRef]
- Tripathi, P.; Dubey, N. Exploitation of natural products as an alternative strategy to control postharvest fungal rotting of fruit and vegetables. Postharvest Biol. Technol. 2004, 32, 235–245. [Google Scholar] [CrossRef]
- Gowtham, H.; Murali, M.; Singh, S.B.; Lakshmeesha, T.; Murthy, K.N.; Amruthesh, K.; Niranjana, S. Plant growth promoting rhizobacteria-Bacillus amyloliquefaciens improves plant growth and induces resistance in chilli against anthracnose disease. Biol. Control 2018, 126, 209–217. [Google Scholar] [CrossRef]
- Heng, J.L.S.; Shah, U.K.; Rahman, N.A.A.; Shaari, K.; Hamzah, H. Streptomyces ambofaciens S2—A potential biological control agent for Colletotrichum gleosporioides the causal agent for anthracnose in red chilli fruits. J. Plant Pathol. Microbiol. 2015, S1, 6. [Google Scholar] [CrossRef]
- Ebrahimi-Zarandi, M.; Bonjar, G.H.S.; Riseh, R.S.; El-Shetehy, M.; Saadoun, I.; Barka, E.A. Exploring two Streptomyces species to control Rhizoctonia solani in tomato. Agronomy 2021, 11, 1384. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.; Qiao, X.; Li, Z.; Li, F.; Chen, M.; Cui, H. Antifungal activity of volatile organic compounds from Streptomyces alboflavus TD-1. FEMS Microbiol. Lett. 2013, 341, 45–51. [Google Scholar] [CrossRef]
- Wu, P.-H.; Tsay, T.-T.; Chen, P. Evaluation of Streptomyces saraciticas as Soil Amendments for Controlling Soil-Borne Plant Pathogens. Plant Pathol. J. 2021, 37, 596. [Google Scholar] [CrossRef]
- Doumbou, C.L.; Hamby, S.; Crawford, D.; Beaulieu, C. Actinomycetes, promising tools to control plant diseases and to promote plant growth. Phytoprotection 2001, 82, 85–102. [Google Scholar] [CrossRef]
- Doumbou, C.; Akimov, V.; Cote, M.; Charest, P.M.; Beaulieu, C. Taxonomic study on nonpathogenic streptomycetes isolated from common scab lesions on potato tubers. Syst. Appl. Microbiol. 2001, 24, 451–456. [Google Scholar] [CrossRef]
- Eccleston, K.L.; Brooks, P.R.; Kurtböke, D.İ. Assessment of the role of local strawberry rhizosphere—Associated Streptomycetes on the bacterially—Induced growth and Botrytis cinerea infection resistance of the fruit. Sustainability 2010, 2, 3831–3845. [Google Scholar] [CrossRef] [Green Version]
- El-Tarabily, K.A.; Sivasithamparam, K. Non-streptomycete actinomycetes as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters. Soil Biol. Biochem. 2006, 38, 1505–1520. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Srinivas, V.; Prasanna, S. Streptomyces . In Beneficial Microbes in Agro-Ecology; Academic Press: Cambridge, MA, USA, 2020; pp. 55–71. [Google Scholar]
- Hong-Thao, P.T.; Mai-Linh, N.V.; Hong-Lien, N.T.; Van Hieu, N. Biological characteristics and antimicrobial activity of endophytic Streptomyces sp. TQR12-4 isolated from elite Citrus nobilis cultivar Ham Yen of Vietnam. Int. J. Microbiol. 2016, 2016, 7207818. [Google Scholar] [CrossRef]
- Jorjandi, M.; Bonjar, G.S.; Baghizadeh, A.; Sirchi, G.S.; Massumi, H.; Baniasadi, F.; Farokhi, P.R. Biocontrol of Botrytis allii Munn the causal agent of neck rot, the post harvest disease in onion, by use of a new Iranian isolate of Streptomyces. Am. J. Agric. Biol. Sci. 2009, 4, 72–78. [Google Scholar] [CrossRef]
- Taechowisan, T.; Chuaychot, N.; Chanaphat, S.; Wanbanjob, A.; Tantiwachwutikul, P. Antagonistic effects of Streptomyces sp. SRM1 on Colletotrichum musae. Biotechnology 2009, 8, 86–92. [Google Scholar] [CrossRef]
- Thilagam, R.; Hemalatha, N. Plant growth promotion and chilli anthracnose disease suppression ability of rhizosphere soil actinobacteria. J. Appl. Microbiol. 2019, 126, 1835–1849. [Google Scholar] [CrossRef]
- Evangelista-Martnnez, Z. Isolation and characterization of soil Streptomyces species as potential biological control agents against fungal plant pathogens. World J. Microbiol. Biotechnol. 2014, 30, 1639–1647. [Google Scholar] [CrossRef]
- Manikandan, A.; Johnson, I.; Jaivel, N.; Krishnamoorthy, R.; SenthilKumar, M.; Raghu, R.; Gopa, N.O.; Mukherjee, P.K.; Anandham, R. Gamma-induced mutants of Bacillus and Streptomyces display enhanced antagonistic activities and suppression of the root rot and wilt diseases in pulses. Biomol. Concepts 2022, 13, 103–118. [Google Scholar] [CrossRef]
- Shanthi, V. Actinomycetes: Implications and prospects in sustainable agriculture. In Biofertilizers: Study and Impact; Wiley: Hoboken, NJ, USA, 2021; pp. 335–370. [Google Scholar]
- Cordero, I.; Balaguer, L.; Rincon, A.; Pueyo, J.J. Inoculation of tomato plants with selected PGPR represents a feasible alternative to chemical fertilization under salt stress. J. Plant Nutr. Soil Sci. 2018, 181, 694–703. [Google Scholar] [CrossRef]
- Vimal, S.R.; Singh, J.S.; Arora, N.K.; Singh, S. Soil-plant-microbe interactions in stressed agriculture management: A review. Pedosphere 2017, 27, 177–192. [Google Scholar] [CrossRef]
- Pappachan, A.; Rahul, K.; Irene, L.; Sivaprasad, V. Molecular identification of fungi associated with mulberry root rot disease in Eastern and North Eastern India. J. Crop Weed 2020, 16, 180–185. [Google Scholar] [CrossRef]
- Anwar, S.; Ali, B.; Sajid, I. Screening of rhizospheric actinomycetes for various in-vitro and in-vivo plant growth promoting (PGP) traits and for agroactive compounds. Front. Microbiol. 2016, 7, 1334. [Google Scholar] [CrossRef] [PubMed]
- Saadoun, I.; Hameed, K.; Moussauui, A. Characterization and analysis of antibiotic activity of some aquatic actinomycetes. Microbios 1999, 99, 173–179. [Google Scholar]
- Kurtboke, D.I. Selective Isolation of Rare Actinomycetes; Queensland Complete Printing Services: Nambour, Australia, 2003. [Google Scholar]
- Taechowisan, T.; Peberdy, J.; Lumyong, S. Chitinase production by endophytic Streptomyces aureofaciens CMUAc130 and its antagonism against phytopathogenic fungi. Ann. Microbiol. 2003, 53, 447–462. [Google Scholar]
- Harsonowati, W.; Astuti, R.I.; Wahyudi, A.T. Leaf blast disease reduction by rice-phyllosphere actinomycetes producing bioactive compounds. J. Gen. Plant Pathol. 2017, 83, 98–108. [Google Scholar] [CrossRef]
- Li, J.; Zhao, G.Z.; Huang, H.Y.; Qin, S.; Zhu, W.Y.; Zhao, L.X.; Xu, L.H.; Zhang, S.; Li, W.J.; Strobel, G. Isolation and characterization of culturable endophytic actinobacteria associated with Artemisia annua L. Antonie Van Leeuwenhoek 2012, 101, 515–527. [Google Scholar] [CrossRef]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; John Innes Foundation: Norwich, UK, 2000; Volume 291. [Google Scholar]
- Khamna, S.; Yokota, A.; Lumyong, S. Actinomycetes isolated from medicinal plant rhizosphere soils: Diversity and screening of antifungal compounds, indole-3-acetic acid and siderophore production. World J. Microbiol. Biotechnol. 2009, 25, 649–655. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R.; Yandigeri, M.S. In vitro biocontrol activity of halotolerant Streptomyces aureofaciens K20: A potent antagonist against Macrophomina phaseolina (Tassi) Goid. Saudi J. Biol. Sci. 2017, 24, 192–199. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hwang, B.K. Diversity of antifungal actinomycetes in various vegetative soils of Korea. Can. J. Microbiol. 2002, 48, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Liotti, R.G.; da Silva Figueiredo, M.I.; Soares, M.A. Streptomyces griseocarneus R132 controls phytopathogens and promotes growth of pepper (Capsicum annuum). Biol. Control 2019, 138, 104065. [Google Scholar] [CrossRef]
- Aneja, K. Experiments in Microbiology, Plant Pathology and Biotechnology; New Age International: New Delhi, India, 2007. [Google Scholar]
- Hankin, L.; Anagnostakis, S.L. Solid media containing carboxymethylcellulose to detect Cx cellulase activity of micro-organisms. Microbiology 1977, 98, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Sadeghian, M.; Bonjar, G.H.S.; Sirchi, G.R.S. Post harvest biological control of apple bitter rot by soil-borne actinomycetes and molecular identification of the active antagonist. Postharvest Biol. Technol. 2016, 112, 46–54. [Google Scholar] [CrossRef]
- Shah, D.; Soni, A. Isolation and screening of actinomycetes from mangrove soil for enzyme production and antimicrobial activity. Int. J. Innov. Sci. Res. 2016, 3, 54. [Google Scholar]
- Sadeghi, A.; Karimi, E.; Dahaji, P.A.; Javid, M.G.; Dalvand, Y.; Askari, H. Plant growth promoting activity of an auxin and siderophore producing isolate of Streptomyces under saline soil conditions. World J. Microbiol. Biotechnol. 2012, 28, 1503–1509. [Google Scholar] [CrossRef]
- Holding, A.; Collee, J. Chapter I Routine biochemical tests. In Methods in Microbiology; Elsevier: Amsterdam, The Netherlands, 1971; Volume 6, pp. 1–32. [Google Scholar]
- Gebily, D.A.; Ghanem, G.A.; Ragab, M.M.; Ali, A.M.; Soliman, N.E.; El-Moity, A.; Tawfik, H. Characterization and potential antifungal activities of three Streptomyces spp. as biocontrol agents against Sclerotinia sclerotiorum (Lib.) de Bary infecting green bean. Egypt. J. Biol. Pest Control 2021, 31, 33. [Google Scholar] [CrossRef]
- Meena, M.; Swapnil, P.; Zehra, A.; Dubey, M.K.; Upadhyay, R. Antagonistic assessment of Trichoderma spp. by producing volatile and non-volatile compounds against different fungal pathogens. Arch. Phytopathol. Plant Prot. 2017, 50, 629–648. [Google Scholar] [CrossRef]
- Sridharan, A.; Thankappan, S.; Karthikeyan, G.; Uthandi, S. Comprehensive profiling of the VOCs of Trichoderma longibrachiatum EF5 while interacting with Sclerotium rolfsii and Macrophomina phaseolina. Microbiol. Res. 2020, 236, 126436. [Google Scholar]
- Passari, A.K.; Mishra, V.K.; Gupta, V.K.; Yadav, M.K.; Saikia, R.; Singh, B.P. In vitro and in vivo plant growth promoting activities and DNA fingerprinting of antagonistic endophytic actinomycetes associates with medicinal plants. PLoS ONE 2015, 10, e0139468. [Google Scholar] [CrossRef]
- El-Sayed, W.S.; Akhkha, A.; El-Naggar, M.Y.; Elbadry, M. In vitro antagonistic activity, plant growth promoting traits and phylogenetic affiliation of rhizobacteria associated with wild plants grown in arid soil. Front. Microbiol. 2014, 5, 651. [Google Scholar] [CrossRef] [PubMed]
- Saratha, M.; Angappan, K.; Karthikeyan, S.; Marimuthu, S.; Chozhan, K. Actinoalloteichus cyanogriseus: A broad spectrum bio-agent against mulberry root rot pathogens. Egypt. J. Biol. Pest Control 2022, 32, 33. [Google Scholar] [CrossRef]
- Samson, R.A.; Stalpers, J.A.; Verkerke, W. A simplified technique to prepare fungal specimens for scanning electron microscopy. Cytobios 1979, 24, 7–11. [Google Scholar] [PubMed]
- Inderiati, S.; Franco, C.M. Isolation and identification of endophytic actinomycetes and their antifungal activity. J. Biotechnol. Res. Trop. Reg. 2008, 1, 1–6. [Google Scholar]
- Gopalakrishnan, S.; Vadlamudi, S.; Bandikinda, P.; Sathya, A.; Vijayabharathi, R.; Rupela, O.; Varshney, R.K. Evaluation of Streptomyces strains isolated from herbal vermicompost for their plant growth-promotion traits in rice. Microbiol. Res. 2014, 169, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Cawoy, H.; Debois, D.; Franzil, L.; De Pauw, E.; Thonart, P.; Ongena, M. Lipopeptides as main ingredients for inhibition of fungal phytopathogens by Bacillus subtilis/amyloliquefaciens. Microb. Biotechnol. 2015, 8, 281–295. [Google Scholar] [CrossRef]
- Strobel, G.; Daisy, B.; Castillo, U.; Harper, J. Natural products from endophytic microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef]
- Sudha, A.; Durgadevi, D.; Archana, S.; Muthukumar, A.; Raj, T.S.; Nakkeeran, S.; Poczai, P.; Nasif, O.; Ansari, M.J.; Sayyed, R.Z. Unraveling the tripartite interaction of volatile compounds of Streptomyces rochei with grain mold pathogens infecting sorghum. Front. Microbiol. 2022, 13, 923360. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Vurukonda, S.S.K.P.; Giovanardi, D.; Stefani, E. Plant growth promoting and biocontrol activity of Streptomyces spp. as endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, P.; Musa, M.Y.; Tan, G.Y.A.; Avin, F.A.; Teo, W.F.A.; Sabaratnam, V. In vitro and in vivo evaluation of Streptomyces suppressions against anthracnose in chili caused by Colletotrichum. Sains Malays. 2014, 43, 697–705. [Google Scholar]
- Chater, K.F. Streptomyces inside-out: A new perspective on the bacteria that provide us with antibiotics. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.; Vickers, J. Detection of actinomycetes in the natural environment: Problems and perspectives. Biol. Actinomycetes 1988, 88, 265–270. [Google Scholar]
- Hyder, S.; Gondal, A.S.; Rizvi, Z.F.; Ahmad, R.; Alam, M.M.; Hannan, A.; Ahmed, W.; Fatima, N.; Inam-ul-Haq, M. Characterization of native plant growth promoting rhizobacteria and their anti-oomycete potential against Phytophthora capsici affecting chilli pepper (Capsicum annum L.). Sci. Rep. 2020, 10, 13859. [Google Scholar] [CrossRef]
- Chaudhry, M.J. Exploring the Biocontrol Potential of Streptomyces isolates Against Pythium violae, a Dominant Carrot Pathogen. Master’s Thesis, California State University, Bakersfield, CA, USA, 2021. [Google Scholar]
- Xu, T.; Cao, L.; Zeng, J.; Franco, C.M.; Yang, Y.; Hu, X.; Bu, Z. The antifungal action mode of the rice endophyte Streptomyces hygroscopicus OsiSh-2 as a potential biocontrol agent against the rice blast pathogen. Pestic. Biochem. Physiol. 2019, 160, 58–69. [Google Scholar] [CrossRef]
- Veliz, E.A.; Martínez-Hidalgo, P.; Hirsch, A.M. Chitinase-producing bacteria and their role in biocontrol. AIMS Microbiol. 2017, 3, 689. [Google Scholar] [CrossRef]
- Jha, S.; Modi, H. Comparative analysis of chitinase activity by four different assay from soil born Actinomycetes. In Proceedings of the 4th International Conference on Multidisciplinary Research and Practice (4ICMRP-2017), Ahmedabad, India, 22 December 2017. [Google Scholar]
- Bhattacharyya, C.; Banerjee, S.; Acharya, U.; Mitra, A.; Mallick, I.; Haldar, A.; Ghosh, A. Evaluation of plant growth promotion properties and induction of antioxidative defense mechanism by tea rhizobacteria of Darjeeling, India. Sci. Rep. 2020, 10, 15536. [Google Scholar] [CrossRef]
- Froes, A.; Macrae, A.; Rosa, J.; Franco, M.; Souza, R.; Soares, R.; Coelho, R. Selection of a Streptomyces strain able to produce cell wall degrading enzymes and active against Sclerotinia sclerotiorum. J. Microbiol. 2012, 50, 798–806. [Google Scholar] [CrossRef]
- Shanmugaiah, V.; Nithya, K.; Harikrishnan, H.; Jayaprakashvel, M.; Balasubramanian, N. Biocontrol mechanisms of siderophores against bacterial plant pathogens. In Sustainable Approaches to Controlling Plant Pathogenic Bacteria; CRC Press: Boca Raton, FL, USA, 2015; pp. 167–190. [Google Scholar]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Wattanasuepsin, W.; Intra, B.; Euanorasetr, J.; Watanabe, Y.; Mingma, R.; Fukasawa, W.; Panbangred, W. 1-Methoxypyrrole-2-carboxamide—A new pyrrole compound isolated from Streptomyces griseocarneus SWW368. J. Gen. Appl. Microbiol. 2017, 63, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Yazawa, S.; Ushijima, Y. A promising strain of endophytic Streptomyces sp. for biological control of cucumber anthracnose. J. Gen. Plant Pathol. 2009, 75, 27–36. [Google Scholar] [CrossRef]
- Ilic, S.; Konstantinovic, S.; Todorovic, Z.; Lazic, M.; Veljkovic, V.; Jokovic, N.; Radovanovic, B. Characterization and antimicrobial activity of the bioactive metabolites in streptomycete isolates. Microbiology 2007, 76, 421–428. [Google Scholar] [CrossRef]
- Rothrock, C.S.; Gottlieb, D. Role of antibiosis in antagonism of Streptomyces hygroscopicus var. geldanus to Rhizoctonia solani in soil. Can. J. Microbiol. 1984, 30, 1440–1447. [Google Scholar] [CrossRef]
- Evangelista-Martinez, Z.; Contreras-Leal, E.A.; Corona-Pedraza, L.F.; Gastelum-Martinez, É. Biocontrol potential of Streptomyces sp. CACIS-1.5 CA against phytopathogenic fungi causing postharvest fruit diseases. Egypt. J. Biol. Pest Control 2020, 30, 117. [Google Scholar] [CrossRef]
- Yasmin, S.; Zaka, A.; Imran, A.; Zahid, M.A.; Yousaf, S.; Rasul, G.; Mirza, M.S. Plant growth promotion and suppression of bacterial leaf blight in rice by inoculated bacteria. PLoS ONE 2016, 11, e0160688. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No | Isolate Code | Antagonistic Activity | Antifungal Mechanisms | Total Assessment Points Out of 24 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mycelial Growth Inhibition (%) | Halo Zone Diameter (cm) | ||||||||||||
| Dual Culture Assay | Paired Antibiosis Assay | ||||||||||||
| C.s | C.t | F.o | C.s | C.t | F.o | Siderophore | Amylase | Cellulase | Chitinase | Protease | |||
| 1. | AR1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 9 |
| 2. | AR10 | 2 | 2 | 1 | 1 | 2 | 2 | 2 | 1 | 1 | 1 | 0 | 15 |
| 3. | AR26 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 0 | 17 |
| 4. | AL5 | 1 | 1 | 1 | 2 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 10 |
| 5. | AL7 | 2 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 0 | 13 |
| 6. | AFE2 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 9 |
| S. No. | Isolate Name | Isolation Source | NCBI Accession Number | Base Pair Length | Closest 16S rRNA Sequence Match (BLASTN) | Per Cent Identity (%) | |
|---|---|---|---|---|---|---|---|
| Organism and Strain | Base Pair Length | ||||||
| 1. | AR1 | Rhizosphere, Pudukottai | OM883984 | 1358 bp | Streptomyces rochei AL14 | 1448 | 99.70 |
| 2. | AR10 | Rhizosphere, Karaikal | ON692910 | 1458 bp | Streptomyces deccanensis WJA64 | 1462 | 99.73 |
| 3. | AR26 | Rhizosphere, Salem | ON140212 | 1432 bp | Streptomyces tuirus PAS9 | 1461 | 99.72 |
| 4. | AL5 | Phyllosphere: Leaf, Coimbatore | ON692752 | 1486 bp | Streptomyces azureus NRRL B-2655 | 1516 | 100.00 |
| 5. | AL7 | Phyllosphere: Leaf, Trichy | ON692754 | 1466 bp | Streptomyces geysiriensis DSD176 | 1466 | 100.00 |
| S. No | Treatments/Pathogens | Lesion Diameter (cm) | Per cent Disease Reduction (%) |
|---|---|---|---|
| 1. | T1: Healthy (uninoculated) control | 0.00 | 100.00 a (89.71) |
| 2. | T2: Antagonist inoculated control (10 mL/L) | 0.00 | 100.00 a (89.71) |
| 3. | T3: Colletotrichum scovillei inoculated control | 2.48 | 0.00 c (0.29) |
| 4. | T4: Colletotrichum truncatum inoculated control | 2.18 | 0.00 c (0.29) |
| 5. | T5: Fusarium oxysporum inoculated control | 2.60 | 0.00 c (0.29) |
| 6. | T6: Co-inoculation of C. scovillei, C. capsici and F. oxysporum | 2.88 | 0.00 c (0.29) |
| 7. | T7: C. scovillei + S. tuirus (5 mL/L) | 0.73 | 70.85 d (57.34) |
| 8. | T8: C. scovillei + S. tuirus (10 mL/L) | 0.30 | 90.32 b (80.17) |
| 9. | T9: C. truncatum + S. tuirus (5 mL/L) | 0.38 | 82.68 c (65.75) |
| 10. | T10: C. truncatum + S. tuirus (10 mL/L) | 0.00 | 100.00 a (89.71) |
| 11. | T11: F. oxysporum + S. tuirus (5 mL/L) | 0.85 | 67.32 d (55.41) |
| 12. | T12: F. oxysporum + S. tuirus (10 mL/L) | 0.00 | 100.00 a (89.71) |
| 13. | T13: Co-inoculation of C. scovillei, C. capsici and F. oxysporum + S. tuirus (5 mL/L) | 0.63 | 77.08 c,d (61.53) |
| 14. | T14: Co-inoculation of C. scovillei, C. capsici and F. oxysporum + S. tuirus (10 mL/L) | 0.00 | 100.00 a (89.71) |
| CD (0.05) | 0.167 | 8.040 | |
| SE (d) | 0.082 | 3.960 |
| S. No | Treatments/Pathogens | Lesion Diameter (cm) | Per cent Disease Reduction (%) |
|---|---|---|---|
| 1. | T1: Healthy (uninoculated) control | 0.0 | 100.00 a (89.71) |
| 2. | T2: Methanol extract inoculated control | 0.0 | 100.00 a (89.71) |
| 3. | T3: C. scovillei inoculated control | 2.93 | 0.00 f (0.29) |
| 4. | T4: C. truncatum inoculated control | 2.50 | 0.00 f (0.29) |
| 5. | T5: F. oxysporum inoculated control | 2.35 | 0.00 f (0.29) |
| 6. | T6: Co-inoculation of C. scovillei, C. capsici and F. oxysporum | 3.40 | 0.00 f (0.29) |
| 7. | T7: C. scovillei + Methanol extract of S. tuirus | 1.10 | 62.45 C (52.22) |
| 8. | T8: C. truncatum + Methanol extract of S. tuirus | 0.75 | 70.10 b (56.99) |
| 9. | T9: F. oxysporum + Methanol extract of S. tuirus | 1.10 | 53.08 d (46.79) |
| 10. | T10: Co-inoculation of C. scovillei, C. capsici and F. oxysporum + methanol extract of S. tuirus | 1.88 | 44.85 e (46.04) |
| CD (0.05) | 0.326 | 3.129 | |
| SE (d) | 0.159 | 1.525 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renuka, R.; Prabakar, K.; Anandham, R.; Pugalendhi, L.; Rajendran, L.; Raguchander, T.; Karthikeyan, G. Exploring the Potentiality of Native Actinobacteria to Combat the Chilli Fruit Rot Pathogens under Post-Harvest Pathosystem. Life 2023, 13, 426. https://doi.org/10.3390/life13020426
Renuka R, Prabakar K, Anandham R, Pugalendhi L, Rajendran L, Raguchander T, Karthikeyan G. Exploring the Potentiality of Native Actinobacteria to Combat the Chilli Fruit Rot Pathogens under Post-Harvest Pathosystem. Life. 2023; 13(2):426. https://doi.org/10.3390/life13020426
Chicago/Turabian StyleRenuka, Rajamuthu, Kupusamy Prabakar, Rangasamy Anandham, Lakshmanan Pugalendhi, Lingam Rajendran, Thiruvengadam Raguchander, and Gandhi Karthikeyan. 2023. "Exploring the Potentiality of Native Actinobacteria to Combat the Chilli Fruit Rot Pathogens under Post-Harvest Pathosystem" Life 13, no. 2: 426. https://doi.org/10.3390/life13020426