
Nematode-Induced Growth Factors Related to Angiogenesis in Autoimmune Disease Attenuation
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Colitis Model
2.2. Burden of Parasitic Infection
2.3. Preparation of Blood, Small Intestine and Colon Mucosa Samples
2.4. ELISA
2.5. EAE Model
2.6. Cerebral Spinal Fluid (CSF)
2.7. Visualization of the Vascular Endothelium
2.8. Antibody Arrays
2.9. Statistical Analysis
3. Results
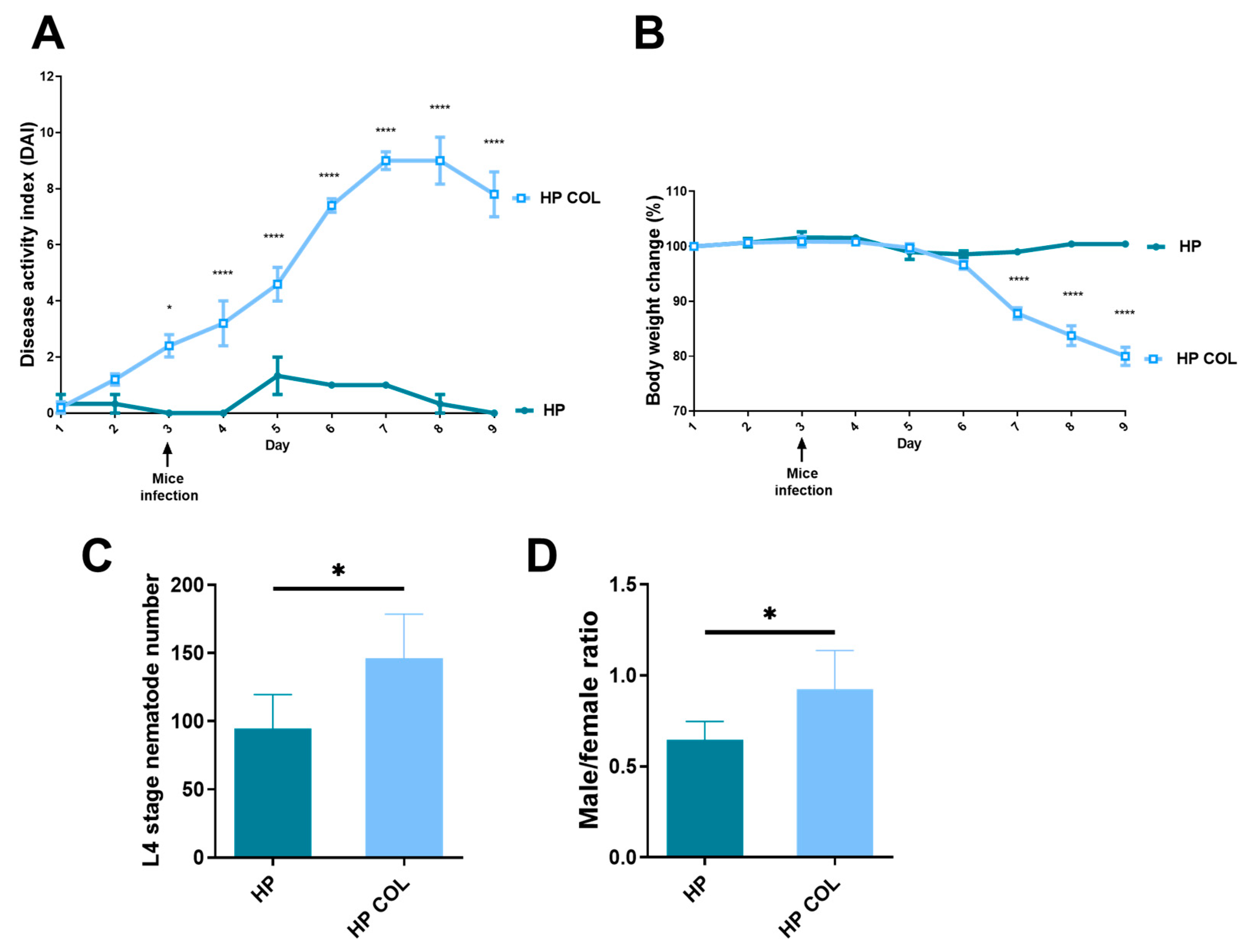
3.1. Symptoms of Colitis
3.2. Nematodes Adapt to Colitis in C57BL/6 Mice
3.3. Nematode Adaptation to Colitis Correlate with Changes in Immune Response
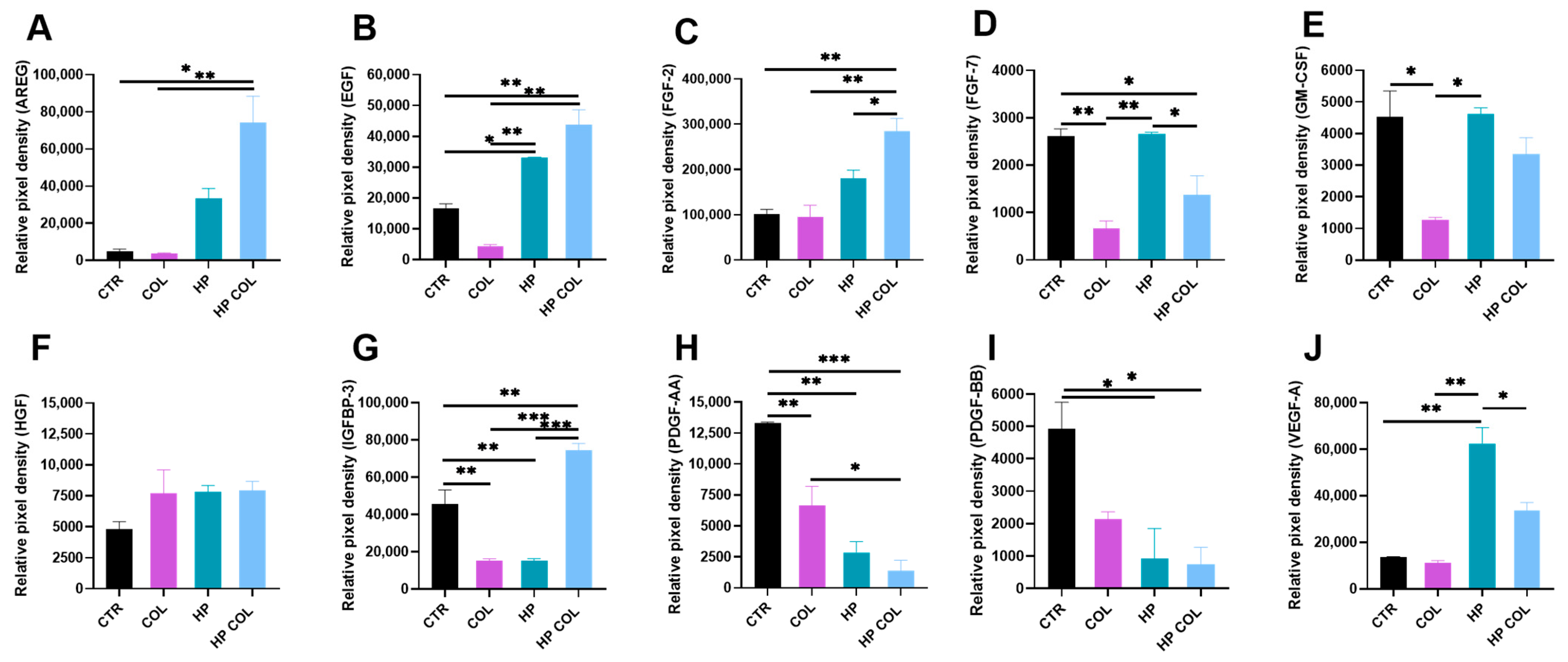
3.4. Colitis Induction and Parasitic Infection Influence Level of Angiogenic Factors in the Intestine
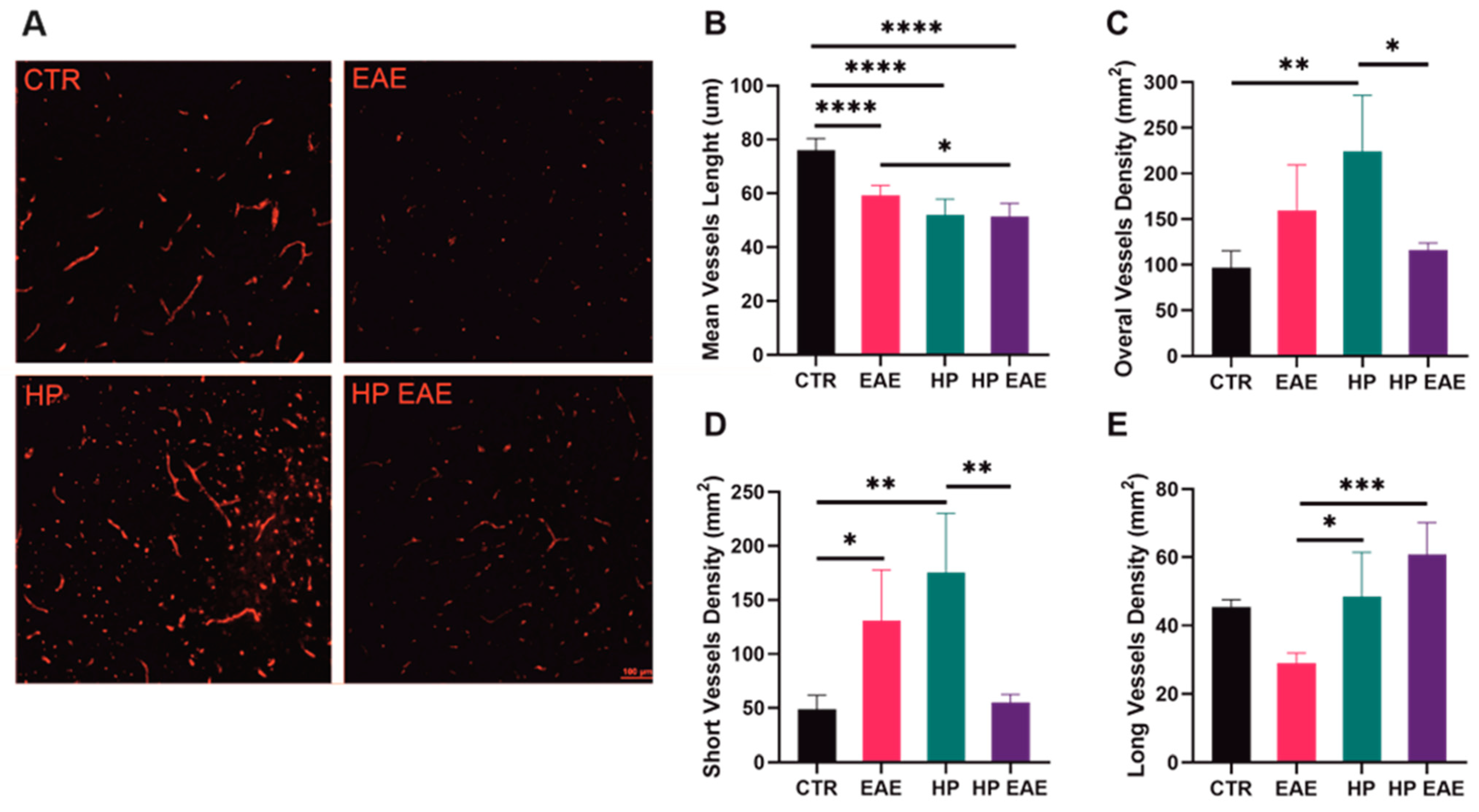
3.5. Nematode Infection Influences Angiogenesis in the Brain
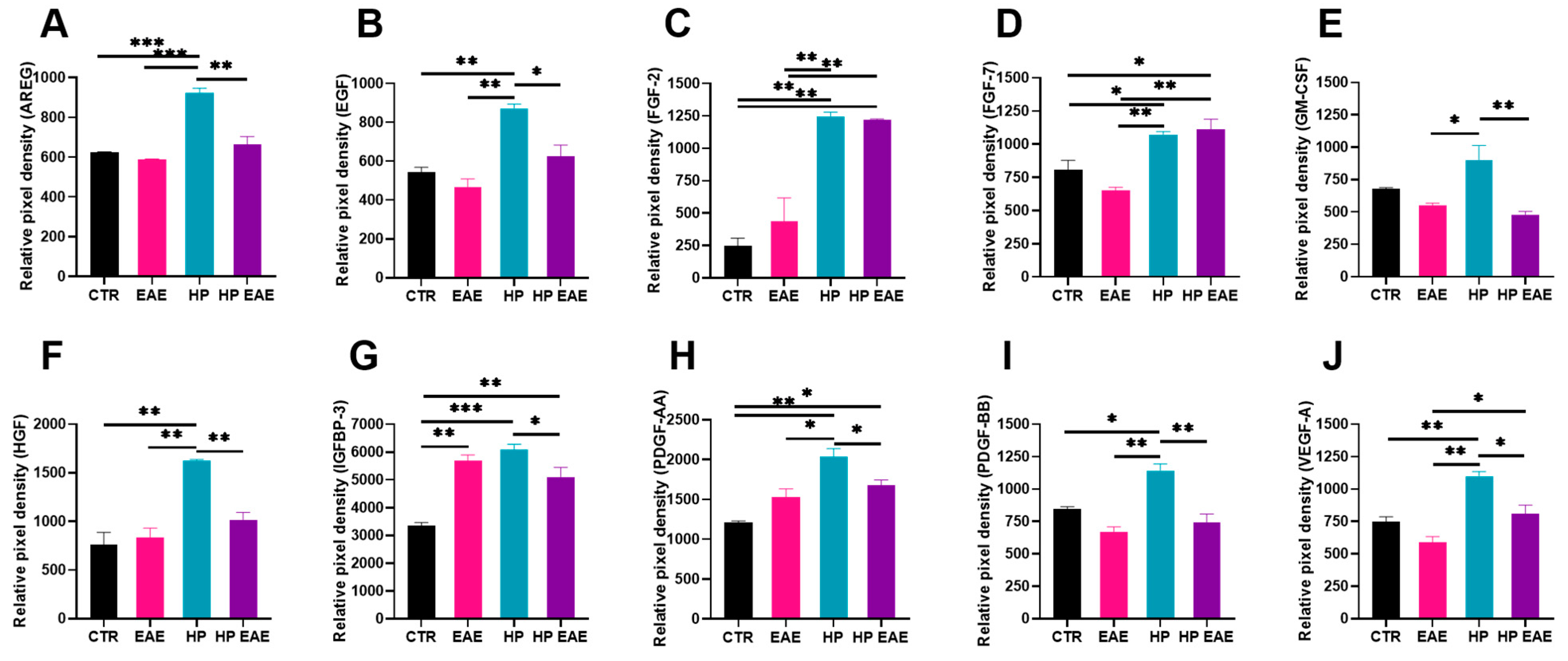
3.6. EAE Induction and Parasitic Infection Influence Level of Angiogenic Factors in the Brain
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhan, H.; Li, H.; Liu, C.; Cheng, L.; Yan, S.; Li, Y. Association of circulating vascular endothelial growth factor levels with Autoimmune diseases: A systematic review and meta-analysis. Front. Immunol. 2021, 27, 674343. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.; Alitalo, K. Molecular regulation of angiogenesis and lymphangiogenesis. Nat. Rev. Mol. Cell Biol. 2007, 8, 464–478. [Google Scholar] [CrossRef]
- Ucuzian, A.A.; Gassman, A.A.; East, A.T.; Greisler, H.P. Molecular mediators of angiogenesis. J. Burn. Care Res. 2010, 31, 158–175. [Google Scholar] [CrossRef]
- Sobotková, K.; Parker, W.; Levá, J.; Růžková, J.; Lukeš, J.; Jirků Pomajbíková, K. Helminth Therapy—From the Parasite Perspective. Trends Parasitol. 2019, 35, 501–515. [Google Scholar] [CrossRef]
- Maruszewska-Cheruiyot, M.; Donskow-Łysoniewska, K.; Doligalska, M. Helminth Therapy: Advances in the use of Parasitic Worms Against Inflammatory Bowel Diseases and its Challenges. Helminthologia 2018, 27, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Shields, V.; Cooper, J. Use of helminth therapy for management of ulcerative colitis and Crohn’s disease: A systematic review. Parasitology 2022, 149, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Fleming, J.O. Helminth therapy and multiple sclerosis. Int. J. Parasitol. 2013, 43, 259–274. [Google Scholar] [CrossRef]
- Gazzinelli-Guimaraes, P.H.; Nutman, T.B. Helminth parasites and immune regulation. F1000Research 2018, 7, 1685. [Google Scholar] [CrossRef] [Green Version]
- Dennis, R.D.; Schubert, U.; Bauer, C. Angiogenesis and parasitic helminth-associated neovascularization. Parasitology 2011, 138, 426–439. [Google Scholar] [CrossRef]
- Wammes, L.J.; Mpairwe, H.; Elliott, A.M.; Yazdanbakhsh, M. Helminth therapy or elimination: Epidemiological, immunological, and clinical considerations. Lancet Infect. Dis. 2014, 14, 1150–1162. [Google Scholar] [CrossRef]
- Maruszewska-Cheruiyot, M.; Donskow-Łysoniewska, K.; Krawczak, K.; Machcińska, M.; Doligalska, M. Immunomodulatory potential of nematodes against dendritic cells is dependent on intestinal inflammation. Dev. Comp. Immunol. 2021, 115, 103879. [Google Scholar] [CrossRef]
- Donskow-Łysoniewska, K.; Krawczak, K.; Doligalska, M. Heligmosomoides polygyrus: EAE remission is correlated with different systemic cytokine profiles provoked by L4 and adult nematodes. Exp. Parasitol. 2012, 132, 243–248. [Google Scholar] [CrossRef]
- Donskow-Łysoniewska, K.; Majewski, P.; Brodaczewska, K.; Jóźwicka, K.; Doligalska, M. Heligmosmoides polygyrus fourth stages induce protection against DSS-induced colitis and change opioid expression in the intestine. Parasite Immunol. 2012, 34, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Melgar, S.; Karlsson, A.; Michaëlsson, E. Acute colitis induced by dextran sulfate sodium progresses to chronicity in C57BL/6 but not in BALB/c mice: Correlation between symptoms and inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G1328–G1338. [Google Scholar] [CrossRef] [PubMed]
- Donskow-Łysoniewska, K.; Bien, J.; Brodaczewska, K.; Krawczak, K.; Doligalska, M. Colitis promotes adaptation of an intestinal nematode: A Heligmosomoides polygyrus mouse model system. PLoS ONE 2013, 22, e78034. [Google Scholar] [CrossRef] [PubMed]
- Cross, M.; Dexter, T.M. Growth factors in development, transformation, and tumorigenesis. Cell 1991, 25, 271–280. [Google Scholar] [CrossRef]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.E.; Sutherland, T.E. Host protective roles of type 2 immunity: Parasite killing and tissue repair, flip sides of the same coin. Semin. Immunol. 2014, 26, 329–340. [Google Scholar] [CrossRef]
- Maruszewska-Cheruiyot, M.; Stear, M.J.; Machcińska, M.; Donskow-Łysoniewska, K. Importance of TGFβ in Cancer and Nematode Infection and Their Interaction-Opinion. Biomolecules 2022, 26, 1572. [Google Scholar] [CrossRef]
- Doligalska, M.; Rzepecka, J.; Drela, N.; Donskow, K.; Gerwel-Wronka, M. The role of TGF-beta in mice infected with Heligmosomoides polygyrus. Parasite Immunol. 2006, 28, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Coakley, G.; Buck, A.H.; Maizels, R.M. Host parasite communications—Messages from helminths for the immune system: Parasite communication and cell-cell interactions. Mol. Biochem. Parasitol. 2016, 208, 33–40. [Google Scholar] [CrossRef]
- King, I.L.; Li, Y. Host-parasite interactions promote disease tolerance to intestinal helminth infection. Front. Immunol. 2018, 9, 2128. [Google Scholar] [CrossRef] [Green Version]
- Szekanecz, Z.; Koch, A.E. Mechanisms of Disease: Angiogenesis in inflammatory diseases. Nat. Clin. Pract. Rheumatol. 2007, 3, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Girolamo, F.; Coppola, C.; Ribatti, D.; Trojano, M. Angiogenesis in multiple sclerosis and experimental autoimmune encephalomyelitis. Acta Neuropathol. Commun. 2014, 2, 84. [Google Scholar] [CrossRef] [PubMed]
- Alkim, C.; Alkim, H.; Koksal, A.R.; Boga, S.; Sen, I. Angiogenesis in Inflammatory Bowel Disease. Int. J. Inflam. 2015, 2015, 970890. [Google Scholar] [CrossRef] [Green Version]
- Tas, S.W.; Maracle, C.X.; Balogh, E.; Szekanecz, Z. Targeting of proangiogenic signalling pathways in chronic inflammation. Nat. Rev. Rheumatol. 2016, 12, 111–122. [Google Scholar] [CrossRef]
- Tham, E.; Gielen, A.W.; Khademi, M.; Martin, C.; Piehl, F. Decreased expression of VEGF-A in rat experimental autoimmune encephalomyelitis and in cerebrospinal fluid mononuclear cells from patients with multiple sclerosis. Scand. J. Immunol. 2006, 64, 609–622. [Google Scholar] [CrossRef]
- Mayhan, W.G. VEGF increases permeability of the blood-brain barrier via a nitric oxide synthase/cGMP-dependent pathway. Am. J. Physiol. 1999, 276, C1148–C1153. [Google Scholar] [CrossRef]
- Argaw, A.T.; Asp, L.; Zhang, J.; Navrazhina, K.; Pham, T.; Mariani, J.N.; Mahase, S.; Dutta, D.J.; Seto, J.; Kramer, E.G.; et al. Astrocyte-derived VEGF-A drives blood–brain barrier disruption in CNS inflammatory disease. J. Clin. Investig. 2012, 122, 2454–2468. [Google Scholar] [CrossRef]
- Quintana, F.J.; Cohen, I.R. Angiogenesis-inflammation cross-talk: Vascular endothelial growth factor is secreted by activated T cells and induces Th1 polarization. J. Immunol. 2004, 172, 4618–4623. [Google Scholar] [CrossRef] [Green Version]
- Scaldaferri, F.; Vetrano, S.; Sans, M.; Arena, V.; Straface, G.; Stigliano, E.; Repici, A.; Sturm, A.; Malesci, A.; Panes, J. VEGF-A links angiogenesis and inflammation in inflammatory bowel disease pathogenesis. Gastroenterology 2009, 136, 585–595.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeffler, D.A.; Lundy, S.K.; Singh, K.P.; Gerard, H.C.; Hudson, A.P.; Boros, D.L. Soluble egg antigens from Schistosoma mansoni induce angiogenesis-related processes by up-regulating vascular endothelial growth factor in human endothelial cells. J. Infect. Dis. 2002, 185, 1650–1656. [Google Scholar] [CrossRef] [Green Version]
- Capó, V.A.; Despommier, D.D.; Polvere, R.I. Trichinella spiralis: Vascular endothelial growth factor is up-regulated within the nurse cell during the early phase of its formation. J. Parasitol. 1998, 84, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Tarsitano, M.; De Falco, S.; Colonna, V.; McGhee, J.D.; Persico, M.G. The C. elegans pvf-1 gene encodes a PDGF/VEGF-like factor able to bind mammalian VEGF receptors and to induce angiogenesis. FASEB J. 2006, 20, 227–233. [Google Scholar] [CrossRef]
- Hirata, A.; Ogawa, S.; Kometani, T.; Kuwano, T.; Naito, S.; Kuwano, M.; Ono, M. ZD1839 (Iressa) induces antiangiogenic effects through inhibition of epidermal growth factor receptor tyrosine kinase. Cancer Res. 2002, 62, 2554–2560. [Google Scholar]
- Zaiss, D.M.W.; Gause, W.C.; Osborne, L.C.; Artis, D. Emerging functions of amphiregulin in orchestrating immunity, inflammation, and tissue repair. Immunity 2015, 42, 216–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaiss, D.M.; Yang, L.; Shah, P.R.; Kobie, J.J.; Urban, J.F.; Mosmann, T.R. Amphiregulin, a TH2 cytokine enhancing resistance to nematodes. Science 2006, 15, 1746. [Google Scholar] [CrossRef]
- Minutti, C.M.; Drube, S.; Blair, N.; Schwartz, C.; McCrae, J.C.; McKenzie, A.N.; Kamradt, T.; Mokry, M.; Coffer, P.J.; Sibilia, M.; et al. Epidermal Growth Factor Receptor Expression Licenses Type-2 Helper T Cells to Function in a T Cell Receptor-Independent Fashion. Immunity 2017, 17, 710–722.e6. [Google Scholar] [CrossRef] [Green Version]
- Zaiss, D.M.; van Loosdregt, J.; Gorlani, A.; Bekker, C.P.; Gröne, A.; Sibilia, M.; van Bergen en Henegouwen, P.M.; Roovers, R.C.; Coffer, P.J.; Sijts, A.J. Amphiregulin Enhances Regulatory T Cell-Suppressive Function via the Epidermal Growth Factor Receptor. Immunity 2013, 38, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Lopez Corcino, Y.; Gonzalez Ferrer, S.; Mantilla, L.E.; Trikeriotis, S.; Yu, J.S.; Kim, S.; Hansen, S.; Portillo, J.C.; Subauste, C.S. Toxoplasma gondii induces prolonged host epidermal growth factor receptor signalling to prevent parasite elimination by autophagy: Perspectives for in vivo control of the parasite. Cell. Microbiol. 2019, 21, e13084. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N.; Ornitz, D.M. Functional evolutionary history of the mouse Fgf gene family. Dev. Dyn. 2008, 237, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Okada-Ban, M.; Thiery, J.P.; Jouanneau, J. Fibroblast growth factor-2. Int. J. Biochem. Cell Biol. 2000, 32, 263–267. [Google Scholar] [CrossRef]
- Klimaschewski, L.; Claus, P. Fibroblast Growth Factor Signalling in the Diseased Nervous System. Mol. Neurobiol. 2021, 58, 3884–3902. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Dai, D.; He, X.; Zhu, S.; Yao, Y.; Gao, H.; Wang, J.; Qu, F.; Qiu, J.; Wang, H.; et al. Growth Factor FGF2 Cooperates with Interleukin-17 to Repair Intestinal Epithelial Damage. Immunity 2015, 15, 488–501. [Google Scholar] [CrossRef] [Green Version]
- Egger, B.; Procaccino, F.; Sarosi, I.; Tolmos, J.; Büchler, M.W.; Eysselein, V.E. Keratinocyte growth factor ameliorates dextran sodium sulfate colitis in mice. Dig. Dis. Sci. 1999, 44, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Förster, S.; Koziol, U.; Schäfer, T.; Duvoisin, R.; Cailliau, K.; Vanderstraete, M.; Dissous, C.; Brehm, K. The role of fibroblast growth factor signalling in Echinococcus multilocularis development and host-parasite interaction. PLoS Negl. Trop. Dis. 2019, 8, e0006959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eubank, T.D.; Roberts, R.; Galloway, M.; Wang, Y.; Cohn, D.E.; Marsh, C.B. GM-CSF induces expression of soluble VEGF receptor-1 from human monocytes and inhibits angiogenesis in mice. Immunity 2004, 21, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Lotfi, N.; Thome, R.; Rezaei, N.; Zhang, G.X.; Rezaei, A.; Rostami, A.; Esmaeil, N. Roles of GM-CSF in the Pathogenesis of Autoimmune Diseases: An Update. Front. Immunol. 2019, 4, 1265. [Google Scholar] [CrossRef]
- Shim, D.S.; Schilter, H.C.; Knott, M.L.; Almeida, R.A.; Short, R.P.; Mackay, C.R.; Dent, L.A.; Sewell, W.A. Protection against Nippostrongylus brasiliensis infection in mice is independent of GM-CSF. Immunol. Cell Biol. 2012, 90, 553–558. [Google Scholar] [CrossRef]
- Xin, X.; Yang, S.; Ingle, G.; Zlot, C.; Rangell, L.; Kowalski, J.; Schwall, R.; Ferrara, N.; Gerritsen, M.E. Hepatocyte growth factor enhances vascular endothelial growth factor-induced angiogenesis in vitro and in vivo. Am. J. Pathol. 2001, 158, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Varma Shrivastav, S.; Bhardwaj, A.; Pathak, K.A.; Shrivastav, A. Insulin-Like Growth Factor Binding Protein-3 (IGFBP-3): Unraveling the Role in Mediating IGF-Independent Effects Within the Cell. Front. Cell Dev. Biol. 2020, 5, 286. [Google Scholar] [CrossRef]
- Kim, S.C.; Hwang, P.H. Up-regulation of IGF Binding Protein-3 Inhibits Colonic Inflammatory Response. J. Korean Med. Sci. 2018, 26, e110. [Google Scholar] [CrossRef] [PubMed]
- Yancu, D.; Blouin, M.J.; Birman, E.; Florianova, L.; Aleynikova, O.; Zakikhani, M.; VanderMeulen, H.; Seidman, E.; Pollak, M. A phenotype of IGFBP-3 knockout mice revealed by dextran sulfate-induced colitis. J. Gastroenterol. Hepatol. 2017, 32, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumagai, S.; Ohtani, H.; Nagai, T.; Funa, K.; Hiwatashi, N.O.; Shimosegawa, T.; Nagura, H. Platelet-derived growth factor and its receptors are expressed in areas of both active inflammation and active fibrosis in inflammatory bowel disease. Tohoku J. Exp. Med. 2001, 195, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Stampanoni Bassi, M.; Iezzi, E.; Marfia, G.A.; Simonelli, I.; Musella, A.; Mandolesi, G.; Fresegna, D.; Pasqualetti, P.; Furlan, R.; Finardi, A.; et al. Platelet-derived growth factor predicts prolonged relapse-free period in multiple sclerosis. J. Neuroinflam. 2018, 15, 108. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokine | Serum | Colon | Small Intestine | |
|---|---|---|---|---|
| IL-1β | HP | 2.9 ± 1.1 | 154.5 ± 54 | 113.5 ± 64 |
| HP COL | 7.5 ± 7.5 | 2518 ± 638 * | 144.3 ± 38.4 | |
| IL-6 | HP | 1.2 ± 2.8 | 206.9 ± 66.2 | not detected |
| HP COL | 241 ± 363.1 * | 360.3 ± 89.7 * | 150 ± 120.1 * | |
| TNF-α | HP | 28.8 ± 4.5 | 148.5 ± 40.2 | 222.6 ± 144.8 |
| HP COL | 73.4 ± 22.2 * | 130.7 ± 32 | 921 ± 608.3 | |
| IL-10 | HP | 3685.1 ± 1829.8 | 8973.6 ± 3336.7 | 4854.2 ± 1362 |
| HP COL | 6735.6 ± 2101.2 * | 9833.1 ± 3740.8 | 5471.3 ± 2256.6 | |
| TGF-β | HP | 977.7 ± 418.4 | 19.7 ± 18.5 | 4655 ± 1267.7 |
| HP COL | 479.7 ± 167 | 38 ± 22.7 | 5099.9 ± 4049.9 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maruszewska-Cheruiyot, M.; Krawczak-Wójcik, K.; Michniowska, M.; Stear, M.J.; Machcińska, M.; Doligalska, M.; Donskow-Łysoniewska, K. Nematode-Induced Growth Factors Related to Angiogenesis in Autoimmune Disease Attenuation. Life 2023, 13, 321. https://doi.org/10.3390/life13020321
Maruszewska-Cheruiyot M, Krawczak-Wójcik K, Michniowska M, Stear MJ, Machcińska M, Doligalska M, Donskow-Łysoniewska K. Nematode-Induced Growth Factors Related to Angiogenesis in Autoimmune Disease Attenuation. Life. 2023; 13(2):321. https://doi.org/10.3390/life13020321
Chicago/Turabian StyleMaruszewska-Cheruiyot, Marta, Katarzyna Krawczak-Wójcik, Martyna Michniowska, Michael James Stear, Maja Machcińska, Maria Doligalska, and Katarzyna Donskow-Łysoniewska. 2023. "Nematode-Induced Growth Factors Related to Angiogenesis in Autoimmune Disease Attenuation" Life 13, no. 2: 321. https://doi.org/10.3390/life13020321