
Soluble and EV-Associated Diagnostic and Prognostic Biomarkers in Knee Osteoarthritis Pathology and Detection

Abstract
:1. Introduction
2. Osteoarthritis—Overall
2.1. Pathophysiology of Osteoarthritis
2.2. Structural Composition of Articular Cartilage
2.3. Synovial Fluid
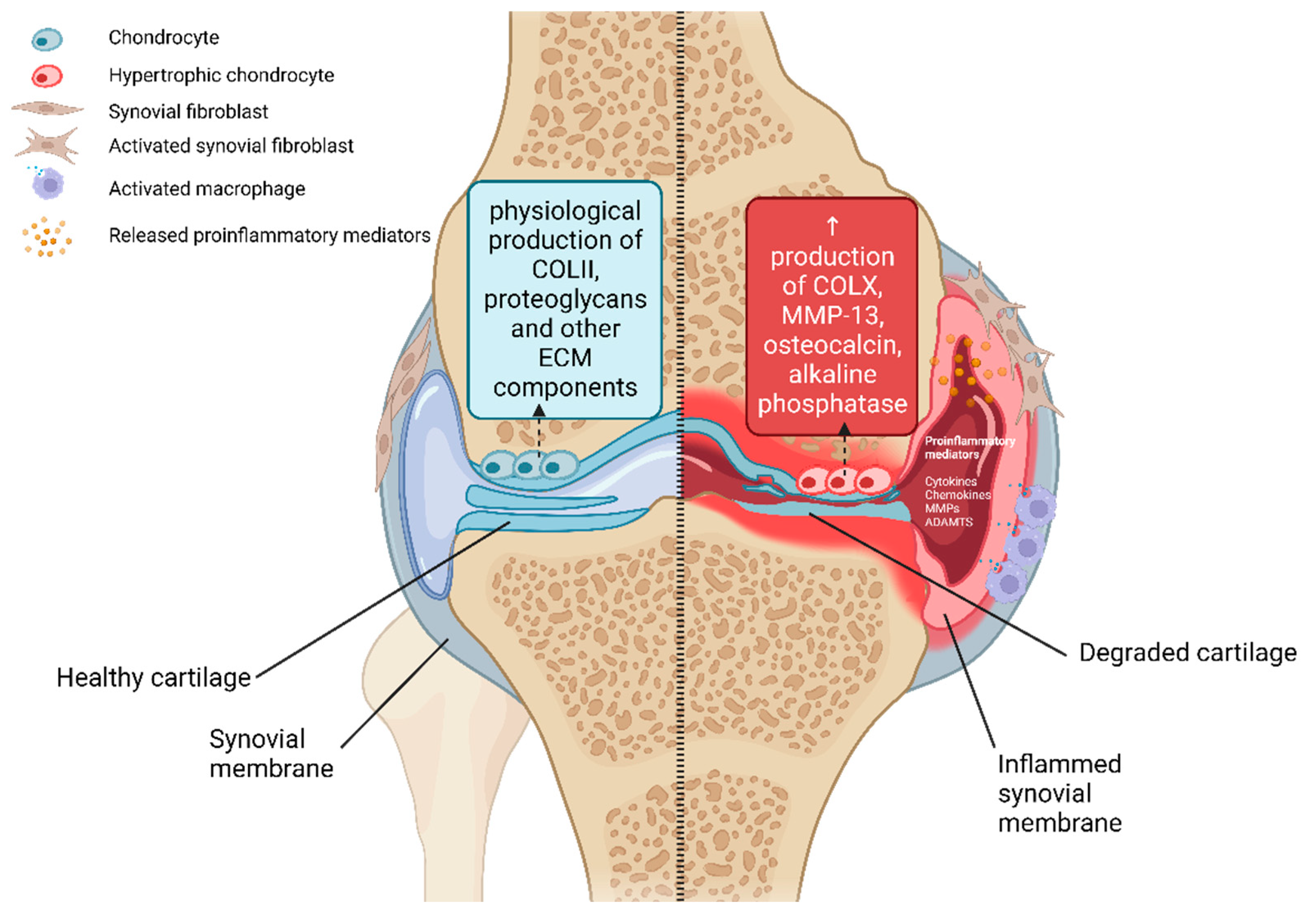
2.4. Synovial Membrane
2.5. Synovial Inflammation
3. Potential Soluble Biomarkers of Osteoarthritis
3.1. Inflammatory Biomarkers
3.1.1. Cytokines and Chemokines
Potential Cytokine/Chemokine Biomarkers in Synovial Fluid
Potential Cytokine/Chemokine Biomarkers in Blood
3.2. Collagenous Biomarkers
3.2.1. C-Terminal Telopeptide of Collagen Type II
3.2.2. Type II Collagen-Specific Biomarker
3.2.3. Type II Collagen Cleavage Product
3.3. Non-Collagenous Biomarkers
3.3.1. Cartilage Oligomeric Matrix Protein
3.3.2. S100 Proteins
3.3.3. C-Terminal End-Product of Vitronectin and C3f Peptide
4. Potential EV-Associated Biomarkers of Osteoarthritis
4.1. EV-Associated Protein/Lipid Biomarkers
4.2. EV-Associated miRNA Biomarkers
4.3. EV-Associated lncRNA Biomarkers
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malemud, C.J. Biologic basis of osteoarthritis: State of the evidence. Curr. Opin. Rheumatol. 2015, 27, 289–294. [Google Scholar] [CrossRef]
- Berenbaum, F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr. Cartil. 2013, 21, 16–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vina, E.R.; Kwoh, C. Epidemiology of osteoarthritis: Literature update. Curr. Opin. Rheumatol. 2018, 30, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Cross, M.; Smith, E.; Hoy, D.; Nolte, S.; Ackerman, I.; Fransen, M.; Bridgett, L.; Williams, S.; Guillemin, F.; Hill, C.; et al. The global burden of hip and knee osteoarthritis: Estimates from the Global Burden of Disease 2010 study. Ann. Rheum. Dis. 2014, 73, 1323. [Google Scholar] [CrossRef] [PubMed]
- Abramoff, B.; Caldera, F. Osteoarthritis: Pathology, Diagnosis, and Treatment Options. Med. Clin. North Am. 2020, 104, 293–311. [Google Scholar] [CrossRef]
- Grunz, J.P.; Gietzen, C. Osteoarthritis of the Wrist: Pathology, Radiology, and Treatment. Semin. Musculoskelet. Radiol. 2021, 25, 294–303. [Google Scholar] [CrossRef]
- Mathiessen, A.; Conaghan, P. Synovitis in osteoarthritis: Current understanding with therapeutic implications. Arthritis Res. Ther. 2017, 19, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culvenor, A.G.; Øiestad, B.; Hart, H.; Stefanik, J.J.; Guermazi, A.; Crossley, K.M. Prevalence of knee osteoarthritis featuRes. on magnetic resonance imaging in asymptomatic uninjured adults: A systematic review and meta-analysis. Br. J. Sport. Med. 2019, 53, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Runhaar, J.; Kloppenburg, M.; Boers, M.; Bijlsma JW, J.; Bierma-Zeinstra SM, A.; CREDO Expert Group. Towards developing diagnostic criteria for early knee osteoarthritis: Data from the CHECK study. Rheumatology 2021, 60, 2448–2455. [Google Scholar] [CrossRef]
- Bijlsma, J.W.J.; Berenbaum, F.; Lafeber, F. Osteoarthritis: An update with relevance for clinical practice. Lancet 2011, 377, 2115–2126. [Google Scholar] [CrossRef] [PubMed]
- Benis, S.; Vanhove, W.; Van Tongel, A.; Hollevoet, N. Non-traumatic primary and secondary osteoarthritis of the distal radioulnar joint. J. Hand Surg. Eur. Vol. 2019, 44, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Taruc-Uy, R.L.; Lynch, S. Diagnosis and treatment of osteoarthritis. Prim Care 2013, 40, 821–836. [Google Scholar] [CrossRef] [PubMed]
- Berenbaum, F.; Griffin, T.; Liu-Bryan, R. Review: Metabolic Regulation of Inflammation in Osteoarthritis. Arthritis Rheumatol. 2017, 69, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashford, S.; Williard, J. Osteoarthritis: A review. Nurse Pract. 2014, 39, 1–8. [Google Scholar] [CrossRef]
- Felson, D.T.; Lawrence, R.; Dieppe, P.; Hirsch, R.; Helmick, C.; Jordan, J.; Kington, R.; Lane, N.; Nevitt, M.; Zhang, Y.; et al. Osteoarthritis: New Insights. Part 1: The Disease and Its Risk Factors. Ann. Intern. Med. 2000, 133, 635–646. [Google Scholar] [CrossRef]
- Berenbaum, F.; Wallace, I.; Lieberman, D.; Felson, D. Modern-day environmental factors in the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2018, 14, 674–681. [Google Scholar] [CrossRef]
- Xia, B.; Di, C.; Zhang, J.; Hu, S.; Jin, H.; Tong, P. Osteoarthritis pathogenesis: A review of molecular mechanisms. Calcif. Tissue Int. 2014, 95, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Carballo, C.B.; Nakagawa, Y.; Sekiya, I.; Rodeo, S. Basic Science of Articular Cartilage. Clin. Sport. Med. 2017, 36, 413–425. [Google Scholar] [CrossRef]
- Brody, L.T. Knee osteoarthritis: Clinical connections to articular cartilage structure and function. Phys. Ther. Sport 2015, 16, 301–316. [Google Scholar] [CrossRef]
- Guilak, F.; Nims, R.; Dicks, A.; Wu, C.-L.; Meulenbelt, I. Osteoarthritis as a disease of the cartilage pericellular matrix. Matrix Biol. J. Int. Soc. Matrix Biol. 2018, 71–72, 40–50. [Google Scholar] [CrossRef]
- Bolduc, J.A.; Collins, J.; Loeser, R. Reactive oxygen species, aging and articular cartilage homeostasis. Free Radic. Biol. Med. 2019, 132, 73–82. [Google Scholar] [CrossRef]
- van der Kraan, P.M.; Davidson, E.B.; van den Berg, W. A role for age-related changes in TGFbeta signaling in aberrant chondrocyte differentiation and osteoarthritis. Arthritis Res. 2010, 12, 201. [Google Scholar] [CrossRef] [Green Version]
- Charlier, E.; Deroyer, C.; Ciregia, F.; Malaise, O.; Neuville, S.; Plener, Z.; Malaise, M.; de Seny, D. Chondrocyte dedifferentiation and osteoarthritis (OA). Biochem. Pharm. 2019, 165, 49–65. [Google Scholar] [CrossRef]
- He, Y.; Manon-Jensen, T.; Arendt-Nielsen, L.; Petersen, K.; Christiansen, T.; Samuels, J.; Abramson, S.; Karsdal, M.; Attur, M.; Bay-Jensen, A. Potential diagnostic value of a type X collagen neo-epitope biomarker for knee osteoarthritis. Osteoarthr. Cartil. 2019, 27, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Munjal, A.; Bapat, S.; Hubbard, D.; Hunter, M.; Kolhe, R.; Fulzele, S. Advances in Molecular biomarker for early diagnosis of Osteoarthritis. Biomol. Concepts 2019, 10, 111–119. [Google Scholar] [CrossRef]
- Bonnevie, E.D.; Galesso, D.; Secchieri, C.; Cohen, I.; Bonassar, L. Elastoviscous Transitions of Articular Cartilage Reveal a Mechanism of Synergy between Lubricin and Hyaluronic Acid. PLoS ONE 2015, 10, e0143415. [Google Scholar] [CrossRef]
- Li, N.; Gao, J.; Mi, L.; Zhang, G.; Zhang, L.; Zhang, N.; Huo, R.; Hu, J.; Xu, K. Synovial membrane mesenchymal stem cells: Past life, current situation, and application in bone and joInt. diseases. Stem Cell Res. 2020, 11, 381. [Google Scholar] [CrossRef]
- Eymard, F.; Pigenet, A.; Citadelle, D.; Flouzat-Lachaniette, C.; Poignard, A.; Benelli, C.; Berenbaum, F.; Chevalier, X.; Houard, X. Induction of an inflammatory and prodegradative phenotype in autologous fibroblast-like synoviocytes by the infrapatellar fat pad from patients with knee osteoarthritis. Arthritis Rheumatol. 2014, 66, 2165–2174. [Google Scholar] [CrossRef]
- Scanzello, C.R.; Goldring, S. The role of synovitis in osteoarthritis pathogenesis. Bone 2012, 51, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Harvanová, D.; Tóthová, T.; Sarišský, M.; Amrichová, J.; Rosocha, J. Isolation and characterization of synovial mesenchymal stem cells. Folia Biol. 2011, 57, 119–124. [Google Scholar]
- Fox, D.B.; Warnock, J. Cell-based meniscal tissue engineering: A case for synoviocytes. Clin. Orthop. Relat. Res. 2011, 469, 2806–2816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haubruck, P.; Pinto, M.; Moradi, B.; Little, C.; Gentek, R. Monocytes, Macrophages, and Their Potential Niches in Synovial Joints—Therapeutic Targets in Post-Traumatic Osteoarthritis? Front. Immunol. 2021, 12, 763702. [Google Scholar] [CrossRef] [PubMed]
- Farinelli, L.; Aquili, A.; Mattioli-Belmonte, M.; Manzotti, S.; D‘Angelo, F.; Ciccullo, C.; Gigante, A. Synovial mast cells from knee and hip osteoarthritis: Histological study and clinical correlations. J. Exp. Orthop. 2022, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Altobelli, E.; Angeletti, P.; Piccolo, D.; De Angelis, R. Synovial Fluid and Serum Concentrations of Inflammatory Markers in Rheumatoid Arthritis, Psoriatic Arthritis and Osteoarthitis: A Systematic Review. Curr. Rheumatol. Rev. 2017, 13, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Dequeker, J.; Luyten, F. The history of osteoarthritis-osteoarthrosis. Ann. Rheum. Dis. 2008, 67, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Felson, D.T. Osteoarthritis as a disease of mechanics. Osteoarthr. Cartil. 2013, 21, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Verbruggen, G.; Wittoek, R.; Cruyssen, B.V.; Elewaut, D. Tumour necrosis factor blockade for the treatment of erosive osteoarthritis of the interphalangeal finger joints: A double blind, randomised trial on structure modification. Ann. Rheum. Dis. 2012, 71, 891–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellam, J.; Berenbaum, F. The role of synovitis in pathophysiology and clinical symptoms of osteoarthritis. Nat. Rev. Rheumatol. 2010, 6, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Potin, I.; Largo, R.; Roman-Blas, J.; Herrero-Beaumont, G.; Walsh, D. Characterization of multinucleated giant cells in synovium and subchondral bone in knee osteoarthritis and rheumatoid arthritis. BMC Musculoskelet. Disord. 2015, 16, 226. [Google Scholar] [CrossRef] [Green Version]
- Klein-Wieringa, I.R.; de Lange-Brokaar, B.; Yusuf, E.; Andersen, S.; Kwekkeboom, J.; Kroon, H.; van Osch, G.; Zuurmond, A.; Stojanovic-Susulic, V.; Nelissen, R.; et al. Inflammatory Cells in Patients with Endstage Knee Osteoarthritis: A Comparison between the Synovium and the Infrapatellar Fat Pad. J. Rheumatol. 2016, 43, 771–778. [Google Scholar] [CrossRef]
- Glyn-Jones, S.; Palmer, A.; Agricola, R.; Price, A.; Vincent, T.; Weinans, H.; Car, A.J. Osteoarthritis. Lancet 2015, 386, 376–387. [Google Scholar] [CrossRef]
- Madry, H.; Kon, E.; Condello, V.; Peretti, G.; Steinwachs, M.; Seil, R.; Berruto, M.; Engebretsen, L.; Filardo, G.; Angele, P. Early osteoarthritis of the knee. Knee Surg. Sport. Traumatol. Arthrosc. 2016, 24, 1753–1762. [Google Scholar] [CrossRef]
- Lotz, M.; Martel-Pelletier, J.; Christiansen, C.; Brandi, M.; Bruyère, O.; Chapurlat, R.; Collette, J.; Cooper, C.; Giacovelli, G.; Kanis, J.; et al. Republished: Value of biomarkers in osteoarthritis: Current status and perspectives. Postgrad Med. J. 2014, 90, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Nordahl, S.; Alstergren, P.; Eliasson, S.; Kopp, S. Interleukin-1beta in plasma and synovial fluid in relation to radiographic changes in arthritic temporomandibular joints. Eur. J. Oral Sci. 1998, 106, 559–563. [Google Scholar] [CrossRef]
- Panina, S.B.; Krolevets, I.; Milyutina, N.; Sagakyants, A.; Kornienko, I.; Ananyan, A.; Zabrodin, M.; Plotnikov, A.; Vnukov, V. Circulating levels of proinflammatory mediators as potential biomarkers of post-traumatic knee osteoarthritis development. J. Orthop. Traumatol. 2017, 18, 349–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konopka, J.; Richbourgh, B.; Liu, C. The role of PGRN in musculoskeletal development and disease. Front. Biosci. Landmark Ed. 2014, 19, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Stannus, O.; Jones, G.; Cicuttini, F.; Parameswaran, V.; Quinn, S.; Burgess, J.; Ding, C. Circulating levels of IL-6 and TNF-α are associated with knee radiographic osteoarthritis and knee cartilage loss in older adults. Osteoarthr. Cartil. 2010, 18, 1441–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monibi, F.; Roller, B.; Stoker, A.; Garner, B.; Bal, S.; Cook, J. Identification of Synovial Fluid Biomarkers for Knee Osteoarthritis and Correlation with Radiographic Assessment. J. Knee Surg. 2016, 29, 242–247. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Yang, Z.; Zhang, Z.; Zhang, Z.; Kang, Y.; Huang, G.; Wang, S.; Huang, H.; Liao, W. CCL3 serves as a potential plasma biomarker in knee degeneration (osteoarthritis). Osteoarthr. Cartil. 2015, 23, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Beekhuizen, M.; Gierman, L.; van Spil, W.; Van Osch, G.; Huizinga, T.; Saris, D.; Creemers, L.; Zuurmond, A. An explorative study comparing levels of soluble mediators in control and osteoarthritic synovial fluid. Osteoarthr. Cartil. 2013, 21, 918–922. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Tian, J.; Pan, H.; Gao, J.; Yao, M. Association of CCL13 levels in serum and synovial fluid with the radiographic severity of knee osteoarthritis. J. Investig. Med. 2015, 63, 545–547. [Google Scholar] [CrossRef]
- Xu, Q.; Sun, X.; Shang, X.; Jiang, H. Association of CXCL12 levels in synovial fluid with the radiographic severity of knee osteoarthritis. J. Investig. Med. 2012, 60, 898–901. [Google Scholar] [CrossRef]
- Meulenbelt, I.; Kloppenburg, M.; Kroon, H.; Houwing-Duistermaat, J.; Garnero, P.; Le Graverand, M.H.; DeGroot, J.; Slagboom, P. Urinary CTX-II levels are associated with radiographic subtypes of osteoarthritis in hip, knee, hand, and facet joints in subject with familial osteoarthritis at multiple sites: The GARP study. Ann. Rheum. Dis. 2006, 65, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arunrukthavon, P.; Heebthamai, D.; Benchasiriluck, P.; Chaluay, S.; Chotanaphuti, T.; Khuangsirikul, S. Can urinary CTX-II be a biomarker for knee osteoarthritis? Arthroplasty 2020, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Valdes, A.M.; Meulenbelt, I.; Chassaing, E.; Arden, N.; Bierma-Zeinstra, S.; Hart, D.; Hofman, A.; Karsdal, M.; Kloppenburg, M.; Kroon, H.; et al. Large scale meta-analysis of urinary C-terminal telopeptide, serum cartilage oligomeric protein and matrix metalloprotease degraded type II collagen and their role in prevalence, incidence and progression of osteoarthritis. Osteoarthr. Cartil. 2014, 22, 683–689. [Google Scholar] [CrossRef] [Green Version]
- Henrotin, Y.; Deberg, M.; Dubuc, J.; Quettier, E.; Christgau, S.; Reginster, J. Type II collagen peptides for measuring cartilage degradation. Biorheology 2004, 41, 543–547. [Google Scholar]
- Poole, A.R.; Ha, N.; Bourdon, S.; Sayre, E.; Guermazi, A.; Cibere, J. Ability of a Urine Assay of Type II Collagen Cleavage by Collagenases to Detect Early Onset and Progression of Articular Cartilage Degeneration: Results from a Population-based Cohort Study. J. Rheumatol. 2016, 43, 1864–1870. [Google Scholar] [CrossRef]
- Plsikova Matejova, J.; Spakova, T. A Preliminary Study of Combined Detection of COMP, TIMP-1, and MMP-3 in Synovial Fluid: Potential Indicators of Osteoarthritis Progression. Cartilage 2021, 13 (Suppl. S2), 1421s–1430s. [Google Scholar] [CrossRef]
- Kluzek, S.; Bay-Jensen, A.; Judge, A.; Karsdal, M.; Shorthose, M.; Spector, T.; Hart, D.; Newton, J.; Arden, N. Serum cartilage oligomeric matrix protein and development of radiographic and painful knee osteoarthritis. A community-based cohort of middle-aged women. Biomarkers 2015, 20, 557–564. [Google Scholar] [CrossRef] [Green Version]
- Ruan, G.; Xu, J.; Wang, K.; Zheng, S.; Wu, J.; Ren, J.; Bian, F.; Chang, B.; Zhu, Z.; Han, W.; et al. Associations between serum S100A8/S100A9 and knee symptoms, joInt. structuRes. and cartilage enzymes in patients with knee osteoarthritis. Osteoarthr. Cartil. 2019, 27, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Zhu, W.; Li, H.; Ma, D.; Liu, W.; Yu, W.; Wang, L.; Cao, Y. Association between cytokines and exosomes in synovial fluid of individuals with knee osteoarthritis. Mod. Rheumatol. 2020, 30, 758–764. [Google Scholar] [CrossRef]
- Zhang, X.; Hsueh, M.; Huebner, J.; Kraus, V. TNF-α Carried by Plasma Extracellular Vesicles Predicts Knee Osteoarthritis Progression. Front. Immunol 2021, 12, 758386. [Google Scholar] [CrossRef] [PubMed]
- Kolhe, R.; Owens, V.; Sharma, A. Sex-Specific Differences in Extracellular Vesicle Protein Cargo in Synovial Fluid of Patients with Osteoarthritis. Life 2020, 10, 337. [Google Scholar] [CrossRef]
- Ben-Trad, L.; Matei, C.; Sava, M.; Filali, S.; Duclos, M.; Berthier, Y.; Guichardant, M.; Bernoud-Hubac, N. Synovial Extracellular Vesicles: Structure and Role in Synovial Fluid Tribological Performances. Int. J. Mol. Sci. 2022, 23, 11998. [Google Scholar] [CrossRef]
- Mao, G.; Hu, S.; Zhang, Z.; Wu, P.; Zhao, X.; Lin, R.; Liao, W.; Kang, Y. Exosomal miR-95-5p regulates chondrogenesis and cartilage degradation via histone deacetylase 2/8. J. Cell. Mol. Med. 2018, 22, 5354–5366. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Li, Z.; Zhang, Z.; Yang, Z.; Kang, Y.; Zhao, X.; Long, D.; Hu, S.; Gu, M.; He, S.; et al. MicroRNA-193b-3p regulates chondrogenesis and chondrocyte metabolism by targeting HDAC3. Theranostics 2018, 8, 2862–2883. [Google Scholar] [CrossRef]
- Mao, G.; Zhang, Z.; Hu, S.; Zhang, Z.; Chang, Z.; Huang, Z.; Liao, W.; Kang, Y. Exosomes derived from miR-92a-3p-overexpressing human mesenchymal stem cells enhance chondrogenesis and suppress cartilage degradation via targeting WNT5A. Stem Cell Res. Ther. 2018, 9, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.C.; Yuan, T.; Zhang, Y.; Yin, W.; Guo, S.; Zhang, C. Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics 2017, 7, 180–195. [Google Scholar] [CrossRef]
- Jin, Z.; Ren, J.; Qi, S. Human bone mesenchymal stem cells-derived exosomes overexpressing microRNA-26a-5p alleviate osteoarthritis via down-regulation of PTGS2. Int. Immunopharmacol. 2020, 78, 105946. [Google Scholar] [CrossRef]
- Wu, J.; Kuang, L.; Chen, C.; Yang, J.; Zeng, W.; Li, T.; Chen, H.; Huang, S.; Fu, Z.; Li, J.; et al. miR-100-5p-abundant exosomes derived from infrapatellar fat pad MSCs protect articular cartilage and ameliorate gait abnormalities via inhibition of mTOR in osteoarthritis. Biomaterials 2019, 206, 87–100. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, J. Synovial fluid-derived exosomal lncRNA PCGEM1 as biomarker for the different stages of osteoarthritis. Int. Orthop. 2018, 42, 2865–2872. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lin, L.; Zou, R.; Wen, C.; Wang, Z.; Lin, F. MSC-derived exosomes promote proliferation and inhibit apoptosis of chondrocytes via lncRNA-KLF3-AS1/miR-206/GIT1 axis in osteoarthritis. Cell Cycle 2018, 17, 2411–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Qiu, S.; Sun, L.; Zuo, J. Knockdown of exosome-mediated lnc-PVT1 alleviates lipopolysaccharide-induced osteoarthritis progression by mediating the HMGB1/TLR4/NF-κB pathway via miR-93-5p. Mol. Med. Rep. 2020, 22, 5313–5325. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Z.; Cui, Y.; Cui, X.; Chen, C.; Wang, Z. Exosomes Isolated From Bone Marrow Mesenchymal Stem Cells Exert a Protective Effect on Osteoarthritis via lncRNA LYRM4-AS1-GRPR-miR-6515-5p. Front. Cell Dev. Biol. 2021, 9, 644380. [Google Scholar] [CrossRef] [PubMed]
- Ene, R.; Sinescu, R.; Ene, P.; Cîrstoiu, M.; Cîrstoiu, F. Synovial inflammation in patients with different stages of knee osteoarthritis. Rom. J. Morphol. Embryol. 2015, 56, 169–173. [Google Scholar]
- Krasnokutsky, S.; Belitskaya-Lévy, I.; Bencardino, J.; Samuels, J.; Attur, M.; Regatte, R.; Rosenthal, P.; Greenberg, J.; Schweitzer, M.; Abramson, S.; et al. Quantitative magnetic resonance imaging evidence of synovial proliferation is associated with radiographic severity of knee osteoarthritis. Arthritis Rheum. 2011, 63, 2983–2991. [Google Scholar] [CrossRef] [Green Version]
- Guermazi, A.; Hayashi, D.; Roemer, F.; Zhu, Y.; Niu, J.; Crema, M.; Javaid, M.; Marra, M.; Lynch, J.; El-Khoury, G.; et al. Synovitis in knee osteoarthritis assessed by contrast-enhanced magnetic resonance imaging (MRI) is associated with radiographic tibiofemoral osteoarthritis and MRI-detected widespread cartilage damage: The MOST study. J. Rheumatol. 2014, 41, 501–508. [Google Scholar] [CrossRef] [Green Version]
- Meehan, R.T.; Regan, E.; Hoffman, E.; Wolf, M.; Gill, M.; Crooks, J. Synovial Fluid Cytokines, Chemokines and MMP Levels in Osteoarthritis Patients with Knee Pain Display a Profile Similar to Many Rheumatoid Arthritis Patients. J. Clin. Med. 2021, 10, 5027. [Google Scholar] [CrossRef]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.-P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33–42. [Google Scholar] [CrossRef]
- Molnar, V.; Matišić, V. Cytokines and Chemokines Involved in Osteoarthritis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 9208. [Google Scholar] [CrossRef]
- Johnson, A.; Huang, Y.-C.; Mao, C.-F.; Chen, C.-K.; Thomas, S.; Kuo, H.-P.; Miao, S.; Kong, Z.-L. Protective effect of ethanolic extract of Echinacea purpurea contained nanoparticles on meniscal/ligamentous injury induced osteoarthritis in obese male rats. Sci. Rep. 2022, 12, 5354. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shah, K.; Luo, J. Strategies for Articular Cartilage Repair and Regeneration. Front. Bioeng. Biotechnol. 2021, 9, 770655. [Google Scholar] [CrossRef] [PubMed]
- Slovacek, H.; Khanna, R.; Poredos, P.; Jezovnik, M.; Hoppensteadt, D.; Fareed, J.; Hopkinson, W. Interrelationship of Osteopontin, MMP-9 and ADAMTS4 in Patients With Osteoarthritis Undergoing Total JoInt. Arthroplasty. Clin. Appl. Thromb. /Hemost. Off. J. Int. Acad. Clin. Appl. Thromb. /Hemost. 2020, 26, 1076029620964864. [Google Scholar] [CrossRef] [PubMed]
- Croft, M.; Siegel, R. Beyond TNF: TNF superfamily cytokines as targets for the treatment of rheumatic diseases. Nat. Rev. Rheumatol. 2017, 13, 217–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Grad, S.; Wen, C.; Lai, Y.; Alini, M.; Qin, L.; Wang, X. An impaired healing model of osteochondral defect in papain-induced arthritis. J. Orthop. Transl. 2020, 26, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Dalal, K. ADAMTS-4 and ADAMTS-5: Key enzymes in osteoarthritis. J. Cell Biochem. 2011, 112, 3507–3514. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Poniatowski, L.; Szukiewicz, D. The Role of Inflammatory and Anti-Inflammatory Cytokines in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; You, Y.; Xi, Y.; Ni, B.; Chu, X.; Zhang, R.; You, H. p-Coumaric Acid Attenuates IL-1β-Induced Inflammatory Responses and Cellular Senescence in Rat Chondrocytes. Inflammation 2020, 43, 619–628. [Google Scholar] [CrossRef]
- Chai, B.; Zheng, Z.; Liao, X.; Li, K.; Liang, J.; Huang, Y.; Tong, C.; Ou, D.; Lu, J. The protective role of omentin-1 in IL-1β-induced chondrocyte senescence. Artif. Cells Nanomed. Biotechnol. 2020, 48, 8–14. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, Q.; Sui, J. Celecoxib prevents tumor necrosis factor-α (TNF-α)-induced cellular senescence in human chondrocytes. Bioengineered 2021, 12, 12812–12820. [Google Scholar] [CrossRef]
- Distel, E.; Cadoudal, T.; Durant, S.; Poignard, A.; Chevalier, X.; Benelli, C. The infrapatellar fat pad in knee osteoarthritis: An important source of interleukin-6 and its soluble receptor. Arthritis Rheum. 2009, 60, 3374–3377. [Google Scholar] [CrossRef] [PubMed]
- Kwan Tat, S.; Padrines, M.; Théoleyre, S.; HeymAnn, D.; Fortun, Y. IL-6, RANKL, TNF-alpha/IL-1: Interrelations in bone resorption pathophysiology. Cytokine Growth Factor Rev. 2004, 15, 49–60. [Google Scholar] [CrossRef]
- Porée, B.; Kypriotou, M.; Chadjichristos, C.; Beauchef, G.; Renard, E.; Legendre, F.; Melin, M.; Gueret, S.; Hartmann, D.; Malléin-Gerin, F.; et al. Interleukin-6 (IL-6) and/or soluble IL-6 receptor down-regulation of human type II collagen gene expression in articular chondrocytes requiRes. a decrease of Sp1.Sp3 ratio and of the binding activity of both factors to the COL2A1 promoter. J. Biol. Chem. 2008, 283, 4850–4865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scanzello, C.R. Chemokines and inflammation in osteoarthritis: Insights from patients and animal models. J. Orthop. Res. 2017, 35, 735–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhavsar, I.; Miller, C.; Al-Sabbagh, M. Macrophage Inflammatory Protein-1 Alpha (MIP-1 alpha)/CCL3: As a Biomarker. Gen. Methods Biomark. Res. Appl. 2015, 223–249. [Google Scholar] [CrossRef]
- Menten, P.; Wuyts, A.; Van Damme, J. Macrophage inflammatory protein-1. Cytokine & Growth Factor Reviews 2002, 13, 455–481. [Google Scholar] [CrossRef]
- Yang, D.-W.; Qian, G.-B.; Jiang, M.-J.; Wang, P.; Wang, K.-Z. Inhibition of microRNA-495 suppresses chondrocyte apoptosis through activation of the NF-κB signaling pathway by regulating CCL4 in osteoarthritis. Gene Ther. 2019, 26, 217–229. [Google Scholar] [CrossRef]
- Pierzchala, A.W.; Kusz, D.; Hajduk, G. CXCL8 and CCL5 Expression in Synovial Fluid and Blood Serum in Patients with Osteoarthritis of the Knee. Arch. Immunol. Ther. Exp. 2011, 59, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Marquez-Curtis, L.A.; Janowska-Wieczorek, A. Enhancing the migration ability of mesenchymal stromal cells by targeting the SDF-1/CXCR4 axis. BioMed. Res. Int. 2013, 2013, 561098. [Google Scholar] [CrossRef] [Green Version]
- Boffa, A.; Merli, G.; Andriolo, L.; Lattermann, C.; SalzmAnn, G.; Filardo, G. Synovial Fluid Biomarkers in Knee Osteoarthritis: A Systematic Review and Quantitative Evaluation Using BIPEDs Criteria. Cartilage 2021, 13 (Suppl. S1), 82S–103S. [Google Scholar] [CrossRef]
- Katz, J.N.; Arant, K.; Loeser, R. Diagnosis and Treatment of Hip and Knee Osteoarthritis: A Review. JAMA 2021, 325, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Blanco, F.J. Osteoarthritis year in review 2014: We need more biochemical biomarkers in qualification phase. Osteoarthr. Cartil. 2014, 22, 2025–2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldring, M.B.; Otero, M.; Plumb, D.; Dragomir, C.; Favero, M.; El Hachem, K.; Hashimoto, K.; Roach, H.; Olivotto, E.; Borzì, R.; et al. Roles of inflammatory and anabolic cytokines in cartilage metabolism: Signals and multiple effectors converge upon MMP-13 regulation in osteoarthritis. Eur. Cell Mater. 2011, 21, 202–220. [Google Scholar] [CrossRef]
- Li, L.; Jiang, B. Serum and synovial fluid chemokine ligand 2/monocyte chemoattractant protein 1 concentrations correlates with symptomatic severity in patients with knee osteoarthritis. Ann. Clin. Biochem. 2015, 52 Pt 2, 276–282. [Google Scholar] [CrossRef]
- Borzì, R.M.; Mazzetti, I.; Cattini, L.; Uguccioni, M.; Baggiolini, M.; Facchini, A. Human chondrocytes express functional chemokine receptors and release matrix-degrading enzymes in response to C-X-C and C-C chemokines. Arthritis Rheum. 2000, 43, 1734–1741. [Google Scholar] [CrossRef]
- Zhao, X.; Gu, M.; Xu, X.; Wen, X.; Yang, G.; Li, L.; Sheng, P.; Meng, F. CCL3/CCR1 mediates CD14(+)CD16(-) circulating monocyte recruitment in knee osteoarthritis progression. Osteoarthr. Cartil. 2020, 28, 613–625. [Google Scholar] [CrossRef]
- Li, L.; Li, Z.; Li, Y.; Hu, X.; Zhang, Y.; Fan, P. Profiling of inflammatory mediators in the synovial fluid related to pain in knee osteoarthritis. BMC Musculoskelet. Disord. 2020, 21, 99. [Google Scholar] [CrossRef] [Green Version]
- Raghu, H.; Lepus, C.; Wang, Q.; Wong, H.; Lingampalli, N.; Oliviero, F.; Punzi, L.; Giori, N.; Goodman, S.; Chu, C.; et al. CCL2/CCR2, but not CCL5/CCR5, mediates monocyte recruitment, inflammation and cartilage destruction in osteoarthritis. Ann. Rheum. Dis. 2017, 76, 914–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitaori, T.; Ito, H.; Schwarz, E.; Tsutsumi, R.; Yoshitomi, H.; Oishi, S.; Nakano, M.; Fujii, N.; Nagasawa, T.; Nakamura, T. Stromal cell-derived factor 1/CXCR4 signaling is critical for the recruitment of mesenchymal stem cells to the fracture site during skeletal repair in a mouse model. Arthritis Rheum. 2009, 60, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Attur, M.; Statnikov, A.; Samuels, J.; Li, Z.; Alekseyenko, A.; Greenberg, J.; Krasnokutsky, S.; Rybak, L.; Lu, Q.; Todd, J.; et al. Plasma levels of interleukin-1 receptor antagonist (IL1Ra) predict radiographic progression of symptomatic knee osteoarthritis. Osteoarthr. Cartil. 2015, 23, 1915–1924. [Google Scholar] [CrossRef] [Green Version]
- Runhaar, J.; Beavers, D.; Miller, G.; Nicklas, B.; Loeser, R.; Bierma-Zeinstra, S.; Messier, S. Inflammatory cytokines mediate the effects of diet and exercise on pain and function in knee osteoarthritis independent of BMI. Osteoarthr. Cartil. 2019, 27, 1118–1123. [Google Scholar] [CrossRef]
- Wang, P. Diagnostic Value of Combined Serum IL-6, TNF-α, and Leptin Levels in Patients with Post-Traumatic Osteoarthritis. Clin. Lab. 2020, 66. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Liu, Z.; Wang, M.; Guo, S.; Cong, H.; Liu, L. Analysis on the expression and value of CCL2 and CCL3 in patients with osteoarthritis. Exp. Mol. Pathol. 2021, 118, 104576. [Google Scholar] [CrossRef]
- Lennerová, T.; Pavelka, K.; Šenolt, L. Biomarkers of hand osteoarthritis. Rheumatol. Int. 2018, 38, 725–735. [Google Scholar] [CrossRef]
- Gabay, C.; Medinger-Sadowski, C.; Gascon, D.; Kolo, F.; Finckh, A. Symptomatic effects of chondroitin 4 and chondroitin 6 sulfate on hand osteoarthritis: A randomized, double-blind, placebo-controlled clinical trial at a single center. Arthritis Rheum. 2011, 63, 3383–3391. [Google Scholar] [CrossRef] [PubMed]
- Freeston, J.E.; Garnero, P.; Wakefield, R.; Hensor, E.; Conaghan, P.; Emery, P. Urinary type II collagen C-terminal peptide is associated with synovitis and predicts structural bone loss in very early inflammatory arthritis. Ann. Rheum. Dis. 2011, 70, 331–333. [Google Scholar] [CrossRef] [PubMed]
- Scarpellini, M.; Lurati, A.; Vignati, G.; Marrazza, M.; Telese, F.; Re, K.; Bellistri, A. Biomarkers, type II collagen, glucosamine and chondroitin sulfate in osteoarthritis follow-up: The “Magenta osteoarthritis study”. J. Orthop. Traumatol. 2008, 9, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Xin, L.; Wu, Z.; Qu, Q.; Wang, R.; Tang, J.; Chen, L. Comparative study of CTX-II, Zn2+, and Ca2+ from the urine for knee osteoarthritis patients and healthy individuals. Medicine 2017, 96, e7593. [Google Scholar] [CrossRef]
- Liu, C.X.; Gao, G.; Qin, X.; Deng, C.; Shen, X. Correlation Analysis of C-terminal telopeptide of collagen type II and Interleukin-1β for Early Diagnosis of Knee Osteoarthritis. Orthop. Surg. 2020, 12, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Duclos, M.E.; Roualdes, O.; Cararo, R.; Rousseau, J.; Roger, T.; Hartmann, D. Significance of the serum CTX-II level in an osteoarthritis animal model: A 5-month longitudinal study. Osteoarthr. Cartil. 2010, 18, 1467–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csifó, E.; Nagy, E.; Horváth, E.; Farr, A.; Muntean, D. Mid-term effects of meloxicam on collagen type II degradation in a rat osteoarthritis model induced by iodoacetate. Farmacia 2015, 63, 556–560. [Google Scholar]
- Park, Y.M.; Kim, S.; Lee, K.; Yang, S.; Min, B.-H.; Yoon, H. Detection of CTX-II in serum and urine to diagnose osteoarthritis by using a fluoro-microbeads guiding chip. Biosens. Bioelectron. 2015, 67, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Lohmander, L.S.; Atley, L.; Pietka, T.; Eyre, D. The release of crosslinked peptides from type II collagen into human synovial fluid is increased soon after joInt. injury and in osteoarthritis. Arthritis Rheum. 2003, 48, 3130–3139. [Google Scholar] [CrossRef]
- Bauer, D.C.; Hunter, D.; Abramson, S.; Attur, M.; Corr, M.; Felson, D.; Heinegård, D.; Jordan, J.; Kepler, T.; Lane, N.; et al. Classification of osteoarthritis biomarkers: A proposed approach. Osteoarthr. Cartil. 2006, 14, 723–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henrotin, Y.; Berenbaum, F.; Chevalier, X.; Marty, M.; Richette, P.; Rannou, F. Reduction of the Serum Levels of a Specific Biomarker of Cartilage Degradation (Coll 2-1) by Hyaluronic Acid (KARTILAGE® CROSS) Compared to Placebo in Painful Knee Osteoarthritis Patients: The EPIKART Study, a Pilot Prospective Comparative Randomized Double Blind Trial. BMC Musculoskelet. Disord. 2017, 18, 222. [Google Scholar] [CrossRef] [Green Version]
- Deberg, M.; Dubuc, J.; Labasse, A.; Sanchez, C.; Quettier, E.; Bosseloir, A.; Crielaard, J.; Henrotin, Y. One-year follow-up of Coll 2-1, Coll 2-1NO2 and myeloperoxydase serum levels in osteoarthritis patients after hip or knee replacement. Ann. Rheum. Dis. 2008, 67, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Coppelman, E.B.; David, F.; Tóth, F.; Ernst, N.; Trumble, T. The association between collagen and bone biomarkers and radiographic osteoarthritis in the distal tarsal joints of horses. Equine Vet. J. 2020, 52, 391–398. [Google Scholar] [CrossRef]
- Kumahashi, N.; Swärd, P.; Larsson, S.; Lohmander, L.; Frobell, R.; Struglics, A. Type II collagen C2C epitope in human synovial fluid and serum after knee injury--associations with molecular and structural markers of injury. Osteoarthr. Cartil. 2015, 23, 1506–1512. [Google Scholar] [CrossRef] [Green Version]
- Posey, K.L.; Coustry, F.; Hecht, J. Cartilage oligomeric matrix protein: COMPopathies and beyond. Matrix Biol. 2018, 71–72, 161–173. [Google Scholar] [CrossRef]
- Bay-Jensen, A.C.; Reker, D.; Kjelgaard-Petersen, C.; Mobasheri, A.; Karsdal, M.; Ladel, C.; Henrotin, Y.; Thudium, C. Osteoarthritis year in review 2015: Soluble biomarkers and the BIPED criteria. Osteoarthr. Cartil. 2016, 24, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, D.J.; Li, J.; LaValley, M.; Bauer, D.; Nevitt, M.; DeGroot, J.; Poole, R.; Eyre, D.; Guermazi, A.; Gale, D.; et al. Cartilage markers and their association with cartilage loss on magnetic resonance imaging in knee osteoarthritis: The Boston Osteoarthritis Knee Study. Arthritis Res. 2007, 9, R108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, P.; Dalal, K. Serum cartilage oligomeric matrix protein (COMP) in knee osteoarthritis: A novel diagnostic and prognostic biomarker. J. Orthop. Res. 2013, 31, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Zhang, Z.; Song, X.; Bai, H.; Li, Y.; Li, X.; Zhao, J.; Ma, Y.; Gao, L. Combined detection of COMP and CS846 biomarkers in experimental rat osteoarthritis: A potential approach for assessment and diagnosis of osteoarthritis. J. Orthop. Surg. Res. 2018, 13, 230. [Google Scholar] [CrossRef] [PubMed]
- Endres, E.; van Drongelen, S.; Meurer, A.; Zaucke, F.; Stief, F. Effect of total joInt. replacement in hip osteoarthritis on serum COMP and its correlation with mechanical-functional parameters of gait analysis. Osteoarthr. Cart. Open 2020, 2, 100034. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Dubuc, J.; Montell, E.; Vergés, J.; Munaut, C.; Noël, A.; Henrotin, Y. Gene expression pattern of cells from inflaMed. and normal areas of osteoarthritis synovial membrane. Arthritis Rheumatol. 2014, 66, 960–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Lent, P.L.; Blom, A.; Schelbergen, R.; Slöetjes, A.; Lafeber, F.; Lems, W.; Cats, H.; Vogl, T.; Roth, J.; van den Berg, W. Active involvement of alarmins S100A8 and S100A9 in the regulation of synovial activation and joInt. destruction during mouse and human osteoarthritis. Arthritis Rheum. 2012, 64, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- Nagy, E.E.; Nagy-Finna, C.; Popoviciu, H. Soluble Biomarkers of Osteoporosis and Osteoarthritis, from Pathway Mapping to Clinical Trials: An Update. Clin. Interv. Aging 2020, 15, 501–518. [Google Scholar] [CrossRef] [Green Version]
- De Seny, D.; Sharif, M.; Fillet, M.; Cobraiville, G.; Meuwis, M.; Marée, R.; Hauzeur, J.; Wehenkel, L.; Louis, E.; Merville, M.; et al. Discovery and biochemical characterisation of four novel biomarkers for osteoarthritis. Ann. Rheum. Dis. 2011, 70, 1144–1152. [Google Scholar] [CrossRef] [Green Version]
- Ourradi, K.; Xu, Y.; de Seny, D.; Kirwan, J.; Blom, A.; Sharif, M. Development and validation of novel biomarker assays for osteoarthritis. PloS ONE 2017, 12, e0181334. [Google Scholar] [CrossRef] [Green Version]
- Yin, B.; Ni, J.; Witherel, C.; Yang, M.; Burdick, J.; Wen, C.; Wong, S. Harnessing Tissue-derived Extracellular Vesicles for Osteoarthritis Theranostics. Theranostics 2022, 12, 207–231. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janockova, J.; Matejova, J.; Moravek, M.; Homolova, L.; Slovinska, L.; Nagyova, A.; Rak, D.; Sedlak, M.; Harvanova, D.; Spakova, T.; et al. Small Extracellular Vesicles Derived from Human Chorionic MSCs as Modern Perspective towards Cell-Free Therapy. Int. J. Mol. Sci. 2021, 22, 13581. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.; Withrow, J.; Hunter, M.; Liu, Y.; Tang, Y.; Fulzele, S.; Hamrick, M. Emerging role of extracellular vesicles in musculoskeletal diseases. Mol. Asp. Med. 2018, 60, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Janockova, J.; Slovinska, L.; Harvanova, D.; Spakova, T.; Rosocha, J. New therapeutic approaches of mesenchymal stem cells-derived exosomes. J. Biomed. Sci. 2021, 28, 39. [Google Scholar] [CrossRef] [PubMed]
- Spakova, T.; Janockova, J. Characterization and Therapeutic Use of Extracellular Vesicles Derived from Platelets. Int. J. Mol. Sci. 2021, 22, 9701. [Google Scholar] [CrossRef]
- Ni, Z.; Kuang, L.; Chen, H.; Xie, Y.; Zhang, B.; Ouyang, J.; Wu, J.; Zhou, S.; Chen, L.; Su, N.; et al. The exosome-like vesicles from osteoarthritic chondrocyte enhanced mature IL-1β production of macrophages and aggravated synovitis in osteoarthritis. Cell Death Dis. 2019, 10, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foers, A.D.; Garnham, A.; Chatfield, S.; Smyth, G. Extracellular Vesicles in Synovial Fluid from Rheumatoid Arthritis Patients Contain miRNAs with Capacity to Modulate Inflammation. Int. J. Mol. Sci. 2021, 22, 4910. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Chubinskaya, S.; Liao, W.; Loeser, R. Wnt5a induces catabolic signaling and matrix metalloproteinase production in human articular chondrocytes. Osteoarthr. Cartil. 2017, 25, 1505–1515. [Google Scholar] [CrossRef] [Green Version]
- Mihanfar, A.; Shakouri, S.; Khadem-Ansari, M.; Fattahi, A.; Latifi, Z.; Nejabati, H.; Nouri, M. Exosomal miRNAs in osteoarthritis. Mol. Biol. Rep. 2020, 47, 4737–4748. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Hu, S.; Zhang, Z.; Lun, J.; Liao, W.; Zhang, Z. Expression of exosomal microRNAs during chondrogenic differentiation of human bone mesenchymal stem cells. J. Cell Biochem. 2019, 120, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Kazimierczyk, M.; Kasprowicz, M.; Kasprzyk, M.; Wrzesinski, J. Human Long Noncoding RNA Interactome: Detection, Characterization and Function. Int. J. Mol. Sci. 2020, 21, 1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolhe, R.; Hunter, M.; Liu, S.; Jadeja, R. Gender-specific differential expression of exosomal miRNA in synovial fluid of patients with osteoarthritis. Sci. Rep. 2017, 7, 2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Type | Category | Molecule | Bodily Fluid/ MSCs Type | Ref. |
|---|---|---|---|---|
| Soluble | Inflammatory | IL-1β | plasma, SF | [44,45] |
| TNFα | N/A | [46] | ||
| IL-6 | SF, serum | [47] | ||
| CCL2 | SF | [48] | ||
| CCL3 | serum | [49] | ||
| CCL5 | SF | [50] | ||
| CCL13 | SF, serum | [51] | ||
| CXCL12 | SF | [52] | ||
| Collagenous | CTX-II | urine | [53,54,55] | |
| Coll 2-1 | urine | [56] | ||
| C2C | urine | [57] | ||
| Non-collagenous | COMP | SF, serum | [58,59] | |
| S100 | serum | [60] | ||
| EV-associated | Protein | IL-1β | SF | [61] |
| IL-17 | SF | [61] | ||
| IL-10 | SF | [61] | ||
| INFγ | SF | [61] | ||
| TNFα | plasma | [62] | ||
| haptoglobin (female) | SF | [63] | ||
| orosomucoid (female) | SF | [63] | ||
| ceruloplasmin (female) | SF | [63] | ||
| β-2-glycoprotein (male) | SF | [63] | ||
| complement component 5 (male) | SF | [63] | ||
| Lipid | phospholipid | SF | [64] | |
| miRNA | miR-95-5p | MSCs-EVs | [65] | |
| miR-193b-3p | plasma-EVs | [66] | ||
| miR-92a-3p | MSCs-EVs | [67] | ||
| miR-140-5p | MSCs-EVs | [68] | ||
| miR-26a-5p | BMSCs-EVs | [69] | ||
| miR-100-5p | IFP-MSCs-EVs | [70] | ||
| lncRNA | PCGEM1 | SF-EVs | [71] | |
| KLF3 AS1 | MSCs-EVs | [72] | ||
| PVT1 | serum-EVs | [73] | ||
| LYRM4-AS1 | BMSCs-EVs | [74] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moravek, M.; Matejova, J.; Spakova, T. Soluble and EV-Associated Diagnostic and Prognostic Biomarkers in Knee Osteoarthritis Pathology and Detection. Life 2023, 13, 342. https://doi.org/10.3390/life13020342
Moravek M, Matejova J, Spakova T. Soluble and EV-Associated Diagnostic and Prognostic Biomarkers in Knee Osteoarthritis Pathology and Detection. Life. 2023; 13(2):342. https://doi.org/10.3390/life13020342
Chicago/Turabian StyleMoravek, Marko, Jana Matejova, and Timea Spakova. 2023. "Soluble and EV-Associated Diagnostic and Prognostic Biomarkers in Knee Osteoarthritis Pathology and Detection" Life 13, no. 2: 342. https://doi.org/10.3390/life13020342