



Food for Thought: Proteomics for Meat Safety
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
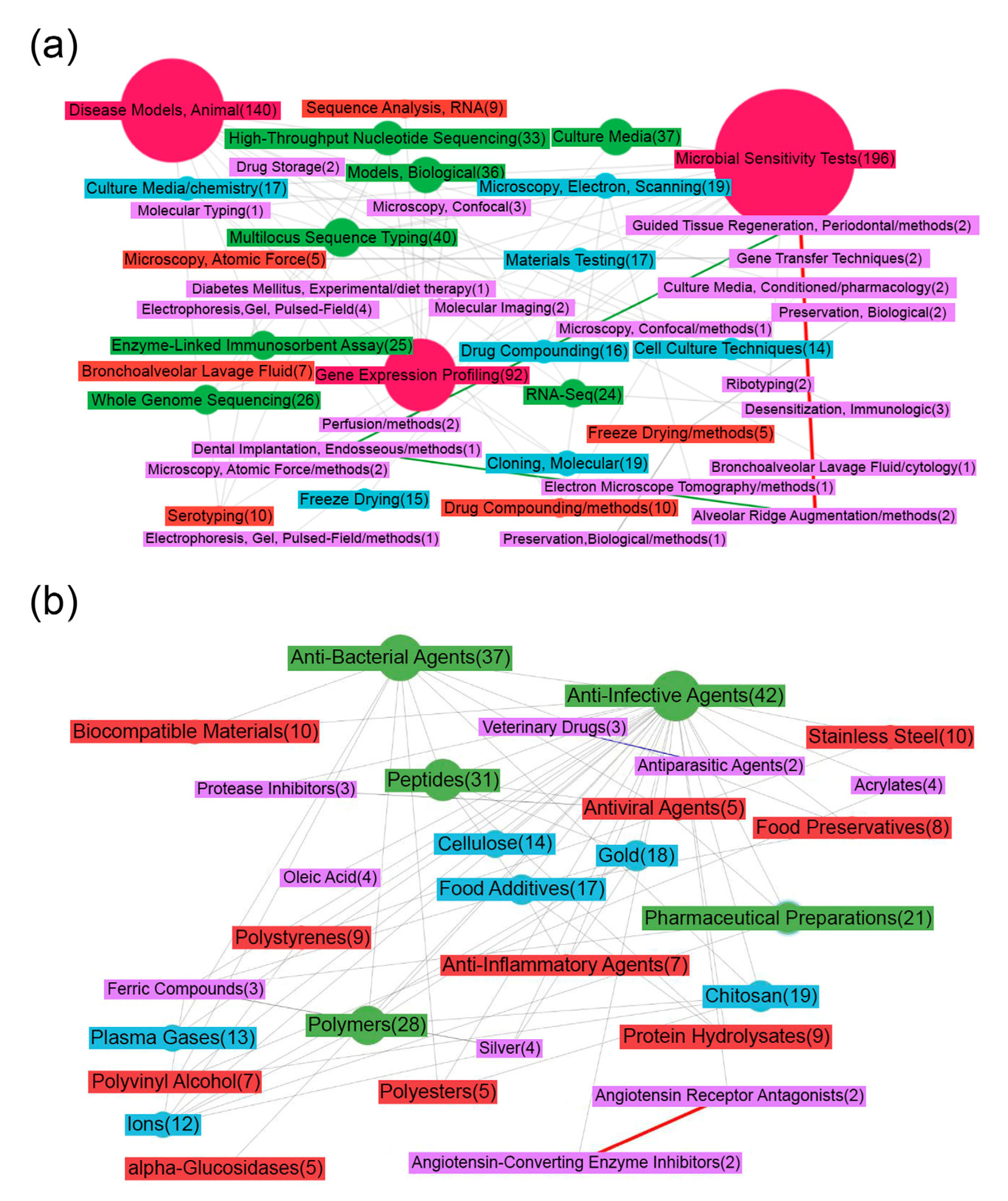
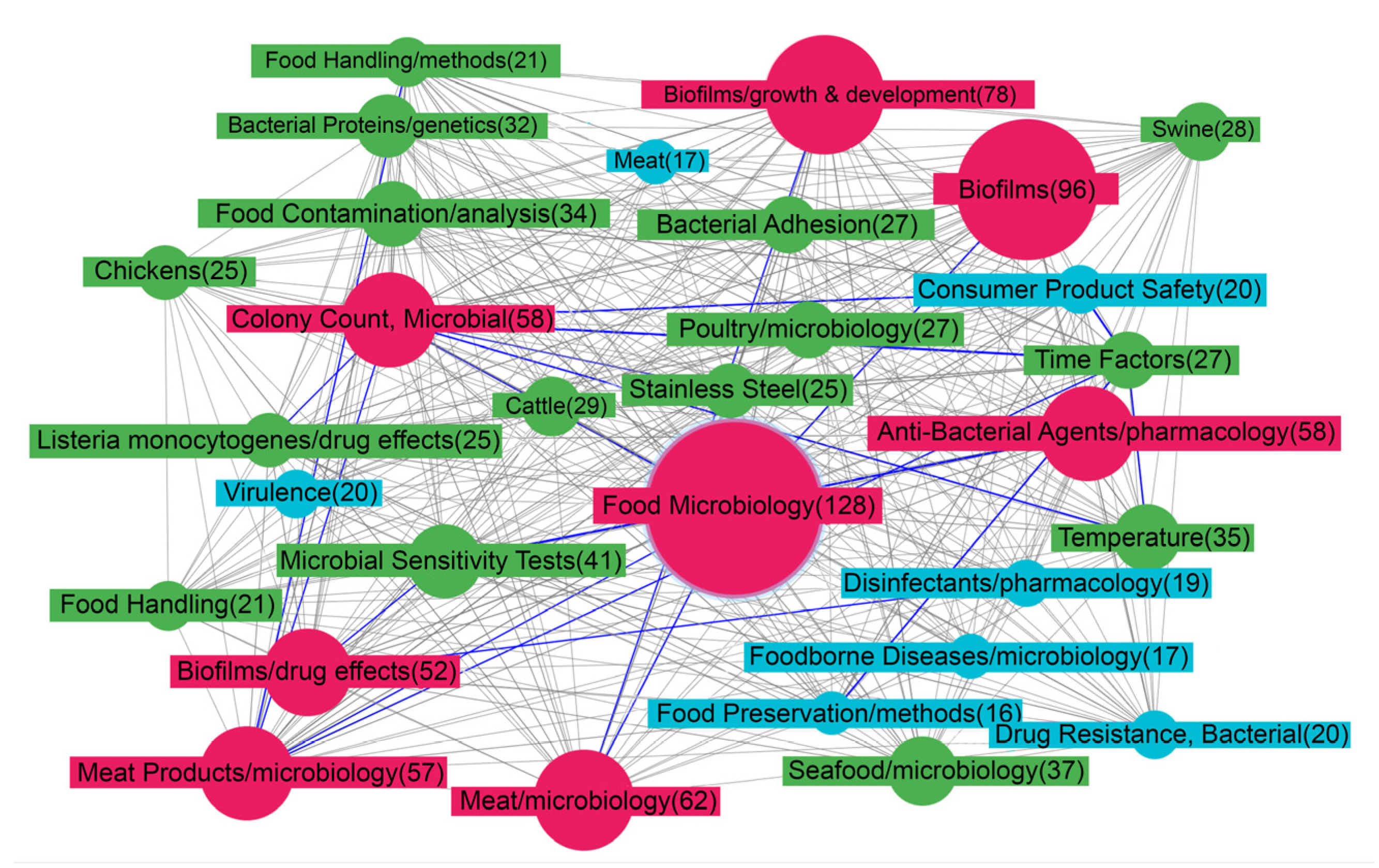
3.1. Subject Area Semantic Network Description
3.2. Genetic Determinants of Resistance
3.3. Proteomic Methods for Studying the Microbiome of Food Industries
- (a)
- Panoramic mass spectrometry
- (b)
- Targeted mass spectrometry
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jadhav, S.R.; Shah, R.M.; Karpe, A.V.; Morrison, P.D.; Kouremenos, K.; Beale, D.J.; Palombo, E.A. Detection of Foodborne Pathogens Using Proteomics and Metabolomics-Based Approaches. Front. Microbiol. 2018, 9, 3132. [Google Scholar] [CrossRef] [PubMed]
- Kataria, R.; Kaundal, R. WeCoNET: A Host–Pathogen Interactome Database for Deciphering Crucial Molecular Networks of Wheat-Common Bunt Cross-Talk Mechanisms. Plant Methods 2022, 18, 73. [Google Scholar] [CrossRef]
- Piras, C.; Roncada, P.; Rodrigues, P.M.; Bonizzi, L.; Soggiu, A. Proteomics in Food: Quality, Safety, Microbes, and Allergens. Proteomics 2016, 16, 799–815. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashton, P.M.; Nair, S.; Peters, T.M.; Bale, J.A.; Powell, D.G.; Painset, A.; Tewolde, R.; Schaefer, U.; Jenkins, C.; Dallman, T.J.; et al. Identification of Salmonella for Public Health Surveillance Using Whole Genome Sequencing. PeerJ 2016, 4, e1752. [Google Scholar] [CrossRef] [Green Version]
- Whole Genome Sequencing (WGS) Program|FDA. Available online: https://www.fda.gov/food/science-research-food/whole-genome-sequencing-wgs-program (accessed on 4 January 2023).
- Jagadeesan, B.; Gerner-Smidt, P.; Allard, M.W.; Leuillet, S.; Winkler, A.; Xiao, Y.; Chaffron, S.; Van Der Vossen, J.; Tang, S.; Katase, M.; et al. The Use of next Generation Sequencing for Improving Food Safety: Translation into Practice. Food Microbiol. 2019, 79, 96–115. [Google Scholar] [CrossRef] [PubMed]
- Sheveleva, S.A.; Kuvaeva, I.B.; Efimochkina, N.R.; Minaeva, L.P. Microbiological Safety of Food: Development of Normative and Methodical Base. Vopr. Pitan 2020, 89, 125–145. [Google Scholar] [CrossRef]
- Khalilpour, A.; Kilic, T.; Khalilpour, S.; Álvarez, M.M.; Yazdi, I.K. Proteomic-Based Biomarker Discovery for Development of next Generation Diagnostics. Appl. Microbiol. Biotechnol. 2017, 101, 475–491. [Google Scholar] [CrossRef]
- Shevchuk, O.; Begonja, A.J.; Gambaryan, S.; Totzeck, M.; Rassaf, T.; Huber, T.B.; Greinacher, A.; Renne, T.; Sickmann, A. Proteomics: A Tool to Study Platelet Function. Int. J. Mol. Sci. 2021, 22, 4776. [Google Scholar] [CrossRef]
- Fornelli, L.; Toby, T.K.; Schachner, L.F.; Doubleday, P.F.; Srzentić, K.; DeHart, C.J.; Kelleher, N.L. Top-down Proteomics: Where We Are, Where We Are Going? J. Proteom. 2018, 175, 3–4. [Google Scholar] [CrossRef]
- Sitammagari, K.K.; Masood, W. Creutzfeldt Jakob Disease; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar] [PubMed]
- Virág, D.; Dalmadi-Kiss, B.; Vékey, K.; Drahos, L.; Klebovich, I.; Antal, I.; Ludányi, K. Current Trends in the Analysis of Post-Translational Modifications. Chromatographia 2020, 83, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Vázquez, R.; Mato, A.; López-Pedrouso, M.; Franco, D.; Sentandreu, M.A.; Zapata, C. Measuring Quantitative Proteomic Distance between Spanish Beef Breeds. Food Chem. 2020, 315, 126293. [Google Scholar] [CrossRef]
- Lasch, P.; Schneider, A.; Blumenscheit, C.; Doellinger, J. Identification of Microorganisms by Liquid Chromatography-Mass Spectrometry (LC-MS1) and in Silico Peptide Mass Libraries. Mol. Cell Proteom. 2020, 19, 2125–2138. [Google Scholar] [CrossRef] [PubMed]
- Ilgisonis, E.; Lisitsa, A.; Kudryavtseva, V.; Ponomarenko, E. Creation of Individual Scientific Concept-Centered Semantic Maps Based on Automated Text-Mining Analysis of PubMed. Adv. Bioinform. 2018, 2018, 4625394. [Google Scholar] [CrossRef]
- Hassani, H.; Beneki, C.; Unger, S.; Mazinani, M.T.; Yeganegi, M.R. Text Mining in Big Data Analytics. Big Data Cogn. Comput. 2020, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Holinski, A.; Burke, M.L.; Morgan, S.L.; McQuilton, P.; Palagi, P.M. Biocuration-Mapping Resources and Needs. F1000Res 2020, 9, 1094. [Google Scholar] [CrossRef]
- Ponomarenko, E.A.; Lisitsa, A.V.; Il’gisonis, E.V.; Archakov, A.I. Construction of Protein Semantic Networks Using PubMed/MEDLINE. Mol. Biol. 2010, 44, 152–161. [Google Scholar] [CrossRef]
- Tarbeeva, S.; Lyamtseva, E.; Lisitsa, A.; Kozlova, A.; Ponomarenko, E.; Ilgisonis, E. ScanBious: Survey for Obesity Genes Using PubMed Abstracts and DisGeNET. J. Pers. Med. 2021, 11, 246. [Google Scholar] [CrossRef] [PubMed]
- Ilgisonis, E.V.; Pyatnitskiy, M.A.; Tarbeeva, S.N.; Aldushin, A.A.; Ponomarenko, E.A. How to Catch Trends Using MeSH Terms Analysis? Scientometrics 2022, 127, 1953–1967. [Google Scholar] [CrossRef]
- Yang, H.; Lee, H.J. Research Trend Visualization by MeSH Terms from PubMed. Int. J. Environ. Res. Public Health 2018, 15, 1113. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-López, P.; Rodríguez-Herrera, J.J.; Cabo, M.L. Tracking Bacteriome Variation over Time in Listeria Monocytogenes-Positive Foci in Food Industry. Int. J. Food Microbiol. 2020, 315, 108439. [Google Scholar] [CrossRef] [PubMed]
- Møretrø, T.; Langsrud, S.; Heir, E.; Møretrø, T.; Langsrud, S.; Heir, E. Bacteria on Meat Abattoir Process Surfaces after Sanitation: Characterisation of Survival Properties of Listeria Monocytogenes and the Commensal Bacterial Flora. Adv. Microbiol. 2013, 3, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Palleroni, N.J. Introduction to the Family Pseudomonadaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 1981; pp. 655–665. [Google Scholar] [CrossRef]
- Hinton, A.; Cason, J.A.; Ingram, K.D. Tracking Spoilage Bacteria in Commercial Poultry Processing and Refrigerated Storage of Poultry Carcasses. Int. J. Food Microbiol. 2004, 91, 155–165. [Google Scholar] [CrossRef]
- Morales, P.A.; Aguirre, J.S.; Troncoso, M.R.; Figueroa, G.O. Phenotypic and Genotypic Characterization of Pseudomonas Spp. Present in Spoiled Poultry Fillets Sold in Retail Settings. LWT 2016, 73, 609–614. [Google Scholar] [CrossRef]
- Algammal, A.M.; Mabrok, M.; Sivaramasamy, E.; Youssef, F.M.; Atwa, M.H.; El-kholy, A.W.; Hetta, H.F.; Hozzein, W.N. Emerging MDR-Pseudomonas Aeruginosa in Fish Commonly Harbor OprL and ToxA Virulence Genes and BlaTEM, BlaCTX-M, and TetA Antibiotic-Resistance Genes. Sci. Rep. 2020, 10, 15961. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.; Yoon, S.S. Virulence Characteristics and an Action Mode of Antibiotic Resistance in Multidrug-Resistant Pseudomonas Aeruginosa. Sci. Rep. 2019, 9, 487. [Google Scholar] [CrossRef] [Green Version]
- Madaha, E.L.; Mienie, C.; Gonsu, H.K.; Bughe, R.N.; Fonkoua, M.C.; Mbacham, W.F.; Alayande, K.A.; Bezuidenhout, C.C.; Ateba, C.N. Whole-Genome Sequence of Multi-Drug Resistant Pseudomonas Aeruginosa Strains UY1PSABAL and UY1PSABAL2 Isolated from Human Broncho-Alveolar Lavage, Yaoundé, Cameroon. PLoS ONE 2020, 15, e0238390. [Google Scholar] [CrossRef]
- de Mesquita Sousa Saraiva, M.; Benevides, V.P.; da Silva, N.M.V.; de Mello Varani, A.; de Freitas Neto, O.C.; Berchieri, Â.; Delgado-Suárez, E.J.; de Lima Rocha, A.D.; Eguale, T.; Munyalo, J.A.; et al. Genomic and Evolutionary Analysis of Salmonella Enterica Serovar Kentucky Sequence Type 198 Isolated From Livestock In East Africa. Front. Cell. Infect. Microbiol. 2022, 12, 1. [Google Scholar] [CrossRef]
- Threlfall, E.J. Antimicrobial Drug Resistance in Salmonella: Problems and Perspectives in Food- and Water-Borne Infections. FEMS Microbiol. Rev. 2002, 26, 141–148. [Google Scholar] [CrossRef]
- Aureli, P.; Fiorucci, G.C.; Caroli, D.; Marchiaro, G.; Novara, O.; Leone, L.; Salmaso, S. An Outbreak of Febrile Gastroenteritis Associated with Corn Contaminated by Listeria Monocytogenes. N. Engl. J. Med. 2000, 342, 1236–1241. [Google Scholar] [CrossRef]
- Kathariou, S. Listeria Monocytogenes Virulence and Pathogenicity, a Food Safety Perspective. J. Food Prot. 2002, 65, 1811–1829. [Google Scholar] [CrossRef] [PubMed]
- Jolivet-Gougeon, A.; Bonnaure-Mallet, M. Biofilms as a Mechanism of Bacterial Resistance. Drug Discov. Today Technol. 2014, 11, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Katsikogianni, M.; Missirlis, Y.F.; Harris, L.; Douglas, J. Concise Review of Mechanisms of Bacterial Adhesion to Biomaterials and of Techniques Used in Estimating Bacteria-Material Interactions. Eur. Cell Mater. 2004, 8, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An Emergent Form of Bacterial Life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.; Falholt, P.; Gram, L. Enzymatic Removal and Disinfection of Bacterial Biofilms. Appl. Environ. Microbiol. 1997, 63, 3724–3728. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.; Bik, E.M.; DiGiulio, D.B.; Relman, D.A.; Brown, P.O. Development of the Human Infant Intestinal Microbiota. PLoS Biol. 2007, 5, 1556–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goode, K.R.; Asteriadou, K.; Robbins, P.T.; Fryer, P.J. Fouling and Cleaning Studies in the Food and Beverage Industry Classified by Cleaning Type. Compr. Rev. Food Sci. Food Saf. 2013, 12, 121–143. [Google Scholar] [CrossRef]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the Mechanism of Bacterial Biofilms Resistance to Antimicrobial Agents. Open Microbiol. J. 2017, 11, 53. [Google Scholar] [CrossRef]
- Goller, C.C.; Romeo, T. Environmental Influences on Biofilm Development. Curr. Top Microbiol. Immunol. 2008, 322, 37–66. [Google Scholar] [CrossRef]
- McDermott, P.F.; Walker, R.D.; White, D.G. Antimicrobials: Modes of Action and Mechanisms of Resistance. Int. J. Toxicol. 2003, 22, 135–143. [Google Scholar] [CrossRef]
- Martínez, J.L.; Rojo, F. Metabolic Regulation of Antibiotic Resistance. FEMS Microbiol. Rev. 2011, 35, 768–789. [Google Scholar] [CrossRef]
- Matle, I.; Mbatha, K.R.; Madoroba, E. A Review of Listeria Monocytogenes from Meat and Meat Products: Epidemiology, Virulence Factors, Antimicrobial Resistance and Diagnosis. Onderstepoort J. Vet. Res. 2020, 87, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Pusic, P.; Sonnleitner, E.; Bläsi, U. Specific and Global RNA Regulators in Pseudomonas Aeruginosa. Int. J. Mol. Sci. 2021, 22, 8632. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.; Li, L.; Girodat, D.; Förstner, K.U.; Said, N.; Corcoran, C.; Śmiga, M.; Papenfort, K.; Reinhardt, R.; Wieden, H.J.; et al. In Vivo Cleavage Map Illuminates the Central Role of RNase E in Coding and Non-Coding RNA Pathways. Mol. Cell 2017, 65, 39–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göpel, Y.; Papenfort, K.; Reichenbach, B.; Vogel, J.; Görke, B. Targeted Decay of a Regulatory Small RNA by an Adaptor Protein for RNase E and Counteraction by an Anti-Adaptor RNA. Genes Dev. 2013, 27, 552–564. [Google Scholar] [CrossRef] [Green Version]
- Fritsche, T.R.; Castanheira, M.; Miller, G.H.; Jones, R.N.; Armstrong, E.S. Detection of Methyltransferases Conferring High-Level Resistance to Aminoglycosides in Enterobacteriaceae from Europe, North America, and Latin America. Antimicrob. Agents Chemother. 2008, 52, 1843–1845. [Google Scholar] [CrossRef] [Green Version]
- Sidoli, S.; Kulej, K.; Garcia, B.A. Why Proteomics Is Not the New Genomics and the Future of Mass Spectrometry in Cell Biology. J. Cell Biol. 2017, 216, 21. [Google Scholar] [CrossRef] [Green Version]
- Manafi, L.; Aliakbarlu, J.; Dastmalchi Saei, H. Antibiotic Resistance and Biofilm Formation Ability of Salmonella Serotypes Isolated from Beef, Mutton, and Meat Contact Surfaces at Retail. J. Food Sci. 2020, 85, 2516–2522. [Google Scholar] [CrossRef]
- Hur, J.; Choi, Y.Y.; Park, J.H.; Jeon, B.W.; Lee, H.S.; Kim, A.R.; Lee, J.H. Antimicrobial Resistance, Virulence-Associated Genes, and Pulsed-Field Gel Electrophoresis Profiles of Salmonella Enterica Subsp. Enterica Serovar Typhimurium Isolated from Piglets with Diarrhea in Korea. Can. J. Vet. Res. 2011, 75, 49. [Google Scholar]
- Anjum, M.F.; Duggett, N.A.; AbuOun, M.; Randall, L.; Nunez-Garcia, J.; Ellis, R.J.; Rogers, J.; Horton, R.; Brena, C.; Williamson, S.; et al. Colistin Resistance in Salmonella and Escherichia Coli Isolates from a Pig Farm in Great Britain. J. Antimicrob. Chemother. 2016, 71, 2306–2313. [Google Scholar] [CrossRef] [Green Version]
- Coldham, N.G.; Randall, L.P.; Piddock, L.J.V.; Woodward, M.J. Effect of Fluoroquinolone Exposure on the Proteome of Salmonella Enterica Serovar Typhimurium. J. Antimicrob. Chemother. 2006, 58, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, W.; Zhang, R.; Xu, J.; Wang, R.; Wang, L.; Zhao, X.; Li, J. First Acetyl-Proteome Profiling of Salmonella Typhimurium Revealed Involvement of Lysine Acetylation in Drug Resistance. Vet. Microbiol. 2018, 226, 1–8. [Google Scholar] [CrossRef]
- Karatzas, K.A.G.; Randall, L.P.; Webber, M.; Piddock, L.J.V.; Humphrey, T.J.; Woodward, M.J.; Coldham, N.G. Phenotypic and Proteomic Characterization of Multiply Antibiotic-Resistant Variants of Salmonella Enterica Serovar Typhimurium Selected Following Exposure to Disinfectants. Appl. Environ. Microbiol. 2008, 74, 1508. [Google Scholar] [CrossRef] [Green Version]
- Qi, C.; Sun, F.; Wei, Q.; Xu, J.; Li, R.; Zhang, L.; Lu, F.; Jiang, X.; Fu, H.; Zhang, C.; et al. Quantitative Phosphoproteomics Reveals the Effect of BaeSR and AcrB Genes on Protein Phosphorylation in Salmonella Enterica Serovar Typhimurium. Res. Microbiol. 2022, 173, 103886. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Omar, M.; Nagaraja, K.V.; Goyal, S.M.; Vidovic, S. Novel Insight into the Effects of Cpxr on Salmonella Enteritidis Cells during the Chlorhexidine Treatment and Non-Stressful Growing Conditions. Int. J. Mol. Sci. 2021, 22, 8938. [Google Scholar] [CrossRef] [PubMed]
- Fillgrove, K.L.; Pakhomova, S.; Schaab, M.R.; Newcomer, M.E.; Armstrong, R.N. Structure and Mechanism of the Genomically Encoded Fosfomycin Resistance Protein, FosX, from Listeria Monocytogenes. Biochemistry 2007, 46, 8110–8120. [Google Scholar] [CrossRef] [PubMed]
- Bergholz, T.M.; Tang, S.; Wiedmann, M.; Boor, K.J. Nisin Resistance of Listeria Monocytogenes Is Increased by Exposure to Salt Stress and Is Mediated via LiaR. Appl. Environ. Microbiol. 2013, 79, 5682–5688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiedler, S.; Bender, J.K.; Klare, I.; Halbedel, S.; Grohmann, E.; Szewzyk, U.; Werner, G. Tigecycline Resistance in Clinical Isolates of Enterococcus Faecium Is Mediated by an Upregulation of Plasmid-Encoded Tetracycline Determinants Tet(L) and Tet(M). J. Antimicrob. Chemother. 2016, 71, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Kode, D.; Nannapaneni, R.; Chang, S. Low-Level Tolerance to Antibiotic Trimethoprim in QAC-Adapted Subpopulations of Listeria Monocytogenes. Foods 2021, 10, 1800. [Google Scholar] [CrossRef]
- Yan, S.; Li, M.; Luque-Sastre, L.; Wang, W.; Hu, Y.; Peng, Z.; Dong, Y.; Gan, X.; Nguyen, S.; Anes, J.; et al. Susceptibility (Re)-Testing of a Large Collection of Listeria Monocytogenes from Foods in China from 2012 to 2015 and WGS Characterization of Resistant Isolates. J. Antimicrob. Chemother. 2019, 74, 1786–1794. [Google Scholar] [CrossRef]
- Kovacevic, J.; Ziegler, J.; Walecka-Zacharska, E.; Reimer, A.; Kitts, D.D.; Gilmour, M.W. Tolerance of Listeria Monocytogenes to Quaternary Ammonium Sanitizers Is Mediated by a Novel Efflux Pump Encoded by EmrE. Appl. Environ. Microbiol. 2016, 82, 939–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Olsen, R.H.; Shi, L.; Ye, L.; He, J.; Meng, H. Characterization of a Plasmid Carrying Cat, ErmB and TetS Genes in a Foodborne Listeria Monocytogenes Strain and Uptake of the Plasmid by Cariogenic Streptococcus Mutans. Int. J. Food Microbiol. 2016, 238, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Lanza, V.F.; Duval, M.; Coque, T.M. Ecogenetics of Antibiotic Resistance in Listeria Monocytogenes. Mol. Microbiol. 2020, 113, 570–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawczyk-Balska, A.; Markiewicz, Z. The Intrinsic Cephalosporin Resistome of Listeria Monocytogenes in the Context of Stress Response, Gene Regulation, Pathogenesis and Therapeutics. J. Appl. Microbiol. 2016, 120, 251–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abril, A.G.; Carrera, M.; Böhme, K.; Barros-Velázquez, J.; Calo-Mata, P.; Sánchez-Pérez, A.; Villa, T.G. Proteomic Characterization of Antibiotic Resistance in Listeria and Production of Antimicrobial and Virulence Factors. Int. J. Mol. Sci. 2021, 22, 8141. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Suárez, J.V.; Ortiz, S.; López-Alonso, V. Potential Impact of the Resistance to Quaternary Ammonium Disinfectants on the Persistence of Listeria Monocytogenes in Food Processing Environments. Front. Microbiol. 2016, 7, 638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Diversity and Antibiotic Resistance in Pseudomonas Spp. from Drinking Water. Sci. Total Environ. 2012, 426, 366–374. [Google Scholar] [CrossRef]
- Ranjan, V.K.; Mukherjee, S.; Basak, C.; Chakraborty, R. Abundance of New Delhi Metallo-β-Lactamase-Producing Acinetobacter, Escherichia, Proteus, and Pseudomonas Spp. in Mahananda and Karala Rivers of India. Microb. Drug Resist. 2021, 27, 1603–1615. [Google Scholar] [CrossRef]
- Ghosh, A.; Saran, N.; Saha, S. Survey of Drug Resistance Associated Gene Mutations in Mycobacterium Tuberculosis, ESKAPE and Other Bacterial Species. Sci. Rep. 2020, 10, 8957. [Google Scholar] [CrossRef]
- Meng, L.; Liu, H.; Lan, T.; Dong, L.; Hu, H.; Zhao, S.; Zhang, Y.; Zheng, N.; Wang, J. Antibiotic Resistance Patterns of Pseudomonas Spp. Isolated From Raw Milk Revealed by Whole Genome Sequencing. Front. Microbiol. 2020, 11, 1005. [Google Scholar] [CrossRef]
- Aoike, N.; Saga, T.; Sakata, R.; Yoshizumi, A.; Kimura, S.; Iwata, M.; Yoshizawa, S.; Sugasawa, Y.; Ishii, Y.; Yamaguchi, K.; et al. Molecular Characterization of Extraintestinal Escherichia Coli Isolates in Japan: Relationship between Sequence Types and Mutation Patterns of Quinolone Resistance-Determining Regions Analyzed by Pyrosequencing. J. Clin. Microbiol. 2013, 51, 1692–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatzeva-Topalova, P.Z.; May, A.P.; Sousa, M.C. Structure and Mechanism of ArnA: Conformational Change Implies Ordered Dehydrogenase Mechanism in Key Enzyme for Polymyxin Resistance. Structure 2005, 13, 929–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengoechea, J.A.; Skurnik, M. Temperature-Regulated Efflux Pump/Potassium Antiporter System Mediates Resistance to Cationic Antimicrobial Peptides in Yersinia. Mol. Microbiol. 2000, 37, 67–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaker, M.; Spanogiannopoulos, P.; Wright, G.D. The Tetracycline Resistome. Cell. Mol. Life Sci. 2010, 67, 419–431. [Google Scholar] [CrossRef]
- Mima, T.; Joshi, S.; Gomez-Escalada, M.; Schweizer, H.P. Identification and Characterization of TriABC-OpmH, a Triclosan Efflux Pump of Pseudomonas Aeruginosa Requiring Two Membrane Fusion Proteins. J. Bacteriol. 2007, 189, 7600–7609. [Google Scholar] [CrossRef] [Green Version]
- Muratovic, A.Z.; Hagstöm, T.; Rosén, J.; Granelli, K.; Hellenäs, K.E. Quantitative Analysis of Staphylococcal Enterotoxins A and B in Food Matrices Using Ultra High-Performance Liquid Chromatography Tandem Mass Spectrometry (UPLC-MS/MS). Toxins 2015, 7, 3637. [Google Scholar] [CrossRef] [Green Version]
- De Groote, V.N.; Verstraeten, N.; Fauvart, M.; Kint, C.I.; Verbeeck, A.M.; Beullens, S.; Cornelis, P.; Michiels, J. Novel Persistence Genes in Pseudomonas Aeruginosa Identified by High-Throughput Screening. FEMS Microbiol. Lett. 2009, 297, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Erdmann, J.; Thöming, J.G.; Pohl, S.; Pich, A.; Lenz, C.; Häussler, S. The Core Proteome of Biofilm-Grown Clinical Pseudomonas Aeruginosa Isolates. Cells 2019, 8, 1129. [Google Scholar] [CrossRef] [Green Version]
- Bao, K.D.; Letellier, A.; Beaudry, F. Analysis of Staphylococcus Enterotoxin B Using Differential Isotopic Tags and Liquid Chromatography Quadrupole Ion Trap Mass Spectrometry. Biomed. Chromatogr. 2012, 26, 1049–1057. [Google Scholar] [CrossRef]
- D’Alessandro, A.; Zolla, L. We Are What We Eat: Food Safety and Proteomics. J. Proteome Res. 2012, 11, 26–36. [Google Scholar] [CrossRef]
- Andjelkovic, M.; Tsilia, V.; Rajkovic, A.; de Cremer, K.; Van Loco, J. Application of LC-MS/MS MRM to Determine Staphylococcal Enterotoxins (SEB and SEA) in Milk. Toxins 2016, 8, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilgisonis, E.V.; Kopylov, A.T.; Ponomarenko, E.A.; Poverennaya, E.V.; Tikhonova, O.V.; Farafonova, T.E.; Novikova, S.; Lisitsa, A.V.; Zgoda, V.G.; Archakov, A.I. Increased Sensitivity of Mass Spectrometry by Alkaline Two-Dimensional Liquid Chromatography: Deep Cover of the Human Proteome in Gene-Centric Mode. J. Proteome Res. 2018, 17, 4258–4266. [Google Scholar] [CrossRef] [PubMed]
- Dupré, M.; Gilquin, B.; Fenaille, F.; Feraudet-Tarisse, C.; Dano, J.; Ferro, M.; Simon, S.; Junot, C.; Brun, V.; Becher, F. Multiplex Quantification of Protein Toxins in Human Biofluids and Food Matrices Using Immunoextraction and High-Resolution Targeted Mass Spectrometry. Anal. Chem. 2015, 87, 8473–8480. [Google Scholar] [CrossRef] [PubMed]
- Afzaal, M.; Saeed, F.; Hussain, M.; Shahid, F.; Siddeeg, A.; Al-Farga, A. Proteomics as a Promising Biomarker in Food Authentication, Quality and Safety: A Review. Food Sci. Nutr. 2022, 10, 2333–2346. [Google Scholar] [CrossRef] [PubMed]
- Jagadeesh, D.S.; Kannegundla, U.; Reddy, R.K. Application of Proteomic Tools in Food Quality and Safety. Adv. Anim. Vet. Sci. 2017, 5, 213–225. [Google Scholar]
- Stryiński, R.; Łopieńska-Biernat, E.; Carrera, M. Proteomic Insights into the Biology of the Most Important Foodborne Parasites in Europe. Foods 2020, 9, 1403. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Organism | Marker (Gene/Protein) | UniProt ID | Resistance | Reference |
|---|---|---|---|---|
| Salmonella enterica | tetA, tetG and tetC, TetR | A0A1Z1VWZ4 *, A3RLS9 *, A0A3G1TUX2 *, A0A1Z1VX01 * | Tetracycline | [51,52] |
| gyrA | A0A447PDY4 * | Nalidixic acid | [51] | |
| mcr-1 | A0A1P8DNG0 * | Colistin | [53] | |
| bla(PSE) and bla(TEM) | K9M2D2 *, A0A7D7QQ45 * | Ampicillin, penicillin antibiotics | [52] | |
| cat1, cat2 and floR | Q3HNN0, Q7BD42, A0A3K0TB41 * | Chloramphenicol | [52] | |
| strA, strB and aadA | A0A5C2D198 *, C4NVB8 *, Q02865 | Streptomycin | [52] | |
| sul1 and sul2 | A0A0F6NWV0 *, A0A1S6KR61 * | Sulfizoxazole | [52] | |
| AcrAB/TolC, including acetylated lysine (PTM) | A0A759DN94 * | Fluoroquinolone | [54] | |
| Quaternary ammonium compounds, chloramphenicol, tetracycline, ampicillin | [55,56] | |||
| AcrB | A0A379QMS3 * | Ciprofloxacin | [57] | |
| FabI | A0A5Z3DTY7 * | Triclosan | [56] | |
| TolB, ElaB, TolC, GrxB, Tpx, Tsx, AhpF, AhpC, NfnB | A0A447MVS8 *, E8XEF3 *, Q54001, A0A6C8EYP6 *, A0A3F3I872 *, A0A379QQ98 *, A0A8E9PK94 *, A0A379WCI5 *, A0A447MWB6 * | Wide range of antimicrobial agents | [56] | |
| AcnA | A0A711ME57 * | Quaternary ammonium compounds | [56] | |
| RpoE, CpxR | D0ZSY9 *, A0A5Z8M962 * | Chlorhexidine | [58] | |
| csgD, bcsA, ardA | O54294, A0A2T8TBA6 *, A0A410J986 * | Quaternary ammonium compounds | [31] | |
| Listeria monocytogenes | FosX | Q8Y6I2 | Fosfomycin | [59] |
| FosE | A0A3T2HNE9 * | Tetronazine | ||
| FosI | A0A5M3ENG4 * | Bleomycin | ||
| LiaR | A0A0E1R5S4 * | Nisin | [60] | |
| TetM, TetS | Q5WMA8, Q48791 | Tetracyclines, tellurite | [61] | |
| dfrD, dfrG, dhfr | Q79CE5 | Trimethoprim | [62] | |
| ErmA(TR), ErmB, ErmC | K4NRN0 *, K4NU76 * | Erythromycin, quaternary ammonium compounds | [63,64] | |
| lnuB | A0A4D6D220 * | Lincosamide | [65] | |
| PBP1 | Q8Y614 | Cephalosporins | [66,67] | |
| LysR, LytR, LytTR, Rgg | A0A5M2ZBL1 *, A0A6W3T4B1 *, A0A5Y7CVQ1 *, A0A3H2VUB6 * | Cationic antimicrobial peptides (CAMP) | [68] | |
| MerR | A0A3T2B4E2 * | Mercury resistance | [61] | |
| bcrA, bcrB, bcrC, qacH, qacA | A0A5Y1L6S6 *, I6ZWK8 *, I7B1C4 *, T2KSX9 *, T1YPL4 * | Quaternary ammonium compounds | [69] | |
| Pseudomonas aeruginosa | gyrB | P13364 | Aminocoumarin | [70] |
| aadA, rpsL | A0A844NVA2 *, Q9HWD0 | Aminoglycosides | [71,72] | |
| blaCTX–M, PBP1a, PBP1b, oprD | A0A0M4CJ048, Q07806, A0A165VXD8 *, P32722 | Beta lactams | [73] | |
| gyrA, gyrB, patA, patB, parC, parE, emrA, emrB, mdtK, mfd | P48372, P13364, A0A5E7MVR1 *, Q5BU34, Q9HUK1, Q9HUJ8, A0A5E7FSX8 *, A0A5M9IUB7 *, A0A0B7DI98 *, Q9HZK3 | Fluoroquinolones | [73,74] | |
| mdtD, mdtG, mdtH, glpT, murA | A0A5E6SQR7 *, A0A5E6YH70 *, A0A5E6QA20 *, Q9HTV5, Q9Z3Z6 | Fosfomycin | [73] | |
| vanA | O05616 | Glycopeptide antibiotics | [73] | |
| arnA, bacA, bcrA, liaR, mprF, phoP, phoQ, pmrA, pmrB, pmrE, pmrF, rosB, floR, lpxA, lpxC, cls, pgsA, rpoC | Q02R25, Q02LA5, Q937U9, A0A8D9KX77 *, A0A1B5E8X0 *, Q9I4F9, Q9I4F8, Q9HV32, Q9HV31, A0A0C6ED02 *, A0A519EUM6 *, A0A485GZQ4 *, A0A4D6RIK8 *, Q9X6P4, P47205, A0A2K9MAE4 *, P45419, P19176 | Lipopeptides | [73,75,76], | |
| carA, macA, macB | Q88DU5, A0A5E7QUJ5 *, Q88F88 | Macrolides | [73] | |
| mefA | A0A383RT01 * | Macrolide-Lincosamide-Streptogramin B | [73] | |
| catA1, catB3, catI, floR | T2HGZ6 *, V5LZV8 *, Q8VPF3, A0A4D6RIK8 * | Fenicol | [73] | |
| rpoB | P19175 | Rifampin | [73] | |
| sul1 | A0A2R4S1K0 * | Sulfonamides | [73] | |
| tetA, tetB(P), tetG, tetM | A0A4P2USE6 *, A0A4Y5T9V5 *, Q9X685, A0MZ57 * | Tetracycline | [73,77] | |
| katG, kasA | Q88GQ0, A0A379J0D7 * | Isoniazid | [73] | |
| triA, triB, triC, opmH | P72156, A0A8F9V618 *, A0A8G1K8C1 *, A0A8G6KPN1 * | Triclosan | [73,78] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarbeeva, S.; Kozlova, A.; Sarygina, E.; Kiseleva, O.; Ponomarenko, E.; Ilgisonis, E. Food for Thought: Proteomics for Meat Safety. Life 2023, 13, 255. https://doi.org/10.3390/life13020255
Tarbeeva S, Kozlova A, Sarygina E, Kiseleva O, Ponomarenko E, Ilgisonis E. Food for Thought: Proteomics for Meat Safety. Life. 2023; 13(2):255. https://doi.org/10.3390/life13020255
Chicago/Turabian StyleTarbeeva, Svetlana, Anna Kozlova, Elizaveta Sarygina, Olga Kiseleva, Elena Ponomarenko, and Ekaterina Ilgisonis. 2023. "Food for Thought: Proteomics for Meat Safety" Life 13, no. 2: 255. https://doi.org/10.3390/life13020255