Figure 1.
Bony structures important for this study. Photos of macerated bones from a male albino rat were taken to provide an overview of the bony structures important for this study. Isolated right hip bone from the lateral (a) and ventral (b) aspects. Isolated right femur from the caudal (c) and medial (d) aspects. Isolated right tibia and fibula from the cranial (e) and medial (f) aspects. a = acetabulum, cb = cranial border of the tibia, cpr = caudal pubic ramus, crpr = cranial pubic ramus, f = fibula, gt = greater trochanter, it = ischial tuber, ll-fa = lateral lip of facies aspera, lst = lateral supracondylar tuberosity, lt = lesser trochanter, mc = medial (tibial) condyle, me = medial (femoral) epicondyle, ml-fa = medial lip of facies aspera, ms = medial surface of tibia, mst = medial supracondylar tuberosity, of = obturator foramen, ri = ramus of ischium, t = tibial tuberostiy, ti = tabula of ischium, tt = third trochanter, asterisk = trochanteric fossa.
Figure 1.
Bony structures important for this study. Photos of macerated bones from a male albino rat were taken to provide an overview of the bony structures important for this study. Isolated right hip bone from the lateral (a) and ventral (b) aspects. Isolated right femur from the caudal (c) and medial (d) aspects. Isolated right tibia and fibula from the cranial (e) and medial (f) aspects. a = acetabulum, cb = cranial border of the tibia, cpr = caudal pubic ramus, crpr = cranial pubic ramus, f = fibula, gt = greater trochanter, it = ischial tuber, ll-fa = lateral lip of facies aspera, lst = lateral supracondylar tuberosity, lt = lesser trochanter, mc = medial (tibial) condyle, me = medial (femoral) epicondyle, ml-fa = medial lip of facies aspera, ms = medial surface of tibia, mst = medial supracondylar tuberosity, of = obturator foramen, ri = ramus of ischium, t = tibial tuberostiy, ti = tabula of ischium, tt = third trochanter, asterisk = trochanteric fossa.
Figure 2.
Muscle attachments of the adductor group. Origin and insertion areas of the adductor muscles are color-coded on photographs of isolated macerated bones from a male albino rat. The caudal half of the hip bone is shown from the ventral aspect (a). The femur is shown from the caudal aspect (b). The proximal part of the tibia is shown from the cranial (c) and medial (d) aspects. Color key: dark blue—cranial gracilis muscle, light blue—caudal gracilis muscle, pink—adductor longus muscle, orange—adductor brevis muscle, light red—adductor magnus muscle, dark red—adductor minimus muscle, dark green—obturator externus muscle, light green—obturator intermedius muscle.
Figure 2.
Muscle attachments of the adductor group. Origin and insertion areas of the adductor muscles are color-coded on photographs of isolated macerated bones from a male albino rat. The caudal half of the hip bone is shown from the ventral aspect (a). The femur is shown from the caudal aspect (b). The proximal part of the tibia is shown from the cranial (c) and medial (d) aspects. Color key: dark blue—cranial gracilis muscle, light blue—caudal gracilis muscle, pink—adductor longus muscle, orange—adductor brevis muscle, light red—adductor magnus muscle, dark red—adductor minimus muscle, dark green—obturator externus muscle, light green—obturator intermedius muscle.
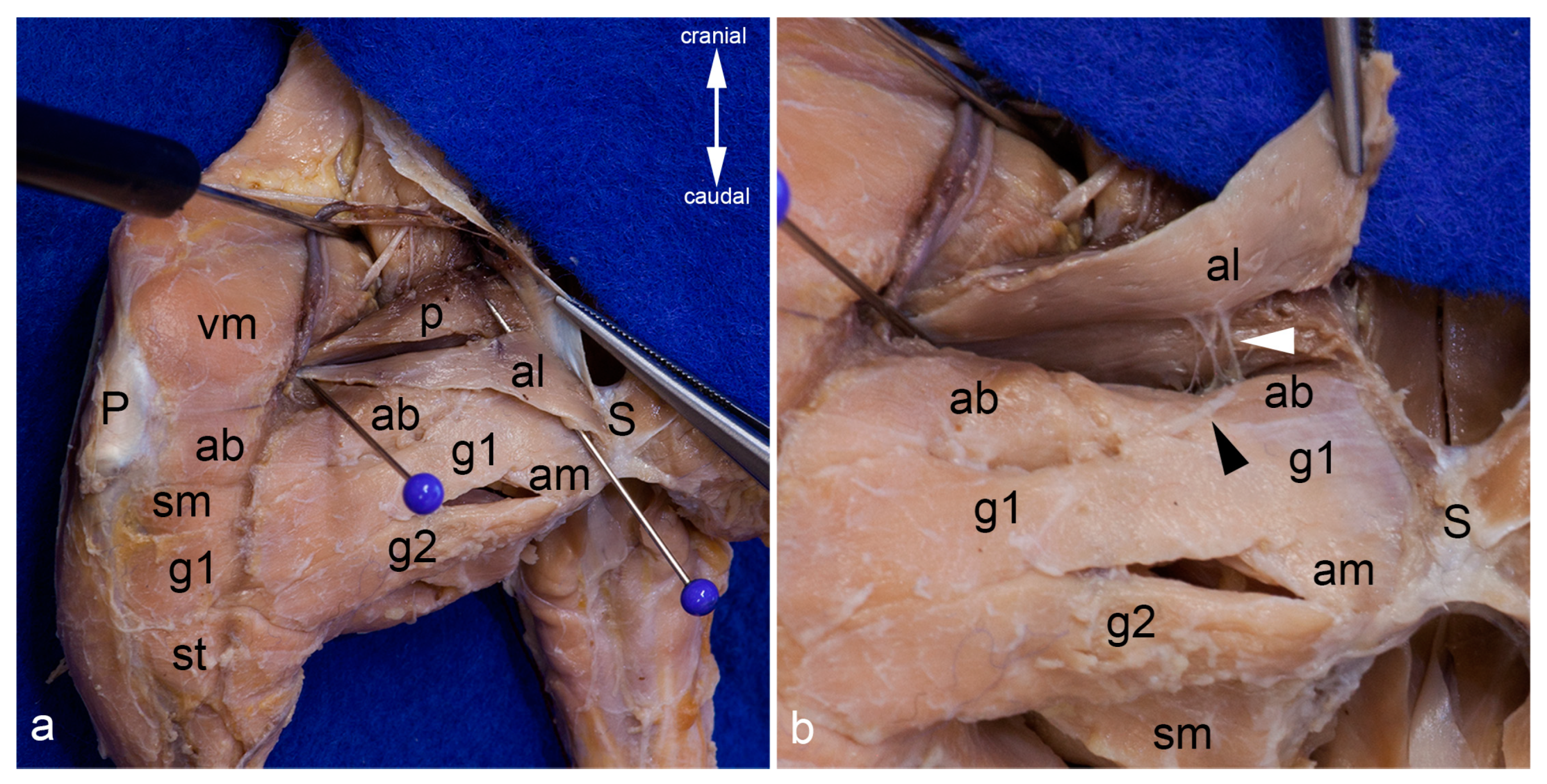
Figure 3.
Gracilis muscles. The adductor longus muscle was detached from its origin and reflected towards its insertion. Thereby its innervating twig from the cranial branch of the obturator nerve has been cut. The cranial gracilis muscle can be observed in toto from the medial aspect of the thigh (a). The cranial gracilis muscle was also detached from its origin and reflected laterally after transecting its innervating twig from the cranial branch of the obturator nerve. The caudal gracilis muscle can be observed in toto (b). In both pictures, the thigh was abducted in the hip joint to show the medial aspect of the thigh. The red pins highlight the origin, the blue pins the insertion, and the white pins the innervation of the gracilis muscles. ab = adductor brevis muscle, al = adductor longus muscle, am = adductor magnus muscle, g1 = cranial gracilis muscle, g2 = caudal gracilis muscle, P = patella/patellar ligament, p = pectineus muscle, S = pelvic symphysis, sm = semimembranosus muscle, st = semitendinosus muscle, vm = vastus medialis muscle.
Figure 3.
Gracilis muscles. The adductor longus muscle was detached from its origin and reflected towards its insertion. Thereby its innervating twig from the cranial branch of the obturator nerve has been cut. The cranial gracilis muscle can be observed in toto from the medial aspect of the thigh (a). The cranial gracilis muscle was also detached from its origin and reflected laterally after transecting its innervating twig from the cranial branch of the obturator nerve. The caudal gracilis muscle can be observed in toto (b). In both pictures, the thigh was abducted in the hip joint to show the medial aspect of the thigh. The red pins highlight the origin, the blue pins the insertion, and the white pins the innervation of the gracilis muscles. ab = adductor brevis muscle, al = adductor longus muscle, am = adductor magnus muscle, g1 = cranial gracilis muscle, g2 = caudal gracilis muscle, P = patella/patellar ligament, p = pectineus muscle, S = pelvic symphysis, sm = semimembranosus muscle, st = semitendinosus muscle, vm = vastus medialis muscle.
![Life 13 02096 g003]()
Figure 4.
Adductor longus muscle. The adductor longus muscle is shown in situ in the medial aspect of the thigh. The blue pins highlight its attachments. The inguinal ligament is elevated by a clamp to show the broad lacunar ligament (a). The adductor longus muscle was mobilized from its origin to show its innervation from the cranial branch of the obturator nerve (white arrowhead). The black arrowhead indicates the course of the cranial branch of the obturator nerve (b). In both pictures, the thigh was abducted in the hip joint. ab = adductor brevis muscle, al = adductor longus muscle, am = adductor magnus muscle, g1 = cranial gracilis muscle g2 = caudal gracilis muscle, P = patella / patellar ligament, p = pectineus muscle, S = pelvic symphysis, sm = semimembranosus muscle, st = semitendinosus muscle, vm = vastus medialis muscle.
Figure 4.
Adductor longus muscle. The adductor longus muscle is shown in situ in the medial aspect of the thigh. The blue pins highlight its attachments. The inguinal ligament is elevated by a clamp to show the broad lacunar ligament (a). The adductor longus muscle was mobilized from its origin to show its innervation from the cranial branch of the obturator nerve (white arrowhead). The black arrowhead indicates the course of the cranial branch of the obturator nerve (b). In both pictures, the thigh was abducted in the hip joint. ab = adductor brevis muscle, al = adductor longus muscle, am = adductor magnus muscle, g1 = cranial gracilis muscle g2 = caudal gracilis muscle, P = patella / patellar ligament, p = pectineus muscle, S = pelvic symphysis, sm = semimembranosus muscle, st = semitendinosus muscle, vm = vastus medialis muscle.
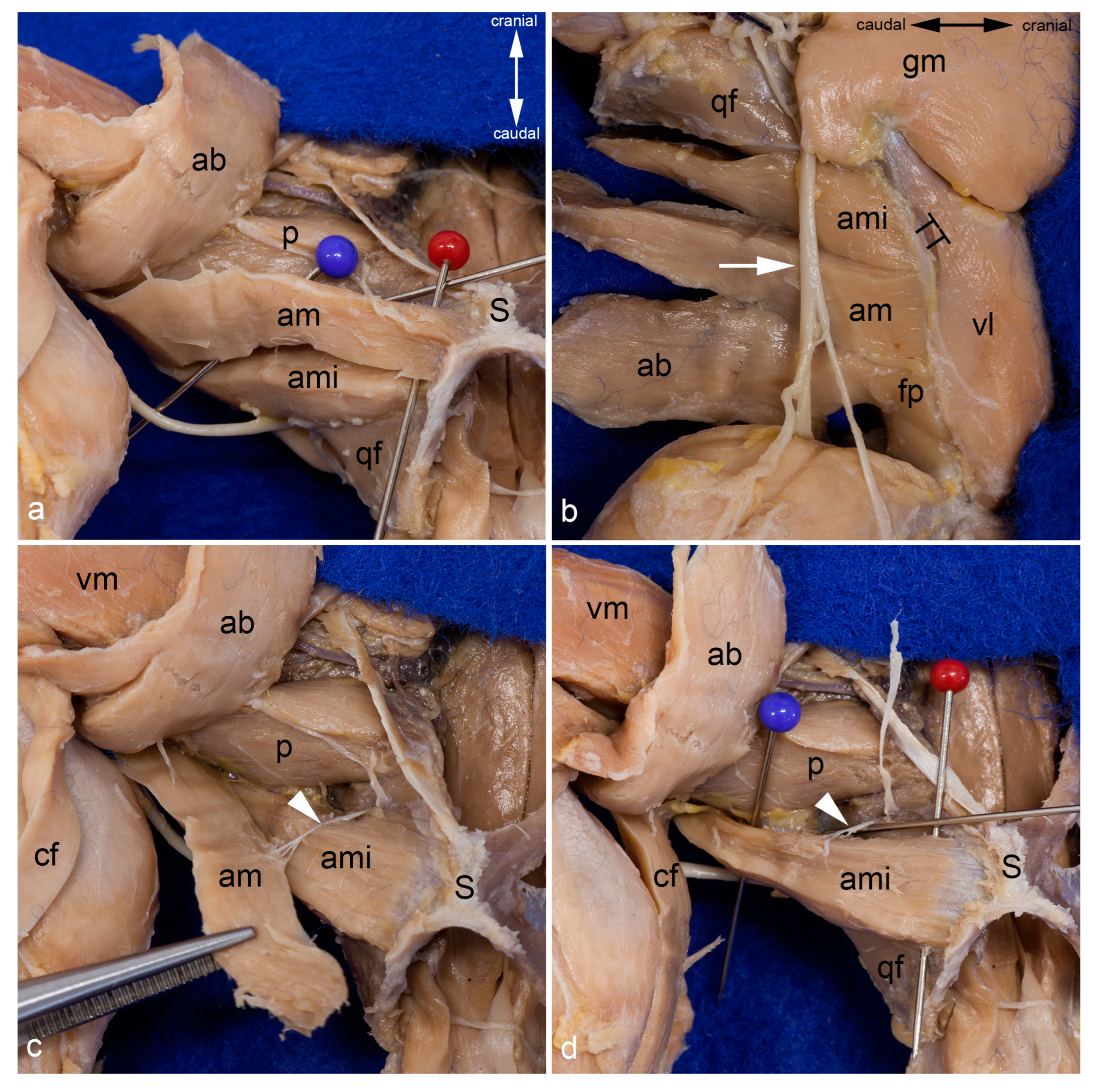
Figure 5.
Adductor brevis muscle. The adductor longus and both gracilis muscles were reflected laterally to show the adductor brevis muscle within the medial aspect of the thigh. The red pin highlights its origin and the blue pins its insertion; only the insertion of the genicular part is visible. The white arrowhead indicates the course of the cranial branch of the obturator nerve (a). The adductor brevis muscle was detached from its origin and pulled distally. The insertion of its femoral part can be observed bordered by the blue pins (b). In both pictures, the thigh was abducted in the hip joint. ab = adductor brevis muscle, al = adductor longus muscle, am = adductor magnus muscle, ami = adductor minimus muscle, fp = femoral part of adductor brevis muscle, gp = genicular part of adductor brevis muscle, P = patella / patellar ligament, p = pectineus muscle, S = pelvic symphysis, sm = semimembranosus muscle, st = semitendinosus muscle, vm = vastus medialis muscle.
Figure 5.
Adductor brevis muscle. The adductor longus and both gracilis muscles were reflected laterally to show the adductor brevis muscle within the medial aspect of the thigh. The red pin highlights its origin and the blue pins its insertion; only the insertion of the genicular part is visible. The white arrowhead indicates the course of the cranial branch of the obturator nerve (a). The adductor brevis muscle was detached from its origin and pulled distally. The insertion of its femoral part can be observed bordered by the blue pins (b). In both pictures, the thigh was abducted in the hip joint. ab = adductor brevis muscle, al = adductor longus muscle, am = adductor magnus muscle, ami = adductor minimus muscle, fp = femoral part of adductor brevis muscle, gp = genicular part of adductor brevis muscle, P = patella / patellar ligament, p = pectineus muscle, S = pelvic symphysis, sm = semimembranosus muscle, st = semitendinosus muscle, vm = vastus medialis muscle.
Figure 6.
Adductor magnus and minimus muscles. The course of the adductor magnus muscle is shown from the medial aspect. The pins separate it from the adductor minimus muscle (a). The insertion of the adductor minimus and adductor magnus muscles can also be observed from the lateral aspect of the thigh, after releasing all hamstring muscles, the femorococcygeus muscle, and the gluteus superficialis muscle. The adductor muscles were separated from the vastus lateralis muscle by the lateral femoral intermuscular septum. The sciatic nerve (white arrow) was in contact only with the adductor minimus muscle as the caudofemoralis and biceps femoris muscles (already removed for this photograph) were situated between the nerve and the adductor magnus and brevis muscles (b). For this photograph the adductor magnus muscle was also released from its origin and pulled distally to show its innervation by the caudal branch of the obturator nerve (white arrowhead) (c). The adductor magnus muscle was removed to show the course of the adductor minimus muscle from the medial aspect. The red pin is inserted near its origin and the blue pin near its insertion. Its innervation by the caudal branch of the obturator nerve (white arrowhead) can be observed. The cranial branch of the obturator nerve was reflected cranially (d). In all pictures, almost all superficial muscles were detached from their origin and reflected towards their insertion or totally removed. In (a,c,d), the thigh was abducted in the hip joint. ab = adductor brevis muscle, am = adductor magnus muscle, ami = adductor minimus muscle, cf = caudofemoralis muscle, fp = femoral part of adductor brevis muscle, gm = gluteus medius muscle, p = pectineus muscle, qf = quadratus femoris muscle, S = pelvic symphysis, TT = third trochanter, vl = vastus lateralis muscle, vm = vastus medialis muscle.
Figure 6.
Adductor magnus and minimus muscles. The course of the adductor magnus muscle is shown from the medial aspect. The pins separate it from the adductor minimus muscle (a). The insertion of the adductor minimus and adductor magnus muscles can also be observed from the lateral aspect of the thigh, after releasing all hamstring muscles, the femorococcygeus muscle, and the gluteus superficialis muscle. The adductor muscles were separated from the vastus lateralis muscle by the lateral femoral intermuscular septum. The sciatic nerve (white arrow) was in contact only with the adductor minimus muscle as the caudofemoralis and biceps femoris muscles (already removed for this photograph) were situated between the nerve and the adductor magnus and brevis muscles (b). For this photograph the adductor magnus muscle was also released from its origin and pulled distally to show its innervation by the caudal branch of the obturator nerve (white arrowhead) (c). The adductor magnus muscle was removed to show the course of the adductor minimus muscle from the medial aspect. The red pin is inserted near its origin and the blue pin near its insertion. Its innervation by the caudal branch of the obturator nerve (white arrowhead) can be observed. The cranial branch of the obturator nerve was reflected cranially (d). In all pictures, almost all superficial muscles were detached from their origin and reflected towards their insertion or totally removed. In (a,c,d), the thigh was abducted in the hip joint. ab = adductor brevis muscle, am = adductor magnus muscle, ami = adductor minimus muscle, cf = caudofemoralis muscle, fp = femoral part of adductor brevis muscle, gm = gluteus medius muscle, p = pectineus muscle, qf = quadratus femoris muscle, S = pelvic symphysis, TT = third trochanter, vl = vastus lateralis muscle, vm = vastus medialis muscle.
![Life 13 02096 g006]()
Figure 7.
Obturator externus and intermedius muscles. The obturator externus and obturator intermedius muscles are shown from the ventrolateral aspect. Between them, the cranial and caudal branches of the obturator nerve exit (a). The obturator externus and intermedius muscles have been detached from their insertion and partly mobilized. Thus, their deep sites of origin can be observed from the ventrolateral aspect. The nerve twig to the obturator intermedius muscle was transected during mobilization (b). The course and insertion of the obturator externus muscle are shown from the dorsal aspect. The obturator intermedius muscle is located ventro-caudally and thus covered by the course of the obturator externus muscle (c). The obturator intermedius muscle has been partly mobilized to show the proper nerve twigs for each muscle (white arrowhead). The cranial branch of the obturator nerve was reflected cranially (d). In all pictures, almost all superficial muscles were detached from their origin and reflected towards their insertion or totally removed. The thigh was abducted and fully rotated laterally in (a,b,d). g = gemellus muscle, GT = greater trochanter, hc = capsule of the hip joint, ip= iliacus and psoas muscles, IT = ischial tuber, LT = lesser trochanter, oe = obturator externus muscle, oim = obturator intermedius muscle, om = obturator membrane, S = pelvic symphysis, white arrow = sciatic nerve.
Figure 7.
Obturator externus and intermedius muscles. The obturator externus and obturator intermedius muscles are shown from the ventrolateral aspect. Between them, the cranial and caudal branches of the obturator nerve exit (a). The obturator externus and intermedius muscles have been detached from their insertion and partly mobilized. Thus, their deep sites of origin can be observed from the ventrolateral aspect. The nerve twig to the obturator intermedius muscle was transected during mobilization (b). The course and insertion of the obturator externus muscle are shown from the dorsal aspect. The obturator intermedius muscle is located ventro-caudally and thus covered by the course of the obturator externus muscle (c). The obturator intermedius muscle has been partly mobilized to show the proper nerve twigs for each muscle (white arrowhead). The cranial branch of the obturator nerve was reflected cranially (d). In all pictures, almost all superficial muscles were detached from their origin and reflected towards their insertion or totally removed. The thigh was abducted and fully rotated laterally in (a,b,d). g = gemellus muscle, GT = greater trochanter, hc = capsule of the hip joint, ip= iliacus and psoas muscles, IT = ischial tuber, LT = lesser trochanter, oe = obturator externus muscle, oim = obturator intermedius muscle, om = obturator membrane, S = pelvic symphysis, white arrow = sciatic nerve.
![Life 13 02096 g007]()
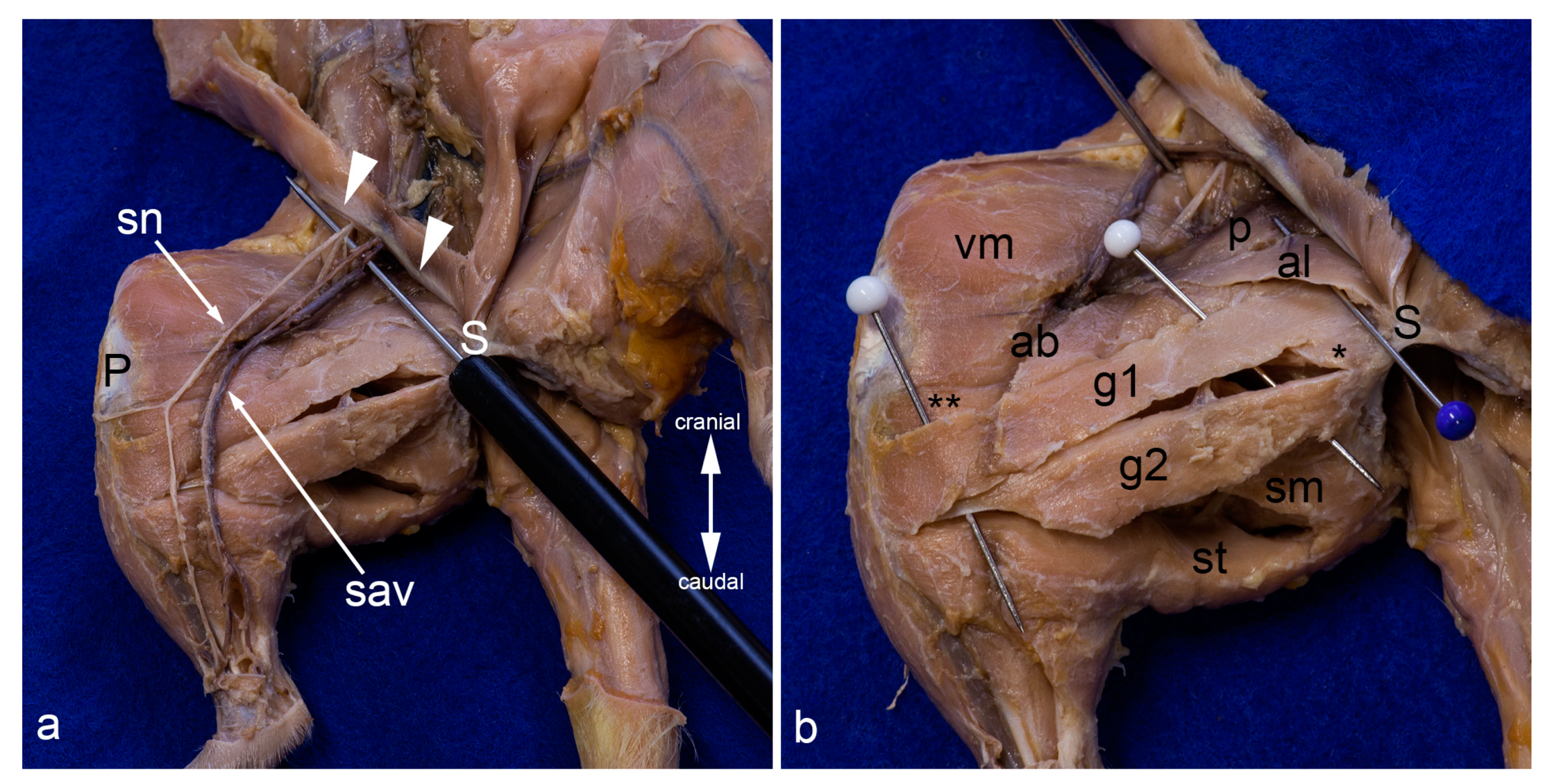
Figure 8.
Superficial layers in the medial aspect of the thigh. On the left leg, only the skin was removed. The superficial structures gleam through the fascia lata. On the right leg, the fascia lata was totally removed. Superficial to the probe, the femoral nerve, the femoral artery, and the femoral vein (from lateral to medial) and their proximal branches (saphenous nerve, saphenous artery, and medial saphenous vein) are visible. The abdominal wall was removed except for the caudal part forming the inguinal ligament (white arrowheads) (a). The superficial muscles in the medial aspect of the thigh were carefully separated by blunt dissection. The saphenous nerve, the saphenous artery, and the medial saphenous vein were partially removed or displaced laterally. The blue pin indicates the most superficial muscle in this region, the adductor longus muscle. The white pins highlight the cranial gracilis and caudal gracilis muscles (b). In both pictures, the thigh was abducted in the hip joint to show the medial aspect of the thigh. ab= adductor brevis muscle, al = adductor longus muscle, g1 = cranial gracilis muscle, g2 = caudal gracilis muscle, P = patella/patellar ligament, p = pectineus muscle, S = pelvic symphysis, sav = saphenous artery and medial saphenous vein), sm and ** = semimembranosus muscle, sn = saphenous nerve, st = semitendinosus muscle, vm = vastus medialis muscle, * = adductor magnus muscle.
Figure 8.
Superficial layers in the medial aspect of the thigh. On the left leg, only the skin was removed. The superficial structures gleam through the fascia lata. On the right leg, the fascia lata was totally removed. Superficial to the probe, the femoral nerve, the femoral artery, and the femoral vein (from lateral to medial) and their proximal branches (saphenous nerve, saphenous artery, and medial saphenous vein) are visible. The abdominal wall was removed except for the caudal part forming the inguinal ligament (white arrowheads) (a). The superficial muscles in the medial aspect of the thigh were carefully separated by blunt dissection. The saphenous nerve, the saphenous artery, and the medial saphenous vein were partially removed or displaced laterally. The blue pin indicates the most superficial muscle in this region, the adductor longus muscle. The white pins highlight the cranial gracilis and caudal gracilis muscles (b). In both pictures, the thigh was abducted in the hip joint to show the medial aspect of the thigh. ab= adductor brevis muscle, al = adductor longus muscle, g1 = cranial gracilis muscle, g2 = caudal gracilis muscle, P = patella/patellar ligament, p = pectineus muscle, S = pelvic symphysis, sav = saphenous artery and medial saphenous vein), sm and ** = semimembranosus muscle, sn = saphenous nerve, st = semitendinosus muscle, vm = vastus medialis muscle, * = adductor magnus muscle.
![Life 13 02096 g008]()
Figure 9.
Topographical relationships of the gracilis muscles. The adductor longus and cranial gracilis muscle were mobilized from their origin and reflected laterally. The course of the cranial branch of the obturator nerve (white pin) and its twigs for the cranial gracilis and caudal gracilis muscles are visualized (a). Both gracilis muscles were reflected laterally to show the bursae underneath their tendons in detail (b). In both pictures, the thigh was abducted in the hip joint to show the medial aspect of the thigh. ab = adductor brevis muscle, al = adductor longus muscle, am = adductor magnus muscle, g1 = cranial gracilis muscle, g2 = caudal gracilis muscle, P = patella / patellar ligament, p = pectineus muscle, S = pelvic symphysis, sm = semimembranosus muscle, st = semitendinosus muscle, T = medial surface of the tibia, vm = vastus medialis muscle.
Figure 9.
Topographical relationships of the gracilis muscles. The adductor longus and cranial gracilis muscle were mobilized from their origin and reflected laterally. The course of the cranial branch of the obturator nerve (white pin) and its twigs for the cranial gracilis and caudal gracilis muscles are visualized (a). Both gracilis muscles were reflected laterally to show the bursae underneath their tendons in detail (b). In both pictures, the thigh was abducted in the hip joint to show the medial aspect of the thigh. ab = adductor brevis muscle, al = adductor longus muscle, am = adductor magnus muscle, g1 = cranial gracilis muscle, g2 = caudal gracilis muscle, P = patella / patellar ligament, p = pectineus muscle, S = pelvic symphysis, sm = semimembranosus muscle, st = semitendinosus muscle, T = medial surface of the tibia, vm = vastus medialis muscle.
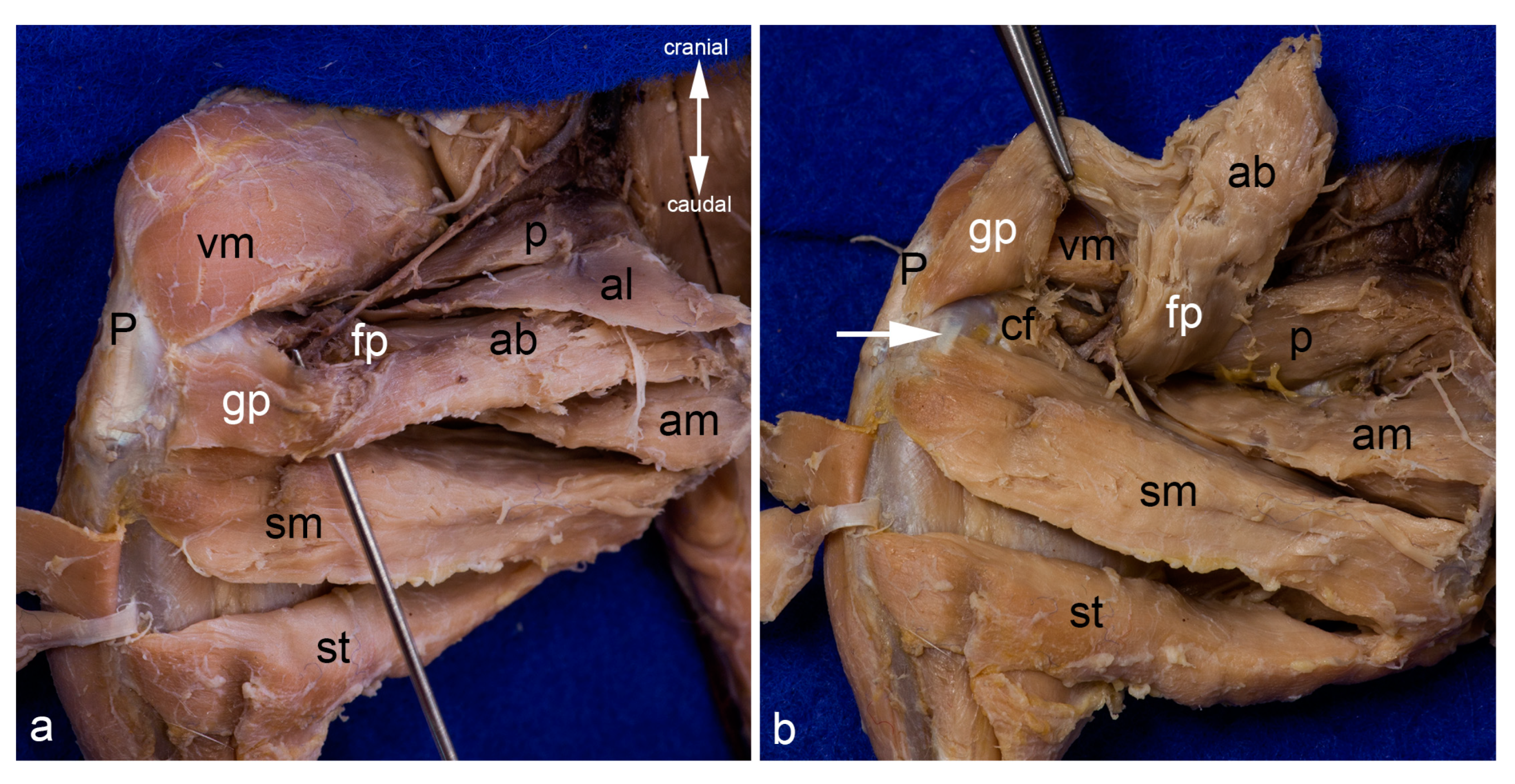
Figure 10.
Adductor canal. The proximal end of the adductor canal between the femoral part and the genicular part of the adductor brevis muscle is indicated with the tip of the probe (a). The distal end of this canal between the femoral part of the adductor brevis muscle and the caudofemoralis muscle can be observed in the deep. The femoral and genicular parts of the adductor brevis muscle are visualized by detaching the muscle from its origin and reflecting it cranially. The white arrow indicates the medial collateral ligament (b). The cranial and caudal gracilis muscles have been detached from their origin and reflected towards their insertion in both pictures. In both pictures, the thigh was abducted in the hip joint to show the medial aspect of the thigh. ab = adductor brevis muscle (fp = femoral part, gp = genicular part), al = adductor longus muscle, am = adductor magnus muscle, cf = caudofemoralis muscle, P = Patella/patellar ligament, p = pectineus muscle, sm = semimembranosus muscle, st = semitendinosus muscle, vm = vastus medialis muscle.
Figure 10.
Adductor canal. The proximal end of the adductor canal between the femoral part and the genicular part of the adductor brevis muscle is indicated with the tip of the probe (a). The distal end of this canal between the femoral part of the adductor brevis muscle and the caudofemoralis muscle can be observed in the deep. The femoral and genicular parts of the adductor brevis muscle are visualized by detaching the muscle from its origin and reflecting it cranially. The white arrow indicates the medial collateral ligament (b). The cranial and caudal gracilis muscles have been detached from their origin and reflected towards their insertion in both pictures. In both pictures, the thigh was abducted in the hip joint to show the medial aspect of the thigh. ab = adductor brevis muscle (fp = femoral part, gp = genicular part), al = adductor longus muscle, am = adductor magnus muscle, cf = caudofemoralis muscle, P = Patella/patellar ligament, p = pectineus muscle, sm = semimembranosus muscle, st = semitendinosus muscle, vm = vastus medialis muscle.
![Life 13 02096 g010]()
Figure 11.
Deepest layers in the medial aspect of the thigh. The adductor magnus and adductor minimus muscles are shown from the medial aspect of the thigh. They were already separated by blunt dissection starting at the entrance of the twigs of the caudal branch of the obturator nerve (white pin). The cranial branch of this nerve was reflected cranially. The red pin was inserted between the origins of these two muscles, the blue pin near their insertions (a). The adductor magnus and adductor minimus muscles were detached from their origin and mobilized towards their insertion. Thus, the deep aspect of the pectineus, the quadratus femoris, part of the iliacus and psoas major muscles, as well as the obturator externus and obturator intermedius muscles can be observed from the ventral aspect (b). In both pictures, almost all superficial muscles were detached from their origin and reflected towards their insertion or totally removed. The thigh was abducted in the hip joint. ab = adductor brevis muscle, al = adductor longus muscle, am = adductor magnus muscle, ami = adductor minimus muscle, cf = caudofemoralis muscle, ip = iliacus and psoas major muscles, LT = lesser trochanter, oe = obturator externus muscle, oim = obturator intermedius muscle, p = pectineus muscle, S = pelvic symphysis, vm = vastus medialis muscle, qf = quadratus femoris muscle.
Figure 11.
Deepest layers in the medial aspect of the thigh. The adductor magnus and adductor minimus muscles are shown from the medial aspect of the thigh. They were already separated by blunt dissection starting at the entrance of the twigs of the caudal branch of the obturator nerve (white pin). The cranial branch of this nerve was reflected cranially. The red pin was inserted between the origins of these two muscles, the blue pin near their insertions (a). The adductor magnus and adductor minimus muscles were detached from their origin and mobilized towards their insertion. Thus, the deep aspect of the pectineus, the quadratus femoris, part of the iliacus and psoas major muscles, as well as the obturator externus and obturator intermedius muscles can be observed from the ventral aspect (b). In both pictures, almost all superficial muscles were detached from their origin and reflected towards their insertion or totally removed. The thigh was abducted in the hip joint. ab = adductor brevis muscle, al = adductor longus muscle, am = adductor magnus muscle, ami = adductor minimus muscle, cf = caudofemoralis muscle, ip = iliacus and psoas major muscles, LT = lesser trochanter, oe = obturator externus muscle, oim = obturator intermedius muscle, p = pectineus muscle, S = pelvic symphysis, vm = vastus medialis muscle, qf = quadratus femoris muscle.
![Life 13 02096 g011]()
Table 1.
Systematic anatomy of the adductor muscles in the albino rat.
Table 1.
Systematic anatomy of the adductor muscles in the albino rat.
| Muscle | Origin | Insertion | Innervation |
|---|
| Cranial gracilis muscle | cranial and caudal pubic rami | cranial border of the tibia between tibial tuberosity and insertion of the semitendinosus muscle | cranial branch of the obturator nerve |
| Caudal gracilis muscle | ramus of ischium | medial surface of the tibia deep to the insertion of the cranial gracilis muscle | cranial branch of the obturator nerve |
| Adductor longus muscle | cranial and caudal pubic rami, ramus of ischium | medial lip of the facies aspera of the femur (fused with distal portion of insertion of pectineus muscle) | cranial branch of the obturator nerve |
| Adductor brevis muscle | cranial and caudal pubic rami | medial and lateral lip of facies aspera,
medial and lateral supracondylar tuberosities | cranial branch of the obturator nerve |
medial femoral epicondyle, medial patellar retinaculum,
medial tibial condyle |
| Adductor magnus muscle | ramus of ischium | distal part of caudal aspect of the third trochanter,
lateral lip of the facies aspera between adductor minimus (proximal) and adductor brevis (distal) | caudal branch of the obturator nerve |
| Adductor minimus muscle | cranial and caudal pubic rami (95% of cases), ramus of ischium (75% of cases) | caudal aspect of the third trochanter | caudal branch of the obturator nerve |
| Obturator externus muscle | cranial and caudal pubic rami,
ramus of ischium,
cranial margin of tabula of ischium,
outer aspect of the obturator membrane | trochanteric fossa,
fibrous capsule of the hip joint | obturator nerve (trunk) |
| Obturator intermedius muscle | cranial pubic ramus | obturator nerve (trunk) |
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}