

Natural Perylenequinone Compounds as Potent Inhibitors of Schistosoma mansoni Glutathione S-Transferase

Abstract
:1. Introduction
2. Methods
2.1. Exploring and Extracting Antihelminth Compounds and SmGST Protein Sequences from Databases
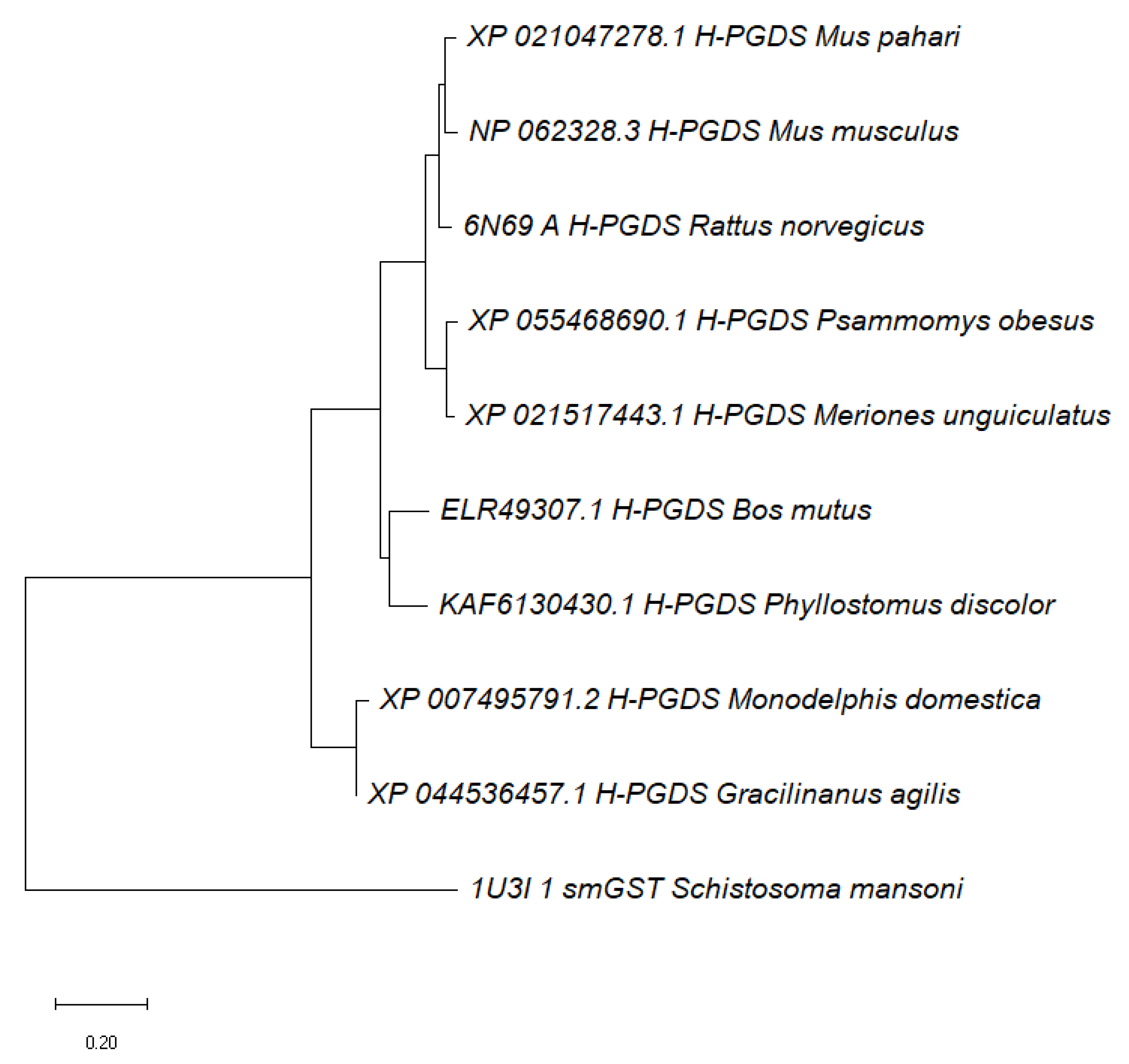
2.2. Alignment of Schistosoma mansoni GST Protein Sequences to Mammalian Homologs
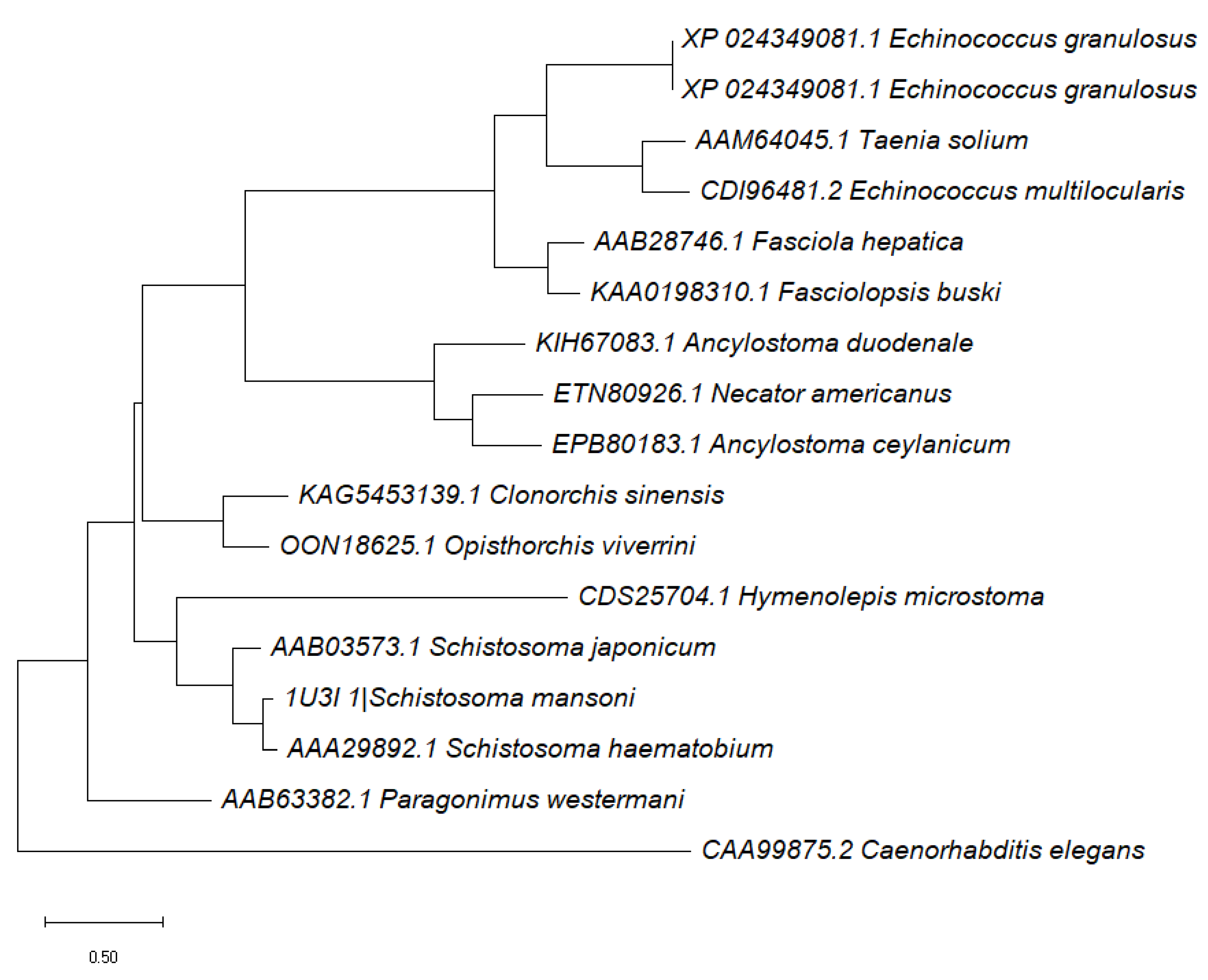
2.3. Schistosoma mansoni GST Relationship to Other Parasitic Worms as Well as Caenorhabditis elegans
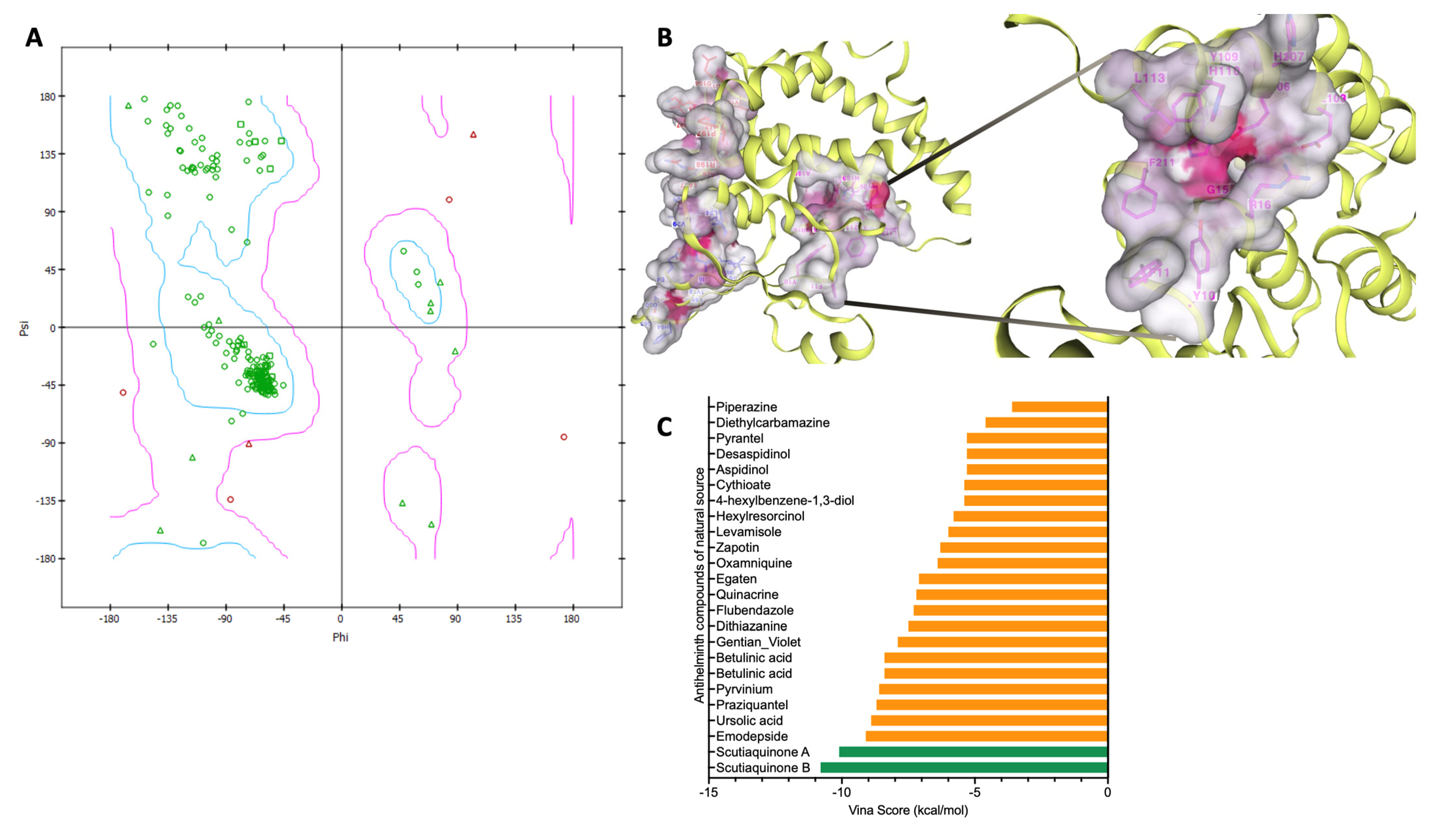
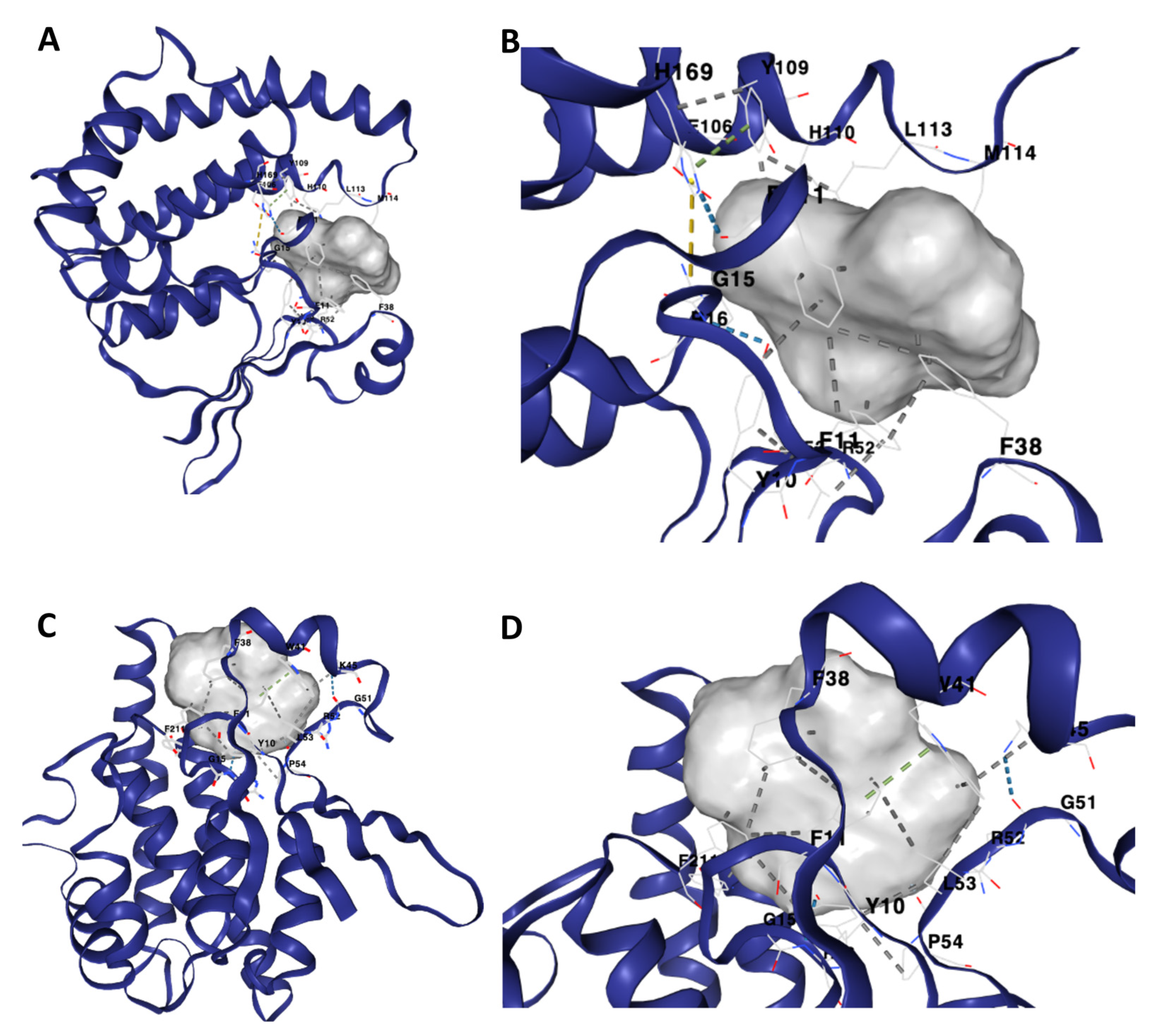
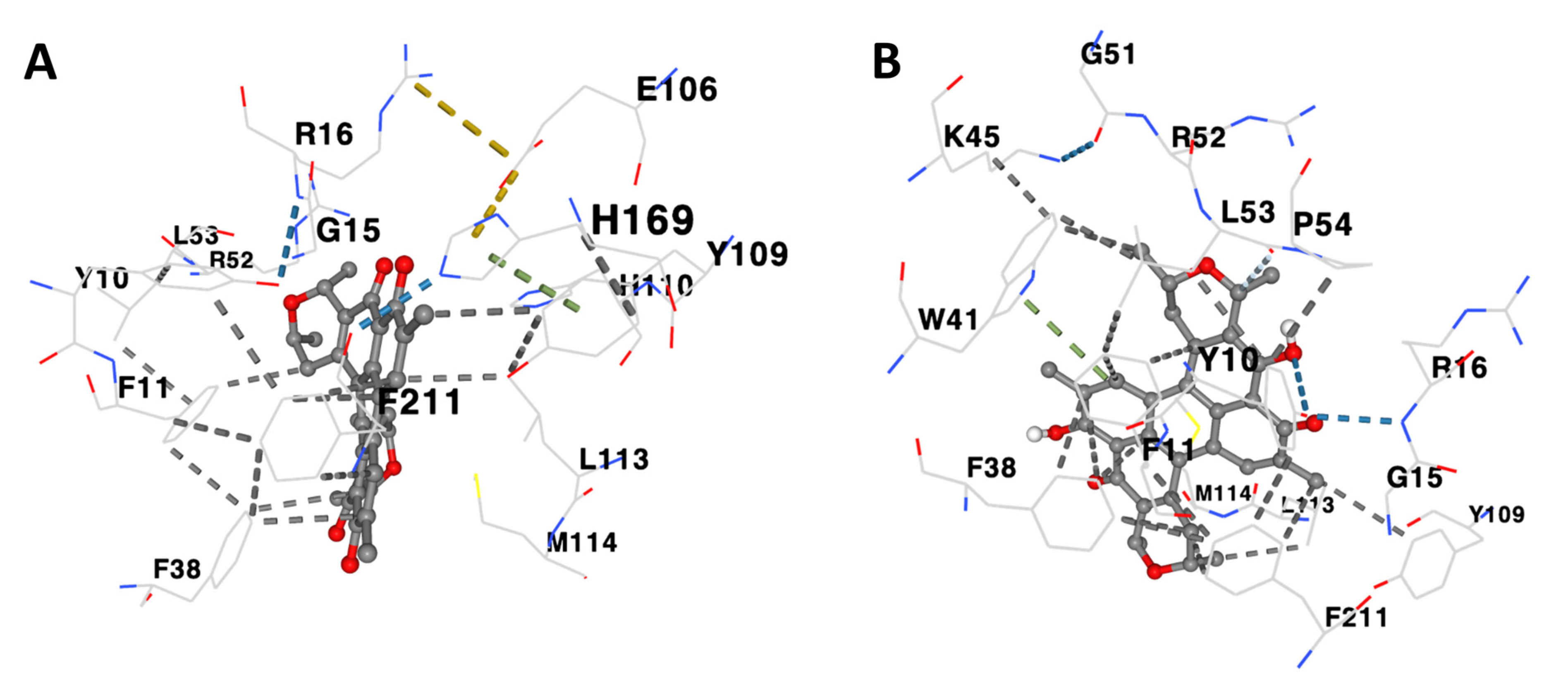
2.4. Protein–Ligand Docking of Compounds against Schistosoma mansoni GST
2.5. Analyses of the Binding Affinity of the SmGST Protein to the Compounds
2.6. Evaluation of Pharmacokinetic and Solubility Profiles for Scutiaquinone A and B
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Colley, D.G.; Bustinduy, A.L.; Secor, W.E.; King, C.H. Human schistosomiasis. Lancet 2014, 383, 2253–2264. [Google Scholar] [CrossRef] [PubMed]
- Lo, N.C.; Addiss, D.G.; Hotez, P.J.; King, C.H.; Stothard, J.R.; Evans, D.S.; Colley, D.G.; Lin, W.; Coulibaly, J.T.; Bustinduy, A.L. A call to strengthen the global strategy against schistosomiasis and soil-transmitted helminthiasis: The time is now. Lancet Infect. Dis. 2017, 17, e64–e69. [Google Scholar] [CrossRef] [PubMed]
- Lo, N.C.; Bezerra, F.S.M.; Colley, D.G.; Fleming, F.M.; Homeida, M.; Kabatereine, N.; Kabole, F.M.; King, C.H.; Mafe, M.A.; Midzi, N. Review of 2022 WHO guidelines on the control and elimination of schistosomiasis. Lancet Infect. Dis. 2022, 22, e327–e335. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, R.E.; Fleming, F.M.; de Vlas, S.J.; Odiere, M.R.; Kinung’hi, S.; King, C.H.; Evans, D.; French, M.D.; Montgomery, S.P.; Straily, A. Defining elimination as a public health problem for schistosomiasis control programmes: Beyond prevalence of heavy-intensity infections. Lancet Glob. Health 2022, 10, e1355–e1359. [Google Scholar] [CrossRef]
- Colley, D.G.; Andros, T.S.; Campbell, C.H. Schistosomiasis is more prevalent than previously thought: What does it mean for public health goals, policies, strategies, guidelines and intervention programs? Infect. Dis. Poverty 2017, 6, 63. [Google Scholar] [CrossRef] [PubMed]
- Cioli, D.; Pica-Mattoccia, L.; Basso, A.; Guidi, A. Schistosomiasis control: Praziquantel forever? Mol. Biochem. Parasitol. 2014, 195, 23–29. [Google Scholar] [CrossRef]
- Fenwick, A.; Webster, J.P.; Bosque-Oliva, E.; Blair, L.; Fleming, F.; Zhang, Y.; Garba, A.; Stothard, J.; Gabrielli, A.F.; Clements, A. The Schistosomiasis Control Initiative (SCI): Rationale, development and implementation from 2002–2008. Parasitology 2009, 136, 1719–1730. [Google Scholar] [CrossRef]
- Stothard, J.R.; Sousa-Figueiredo, J.C.; Betson, M.; Green, H.K.; Seto, E.Y.; Garba, A.; Sacko, M.; Mutapi, F.; Nery, S.V.; Amin, M.A. Closing the praziquantel treatment gap: New steps in epidemiological monitoring and control of schistosomiasis in African infants and preschool-aged children. Parasitology 2011, 138, 1593–1606. [Google Scholar] [CrossRef]
- Caffrey, C.R. Chemotherapy of schistosomiasis: Present and future. Curr. Opin. Chem. Biol. 2007, 11, 433–439. [Google Scholar] [CrossRef]
- Summers, S.; Bhattacharyya, T.; Allan, F.; Stothard, J.R.; Edielu, A.; Webster, B.L.; Miles, M.A.; Bustinduy, A.L. A review of the genetic determinants of praziquantel resistance in Schistosoma mansoni: Is praziquantel and intestinal schistosomiasis a perfect match? Front. Trop. Dis. 2022, 3, 933097. [Google Scholar] [CrossRef]
- Botros, S.; Sayed, H.; Amer, N.; El-Ghannam, M.; Bennett, J.L.; Day, T.A. Current status of sensitivity to praziquantel in a focus of potential drug resistance in Egypt. Int. J. Parasitol. 2005, 35, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-M.; Coultas, K.A. Identification of plumbagin and sanguinarine as effective chemotherapeutic agents for treatment of schistosomiasis. Int. J. Parasitol. Drugs Drug Resist. 2013, 3, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Engels, D.; Chitsulo, L.; Montresor, A.; Savioli, L. The global epidemiological situation of schistosomiasis and new approaches to control and research. Acta Trop. 2002, 82, 139–146. [Google Scholar] [CrossRef]
- Faust, C.L.; Osakunor, D.N.; Downs, J.A.; Kayuni, S.; Stothard, J.R.; Lamberton, P.H.; Reinhard-Rupp, J.; Rollinson, D. Schistosomiasis control: Leave no age group behind. Trends Parasitol. 2020, 36, 582–591. [Google Scholar] [CrossRef]
- Nogueira, R.A.; Lira, M.G.S.; Licá, I.C.L.; Frazão, G.C.C.G.; Dos Santos, V.A.F.; Mendes Filho, A.C.C.; Rodrigues, J.G.M.; Miranda, G.S.; Carvalho, R.C.; Nascimento, F.R.F. Praziquantel: An update on the mechanism of its action against schistosomiasis and new therapeutic perspectives. Mol. Biochem. Parasitol. 2022, 252, 111531. [Google Scholar] [CrossRef] [PubMed]
- Kremer, L.; Riveau, G.; Baulard, A.; Capron, A.; Locht, C. Neutralizing antibody responses elicited in mice immunized with recombinant bacillus Calmette-Guérin producing the Schistosoma mansoni glutathione S-transferase. J. Immunol. 1996, 156, 4309–4317. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Q.; Liu, S.S.; Cheng, C.; Cui, J.; Wang, Z.Q.; Zhang, X. Molecular characteristics of glutathione transferase gene family in a neglect medical Spirometra tapeworm. Front. Vet. Sci. 2022, 9, 1035767. [Google Scholar] [CrossRef]
- Adegboye, O.; Field, M.A.; Kupz, A.; Pai, S.; Sharma, D.; Smout, M.J.; Wangchuk, P.; Wong, Y.; Loiseau, C. Natural-product-based solutions for tropical infectious diseases. Clin. Microbiol. Rev. 2021, 34, e00348-20. [Google Scholar] [CrossRef]
- Wangchuk, P.; Loukas, A. Techniques and technologies for the biodiscovery of novel small molecule drug lead compounds from natural products. In Natural Products and Drug Discovery; Elsevier: Amsterdam, The Netherlands, 2018; pp. 435–465. [Google Scholar]
- Neves, B.J.; Andrade, C.H.; Cravo, P.V. Natural products as leads in schistosome drug discovery. Molecules 2015, 20, 1872–1903. [Google Scholar] [CrossRef]
- Mtemeli, F.; Ndlovu, J.; Mugumbate, G.; Makwikwi, T.; Shoko, R. Advances in schistosomiasis drug discovery based on natural products. All Life 2022, 15, 608–623. [Google Scholar] [CrossRef]
- Wangchuk, P. Therapeutic applications of natural products in herbal medicines, biodiscovery programs, and biomedicine. J. Biol. Act. Prod. Nat. 2018, 8, 1–20. [Google Scholar] [CrossRef]
- Cheuka, P.M.; Mayoka, G.; Mutai, P.; Chibale, K. The role of natural products in drug discovery and development against neglected tropical diseases. Molecules 2016, 22, 58. [Google Scholar] [CrossRef] [PubMed]
- Racz, L.Z.; Racz, C.P.; Pop, L.-C.; Tomoaia, G.; Mocanu, A.; Barbu, I.; Sárközi, M.; Roman, I.; Avram, A.; Tomoaia-Cotisel, M. Strategies for improving bioavailability, bioactivity, and physical-chemical behavior of curcumin. Molecules 2022, 27, 6854. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, A.; Javed, S.; Wahab, S.; Azmi, L.; Tabish, M.; Sultan, M.H.; Abdelsalam, K.; Alqahtani, S.S.; Ahmad, M.F. Cellular, Molecular, Pharmacological, and Nano-Formulation Aspects of Thymoquinone—A Potent Natural Antiviral Agent. Molecules 2023, 28, 5435. [Google Scholar] [CrossRef]
- Adekiya, T.A.; Aruleba, R.T.; Klein, A.; Fadaka, A.O. In silico inhibition of SGTP4 as a therapeutic target for the treatment of schistosomiasis. J. Biomol. Struct. Dyn. 2022, 40, 3697–3705. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.G.; Oliva, G.; Andricopulo, A.D. Target-based molecular modeling strategies for schistosomiasis drug discovery. Future Med. Chem. 2015, 7, 753–764. [Google Scholar] [CrossRef]
- Huang, H.-H.; Rigouin, C.; L Williams, D. The redox biology of schistosome parasites and applications for drug development. Curr. Pharm. Des. 2012, 18, 3595–3611. [Google Scholar] [CrossRef]
- Anibogwu, R.; Jesus, K.D.; Pradhan, S.; Pashikanti, S.; Mateen, S.; Sharma, K. Extraction, isolation and characterization of bioactive compounds from Artemisia and their biological significance: A review. Molecules 2021, 26, 6995. [Google Scholar] [CrossRef]
- Diallo, B.; Glenister, M.; Musyoka, T.M.; Lobb, K.; Tastan Bishop, Ö. SANCDB: An update on South African natural compounds and their readily available analogs. J. Cheminform. 2021, 13, 37. [Google Scholar] [CrossRef]
- Hatherley, R.; Brown, D.K.; Musyoka, T.M.; Penkler, D.L.; Faya, N.; Lobb, K.A.; Tastan Bishop, Ö. SANCDB: A South African natural compound database. J. Cheminform. 2015, 7, 29. [Google Scholar] [CrossRef]
- Sayers, E.W.; Beck, J.; Bolton, E.E.; Bourexis, D.; Brister, J.R.; Canese, K.; Comeau, D.C.; Funk, K.; Kim, S.; Klimke, W. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2021, 49, D10. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022, 50, D20. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tamura, K.; Nei, M. MEGA: Molecular evolutionary genetics analysis software for microcomputers. Bioinformatics 1994, 10, 189–191. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, X.; Gan, J.; Chen, S.; Xiao, Z.-X.; Cao, Y. CB-Dock2: Improved protein–ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Res. 2022, 50, W159–W164. [Google Scholar] [CrossRef]
- Gan, J.-H.; Liu, J.-X.; Liu, Y.; Chen, S.-W.; Dai, W.-T.; Xiao, Z.-X.; Cao, Y. DrugRep: An automatic virtual screening server for drug repurposing. Acta Pharmacol. Sin. 2023, 44, 888–896. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Riveau, G.; Poulain-Godefroy, O.; Dupré, L.; Remoué, F.; Mielcarek, N.; Locht, C.; Capron, A. Glutathione S-transferases of 28kDa as major vaccine candidates against schistosomiasis. Mem. Inst. Oswaldo Cruz 1998, 93, 87–94. [Google Scholar] [CrossRef]
- Angelucci, F.; Baiocco, P.; Brunori, M.; Gourlay, L.; Morea, V.; Bellelli, A. Insights into the catalytic mechanism of glutathione S-transferase: The lesson from Schistosoma haematobium. Structure 2005, 13, 1241–1246. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D.; Meade, G.; Foley, V.M.; Dowd, C.A. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem. J. 2001, 360, 1–16. [Google Scholar] [CrossRef] [PubMed]
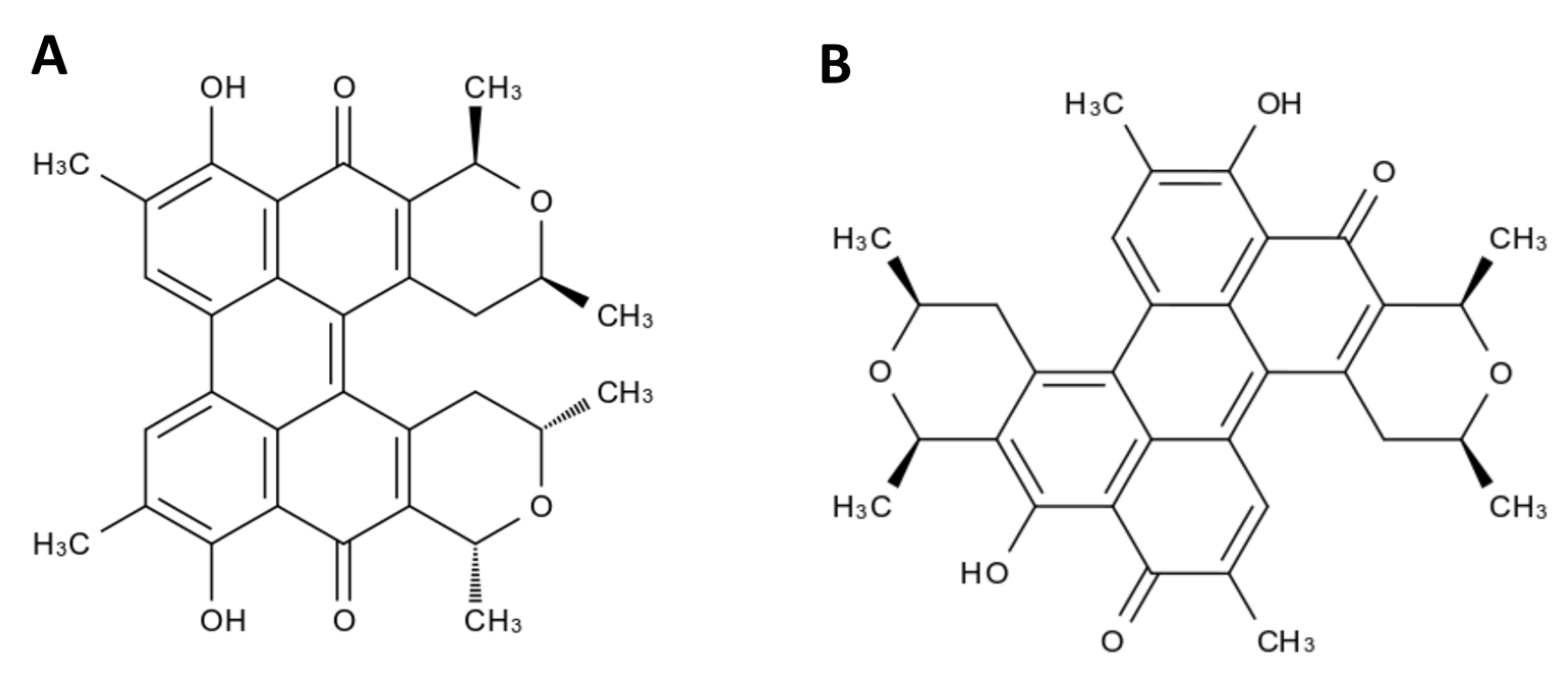
- Ayers, S.; Zink, D.L.; Mohn, K.; Powell, J.S.; Brown, C.M.; Murphy, T.; Brand, R.; Pretorius, S.; Stevenson, D.; Thompson, D. Scutiaquinones A and B, Perylenequinones from the Roots of Scutia myrtina with Anthelmintic Activity. J. Nat. Prod. 2007, 70, 425–427. [Google Scholar] [CrossRef]
- Mulrooney, C.A.; O’Brien, E.M.; Morgan, B.J.; Kozlowski, M.C. Perylenequinones: Isolation, synthesis, and biological activity. Eur. J. Org. Chem. 2012, 2012, 3887–3904. [Google Scholar] [CrossRef] [PubMed]
- Mulrooney, C.A.; Morgan, B.J.; Li, X.; Kozlowski, M.C. Perylenequinone natural products: Enantioselective synthesis of the oxidized pentacyclic core. J. Org. Chem. 2010, 75, 16–29. [Google Scholar] [CrossRef]
- Emery, D.L.; Hunt, P.W.; Le Jambre, L.F. Haemonchus contortus: The then and now, and where to from here? Int. J. Parasitol. 2016, 46, 755–769. [Google Scholar] [CrossRef]
- Bickerton, G.R.; Paolini, G.V.; Besnard, J.; Muresan, S.; Hopkins, A.L. Quantifying the chemical beauty of drugs. Nat. Chem. 2012, 4, 90–98. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A boiled-egg to predict gastrointestinal absorption and brain penetration of small molecules. ChemMedChem 2016, 11, 1117–1121. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. iLOGP: A simple, robust, and efficient description of n-octanol/water partition coefficient for drug design using the GB/SA approach. J. Chem. Inf. Model. 2014, 54, 3284–3301. [Google Scholar] [CrossRef]
- Lautens, M.J.; Tan, J.H.; Serrat, X.; Del Borrello, S.; Schertzberg, M.R.; Fraser, A.G. Identification of enzymes that have helminth-specific active sites and are required for Rhodoquinone-dependent metabolism as targets for new anthelmintics. PLoS Neglected Trop. Dis. 2021, 15, e0009991. [Google Scholar] [CrossRef]
- Rana, A.K.; Misra-Bhattacharya, S. Current drug targets for helminthic diseases. Parasitol. Res. 2013, 112, 1819–1831. [Google Scholar] [CrossRef] [PubMed]
- J Timson, D. Metabolic enzymes of helminth parasites: Potential as drug targets. Curr. Protein Pept. Sci. 2016, 17, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Alzain, A.A.; Mukhtar, R.M.; Abdelmoniem, N.; Elbadwi, F.A.; Hussien, A.; Garelnabi, E.A.; Osman, W.; Sherif, A.E.; Khedr, A.I.; Ghazawi, K.F. Computational Insights into Natural Antischistosomal Metabolites as SmHDAC8 Inhibitors: Molecular Docking, ADMET Profiling, and Molecular Dynamics Simulation. Metabolites 2023, 13, 658. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Pandey, P.N. Molecular docking and molecular dynamics study on SmHDAC1 to identify potential lead compounds against Schistosomiasis. Mol. Biol. Rep. 2015, 42, 689–698. [Google Scholar] [CrossRef]
- Akachukwu, I.; O Olubiyi, O.; Kosisochukwu, A.; John, M.C.; Justina, N.N. Structure-Based study of Natural products with anti-Schistosoma activity. Curr. Comput.-Aided Drug Des. 2017, 13, 91–100. [Google Scholar] [CrossRef]
- Mafud, A.C.; Ferreira, L.G.; Mascarenhas, Y.P.; Andricopulo, A.D.; de Moraes, J. Discovery of novel antischistosomal agents by molecular modeling approaches. Trends Parasitol. 2016, 32, 874–886. [Google Scholar] [CrossRef]
- Menezes, R.P.B.d.; Viana, J.d.O.; Muratov, E.; Scotti, L.; Scotti, M.T. Computer-assisted discovery of alkaloids with schistosomicidal activity. Curr. Issues Mol. Biol. 2022, 44, 383–408. [Google Scholar] [CrossRef]
- Rocha, J.A.; Rego, N.C.; Carvalho, B.T.; Silva, F.I.; Sousa, J.A.; Ramos, R.M.; Passos, I.N.; de Moraes, J.; Leite, J.R.; Lima, F.C. Computational quantum chemistry, molecular docking, and ADMET predictions of imidazole alkaloids of Pilocarpus microphyllus with schistosomicidal properties. PLoS ONE 2018, 13, e0198476. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
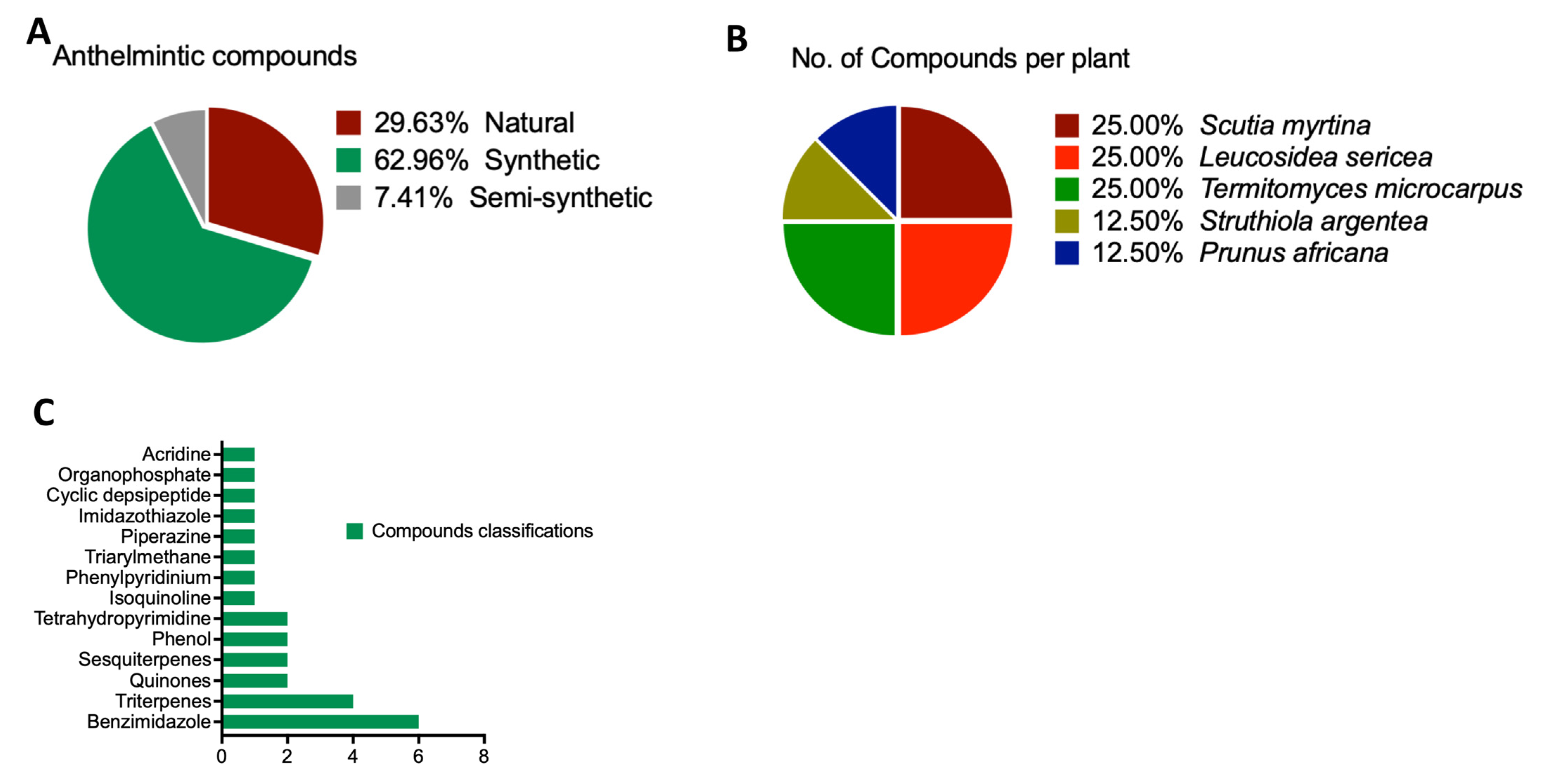
| Database | Activities | S/N | Compound Name | Accession No | Sources | Class | Vina Score | Cavity Volume (Å3) | Center (x, y, z) | Docking Size (x, y, z) |
|---|---|---|---|---|---|---|---|---|---|---|
| SANCDB | Anthelmintic Compound | 1 | Scutiaquinone A | SANC00584 | Scutia myrtina | Quinones | −10.1 | 297 | 16, 51, 34 | 22, 22, 22 |
| 2 | Scutiaquinone B | SANC00585 | Scutia myrtina | Quinones | −10.8 | 297 | 16, 51, 34 | 23, 23, 23 | ||
| 3 | Aspidinol | SANC00741 | Leucosidea sericea | Sesquiterpenes | −5.3 | 117 | 14, 56, 13 | 19, 19, 19 | ||
| 4 | Desaspidinol | SANC00742 | Leucosidea sericea | Sesquiterpenes | −5.3 | 117 | 14, 56, 13 | 19, 19, 19 | ||
| 5 | Betulinic acid | SANC00743 | Termitomyces microcarpus | Triterpenes | −8.4 | 117 | 14, 56, 13 | 22, 22, 22 | ||
| 6 | Zapotin | SANC01031 | Struthiola argentea | Triterpenes | −6.3 | 117 | 14, 56, 13 | 21, 21, 21 | ||
| 7 | Betulinic acid | SANC00743 | Termitomyces microcarpus | Triterpenes | −8.4 | 117 | 14, 56, 13 | 22, 22, 22 | ||
| 8 | Ursolic acid | SANC00744 | Prunus africana | Triterpenes | −8.9 | 117 | 14, 56, 13 | 23, 23, 23 | ||
| pubchem | 9 | 4-hexylbenzene-1,3-diol | Compound CID: 3610 | Anacardium occidentale, Sargassum muticum | Phenol | −5.4 | 117 | 14, 56, 13 | 20, 20, 20 | |
| drugbank | 10 | Albendazole | DB00518 | Synthetic Source | Benzimidazole | |||||
| 11 | Pyrantel | DB11156 | Synthetic Source | Tetrahydropyrimidine | −5.3 | 117 | 14, 56, 13 | 20, 20, 20 | ||
| 12 | Piperazine | DB00592 | Synthetic Source | Piperazine | −3.6 | 297 | 16, 51, 34 | 14, 14, 14 | ||
| 13 | Mebendazole | DB00643 | Synthetic Source | Benzimidazole | ||||||
| 14 | Praziquantel | DB01058 | Synthetic Source | Isoquinoline | −8.7 | 297 | 16, 51, 34 | 22, 22, 22 | ||
| 15 | Oxamniquine | DB01096 | Semi-synthetic | Tetrahydroquinoline | −6.4 | 297 | 16, 51, 34 | 21, 21, 21 | ||
| 16 | Egaten | DB12245 | Synthetic Source | Benzimidazole | −7.1 | 297 | 16, 51, 34 | 22, 22, 22 | ||
| 17 | Pyrvinium | DB06816 | Synthetic Source | Phenylpyridinium | −8.6 | 297 | 16, 51, 34 | 24, 24, 24 | ||
| 18 | Gentian_Violet | DB00406 | Synthetic Source | Triarylmethane | −7.9 | 297 | 16, 51, 34 | 23, 23, 23 | ||
| 19 | Diethylcarbamazine | DB00711 | Synthetic Source | Piperazine | −4.6 | 117 | 14, 56, 13 | 18, 18, 18 | ||
| 20 | Levamisole | DB00848 | Synthetic Source | Imidazothiazole | −6 | 117 | 14, 56, 13 | 19, 19, 19 | ||
| 21 | Hexylresorcinol | DB11254 | Synthetic Source | Phenol | −5.8 | 297 | 16, 51, 34 | 20, 20, 20 | ||
| 22 | Emodepside | DB11403 | Semi-synthetic | Cyclic depsipeptide | −9.1 | 117 | 14, 56, 13 | 29, 29, 29 | ||
| 23 | Flubendazole | DB08974 | Synthetic Source | Benzimidazole | −7.3 | 297 | 16, 51, 34 | 23, 23, 23 | ||
| 24 | Cythioate | DB11392 | Synthetic Source | Organophosphate | −5.4 | 297 | 16, 51, 34 | 20, 20, 20 | ||
| 25 | Quinacrine | DB01103 | Synthetic Source | Acridine | −7.2 | 297 | 16, 51, 34 | 22, 22, 22 | ||
| 26 | Fenbendazole | DB11410 | Synthetic Source | Benzimidazole | ||||||
| 27 | Dithiazanine | DB11516 | Synthetic Source | Benzothiazole | −7.5 | 297 | 16, 51, 34 | 25, 25, 25 |
| Scutiaquinone A | Scutiaquinone B | ||
|---|---|---|---|
| Lipophilicity | Log Po/w (iLOGP) | 4.38 | 4.39 |
| Log Po/w (XLOGP3) | 5.72 | 6.17 | |
| Log Po/w (WLOGP) | 6.32 | 6.05 | |
| Log Po/w (MLOGP) | 2.62 | 2.62 | |
| Log Po/w (SILICOS-IT) | 6.68 | 6.71 | |
| Consensus Log Po/w | 5.15 | 5.19 | |
| Water Solubility | Log S (ESOL) | −6.88 | −7.17 |
| Solubility | 6.7 × 10−5 mg/mL; 1.31 × 10−7 mol/L | 3.49 × 10−5 mg/mL; 6.83 × 10−8 mol/L | |
| Class | Poorly soluble | Poorly soluble | |
| Log S (Ali) | −7.44 | −7.91 | |
| Solubility | 1.85 × 10−5 mg/mL; 3.62 × 10−8 mol/L | 6.31 × 10−6 mg/mL; 1.24 × 10−8 mol/L | |
| Class | Poorly soluble | Poorly soluble | |
| Log S (SILICOS-IT) | −8.33 | −8.11 | |
| Solubility | 2.39 × 10−6 mg/mL; 4.69 × 10−9 mol/L | 3.96 × 10−6 mg/mL; 7.76 × 10−9 mol/L | |
| Class | Poorly soluble | Poorly soluble | |
| Druglikeness | Lipinski | Yes | Yes |
| Veber | Yes | Yes | |
| Bioavailability Score | 0.55 | 0.55 | |
| Pharmacokinetics | GI absorption | Low | High |
| BBB permeant | No | No | |
| P-gp substrate | No | No | |
| CYP1A2 inhibitor | No | No | |
| CYP2C19 inhibitor | Yes | No | |
| CYP2C9 inhibitor | Yes | Yes | |
| CYP2D6 inhibitor | No | No | |
| CYP3A4 inhibitor | Yes | No | |
| Log Kp (skin permeation) | −5.35 cm/s | −5.03 cm/s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otarigho, B.; Falade, M.O. Natural Perylenequinone Compounds as Potent Inhibitors of Schistosoma mansoni Glutathione S-Transferase. Life 2023, 13, 1957. https://doi.org/10.3390/life13101957
Otarigho B, Falade MO. Natural Perylenequinone Compounds as Potent Inhibitors of Schistosoma mansoni Glutathione S-Transferase. Life. 2023; 13(10):1957. https://doi.org/10.3390/life13101957
Chicago/Turabian StyleOtarigho, Benson, and Mofolusho O. Falade. 2023. "Natural Perylenequinone Compounds as Potent Inhibitors of Schistosoma mansoni Glutathione S-Transferase" Life 13, no. 10: 1957. https://doi.org/10.3390/life13101957