
Naringin and Naringenin Polyphenols in Neurological Diseases: Understandings from a Therapeutic Viewpoint

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Methodology
3. Naringin and Naringenin
4. Botanical Sources
5. Neuroprotective Action
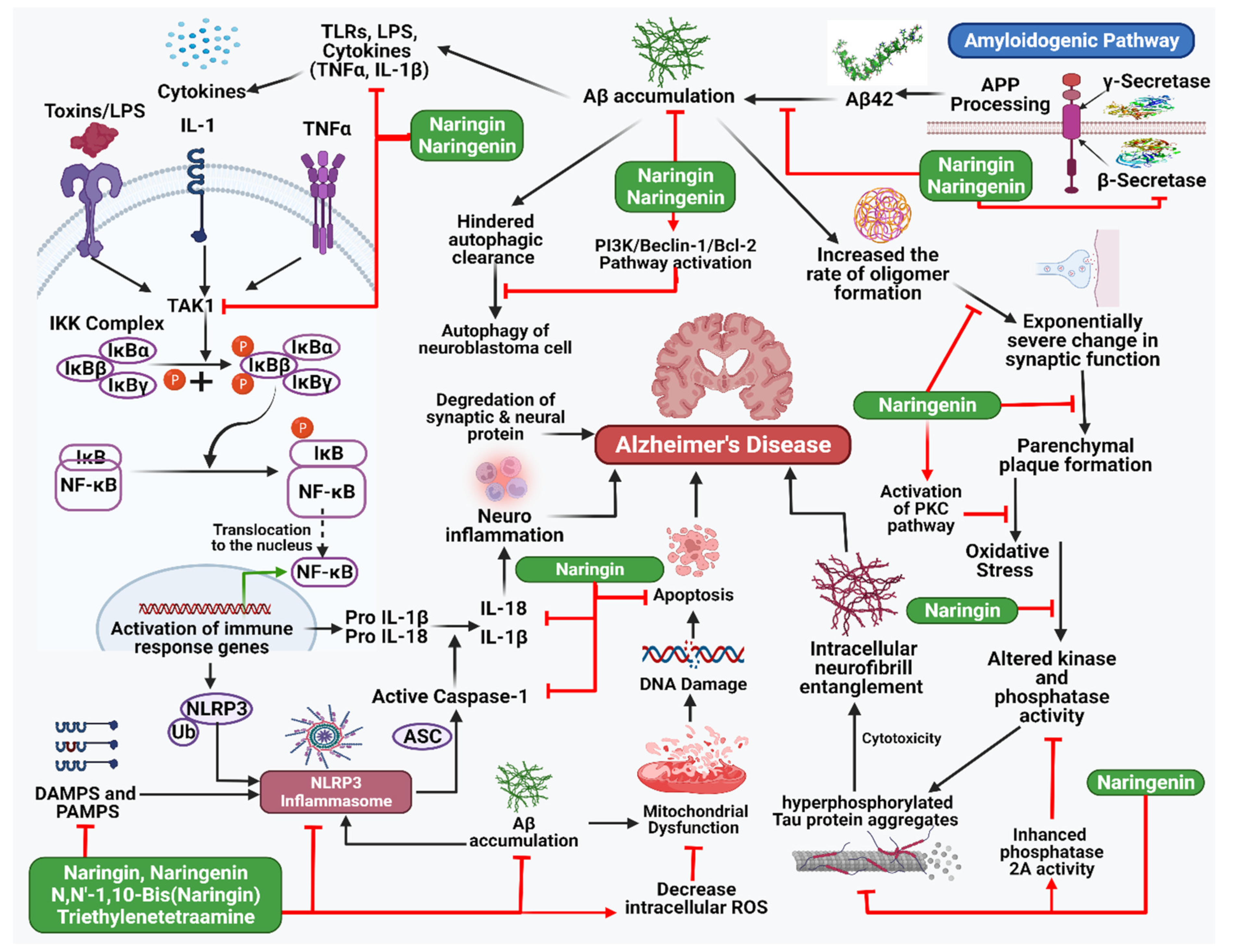
5.1. Alzheimer’s Disease
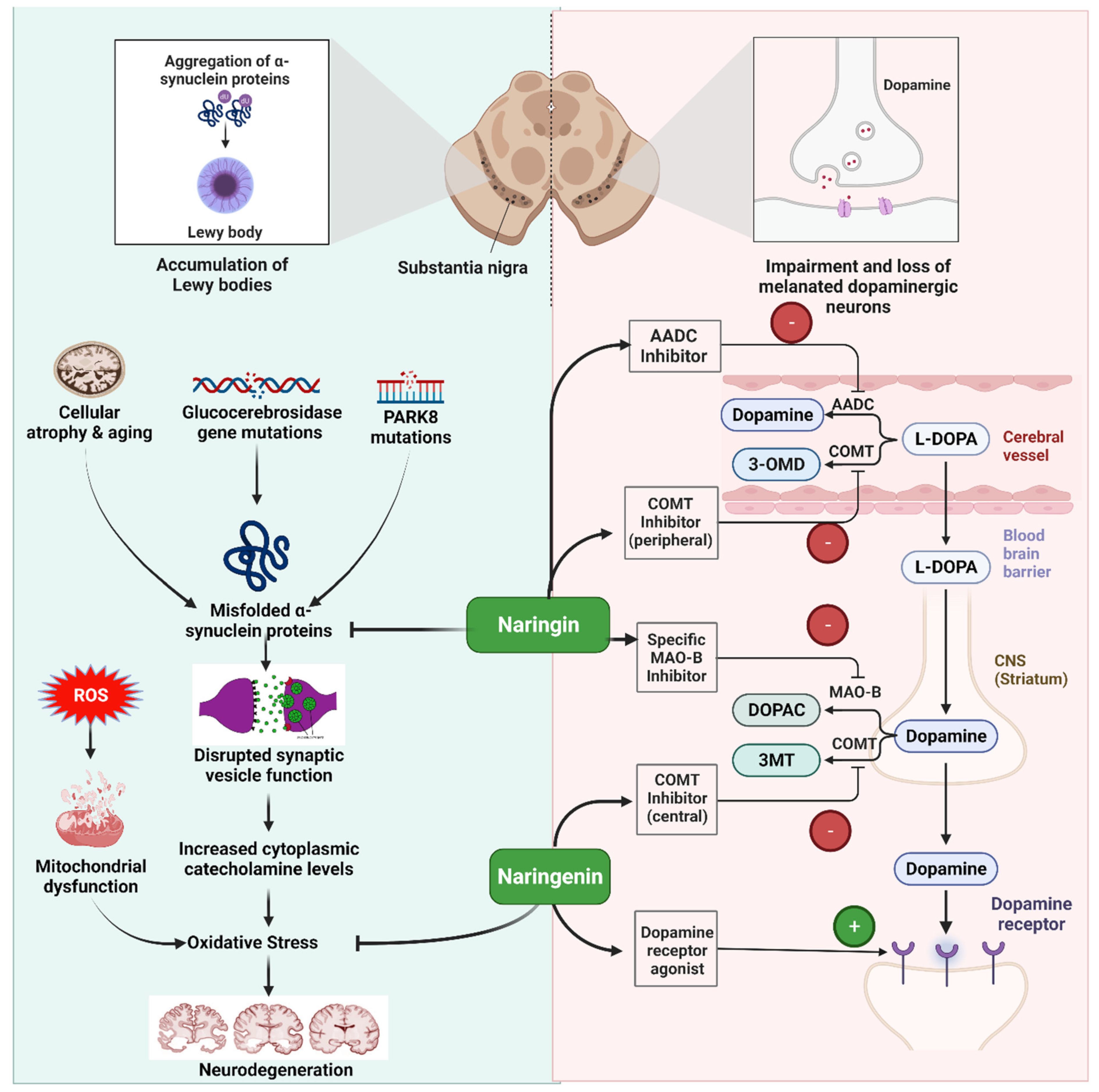
5.2. Parkinson’s Disease

{kind=link}
{kind=link}
{kind=link}
| Disease | Compound | Dose/Conc. | Study Model | Findings | References |
|---|---|---|---|---|---|
| Alzheimer Disease | Naringin | 50, 100 and 200 mg/kg; PO) for twentyone days | ICV-STZ rats | Restoration of cognitive deficits in ICV-STZ rat along with mitigation of mitochondrial dysfunction mediated oxido-nitrosative stress and cytokine release | [58] |
| Naringin | 50 or 100 mg/kg/day | APPswe/PSΔE9 transgenic mice | Reduction in plaque burden and an increase in glucose uptake through the inhibition of GSK-3β activity | [59] | |
| Naringin | 40 and 80 mg/kg | Wistar rats | Protection against ICV β-A1–42 and intranasal manganese induced memory dysfunction possibly due to its antioxidant, anti-inflammatory, anti-amyloidogenesis | [60] | |
| Naringin | 100 mg/kg/day | Mice | Neuroprotective effects through a variety of mechanisms, including amyloid β metabolism, Tau protein hyperphosphorylation, acetyl cholinergic system, glutamate receptor system, oxidative stress and cell apoptosis | [61] | |
| Naringenin | 70–210 µg/mL | PC12 cells | Inhibition of AChE activity | [62] | |
| Naringenin | 50 mg/kg | Male albino Wistar rats | Reduced oxidative stress markers: 4-HNE, MDA, TBARS, H2O2, PC, GSH in the hippocampus; Increase antioxidant level: GPx, GR, GST, SOD, CAT and Na+/K+-ATPase in the hippocampus | [63] | |
| Naringenin | 25, 50 and 100 mg/kg | Male Sprague-Dawley rats | Increased the mRNA expression of INS and INSR in cerebral cortex and hippocampus. In addition, NAR reversed ICV-STZ induced Tau hyper-phosphorylation in both hippocampus and cerebral cortex through downregulation of GSK-3β activity | [41] | |
| Naringenin | 25, 50 and 100 μM and 1.5, 3.0 and 4.5 mg/kg | PC12 cells and male ICR mice | Decreased ROS level and LDH activity | [64] | |
| Naringenin | 25 and 50 mg/kg | Male Sprague-Dawley rats | Decreased oxidative stress by depleting elevated lipid peroxide and nitric oxide and elevating reduced glutathione levels and exert cholinergic function through the inhibition of elevated ChE activity | [65] | |
| Naringin dihydrochalcone | 100 mg/kg | APPswe/PS1ΔE9 (APP/PS1) transgenic mice | Reduction in amyloid plaque burden and Aβ levels, suppression of neuroinflammation and promotion of neurogenesis | [66] | |
| Naringin | 40 and 80 mg/kg, PO | Male Wistar rats | Improvement in the cognitive performance and attenuated oxidative damage, as evidenced by lowering of malondialdehyde level and nitrite concentration and restoration of superoxide dismutase, catalase, glutathione S-transferase, and reduced glutathione levels, and acetylcholinesterase activity | [67] | |
| Naringin | 30 or 60 mg/kg/day | NMRI male mice | Reduction of Aβ plaque numbers in CA1, CA3, and DG areas of the hippocampus | [68] | |
| Naringin | 25, 50 and 100 mg/kg PO | Westar rats | Reduced lipid peroxidation, restored reduced superoxide dismutase and catalase) and acetylcholine esterase activity were significantly decreased | [69] | |
| N,N′-1,10-Bis(Naringin) Triethylenetetraamine | 10–200 μM | PC12 cells | Deceased the level of ROS in Cu2+-Aβ1-42-treated PC12 cells and elevate the SOD activity in Cu2+-Aβ1-42-treated PC12 cells | [70] | |
| Naringin | 2.5, 5 and 10 mg/kg | Swiss mice | Increased the activities of superoxide dismutase and catalase, and glutathione and decreased malondialdehyde and nitrite contents, and reduced brain acetylcholinesterase activity in mice brains | [71] | |
| Naringin | 80 mg/kg | Wistar albino rats | Improvement of the Aβ-induced cholinergic dysfunction and increase in the activity of AChE in rat hippocampus, prefrontal cortex, and amygdala. Furthermore, naringin attenuated Aβ-induced decrease in mitochondrial function, integrity, and bioenergetics as well as mitochondrial and cytosolic calcium level in all the brain regions. Moreover, reversal of Aβ-induced increase in apoptosis and level of mitochondrial calcium uniporter and decrease in the level of hemeoxygenase-1 | [72] | |
| Naringenin | 100 mg/kg, orally | male Wistar rats | Lowered hippocampal MDA content | [73] | |
| Parkinson Disease | Naringin | 80 mg/kg | rat model | Protection of the nigrostriatal DA projection by increasing glial cell line-derived neurotrophic factor expression and decreasing TNF-a expression in DA neurons and microglia | [74] |
| Naringin | 8 or 80 mg/kg per day | Female Sprague Dawley (SD) rats | Increased the level of GDNF in DA neurons, contributing to neuroprotection in the MPP+ rat model of PD, with activation of mammalian target of rapamycin complex 1 and pre-treatment with naringin could attenuate the level of TNF-α in the substantia nigra of MPP+-treated brains | [75] | |
| Naringin | 80 mg/kg | male Wistar albino rats | Neuroprotective activity against rotenone-induced toxicity in the animals possibly through Nrf2-mediated pathway | [76] | |
| Naringin | 50, 100 and 200 mg/kg | Swiss albino mice | Reduction in haloperidol-induced cataleptic scores in both bar test and block test | [77] | |
| Naringenin | 25, 50, 100 mg/kg/b.w, PO | male C57BL/6J mice | Reversed the toxic effects of MPTP by reducing LPO levels and increasing the activities of glutathione reductase and catalase along with improved behavioral performance | [78] | |
| Naringenin | 50 mg/kg, orally | albino Wister rats | Improved oxidative stress status by decreasing MDA and increasing glutathione content | [79] | |
| Naringenin | 50, 100 mg/kg | male Sprague-Dawley rats BV-2 and MN9D cell lines | Inhibition of microglia-induced neuroinflammation via NLRP3 inflammasome inactivation | [80] | |
| Naringenin | 25, 50, and 100 mM | SH-SY5Y Cell Line | Reduction of the ROS production by decreasing oxidative stress markers such as LPO and NO and increasing SOD level. In addition, pretreatment with NGN decreased the inflammatory markers such as TNF-α and NF-κβ in MPP+-treated SH-SY5Y cells. Further, NGN decreased the pro-apoptotic marker—Bax—and increased the anti-apoptotic marker—Bcl-2—in MPP+-induced SH-SY5Y cells | [81] | |
| Naringenin | 40 μM | Primary rat mesencephalic cultures | Decreased TH-positive neurons and TUNEL positive neurons | [82] | |
| Naringenin | 50 mg/kg | Male Sprague-Dawley rats | Restoration of dopamine concentrations due to neuroprotective effects rather than compensatory effects by remaining TH-positive cells after 6-OHDA lesioning | [83] | |
| Naringenin | 20, 40 and 80 mM (in vitro) 70 mg/kg, orally (in vivo) | Human neuroblastoma SH-SY5Y cells and male C57BL/6 mice | Activated Nrf2/ARE pathway in dopaminergic (in vitro) Up regulated protein levels of Nrf2/ARE genes (in vivo) Reduced striatal oxidative stress and subsequent apoptotic signalling cascades in striatum (in vivo) | [84] | |
| Naringenin | 12.5 μM and 25 μM | SH-SY5Y Human Neuroblastoma cell line | Downregulation of the expression of some Parkinsonian genes such as casp9, lrrk2, and polg and upregulate pink1 | [85] | |
| Naringenin | 25, 50, and 100 mg/ kg/p.o | Male C57BL/6J mice | Reduced NO content and restored SOD activity, also downregulated TNF-α and IL-1β expression | [86] | |
| Naringenin | 10 and 50 µM | Female Wistar rats | Enhanced astroglial neurotrophic effects on DA neurons through the regulation of Nrf2 activation, | [87] | |
| Anxiety and depression | Naringenin | 5, 10 and 20 mg/kg | Male ICR mice | Increased hippocampal 5-HT, NE and GR levels, and reduced serum corticosterone levels | [88] |
| Naringenin | 5, 10 and 20 mg/kg | Male ICR mice | Up-regulation of BDNF | [89] | |
| Naringenin | 10, 25 and 50 mg/kg | Swiss mice | Naringenin (25–50 mg/ kg) ameliorated the hypolocomotion, depressive- and anxiety-like behaviors in hypoxic mice Naringenin (10 mg/kg) increases BDNF expression but did not significantly (p < 0.05) alter corticosterone and catalase contents. The increased expressions of iNOS and NF-kB as well as loss of amygdala neuronal cells were reduced by naringenin (10 mg/kg) | [90] | |
| Naringin | 10 mg/kg | Adult male Swiss mice | Alleviation of the depressive and anxiogenic behaviors evidenced by the increased preference to sucrose and open arm entries and duration in SPT and EPM respectively | [91] | |
| Naringin | 25–100 mg/kg, i.p | Swiss mice | Increased the levels of GAD67, glutathione and decrease AChE activities, pro-inflammatory cytokines (TNF-α, IL-6), malondialdehyde, nitrite concentrations | [92] | |
| Naringenin | 50 mg/kg/day | Adult male Wistar rats | Mitigation of morphological anomalies in the hippocampal CA1 region and cortex and upregulation of BDNF, Shh, GLI1, NKX2.2, and PAX6 | [93] | |
| Huntington’s disease | Naringenin | 0.2, 0.4 mM | C3H10T1/2 cells | Suppression of the protein aggregation caused by EGFP-polyQ97 in mammalian cells. | [94] |
| Naringin | 50, 100 mg/kg | Male Wistar rats | Protection against 3-nitropropionic acid induced neurotoxicity via nitric oxide mechanism | [95] | |
| Naringenin | 50 mg/kg b.w, PO | Albino Wistar rats | Improvement of the behavioral function and restored the activity of MAO and 5-HT levels and reduction of the activation of astrocytes against 3-NP induced neurotoxicity | [96] | |
| Naringin | 40, and 80 mg/kg | Adult male Sprague-Dawley rats | Modulation of oxido-nitrosative stress, neuroinflammatory, apoptotic markers and mitochondrial complex activity | [97] | |
| Naringin | (80 mg/kg b.w/day, orally) | Male Wistar rats | Enhancement of phase II and antioxidant gene expressions via Nrf2 activation | [98] | |
| Naringin | 10 µM | PC12 cells | Modulation in expressions of B-cell lymphoma 2 and Bcl-2-associated X protein and enhancement of the nuclear translocation of Nrf2 | [99] | |
| Ischemic brain injury | Naringin | 50 and 100 mg/kg | Male Wistar rats | Restoration of reduced glutathione and catalase activity and mitochondrial enzyme activities in cortex, striatum, cerebellum | [100] |
| Naringin | 106 mg/kg/day | male C57BL/6 strain mice | Suppression of neuronal cell death, reversed the reduction in the level of phosphorylated calcium-calmodulin-dependent protein kinase II, had the tendency to reverse the reduction in the level of glutathione, and blockade of excessive activation of microglia and astrocytes | [101] | |
| Naringin | 40, 80 mg/kg | Male Sprague-Dawley rats | Improvement of early brain injury (EBI), including subarachnoid hemorrhage (SAH) severity, neurologic deficits, brain edema and blood-brain barrier integrity by attenuating SAH-induced oxidative stress and apoptosis, and reduction of the oxidative damage and apoptosis by inhibiting the activation of MAPK signaling pathway | [102] | |
| Naringenin | 50 and 100 mg/kg | Male Sprague–Dawley rats | Down-regulation of NOD2, RIP2, NF-κB, MMP-9 and up-regulation of claudin-5 expression | [103] | |
| Naringin | 80, 120, or 160 mg/kg/ day | SH-SY5Y cells | Reduced 3-nitrotyrosine formation, NADPH oxidase, and iNOS expression. Increased nNOS, p47, and p67 expression. Decreased mitophagy | [104] | |
| Naringin | 100 mg/kg/day | Adult Wistar male rats | Continual treatment increased SOD activity, decreased MDA, NO, iNOS, and IL-1β. It also improved rats’ behavioral performance | [105] | |
| Spinal cord injury | Naringin | 20, 40 mg/kg | Female Sprague-Dawley rat | Upregulation of the expression of NKx2.2 and 2′3′-cyclic nucleotide 3′-phosphodiesterase, and inhibition of β-catenin expression and GSK-3β phosphorylation | [106] |
| Naringenin | 5, 10, 15 mM | Male Wistar rats | Suppression of MMP-9 activity and upregulation of GSH, catalase and MMP-2 activation | [107] | |
| Naringenin | 50–100 mg/kg | Female Wistar rats | Repression of miR-223 | [108] | |
| Naringin | 25, 50, and 100 mg/kg | Adult Sprague Dawley rats | Reduction of TNF-α, IL-8 as well as MDAcontent and elevation of IL-10 as well as SOD activity | [109] | |
| Chronic hyperglycemic peripheral neuropathy | Naringenin | 50, 100 and 200 mg/kg | Male Sprague Dawley rats | Inhibition of upregulated expression of TNF-α, IL-1β and MCP-1 level; GFAP and Mac-1 mRNA expression | [110] |
| Naringenin | 25 and 50 mg/kg | Male Sprague Dawley rats | Increase GSH level and decrease MDA and NO level | [111] | |
| Naringin | 50 and 100 mg/kg, b.w | Rat model of OXL- induced peripheral neuropathy | Improved the level of superoxide dismutase, catalase, glutathione peroxidase, nuclear factor erythroid 2-related factor 2, Heme oxygenase-1, nuclear factor-κ B, tumor necrosis factor-α, interleukin-1β, Bax, Bcl-2, caspase-3, paraoxonase, mitogen-activated protein kinase 14, neuronal nitric oxide synthase (nNOS), acetylcholinesterase, and arginase 2 | [112] |
5.3. Cerebral Ischemia
5.4. Anxiety and Depression
5.5. Schizophrenia
6. Concluding Remarks and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PD | Parkinson’s disease |
| TRP | Transient receptor potential |
| NO | Nitric oxide |
| cGMP | Cyclic guanosine monophosphate |
| PKG | Protein kinase G |
| MAPK | Mitogen-activated protein kinase |
| GDNF | Growth differentiation and neurotrophic factor |
| AD | Alzheimer’s disease |
| Aβ | Amyloid-β |
| AChE | Acetylcholinesterase |
| CaMKII | Calcium/calmodulin-dependent protein kinase II |
| GSH | Reduced glutathione |
| GPx | Glutathione peroxidase |
| GST | Glutathione-S-transferase |
| SOD | Superoxide dismutase |
| ROS | Reactive oxygen species |
| LPO | Lipid peroxidation |
| MDA | Malondialdehyde |
| 5-HT | Serotonin |
| NE | Norepinephrine |
| BBB | Blood–brain barrier |
| iNOS | Inducible nitric oxide synthase |
References
- Patel, V.; Chisholm, D.; Dua, T.; Laxminarayan, R.; Medina-Mora, M.E. Mental, Neurological, and Substance Use Disorders: Disease Control Priorities; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2016; Volume 4. [Google Scholar]
- Zis, P.; Hadjivassiliou, M. Treatment of Neurological Manifestations of Gluten Sensitivity and Coeliac Disease. Curr. Treat. Options Neurol. 2019, 21, 10. [Google Scholar] [CrossRef] [PubMed]
- Ludvigsson, J.F.; Bai, J.C.; Biagi, F.; Card, T.R.; Ciacci, C.; Ciclitira, P.J.; Green, P.H.R.; Hadjivassiliou, M.; Holdoway, A.; Van Heel, D.A. Diagnosis and Management of Adult Coeliac Disease: Guidelines from the British Society of Gastroenterology. Gut 2014, 63, 1210–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapone, A.; Bai, J.C.; Ciacci, C.; Dolinsek, J.; Green, P.H.R.; Hadjivassiliou, M.; Kaukinen, K.; Rostami, K.; Sanders, D.S.; Schumann, M. Spectrum of Gluten-Related Disorders: Consensus on New Nomenclature and Classification. BMC Med. 2012, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuhr, U.; Kummert, A.L. The Fate of Naringin in Humans: A Key to Grapefruit Juice-drug Interactions? Clin. Pharmacol. Ther. 1995, 58, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Subhan, N.; Rahman, M.M.; Uddin, S.J.; Reza, H.M.; Sarker, S.D. Effect of Citrus Flavonoids, Naringin and Naringenin, on Metabolic Syndrome and Their Mechanisms of Action. Adv. Nutr. 2014, 5, 404–417. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Gao, W.; Liu, Z.; Zhang, Z.; Liu, C. Systematic Analysis of Main Constituents in Rat Biological Samples after Oral Administration of the Methanol Extract of Fructus aurantii by HPLC-ESI-MS/MS. Iran. J. Pharm. Res. 2014, 13, 493–503. [Google Scholar] [CrossRef]
- Ramesh, E.; Alshatwi, A.A. Naringin Induces Death Receptor and Mitochondria-Mediated Apoptosis in Human Cervical Cancer (SiHa) Cells. Food Chem. Toxicol. 2013, 51, 97–105. [Google Scholar] [CrossRef]
- Romagnolo, D.F.; Selmin, O.I. Flavonoids and Cancer Prevention: A Review of the Evidence. J. Nutr. Gerontol. Geriatr. 2012, 31, 206–238. [Google Scholar] [CrossRef]
- Bharti, S.; Rani, N.; Krishnamurthy, B.; Arya, D.S. Preclinical Evidence for the Pharmacological Actions of Naringin: A Review. Planta Med. 2014, 80, 437–451. [Google Scholar] [CrossRef] [Green Version]
- Esaki, S.; Nishiyama, K.; Sugiyama, N.; Nakajima, R.; Takao, Y.; Kamiya, S. Preparation and Taste of Certain Glycosides of Flavanones and of Dihydrochalcones. Biosci. Biotechnol. Biochem. 1994, 58, 1479–1485. [Google Scholar] [CrossRef]
- Felgines, C.; Texier, O.; Morand, C.; Manach, C.; Scalbert, A.; Régerat, F.; Remesy, C.; Felgines, C.; Texier, O.; Morand, C.; et al. Bioavailability of the Flavanone Naringenin and Its Glycosides in Rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G1148–G1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jadeja, R.N.; Devkar, R.V. Chapter 47—Polyphenols and Flavonoids in Controlling Non-Alcoholic Steatohepatitis. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 615–623. ISBN 978-0-12-398456-2. [Google Scholar]
- Manchope, M.F.; Ferraz, C.R.; Borghi, S.M.; Rasquel-Oliveira, F.S.; Franciosi, A.; Bagatim-Souza, J.; Dionisio, A.M.; Casagrande, R.; Verri, W.A. Chapter 38—Therapeutic Role of Naringenin to Alleviate Inflammatory Pain. In Treatments, Mechanisms, and Adverse Reactions of Anesthetics and Analgesics; Rajendram, R., Patel, V.B., Preedy, V.R., Martin, C.R., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 443–455. ISBN 978-0-12-820237-1. [Google Scholar]
- Li, Z.-X.; Cao, Y.; Yan, P. Topological Insulators and Semimetals in Classical Magnetic Systems. Phys. Rep. 2021, 915, 1–64. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Abaza, M.S.I.; Orabi, K.Y.; Al-Quattan, E.; Al-Attiyah, R.J. Growth Inhibitory and Chemo-Sensitization Effects of Naringenin, a Natural Flavanone Purified from Thymus Vulgaris, on Human Breast and Colorectal Cancer. Cancer Cell Int. 2015, 15, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ademosun, A.O.; Oboh, G.; Passamonti, S.; Tramer, F.; Ziberna, L.; Boligon, A.A.; Athayde, M.L. Inhibition of Metalloproteinase and Proteasome Activities in Colon Cancer Cells by Citrus Peel Extracts. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Hsiu, S.-L.; Huang, T.-Y.; Hou, Y.-C.; Chin, D.-H.; Chao, P.-D.L. Comparison of Metabolic Pharmacokinetics of Naringin and Naringenin in Rabbits. Life Sci. 2002, 70, 1481–1489. [Google Scholar] [CrossRef]
- Moghaddam, R.H.; Samimi, Z.; Moradi, S.Z.; Little, P.J.; Xu, S.; Farzaei, M.H. Naringenin and Naringin in Cardiovascular Disease Prevention: A Preclinical Review. Eur. J. Pharmacol. 2020, 887, 173535. [Google Scholar] [CrossRef]
- Zhang, L.; Song, L.; Zhang, P.; Liu, T.; Zhou, L.; Yang, G.; Lin, R.; Zhang, J. Solubilities of Naringin and Naringenin in Different Solvents and Dissociation Constants of Naringenin. J. Chem. Eng. Data 2015, 60, 932–940. [Google Scholar] [CrossRef]
- Fujioka, K.; Greenway, F.; Sheard, J.; Ying, Y. The Effects of Grapefruit on Weight and Insulin Resistance: Relationship to the Metabolic Syndrome. J. Med. Food 2006, 9, 49–54. [Google Scholar] [CrossRef]
- Ishii, K.; Furuta, T.; Kasuya, Y. Determination of Naringin and Naringenin in Human Plasma by High-Performance Liquid Chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1996, 683, 225–229. [Google Scholar] [CrossRef]
- Lee, C.-H.; Jeong, T.-S.; Choi, Y.-K.; Hyun, B.-H.; Oh, G.-T.; Kim, E.-H.; Kim, J.-R.; Han, J.-I.; Bok, S.-H. Anti-Atherogenic Effect of Citrus Flavonoids, Naringin and Naringenin, Associated with Hepatic ACAT and Aortic VCAM-1 and MCP-1 in High Cholesterol-Fed Rabbits. Biochem. Biophys. Res. Commun. 2001, 284, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Renugadevi, J.; Prabu, S.M. Naringenin Protects against Cadmium-Induced Oxidative Renal Dysfunction in Rats. Toxicology 2009, 256, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Cook, N.C.; Samman, S. Flavonoids—Chemistry, Metabolism, Cardioprotective Effects, and Dietary Sources. J. Nutr. Biochem. 1996, 7, 66–76. [Google Scholar] [CrossRef]
- Choudhury, R.; Chowrimootoo, G.; Srai, K.; Debnam, E.; Rice-Evans, C.A. Interactions of the Flavonoid Naringenin in the Gastrointestinal Tract and the Influence of Glycosylation. Biochem. Biophys. Res. Commun. 1999, 265, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Croft, K.D. The Chemistry and Biological Effects of Flavonoids and Phenolic Acids A. Ann. N. Y. Acad. Sci. 1998, 854, 435–442. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary Intake and Bioavailability of Polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef] [Green Version]
- Pietta, P.; Minoggio, M.; Bramati, L. Plant Polyphenols: Structure, Occurrence and Bioactivity. Stud. Nat. Prod. Chem. 2003, 28, 257–312. [Google Scholar] [CrossRef]
- Navipour, E.; Neamatshahi, M.; Barabadi, Z.; Neamatshahi, M.; Keykhosravi, A. Epidemiology and Risk Factors of Alzheimer’s Disease in Iran: A Systematic Review. Iran. J. Public Health 2019, 48, 2133–2139. [Google Scholar] [CrossRef]
- Pedersen, W.A.; McMillan, P.J.; Kulstad, J.J.; Leverenz, J.B.; Craft, S.; Haynatzki, G.R. Rosiglitazone Attenuates Learning and Memory Deficits in Tg2576 Alzheimer Mice. Exp. Neurol. 2006, 199, 265–273. [Google Scholar] [CrossRef]
- Rajmohan, R.; Reddy, P.H. Amyloid-Beta and Phosphorylated Tau Accumulations Cause Abnormalities at Synapses of Alzheimer’s Disease Neurons. J. Alzheimer’s Dis. 2017, 57, 975–999. [Google Scholar] [CrossRef]
- Jin, H.; Wang, W.; Zhao, S.; Yang, W.; Qian, Y.; Jia, N.; Feng, G. Aβ-HBc Virus-like Particles Immunization without Additional Adjuvant Ameliorates the Learning and Memory and Reduces Aβ Deposit in PDAPP Mice. Vaccine 2014, 32, 4450–4456. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, S.; Scheltens, P.; Cummings, J. Alzheimer’s Disease and Related Disorders; CRC Press: Boca Raton, FL, USA, 2005; ISBN 0203931742. [Google Scholar]
- Mimura, M.; Yano, M. Memory Impairment and Awareness of Memory Deficits in Early-Stage Alzheimer’s Disease. Rev. Neurosci. 2006, 17, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, S.; Abbaszadeh, F.; Dargahi, L.; Jorjani, M. Astaxanthin: A Mechanistic Review on Its Biological Activities and Health Benefits. Pharmacol. Res. 2018, 136, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.-Q.; Li, N.; Wang, T.; Kong, X.-C.; Tai, W.-J.; Sun, H.; Zhang, D. FLZ Alleviates the Memory Deficits in Transgenic Mouse Model of Alzheimer’s Disease via Decreasing Beta-Amyloid Production and Tau Hyperphosphorylation. PLoS ONE 2013, 8, e78033. [Google Scholar] [CrossRef]
- Mimura, M. Memory Impairment and Awareness of Memory Deficits in Early-Stage Alzheimer’s Disease. Tohoku J. Exp. Med. 2008, 215, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Obulesu, M.; Jhansilakshmi, M. Neuroinflammation in Alzheimer’s Disease: An Understanding of Physiology and Pathology. Int. J. Neurosci. 2014, 124, 227–235. [Google Scholar] [CrossRef]
- Yang, W.; Ma, J.; Liu, Z.; Lu, Y.; Hu, B.; Yu, H. Effect of Naringenin on Brain Insulin Signaling and Cognitive Functions in ICV-STZ Induced Dementia Model of Rats. Neurol. Sci. 2014, 35, 741–751. [Google Scholar] [CrossRef]
- Burke, R.E.; O’Malley, K. Axon Degeneration in Parkinson’s Disease. Exp. Neurol. 2013, 246, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Savitt, J.M.; Dawson, V.L.; Dawson, T.M. Diagnosis and Treatment of Parkinson Disease: Molecules to Medicine. J. Clin. Investig. 2006, 116, 1744–1754. [Google Scholar] [CrossRef] [Green Version]
- Shulman, J.M.; De Jager, P.L.; Feany, M.B. Parkinson’s Disease: Genetics and Pathogenesis. Annu. Rev. Pathol. Mech. Dis. 2011, 6, 193–222. [Google Scholar] [CrossRef]
- Dexter, D.T.; Jenner, P. Parkinson Disease: From Pathology to Molecular Disease Mechanisms. Free Radic. Biol. Med. 2013, 62, 132–144. [Google Scholar] [CrossRef] [PubMed]
- AlDakheel, A.; Kalia, L.V.; Lang, A.E. Pathogenesis-Targeted, Disease-Modifying Therapies in Parkinson Disease. Neurotherapeutics 2014, 11, 6–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olanow, C.W.; Tatton, W.G. Etiology and Pathogenesis of Parkinson’s Disease. Annu. Rev. Neurosci. 1999, 22, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zbarsky, V.; Datla, K.P.; Parkar, S.; Rai, D.K.; Aruoma, O.I.; Dexter, D.T. Neuroprotective Properties of the Natural Phenolic Antioxidants Curcumin and Naringenin but Not Quercetin and Fisetin in a 6-OHDA Model of Parkinson’s Disease. Free Radic. Res. 2005, 39, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Golechha, M.; Chaudhry, U.; Bhatia, J.; Saluja, D.; Arya, D.S. Naringin Protects against Kainic Acid-Induced Status Epilepticus in Rats: Evidence for an Antioxidant, Anti-Inflammatory and Neuroprotective Intervention. Biol. Pharm. Bull. 2011, 34, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Khan, H.; Aschner, M.; Hasan, M.M.; Hassan, S.T.S. Therapeutic Potential of Naringin in Neurological Disorders. Food Chem. Toxicol. 2019, 132, 110646. [Google Scholar] [CrossRef]
- Raza, S.S.; Khan, M.M.; Ahmad, A.; Ashafaq, M.; Islam, F.; Wagner, A.P.; Safhi, M.M. Neuroprotective Effect of Naringenin Is Mediated through Suppression of NF-ΚB Signaling Pathway in Experimental Stroke. Neuroscience 2013, 230, 157–171. [Google Scholar] [CrossRef]
- Harrison, W.T.A.; Yathirajan, H.S.; Sarojini, B.K.; Narayana, B.; Anilkumar, H.G. Do C—H⋯ O and C—H⋯ π Interactions Help to Stabilize a Non-Centrosymmetric Structure for Racemic 2, 3-Dibromo-1, 3-Diphenylpropan-1-One? Acta Crystallogr. Sect. C Cryst. Struct. Commun. 2005, 61, o728–o730. [Google Scholar] [CrossRef]
- Gopinathan, G.; Teravainen, H.; Dambrosia, J.M.; Ward, C.D.; Sanes, J.N.; Stuart, W.K.; Evarts, E.V.; Calne, D.B. Lisuride in Parkinsonism. Neurology 1981, 31, 371. [Google Scholar] [CrossRef]
- Kim, H.D.; Jeong, K.H.; Jung, U.J.; Kim, S.R. Naringin Treatment Induces Neuroprotective Effects in a Mouse Model of Parkinson’s Disease in Vivo, but Not Enough to Restore the Lesioned Dopaminergic System. J. Nutr. Biochem. 2016, 28, 140–146. [Google Scholar] [CrossRef]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell Survival Responses to Environmental Stresses via the Keap1-Nrf2-ARE Pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-M.; Johnson, J.A. An Important Role of Nrf2-ARE Pathway in the Cellular Defense Mechanism. BMB Rep. 2004, 37, 139–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eagling, V.A.; Profit, L.; Back, D.J. Inhibition of the CYP3A4-Mediated Metabolism and P-Glycoprotein-Mediated Transport of the HIV-1 Protease Inhibitor Saquinavir by Grapefruit Juice Components. Br. J. Clin. Pharmacol. 1999, 48, 543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachdeva, A.K.; Kuhad, A.; Chopra, K. Naringin Ameliorates Memory Deficits in Experimental Paradigm of Alzheimer’s Disease by Attenuating Mitochondrial Dysfunction. Pharmacol. Biochem. Behav. 2014, 127, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Gao, K.; Li, X.; Shen, X.; Zhang, X.; Ma, C.; Qin, C.; Zhang, L. Long-Term Naringin Consumption Reverses a Glucose Uptake Defect and Improves Cognitive Deficits in a Mouse Model of Alzheimer’s Disease. Pharmacol. Biochem. Behav. 2012, 102, 13–20. [Google Scholar] [CrossRef]
- Kaur, G.; Prakash, A. Involvement of the Nitric Oxide Signaling in Modulation of Naringin against Intranasal Manganese and Intracerbroventricular β-Amyloid Induced Neurotoxicity in Rats. J. Nutr. Biochem. 2020, 76, 108255. [Google Scholar] [CrossRef]
- Meng, X.; Fu, M.; Wang, S.; Chen, W.; Wang, J.; Zhang, N. Naringin Ameliorates Memory Deficits and Exerts Neuroprotective Effects in a Mouse Model of Alzheimer’s Disease by Regulating Multiple Metabolic Pathways. Mol. Med. Rep. 2021, 23, 332. [Google Scholar] [CrossRef]
- Heo, H.J.; Kim, M.J.; Lee, J.M.; Choi, S.J.; Cho, H.Y.; Hong, B.; Kim, H.K.; Kim, E.; Shin, D.H. Naringenin from Citrus Junos Has an Inhibitory Effect on Acetylcholinesterase and a Mitigating Effect on Amnesia. Dement. Geriatr. Cogn. Disord. 2004, 17, 151–157. [Google Scholar] [CrossRef]
- Khan, M.B.; Khan, M.M.; Khan, A.; Ahmed, M.E.; Ishrat, T.; Tabassum, R.; Vaibhav, K.; Ahmad, A.; Islam, F. Naringenin Ameliorates Alzheimer’s Disease (AD)-Type Neurodegeneration with Cognitive Impairment (AD-TNDCI) Caused by the Intracerebroventricular- Streptozotocin in Rat Model. Neurochem. Int. 2012, 61, 1081–1093. [Google Scholar] [CrossRef]
- Heo, H.J.; Kim, D.O.; Shin, S.C.; Kim, M.J.; Kim, B.G.; Shin, D.H. Effect of Antioxidant Flavanone, Naringenin, from Citrus Junos on Neuroprotection. J. Agric. Food Chem. 2004, 52, 1520–1525. [Google Scholar] [CrossRef]
- Rahigude, A.; Bhutada, P.; Kaulaskar, S.; Aswar, M.; Otari, K. Participation of Antioxidant and Cholinergic System in Protective Effect of Naringenin against Type-2 Diabetes-Induced Memory Dysfunction in Rats. Neuroscience 2012, 226, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Zhou, K.; Zhou, Y.; An, Y.; Hu, T.; Lu, J.; Huang, S.; Pei, G. Naringin Dihydrochalcone Ameliorates Cognitive Deficits and Neuropathology in APP/PS1 Transgenic Mice. Front. Aging Neurosci. 2018, 10, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Dogra, S.; Prakash, A. Protective Effect of Naringin, a Citrus Flavonoid, against Colchicine-Induced Cognitive Dysfunction and Oxidative Damage in Rats. J. Med. Food 2010, 13, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Jahanshahi, M.; Khalili, M.; Margedari, A. Naringin Chelates Excessive Iron and Prevents the Formation of Amyloid-Beta Plaques in the Hippocampus of Iron-Overloaded Mice. Front. Pharmacol. 2021, 12, 518. [Google Scholar] [CrossRef]
- Prabhakar, O. Naringin Attenuates Aluminum Induced Cognitive Deficits in Rats. Indian J. Pharm. Educ. Res. 2020, 54, 674–681. [Google Scholar] [CrossRef]
- Guo, L.; Sun, B. N,N’-1,10-Bis(Naringin) Triethylenetetraamine, Synthesis and as a Cu(II) Chelator for Alzheimer’s Disease Therapy. Biol. Pharm. Bull. 2021, 44, 51–56. [Google Scholar] [CrossRef]
- Ben-Azu, B.; Nwoke, E.; Aderibigbe, A.; Omogbiya, I.; Ajayi, A.; Olonode, E.; Umukoro, S.; Iwalewa, E. Possible Neuroprotective Mechanisms of Action Involved in the Neurobehavioral Property of Naringin in Mice. Biomed. Pharmacother. 2019, 109, 536–546. [Google Scholar] [CrossRef]
- Varshney, V.; Garabadu, D. Naringin Exhibits Mas Receptor-Mediated Neuroprotection Against Amyloid Beta-Induced Cognitive Deficits and Mitochondrial Toxicity in Rat Brain. Neurotox. Res. 2021, 39, 1023–1043. [Google Scholar] [CrossRef]
- Ghofrani, S.; Joghataei, M.; Mohseni, S.; Baluchnejadmojarad, T.; Bagheri, M.; Khamse, S.; Roghani, M. Naringenin Improves Learning and Memory in an Alzheimer’s Disease Rat Model: Insights into the Underlying Mechanisms. Eur. J. Pharmacol. 2015, 764, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Jung, U.; Kim, S. Effects of Naringin, a Flavanone Glycoside in Grapefruits and Citrus Fruits, on the Nigrostriatal Dopaminergic Projection in the Adult Brain. Neural Regen. Res. 2014, 9, 1514. [Google Scholar]
- Leem, E.; Nam, J.; Jeon, M.; Shin, W.; Won, S.; Park, S.; Choi, M.; Jin, B.; Jung, U.; Kim, S. Naringin Protects the Nigrostriatal Dopaminergic Projection through Induction of GDNF in a Neurotoxin Model of Parkinson’s Disease. J. Nutr. Biochem. 2014, 25, 801–806. [Google Scholar] [CrossRef]
- Garabadu, D.; Agrawal, N. Naringin Exhibits Neuroprotection Against Rotenone-Induced Neurotoxicity in Experimental Rodents. NeuroMolecular Med. 2020, 22, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; James, R.; Sinha, S.; Kinra, M.; Anuranjana, P.V.; Nandakumar, K. Naringin Exhibited Anti-Parkinsonian like Effect against Haloperidol-Induced Catalepsy in Mice. Res. J. Pharm. Technol. 2021, 14, 662–666. [Google Scholar] [CrossRef]
- Sugumar, M.; Sevanan, M.; Sekar, S. Neuroprotective Effect of Naringenin against MPTP-Induced Oxidative Stress. Int. J. Neurosci. 2018, 129, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Madani, M.; Attia, A.; ElSalalm, R.; El-Shenawy, S.; Arbid, M. Neuropharmacological Effects of Naringenin, Harmine and Adenosine on Parkinsonism Induced in Rats. Der Pharm. Lett. 2016, 8, 45–57. [Google Scholar]
- Chen, C.; Wei, Y.; He, X.; Li, D.; Wang, G.; Li, J. Naringenin Produces Neuroprotection Against LPS-Induced Dopamine Neurotoxicity via the Inhibition of Microglial NLRP3 Inflammasome Activation. Front. Immunol. 2019, 10, 936. [Google Scholar] [CrossRef] [Green Version]
- Mani, S.; Sekar, S.; Chidambaram, S.B.; Sevanan, M. Naringenin Protects against 1-Methyl-4-Phenylpyridinium-Induced Neuroinflammation and Resulting Reactive Oxygen Species Production in SH - SY5Y Cell Line: An In Vitro Model of Parkinson’s Disease. Pharmacogn. Mag. 2018, 458–464. [Google Scholar] [CrossRef]
- Mercer, L.; Kelly, B.; Horne, M.; Beart, P. Dietary Polyphenols Protect Dopamine Neurons from Oxidative Insults and Apoptosis: Investigations in Primary Rat Mesencephalic Cultures. Biochem. Pharmacol. 2005, 69, 339–345. [Google Scholar] [CrossRef]
- Kabir, M.S.H.; Hossain, M.M.; Kabir, M.I.; Rahman, M.M.; Hasanat, A.; Emran, T.B.; Rahman, M.A. Phytochemical screening, Antioxidant, Thrombolytic, alpha-amylase inhibition and cytotoxic activities of ethanol extract of Steudnera colocasiifolia K. Koch leaves. J. Young Pharm. 2016, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Lou, H.; Jing, X.; Wei, X.; Shi, H.; Ren, D.; Zhang, X. Naringenin Protects against 6-OHDA-Induced Neurotoxicity via Activation of the Nrf2/ARE Signaling Pathway. Neuropharmacology 2014, 79, 380–388. [Google Scholar] [CrossRef]
- Kesh, S.; Rajesh, R.; Balakrishnan, A. Naringenin Alleviates 6-Hydroxydopamine Induced Parkinsonism in SHSY5Y Cells and Zebrafish Model. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 239, 108893. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.; Sekar, S.; Barathidasan, R.; Manivasagam, T.; Thenmozhi, A.J.; Sevanan, M.; Chidambaram, S.B.; Essa, M.M.; Guillemin, G.J.; Sakharkar, M.K. Naringenin Decreases α -Synuclein Expression and Neuroinflammation in MPTP-Induced Parkinson’s Disease Model in Mice. Neurotox. Res. 2018, 33, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, B.; He, X.; Li, D.; Shi, J.; Zhang, F. Naringenin Targets on Astroglial Nrf2 to Support Dopaminergic Neurons. Pharmacol. Res. 2018, 139, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Li, J.; Li, H.; Su, D.; Quan, X.; He, X.; Wang, X. Antidepressant-like Behavioral, Neurochemical and Neuroendocrine Effects of Naringenin in the Mouse Repeated Tail Suspension Test. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 39, 175–181. [Google Scholar] [CrossRef]
- Yi, L.; Liu, B.; Li, J.; Luo, L.; Liu, Q.; Geng, D.; Tang, Y.; Xia, Y.; Wu, D. BDNF Signaling Is Necessary for the Antidepressant-like Effect of Naringenin. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 48, 135–141. [Google Scholar] [CrossRef]
- Olugbemide, A.S.; Ben-Azu, B.; Bakre, A.G.; Ajayi, A.M.; Femi-Akinlosotu, O.; Umukoro, S. Naringenin Improves Depressive- and Anxiety-like Behaviors in Mice Exposed to Repeated Hypoxic Stress through Modulation of Oxido-Inflammatory Mediators and NF-KB/BDNF Expressions. Brain Res. Bull. 2021, 169, 214–227. [Google Scholar] [CrossRef]
- Anyanwu, G.E.; Atuadu, V.O.; Esom, E.A.; Nto, J.N.; Katchy, A.U. Putative Role of Monoaminergic Systems in Antidepressant and Anxiolytic Effects of Naringin in Mice: An Interaction Study with Receptor Antagonists. J. Pharm. Res. Int. 2021, 33, 661–676. [Google Scholar] [CrossRef]
- Oladapo, O.M.; Ben-azu, B.; Ajayi, A.M.; Emokpae, O.; Eneni, A.O. Naringin Confers Protection against Psychosocial Defeat Stress-Induced Neurobehavioral Deficits in Mice: Involvement of Glutamic Acid Decarboxylase Isoform-67, Oxido-Nitrergic Stress, and Neuroinflammatory Mechanisms. J. Mol. Neurosci. 2020, 71, 431–445. [Google Scholar] [CrossRef]
- Tayyab, M.; Farheen, S.; Mariyath, M.; Khanam, P.M.N.; Hossain, M.M. Antidepressant and Neuroprotective Effects of Naringenin via Sonic Hedgehog-GLI1 Cell Signaling Pathway in a Rat Model of Chronic Unpredictable Mild Stress. Neuromol. Med. 2019, 21, 250–261. [Google Scholar] [CrossRef]
- Yamagishi, N.; Yamamoto, Y.; Noda, C.; Hatayama, T. Naringenin Inhibits the Aggregation of Expanded Polyglutamine Tract-Containing Protein through the Induction of Endoplasmic Reticulum Chaperone GRP78. Biol. Pharm. Bull. 2012, 35, 1836–1840. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Kumar, A. Protective Effect of Hesperidin and Naringin against 3-Nitropropionic Acid Induced Huntington’ s like Symptoms in Rats: Possible Role of Nitric Oxide. Behav. Brain Res. 2010, 206, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Salman, M.; Sharma, P.; Alam, I.; Tabassum, H. Naringenin Mitigates Behavioral Alterations and Provides Neuroprotection against 3-Nitropropinoic Acid-Induced Huntington’s Disease like Symptoms in Rats. Nutr. Neurosci. 2021, 25, 1–11. [Google Scholar] [CrossRef]
- Cui, J.; Wang, G.; Kandhare, A.D.; Mukherjee, A.A.; Bodhankar, S.L. Neuroprotective Effect of Naringin, a Flavone Glycoside in Quinolinic Acid-Induced Neurotoxicity: Possible Role of PPAR-γ, Bax/Bcl-2, and Caspase-3. Food Chem. Toxicol. 2018, 121, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, K.; Sudhandiran, G. Naringin modulates oxidative stress and inflammation in 3-nitropropionic acid-induced neurodegeneration through the activation of nuclear factor-erythroid 2-related factor-2 signalling pathway. Neuroscience 2012, 227, 134–143. [Google Scholar] [CrossRef]
- Kulasekaran, G.; Ganapasam, S. Neuroprotective Efficacy of Naringin on 3-Nitropropionic Acid- Induced Mitochondrial Dysfunction through the Modulation of Nrf2 Signaling Pathway in PC12 Cells. Mol. Cell. Biochem. 2015, 409, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Gaur, V.; Aggarwal, A.; Kumar, A. Protective Effect of Naringin against Ischemic Reperfusion Cerebral Injury: Possible Neurobehavioral, Biochemical and Cellular Alterations in Rat Brain. Eur. J. Pharmacol. 2009, 616, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Yamamoto, K.; Mori, H.; Sawamoto, A.; Amakura, Y. Neuroprotective Effect of Citrus Kawachiensis (Kawachi Bankan) Peels, a Rich Source of Naringin, against Global Cerebral Ischemia/Reperfusion Injury in Mice. Biosci. Biotechnol. Biochem. 2018, 82, 1216–1224. [Google Scholar] [CrossRef]
- Han, Y.; Su, J.; Liu, X.; Zhao, Y.; Wang, C.; Li, X. Naringin Alleviates Early Brain Injury after Experimental Subarachnoid Hemorrhage by Reducing Oxidative Stress and Inhibiting Apoptosis. Brain Res. Bull. 2016, 133, 42–50. [Google Scholar] [CrossRef]
- Bai, X.; Zhang, X.; Chen, L. Protective Effect of Naringenin in Experimental Ischemic Stroke: Down-Regulated NOD2, RIP2, NF-KappaB, MMP-9 and Up-Regulated Claudin-5 Expression. Neurochem. Res. 2014, 39, 1405–1415. [Google Scholar] [CrossRef]
- Feng, J.; Chen, X.; Lu, S.; Li, W.; Yang, D.; Su, W.; Wang, X. Naringin Attenuates Cerebral Ischemia-Reperfusion Injury Through Inhibiting Peroxynitrite-Mediated Mitophagy Activation. Mol. Neurobiol. 2018, 55, 9029–9042. [Google Scholar] [CrossRef]
- Cui, Q.; Wang, L.; Wei, Z.; Qu, W. Continual Naringin Treatment Benefits the Recovery of Traumatic Brain Injury in Rats Through Reducing Oxidative and Inflammatory Alterations. Neurochem. Res. 2014, 39, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Rong, W.; Pan, Y.; Cai, X.; Song, F.; Zhao, Z.; Xiao, S.; Zhang, C. The Mechanism of Naringin-Enhanced Remyelination after Spinal Cord Injury. Neural Regen. Res. 2017, 12, 470. [Google Scholar] [PubMed]
- Fakhri, S.; Sabouri, S.; Kiani, A.; Farzaei, M.H.; Rashidi, K. Intrathecal Administration of Naringenin Improves Motor Dysfunction and Neuropathic Pain Following Compression Spinal Cord Injury in Rats: Relevance to Its Antioxidant and Anti-Inflammatory Activities. Korean J. Pain 2022, 35, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Tang, P.; Zhang, W.; Zhao, Y.; Zhang, L.; Zhang, H. Naringenin Inhibits Spinal Cord Injury-Induced Activation of Neutrophils through MiR-223. Gene 2016, 592, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Z.; Wang, H. Naringin Attenuates Cerebral Ischemia-Reperfusion Injury in Rats by Inhibiting Endoplasmic Reticulum Stress. Transl. Neurosci. 2021, 12, 190–197. [Google Scholar] [CrossRef]
- Hu, C.; Zhao, Y. Analgesic Effects of Naringenin in Rats with Spinal Nerve Ligation-Induced Neuropathic Pain. Biomed. Rep. 2014, 2, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Kaulaskar, S.; Bhutada, P.; Rahigude, A.; Jain, D.; Harle, U. Effects of Naringenin on Allodynia and Hyperalgesia in Rats with Chronic Constriction Injury-Induced Neuropathic Pain. J. Chin. Integr. Med. 2012, 10, 1482–1489. [Google Scholar] [CrossRef]
- Semis, H.; Kandemir, F.; Caglayan, C.; Kaynar, O.; Genc, A.; Arıkan, S. Protective Effect of Naringin against Oxaliplatin-induced Peripheral Neuropathy in Rats: A Behavioral and Molecular Study. J. Biochem. Mol. Toxicol. 2022, 36, e23121. [Google Scholar] [CrossRef]
- Yang, J.; Yuan, L.; Wen, Y.; Zhou, H.; Jiang, W.; Xu, D.; Wang, M. Protective Effects of Naringin in Cerebral Infarction and Its Molecular Mechanism. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e918772. [Google Scholar] [CrossRef]
- Sveinsson, O.A.; Kjartansson, O.; Valdimarsson, E.M. Cerebral Ischemia/Infarction-Diagnosis and Treatment. Laeknabladid 2014, 100, 393–401. [Google Scholar]
- Olmez, I.; Ozyurt, H. Reactive Oxygen Species and Ischemic Cerebrovascular Disease. Neurochem. Int. 2012, 60, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Tsung, A.; Klune, J.R.; Zhang, X.; Jeyabalan, G.; Cao, Z.; Peng, X.; Stolz, D.B.; Geller, D.A.; Rosengart, M.R.; Billiar, T.R. HMGB1 Release Induced by Liver Ischemia Involves Toll-like Receptor 4–Dependent Reactive Oxygen Species Production and Calcium-Mediated Signaling. J. Exp. Med. 2007, 204, 2913–2923. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.L.; Song, L.; Yu, X.D.; Yu, T.X.; Liang, H.; Qiu, J.X. The Clinical Study on the Treatment for Acute Cerebral Infarction by Intra-Arterial Thrombolysis Combined with Mild Hypothermia. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1999–2006. [Google Scholar]
- Jeong, H.S.; Song, H.-J.; Kim, S.-B.; Lee, J.; Kang, C.W.; Koh, H.-S.; Shin, J.E.; Lee, S.H.; Kwon, H.J.; Kim, J. A Comparison of Stent-Assisted Mechanical Thrombectomy and Conventional Intra-Arterial Thrombolysis for Acute Cerebral Infarction. J. Clin. Neurol. 2013, 9, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.G.; Arnold, J.M.O.; Strong, H.A.; Munoz, C.; Spence, J.D. Effect of Grapefruit Juice and Naringin on Nisoldipine Pharmacokinetics. Clin. Pharmacol. Ther. 1993, 54, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Feng, S.-J.; Kan, M.-C. Naringin Targets NFKB1 to Alleviate Oxygen-Glucose Deprivation/Reoxygenation–Induced Injury in PC12 Cells via Modulating HIF-1α/AKT/MTOR-Signaling Pathway. J. Mol. Neurosci. 2021, 71, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Wu, C.; Guo, S.; Su, J.; Zhao, W.; Xing, H. Higenamine Protects Neuronal Cells from Oxygen-glucose Deprivation/Reoxygenation-induced Injury. J. Cell. Biochem. 2019, 120, 3757–3764. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Zhang, Y. Protodioscin Protects PC12 Cells against Oxygen and Glucose Deprivation-Induced Injury through MiR-124/AKT/Nrf2 Pathway. Cell Stress Chaperones 2019, 24, 1091–1099. [Google Scholar] [CrossRef]
- Wang, C.; Yang, Y.-H.; Zhou, L.; Ding, X.-L.; Meng, Y.-C.; Han, K. Curcumin Alleviates OGD/R-Induced PC12 Cell Damage via Repressing CCL3 and Inactivating TLR4/MyD88/MAPK/NF-ΚB to Suppress Inflammation and Apoptosis. J. Pharm. Pharmacol. 2020, 72, 1176–1185. [Google Scholar] [CrossRef]
- Rahman, J.; Tareq, A.M.; Hossain, M.; Sakib, S.A.; Islam, M.N.; Ali, M.H.; Uddin, A.B.M.N.; Hoque, M.; Nasrin, M.S.; Emran, T.B.; et al. Biological evaluation, DFT calculations and molecular docking studies on the antidepressant and cytotoxicity activities of Cycas pectinata Buch. - Ham. Compounds. Pharmaceuticals 2020, 13, 232. [Google Scholar] [CrossRef]
- Tejera, D.; Heneka, M.T. Microglia in Neurodegenerative Disorders. In Microglia; Springer: New York, NY, USA, 2019; pp. 57–67. [Google Scholar]
- Abdel-Magied, N.; Shedid, S.M. The Effect of Naringenin on the Role of Nuclear Factor (Erythroid-derived 2)-like2 (Nrf2) and Haem Oxygenase 1 (HO-1) in Reducing the Risk of Oxidative Stress-related Radiotoxicity in the Spleen of Rats. Environ. Toxicol. 2019, 34, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Drishya, S.; Dhanisha, S.S.; Raghukumar, P.; Guruvayoorappan, C. Amomum Subulatum Mitigates Total Body Irradiation-Induced Oxidative Stress and Associated Inflammatory Responses by Enhancing the Antioxidant Status and Regulating pro-Inflammatory Cytokines. J. Nutr. Biochem. 2022, 107, 109064. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Fu, Q.; Li, Y.; Li, S.; Xue, J.; Ma, S. Emodin Opposes Chronic Unpredictable Mild Stress Induced Depressive-like Behavior in Mice by Upregulating the Levels of Hippocampal Glucocorticoid Receptor and Brain-Derived Neurotrophic Factor. Fitoterapia 2014, 98, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Anjum, J.; Muni, M.; Das, R.; Rauf, A.; Islam, F.; Emran, T.B.; Semwal, P.; Hemeg, H.A.; Alhumaydhi, F.A. Exploring the Journey of Emodin as a Potential Neuroprotective Agent: Novel Therapeutic Insights with Molecular Mechanism of Action. Biomed. Pharmacother. 2022, 149, 112877. [Google Scholar] [CrossRef]
- Mitra, S.; Lami, M.S.; Uddin, T.M.; Das, R.; Islam, F.; Anjum, J.; Hossain, M.J.; Emran, T.B. Prospective multifunctional roles and pharmacological potential of dietary flavonoid narirutin. Biomed. Pharmacother. 2022, 150, 112932. [Google Scholar] [CrossRef]
- Halbreich, U.; Kahn, L.S. Role of Estrogen in the Aetiology and Treatment of Mood Disorders. CNS Drugs 2001, 15, 797–817. [Google Scholar] [CrossRef]
- Bruce, M.L.; Raue, P.J. Mental Illness as Psychiatric Disorder. In Handbook of the Sociology of Mental Health; Springer: Berlin/Heidelberg, Germany, 2013; pp. 41–59. [Google Scholar]
- Evans, D.L.; Foa, E.B.; Gur, R.E.; Hendin, H.; O’Brien, C.P.; Seligman, M.E.P.; Walsh, B.T. Treating and Preventing Adolescent Mental Health Disorders: What We Know and What We Don’t Know; Oxford University Press: Oxford, UK, 2005; ISBN 0198038712. [Google Scholar]
- Chtourou, Y.; Gargouri, B.; Kebieche, M.; Fetoui, H. Naringin Abrogates Cisplatin-Induced Cognitive Deficits and Cholinergic Dysfunction through the down-Regulation of AChE Expression and INOS Signaling Pathways in Hippocampus of Aged Rats. J. Mol. Neurosci. 2015, 56, 349–362. [Google Scholar] [CrossRef]
- Kwatra, M.; Jangra, A.; Mishra, M.; Sharma, Y.; Ahmed, S.; Ghosh, P.; Kumar, V.; Vohora, D.; Khanam, R. Naringin and Sertraline Ameliorate Doxorubicin-Induced Behavioral Deficits through Modulation of Serotonin Level and Mitochondrial Complexes Protection Pathway in Rat Hippocampus. Neurochem. Res. 2016, 41, 2352–2366. [Google Scholar] [CrossRef]
- Tallei, T.E.; Niode, N.J.; Idroes, R.; Zidan, B.M.R.M.; Mitra, S.; Celik, I.; Nainu, F.; Ağagündüz, D.; Emran, T.B.; Capasso, R.A. Comprehensive Review of the Potential Use of Green Tea Polyphenols in the Management of COVID-19 Evid-based Complement. Altern. Med. 2021, 2021, 7170736. [Google Scholar] [CrossRef]
- D’Angelo, S.; Mele, E.; Di Filippo, F.; Viggiano, A.; Meccariello, R. Sirt1 Activity in the Brain: Simultaneous Effects on Energy Homeostasis and Reproduction. Int. J. Environ. Res. Public Health 2021, 18, 1243. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, I.; Domínguez, A.P.; Drägert, K.; Haas, E.; Battegay, E.J. Hypoxia Potentiates Tumor Necrosis Factor-α Induced Expression of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 in White and Brown Adipocytes. Biochem. Biophys. Res. Commun. 2015, 461, 287–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matheus, M.E.; de Almeida Violante, F.; Garden, S.J.; Pinto, A.C.; Fernandes, P.D. Isatins Inhibit Cyclooxygenase-2 and Inducible Nitric Oxide Synthase in a Mouse Macrophage Cell Line. Eur. J. Pharmacol. 2007, 556, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-S.; Park, Y.M.; Choi, J.-H.; Park, H.-J.; Shin, M.C.; Lee, Y.S.; Lee, K.-T. Sulfuretin Isolated from Heartwood of Rhus Verniciflua Inhibits LPS-Induced Inducible Nitric Oxide Synthase, Cyclooxygenase-2, and pro-Inflammatory Cytokines Expression via the down-Regulation of NF-ΚB in RAW 264.7 Murine Macrophage Cells. Int. Immunopharmacol. 2010, 10, 943–950. [Google Scholar] [CrossRef]
- Feifel, D.; Shilling, P.D.; MacDonald, K. A Review of Oxytocin’s Effects on the Positive, Negative, and Cognitive Domains of Schizophrenia. Biol. Psychiatry 2016, 79, 222–233. [Google Scholar] [CrossRef] [Green Version]
- Lisman, J.E.; Coyle, J.T.; Green, R.W.; Javitt, D.C.; Benes, F.M.; Heckers, S.; Grace, A.A. Circuit-Based Framework for Understanding Neurotransmitter and Risk Gene Interactions in Schizophrenia. Trends Neurosci. 2008, 31, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Obrosova, I.G. Diabetes and the Peripheral Nerve. Biochim. Biophys. Acta Mol. Basis Dis. 2009, 1792, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Howes, O.; McCutcheon, R.; Stone, J. Glutamate and Dopamine in Schizophrenia: An Update for the 21st Century. J. Psychopharmacol. 2015, 29, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Li, P.-F.; Dietz, R.; von Harsdorf, R. P53 Regulates Mitochondrial Membrane Potential through Reactive Oxygen Species and Induces Cytochrome C-Independent Apoptosis Blocked by Bcl-2. EMBO J. 1999, 18, 6027–6036. [Google Scholar] [CrossRef] [Green Version]
- Zou, W.; Xiao, Z.; Wen, X.; Luo, J.; Chen, S.; Cheng, Z.; Xiang, D.; Hu, J.; He, J. The Anti-Inflammatory Effect of Andrographis paniculata (Burm. f.) Nees on Pelvic Inflammatory Disease in Rats through down-Regulation of the NF-ΚB Pathway. BMC Complement. Altern. Med. 2016, 16, 483. [Google Scholar] [CrossRef] [Green Version]
- Stenvinkel, P.; Ketteler, M.; Johnson, R.J.; Lindholm, B.; Pecoits-Filho, R.; Riella, M.; Heimbürger, O.; Cederholm, T.; Girndt, M. IL-10, IL-6, and TNF-α: Central Factors in the Altered Cytokine Network of Uremia—The Good, the Bad, and the Ugly. Kidney Int. 2005, 67, 1216–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Q.; Pecoits-Filho, R.; Lindholm, B.; Stenvinkel, P. Traditional and Non-Traditional Risk Factors as Contributors to Atherosclerotic Cardiovascular Disease in End-Stage Renal Disease. Scand. J. Urol. Nephrol. 2004, 38, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Higley, M.J.; Picciotto, M.R. Neuromodulation by Acetylcholine: Examples from Schizophrenia and Depression. Curr. Opin. Neurobiol. 2014, 29, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Futamura, T.; Toyooka, K.; Iritani, S.; Niizato, K.; Nakamura, R.; Tsuchiya, K.; Someya, T.; Kakita, A.; Takahashi, H.; Nawa, H. Abnormal Expression of Epidermal Growth Factor and Its Receptor in the Forebrain and Serum of Schizophrenic Patients. Mol. Psychiatry 2002, 7, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Yu, R.; Zhou, Q.; Liu, P.; Liu, Z.; Bian, Y. Naringenin Prevents TNF-α-Induced Gut-Vascular Barrier Disruption Associated with Inhibiting the NF-ΚB-Mediated MLCK/p-MLC and NLRP3 Pathways. Food Funct. 2021, 12, 2715–2725. [Google Scholar] [CrossRef]
- Bitanihirwe, B.K.Y.; Woo, T.-U.W. Oxidative Stress in Schizophrenia: An Integrated Approach. Neurosci. Biobehav. Rev. 2011, 35, 878–893. [Google Scholar] [CrossRef] [Green Version]
- Jarskog, L.F.; Glantz, L.A.; Gilmore, J.H.; Lieberman, J.A. Apoptotic Mechanisms in the Pathophysiology of Schizophrenia. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2005, 29, 846–858. [Google Scholar] [CrossRef]
- Missonnier, P.; Hasler, R.; Perroud, N.; Herrmann, F.R.; Millet, P.; Richiardi, J.; Malafosse, A.; Giannakopoulos, P.; Baud, P. EEG Anomalies in Adult ADHD Subjects Performing a Working Memory Task. Neuroscience 2013, 241, 135–146. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emran, T.B.; Islam, F.; Nath, N.; Sutradhar, H.; Das, R.; Mitra, S.; Alshahrani, M.M.; Alhasaniah, A.H.; Sharma, R. Naringin and Naringenin Polyphenols in Neurological Diseases: Understandings from a Therapeutic Viewpoint. Life 2023, 13, 99. https://doi.org/10.3390/life13010099
Emran TB, Islam F, Nath N, Sutradhar H, Das R, Mitra S, Alshahrani MM, Alhasaniah AH, Sharma R. Naringin and Naringenin Polyphenols in Neurological Diseases: Understandings from a Therapeutic Viewpoint. Life. 2023; 13(1):99. https://doi.org/10.3390/life13010099
Chicago/Turabian StyleEmran, Talha Bin, Fahadul Islam, Nikhil Nath, Hriday Sutradhar, Rajib Das, Saikat Mitra, Mohammed Merae Alshahrani, Abdulaziz Hassan Alhasaniah, and Rohit Sharma. 2023. "Naringin and Naringenin Polyphenols in Neurological Diseases: Understandings from a Therapeutic Viewpoint" Life 13, no. 1: 99. https://doi.org/10.3390/life13010099