
An Anatomically Preserved Cone-like Flower from the Lower Cretaceous of China


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
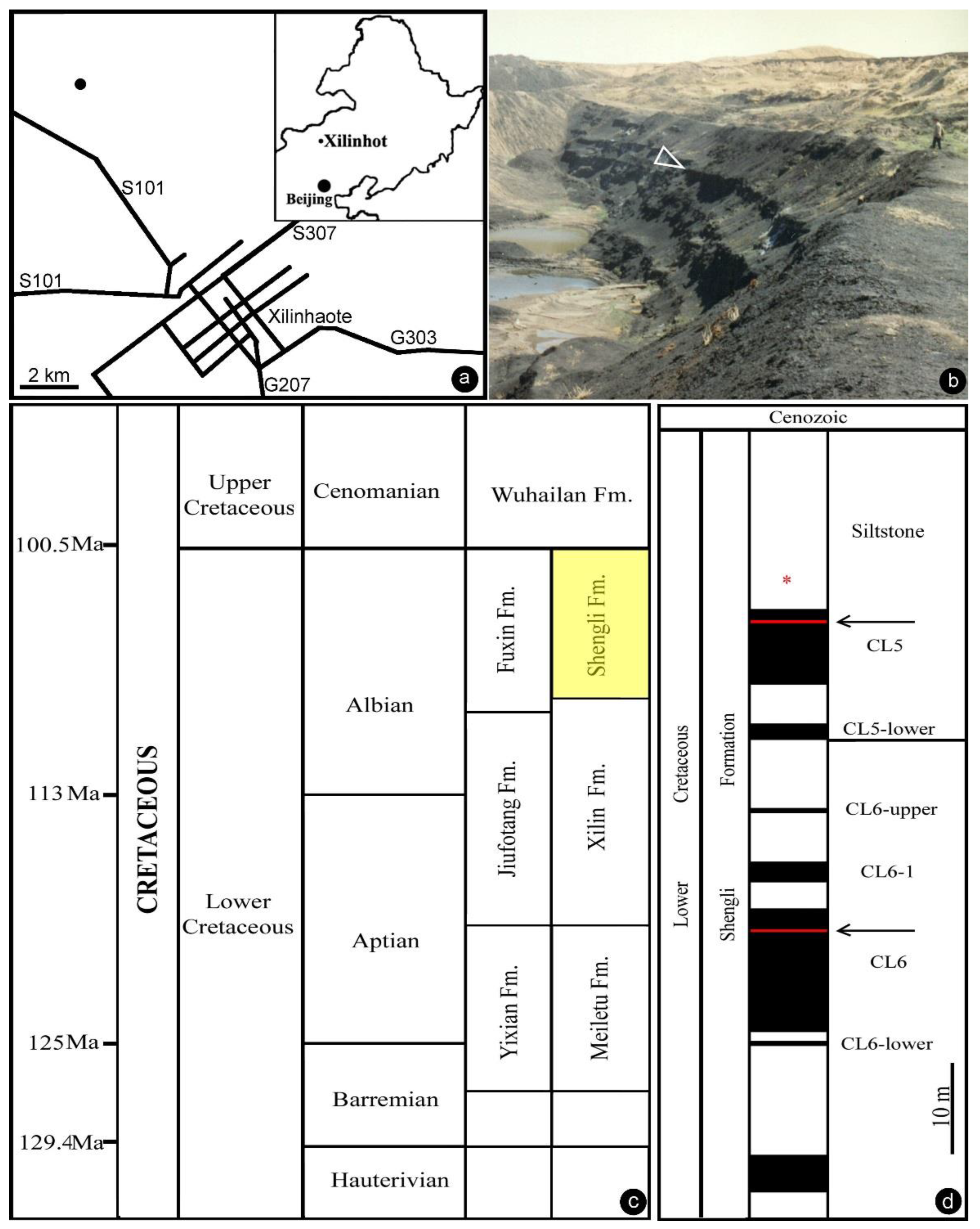
2. Materials and Methods
3. Results
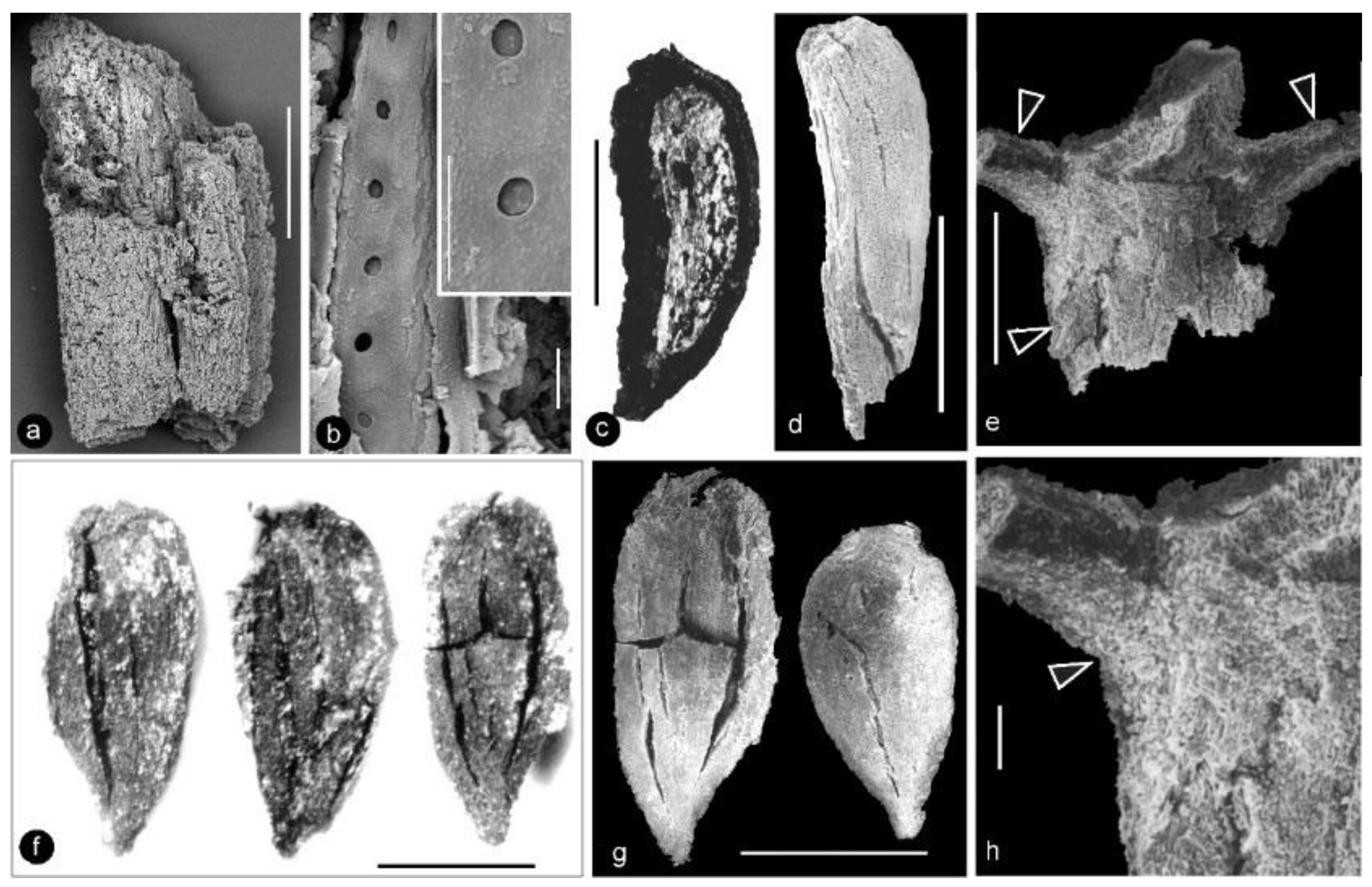
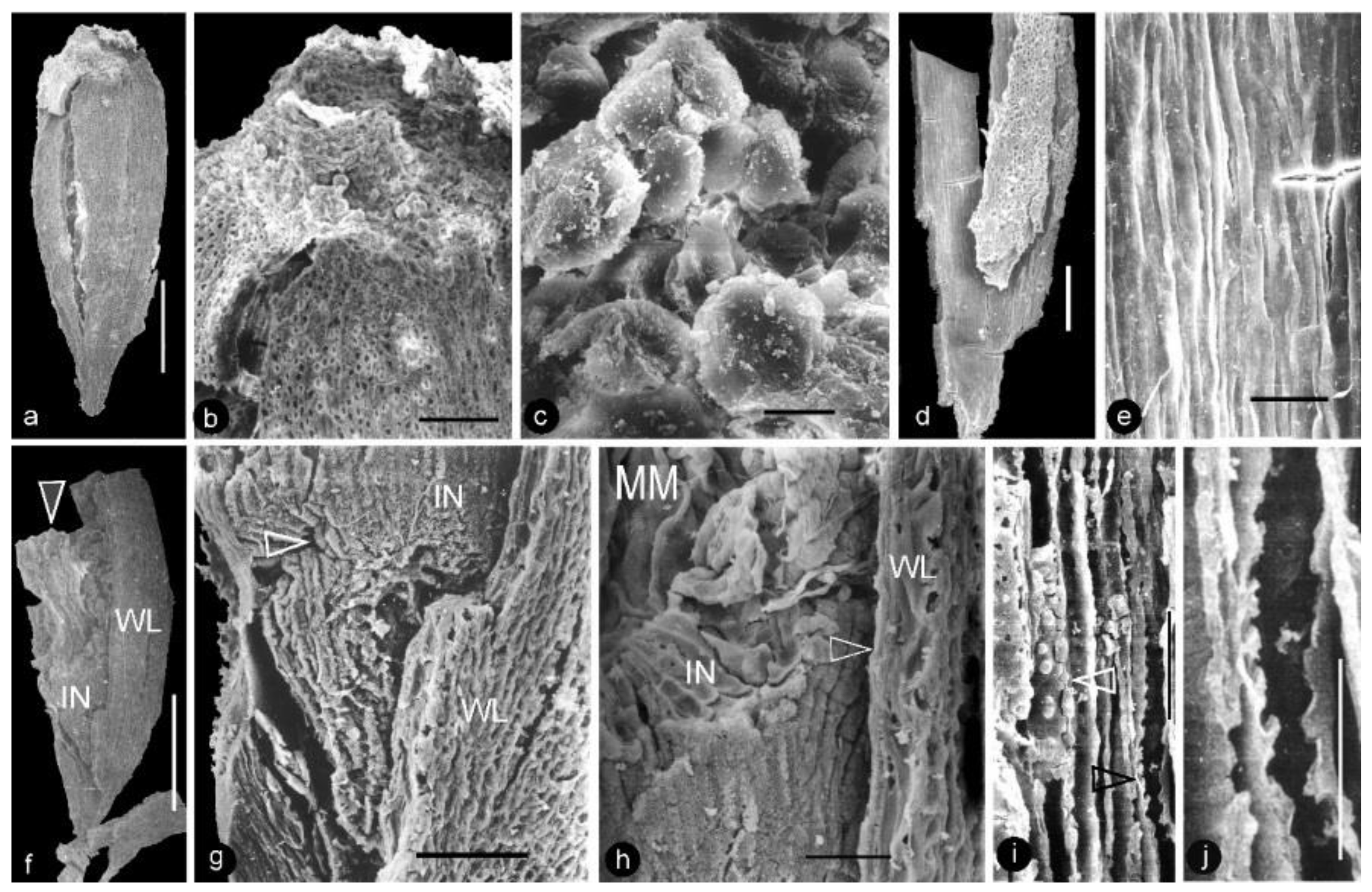
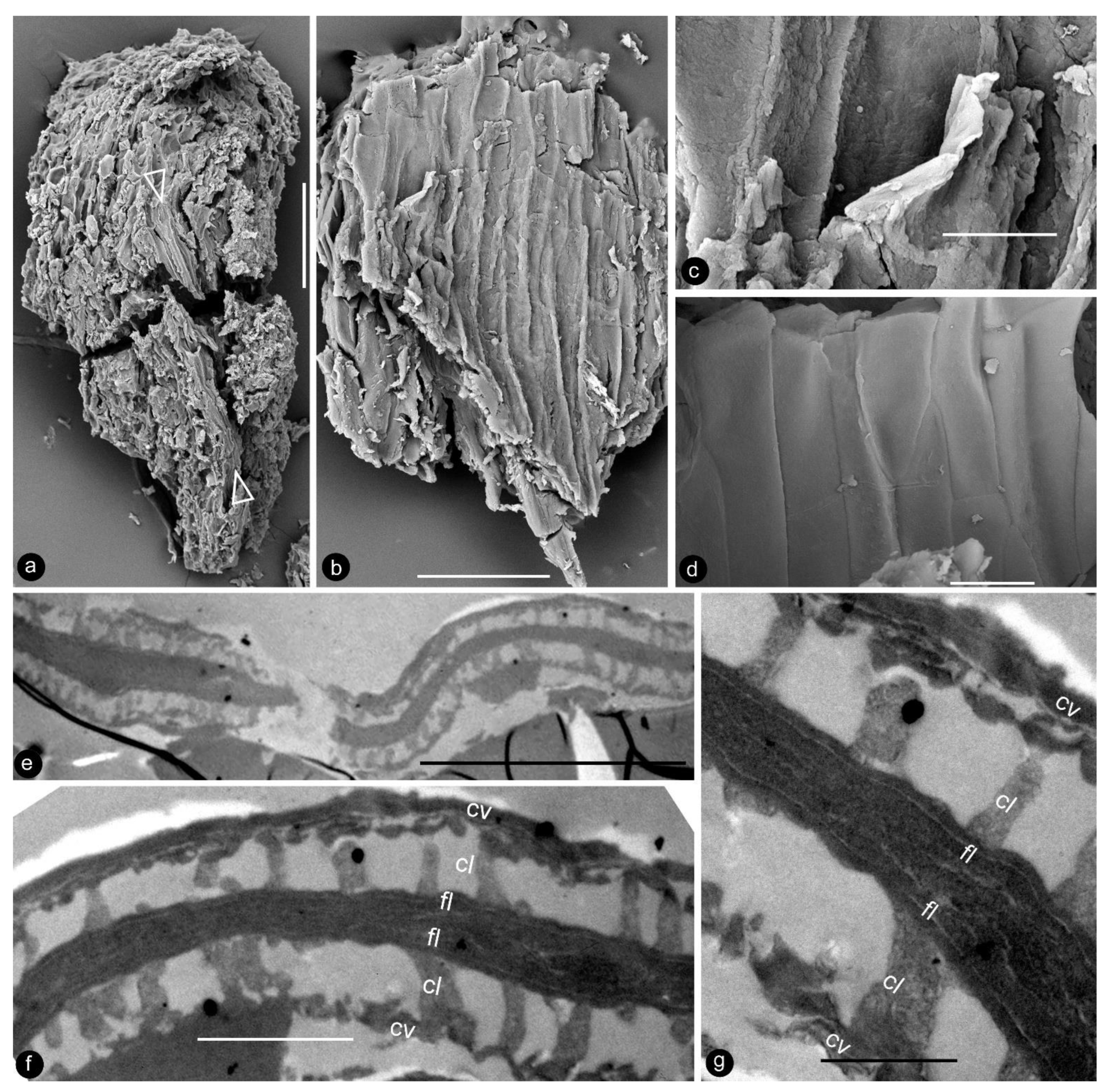
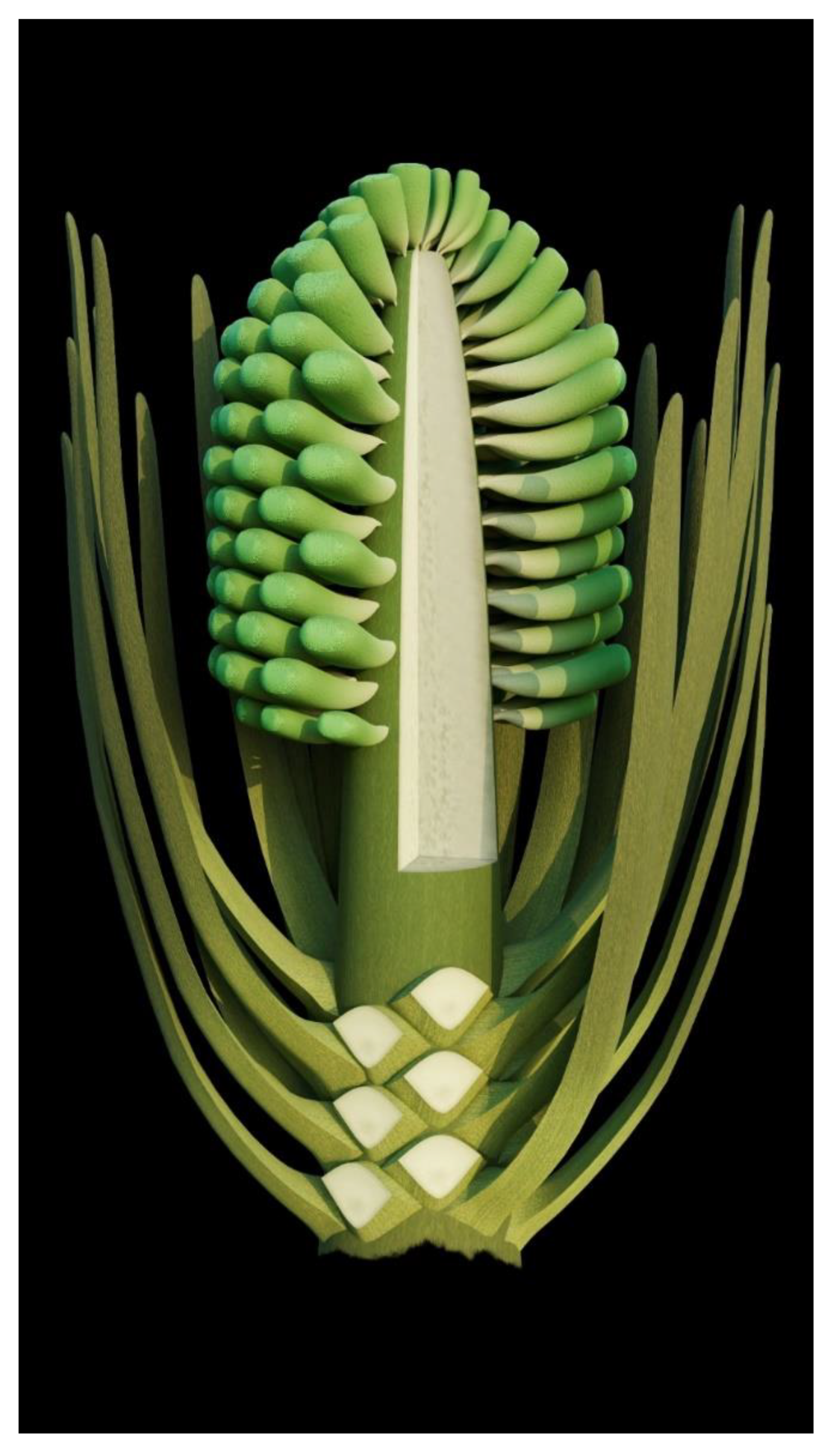
- Generic diagnosis: Female organ cone-like, pedicellate, with helically arranged carpels and perianth. Carpel composed of an ovary wall and an anatropous ovule. Ovular membrane sac-like with an extra layer in the distal portion, three-layered.
- Type species: Xilinia shengliensis gen. et sp. nov.
- Etymology: Xilin, for the city of Xilinhot where the fossil was collected.
- Species diagnosis: (In addition to that of the genus), organ up to 23 mm long and 14 mm wide. Pedicel up to 3.5 mm in diameter. Perianth element linear, narrow, up to 22 mm long. Gynoecium oval, about 1 cm in diameter, with multiple carpels. Carpel inverted triangular, up to 2.3 mm long and 1.1 mm wide, three-dimensionally preserved, papillate apically. Ovule single in a carpel, anatropous, with sac-like ovular membrane of amber color.
- Etymology: shengli, for the name of the formation from which the fossil was collected.
- Holotype specimen: 9222.
- Depository: Institute of Botany, Chinese Academy of Sciences, Beijing, China.
- Age and horizon: the Albian, Early Cretaceous; the Shengli Formation.
- Locality: Xilinhot, Inner Mongolia, China.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arber, E.A.N.; Parkin, J. On the origin of angiosperms. J. Linn. Soc. Lond. Bot. 1907, 38, 29–80. [Google Scholar] [CrossRef] [Green Version]
- Wang, X. The Dawn Angiosperms, 2nd ed.; Springer: Cham, Switzerland, 2018; p. 407. [Google Scholar]
- Duan, S. The oldest angiosperm—A tricarpous female reproductive fossil from western Liaoning Province, NE China. Sci. China D 1998, 41, 14–20. [Google Scholar] [CrossRef]
- Sun, G.; Dilcher, D.L.; Zheng, S.; Zhou, Z. In search of the first flower: A Jurassic angiosperm, Archaefructus, from Northeast China. Science 1998, 282, 1692–1695. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Ji, Q.; Dilcher, D.L.; Zheng, S.; Nixon, K.C.; Wang, X. Archaefructaceae, a new basal angiosperm family. Science 2002, 296, 899–904. [Google Scholar] [CrossRef] [Green Version]
- Ji, Q.; Li, H.; Bowe, M.; Liu, Y.; Taylor, D.W. Early Cretaceous Archaefructus eoflora sp. nov. with bisexual flowers from Beipiao, Western Liaoning, China. Acta Geol. Sin. Engl. Ed. 2004, 78, 883–896. [Google Scholar]
- Wang, X.; Zheng, X.-T. Reconsiderations on two characters of early angiosperm Archaefructus. Palaeoworld 2012, 21, 193–201. [Google Scholar] [CrossRef]
- Leng, Q.; Friis, E.M. Sinocarpus decussatus gen. et sp. nov., a new angiosperm with basally syncarpous fruits from the Yixian Formation of Northeast China. Plant Syst. Evol. 2003, 241, 77–88. [Google Scholar] [CrossRef]
- Leng, Q.; Friis, E.M. Angiosperm leaves associated with Sinocarpus infructescences from the Yixian Formation (Mid-Early Cretaceous) of NE China. Plant Syst. Evol. 2006, 262, 173–187. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, S. The earliest normal flower from Liaoning Province, China. J. Integr. Plant Biol. 2009, 51, 800–811. [Google Scholar] [CrossRef]
- Wang, X.; Shih, C.; Liu, Z.-J.; Lin, L.; Singh, K.J. Reconstructing the Callianthus plant—An early aquatic angiosperm from the Lower Cretaceous of China. Cretac. Res. 2021, 128, 104983. [Google Scholar] [CrossRef]
- Wang, X.; Han, G. The earliest ascidiate carpel and its implications for angiosperm evolution. Acta Geol. Sin. Engl. Ed. 2011, 85, 998–1002. [Google Scholar] [CrossRef]
- Liu, Z.-J.; Chen, L.-J.; Wang, X. A whole-plant monocot from Lower Cretaceous. Palaeoworld 2021, 30, 169–175. [Google Scholar] [CrossRef]
- Liu, X.; Ma, L.; Liu, B.; Liu, Z.-J.; Wang, X. A novel angiosperm including various parts from the Early Cretaceous sheds new light on flower evolution. Hist. Biol. 2021, 33, 2706–2714. [Google Scholar] [CrossRef]
- Retallack, G.; Dilcher, D.L. Early angiosperm reproduction: Prisca reynoldsii, gen. et sp. nov. from the mid-Cretaceous coastal deposits in Kansas, U.S.A. Paläontograph. B 1981, 179, 103–107. [Google Scholar]
- Crane, P.R.; Dilcher, D.L. Lesqueria: An early angiosperm fruiting axis from the Mid-Cretaceous. Ann. Mo. Bot. Gard. 1984, 71, 384–402. [Google Scholar] [CrossRef]
- Dilcher, D.L.; Crane, P.R. Archaeanthus: An early angiosperm from the Cenomanian of the Western Interior of North America. Ann. Mo. Bot. Gard. 1984, 71, 351–383. [Google Scholar] [CrossRef]
- Fu, Q.; Diez, J.B.; Pole, M.; Garcia-Avila, M.; Liu, Z.-J.; Chu, H.; Hou, Y.; Yin, P.; Zhang, G.-Q.; Du, K.; et al. An unexpected noncarpellate epigynous flower from the Jurassic of China. eLife 2018, 7, e38827. [Google Scholar] [CrossRef]
- Friis, E.M.; Crane, P.R.; Pedersen, K.R. The Early Flowers and Angiosperm Evolution; Cambridge University Press: Cambridge, UK, 2011; p. 596. [Google Scholar]
- Herendeen, P.S.; Friis, E.M.; Pedersen, K.R.; Crane, P.R. Palaeobotanical redux: Revisiting the age of the angiosperms. Nat. Plants 2017, 3, 17015. [Google Scholar] [CrossRef] [Green Version]
- Sokoloff, D.D.; Remizowa, M.V.; El, E.S.; Rudall, P.J.; Bateman, R.M. Supposed Jurassic angiosperms lack pentamery, an important angiosperm-specific feature. New Phytol. 2020, 228, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Coiro, M.; Doyle, J.A.; Hilton, J. How deep is the conflict between molecular and fossil evidence on the age of angiosperms? New Phytol. 2019, 223, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Bateman, R.M. Hunting the snark: The flawed search for mythical Jurassic angiosperms. J. Exp. Bot. 2020, 71, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Diez, J.B.; Pole, M.; García-Ávila, M.; Wang, X. Nanjinganthus is an angiosperm, isn’t it? China Geol. 2020, 3, 359–361. [Google Scholar] [CrossRef]
- Song, Z.-C.; Liu, G.-W.; Jia, B.-L.; Hua, R.-H. Early Cretaceous palynological assemblages from Eren Basin, Nei Mongol. In Cretaceous Ostracod and Sporo-Pollen Fossils of Eren Basin, Nei Mongol; Nanjing Institute of Geology and Palaeontology, The First Exploration Company, Ministry of Oil Industry, Eds.; Anhui Science and Technology Publishing House: Hefei, China, 1986; Volume 2, pp. 105–335. [Google Scholar]
- Shen, H.X.; Zhang, C.H.; Wu, Y.F.; Lü, M.; Wang, X. The first radiometric age of the Shengli Formation (Lower Cretaceous) of Inner Mongolia, China. J. Stratigr. 2021, 45, 204–209. [Google Scholar] [CrossRef]
- Song, Z.-C. A review on the study of Early Cretaceous angiosperm pollen in China. Acta Micropalaeontol. Sin. 1986, 3, 373–386. [Google Scholar]
- Sha, J. Current research on Cretaceous lake systems in northeast China. Cretac. Res. 2007, 28, 143–145. [Google Scholar] [CrossRef]
- Gou, Y.-X.; Wang, Z.-Z.; Yang, J.-D.; Wang, W.-S. Cretaceous Ostracoda and Pollen/Spores from Eren Basin of Nei Mongol along with Sedimentary Environments; Anhui Science and Technology Press: Hefei, China, 1986; Volume 2, p. 104. [Google Scholar]
- Sha, J. Cretaceous stratigraphy of northeast China: Non-marine and marine correlation. Cretac. Res. 2007, 28, 146–170. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Company: New York, NY, USA, 1940; p. 523. [Google Scholar]
- Jensen, W.A. Botanical Histochemistry, Principles and Practice; W. H. Freeman and Company: San Francisco, CA, USA, 1962; p. 408. [Google Scholar]
- Tomlinson, P.B.; Takaso, T. Seed cone structure in conifers in relation to development and pollination: A biological approach. Can. J. Bot. 2002, 80, 1250–1273. [Google Scholar] [CrossRef]
- Crane, P.R. Phylogenetic analysis of seed plants and the origin of angiosperms. Ann. Mo. Bot. Gard. 1985, 72, 716–793. [Google Scholar] [CrossRef]
- Hemsley, A.R. The ultrastructure of the exine of the megaspores in two Palaeozoic seed-like structures. Rev. Palaeobot. Palynol. 1990, 63, 137–152. [Google Scholar] [CrossRef]
- Kovach, W.L.; Dilcher, D.L. Megaspores and other dispersed plant remains from the Dakota Formation (Cenomanian) of Kansas, USA. Palynology 1988, 12, 89–119. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, B.; Wang, Y.; Guignard, G. A new Karkenia (Ginkgoales) from the Jurassic Yima Formation, Henan, China and its megaspore membrane ultrastructure. Rev. Palaeobot. Palynol. 2002, 120, 91–105. [Google Scholar] [CrossRef]
- Taylor, T.N.; Brauer, D.F. Ultrastructural studies of in situ Devonian spores: Barinophyton citrulliforme . Am. J. Bot. 1983, 70, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Taylor, T.N. Scanning electron microscopy of fossil megaspores: Wall development. In Proceedings of the 7th Annual Scanning Electron Microscope Symposium, Chicago, IL, USA, 8–9 April 1974; pp. 359–366. [Google Scholar]
- Zhou, Z. Comparative ultrastructure of fossil and living ginkgoacean megaspore membranes. Rev. Palaeobot. Palynol. 1993, 78, 167–182. [Google Scholar]
- Zhou, Z. Stalagma samara, a new podocarpaceous conifer with monocolpate pollen from the Upper Triassic of Hunan, China. Paläontograph. B 1983, 185, 56–78. [Google Scholar]
- Batten, D.J.; Collinson, M.E.; Brain, A.P.R. Megaspores and microspores of the extant and Paleogene marsileaceous fern Regnellidium and Cretaceous Molaspora: Evolutionary and phytogeographic implications. Int. J. Plant Sci. 2011, 172, 1087–1100. [Google Scholar] [CrossRef]
- Archangelsky, S.; Cuneo, R. Ferugliocladaceae, a new conifer family from the Permian of Gondwana. Rev. Palaeobot. Palynol. 1987, 51, 3–30. [Google Scholar] [CrossRef]
- Lupia, R. Megaspores and palynomorphs from the lower Potomac Group of Maryland, U.S.A. Int. J. Plant Sci. 2004, 165, 651–670. [Google Scholar] [CrossRef]
- Krings, M.; Grewing, A.; Taylor, T.N.; Kerp, H.; Galtier, J. Lageniastrum macrosporae (fossil Volvocales, Lagniastraceae nov. fam.), an endophyte in megaspores from the Carboniferous of the French Massif Central. Geobios 2005, 38, 451–465. [Google Scholar] [CrossRef]
- Zhou, Z.; Zheng, S.; Zhang, L. Morphology and age of Yimaia (Ginkgoales) from Daohugou Village, Ningcheng, Inner Mongolia, China. Cretac. Res. 2007, 28, 348–362. [Google Scholar] [CrossRef]
- Cúneo, N.R.; Hermsen, E.J.; Gandolfo, M.A. Regnellidium (Salviniales, Marsileaceae) macrofossils and associated spores from the Late Cretaceous of South America. Int. J. Plant Sci. 2013, 174, 340–349. [Google Scholar] [CrossRef]
- Wilson, A.T.; Gullickson, K.A. Ultrastructure of transitional dispersed megaspores from the Middle Devonian of New York. Int. J. Plant Sci. 2013, 174, 309–316. [Google Scholar] [CrossRef]
- Pettitt, J.M. Pteridophytic features in some Lower Carboniferous seed megaspores. Bot. J. Linn. Soc. 1969, 62, 233–239. [Google Scholar] [CrossRef]
- Taylor, W.A. Comparative analysis of megaspore ultrastructure in Pennsylvanian lycophytes. Rev. Palaeobot. Palynol. 1990, 62, 65–78. [Google Scholar] [CrossRef]
- Taylor, T.N.; Brack-Hanes, S.D. Achlamydocarpon varius comb. nov.: Morphology and reproductive biology. Am. J. Bot. 1976, 63, 1257–1265. [Google Scholar] [CrossRef]
- Zimmermann, R.P.; Taylor, T.N. The ultrastructure of Paleozoic megaspore membranes. Pollen Spores 1970, 12, 451–468. [Google Scholar]
- Grauvogel-Stamm, L.; Lugardon, B. The spores of the Triassic lycopsid Pleuromeia sternbergii (Muenster) Corda: Morphology, ultrastructure, phyologenetic implications, and chronostratigraphic inferences. Int. J. Plant Sci. 2004, 165, 631–650. [Google Scholar] [CrossRef]
- Del Fueyo, G.M.; Archangelsky, S.; Llorens, M.; Cuneo, R. Coniferous ovulate cones from the Lower Cretaceous of Santa Cruz Province, Argentina. Int. J. Plant Sci. 2008, 169, 799–813. [Google Scholar] [CrossRef]
- Liu, W.-Z.; Hilu, K.; Wang, Y.-L. From leaf and branch into a flower: Magnolia tells the story. Bot. Stud. 2014, 55, 28. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, C.J. Gymnosperms, Structure and Evolution; Johnson Reprint Corporation: New York, NY, USA, 1957; p. 484. [Google Scholar]
- Bierhorst, D.W. Morphology of Vascular Plants; Macmillan Company: New York, NY, USA, 1971; p. 560. [Google Scholar]
- Biswas, C.; Johri, B.M. The Gymnosperms; Springer: Berlin/Heidelberg, Germany, 1997; p. 494. [Google Scholar]
- Cronquist, A. The Evolution and Classification of Flowering Plants, 2nd ed.; New York Botanical Garden: New York, NY, USA, 1988; p. 555. [Google Scholar]
- Han, G.; Wang, X. A new infructescence of angiosperms from the Early Cretaceous of China. Acta Geol. Sin. Engl. Ed. 2020, 94, 1711–1713. [Google Scholar] [CrossRef]
- Cui, D.-F.; Hou, Y.; Yin, P.; Wang, X. A Jurassic flower bud from China. Geol. Soc. Lond. Spec. Publ. 2022, 521, 81–93. [Google Scholar] [CrossRef]
- Chen, L.-J.; Wang, X. A flower bud from the Lower Cretaceous of China. Biology 2022, 11, 1598. [Google Scholar] [CrossRef] [PubMed]
- Farjon, A. A Monograph of Cupressaceae and Sciadopitys; Royal Botanic Gardens: Richmond, UK, 2005; p. 643. [Google Scholar]
- Eckenwalder, J.E. Conifers of the World, the Complete Reference; Timber Press: Portland, OR, USA, 2009; p. 720. [Google Scholar]
- Martens, P. Les Gnetophytes; Gebrueder Borntraeger: Berlin, Germany, 1971; p. 295. [Google Scholar]
- Li, H.; Taylor, D.W. Vessel-bearing stems of Vasovinea tianii gen. et sp. nov. (Gigantopteridales) from the upper Permian of Guizhou Province, China. Am. J. Bot. 1999, 86, 1563–1575. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Taylor, E.L.; Taylor, T.N. Permian vessel elements. Science 1996, 271, 188–189. [Google Scholar] [CrossRef]
- Meeuse, A.D.J. From ovule to ovary: A contribution to the phylogeny of the megasporangium. Acta Biotheor. 1963, 16, 127–182. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Diez, J.B.; Pole, M.; García-Ávila, M. An Anatomically Preserved Cone-like Flower from the Lower Cretaceous of China. Life 2023, 13, 129. https://doi.org/10.3390/life13010129
Wang X, Diez JB, Pole M, García-Ávila M. An Anatomically Preserved Cone-like Flower from the Lower Cretaceous of China. Life. 2023; 13(1):129. https://doi.org/10.3390/life13010129
Chicago/Turabian StyleWang, Xin, José B. Diez, Mike Pole, and Manuel García-Ávila. 2023. "An Anatomically Preserved Cone-like Flower from the Lower Cretaceous of China" Life 13, no. 1: 129. https://doi.org/10.3390/life13010129