
Inhibitory Effect of Bacillus licheniformis Strains Isolated from Canine Oral Cavity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Bacterial Strains
2.2. Preparation of Neutralized Cell-Free Supernatants
2.3. Antimicrobial Effect of B. licheniformis Strains
2.4. Bacitracin Susceptibility Test of Oral Pathogens
2.5. Evaluation of the Ability to Produce Bioactive Substances
2.6. Effect of Enzymes on Antimicrobial Activity
2.7. Statistical Analyses
3. Results
3.1. Identification of Bacterial Strains
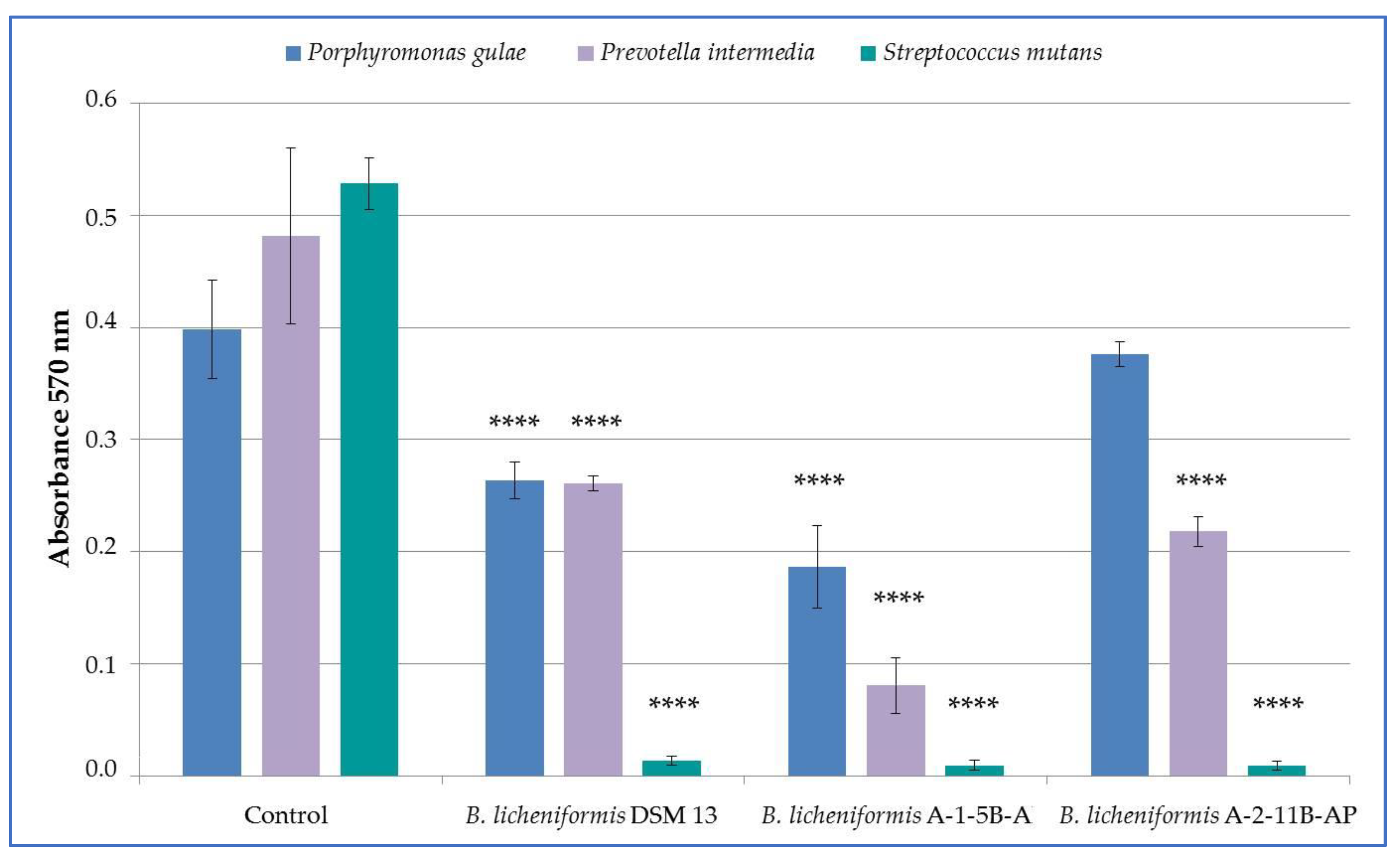
3.2. Antimicrobial Effect of B. licheniformis Strains
3.3. Bacitracin Susceptibility Test
3.4. Production of Bioactive Substances
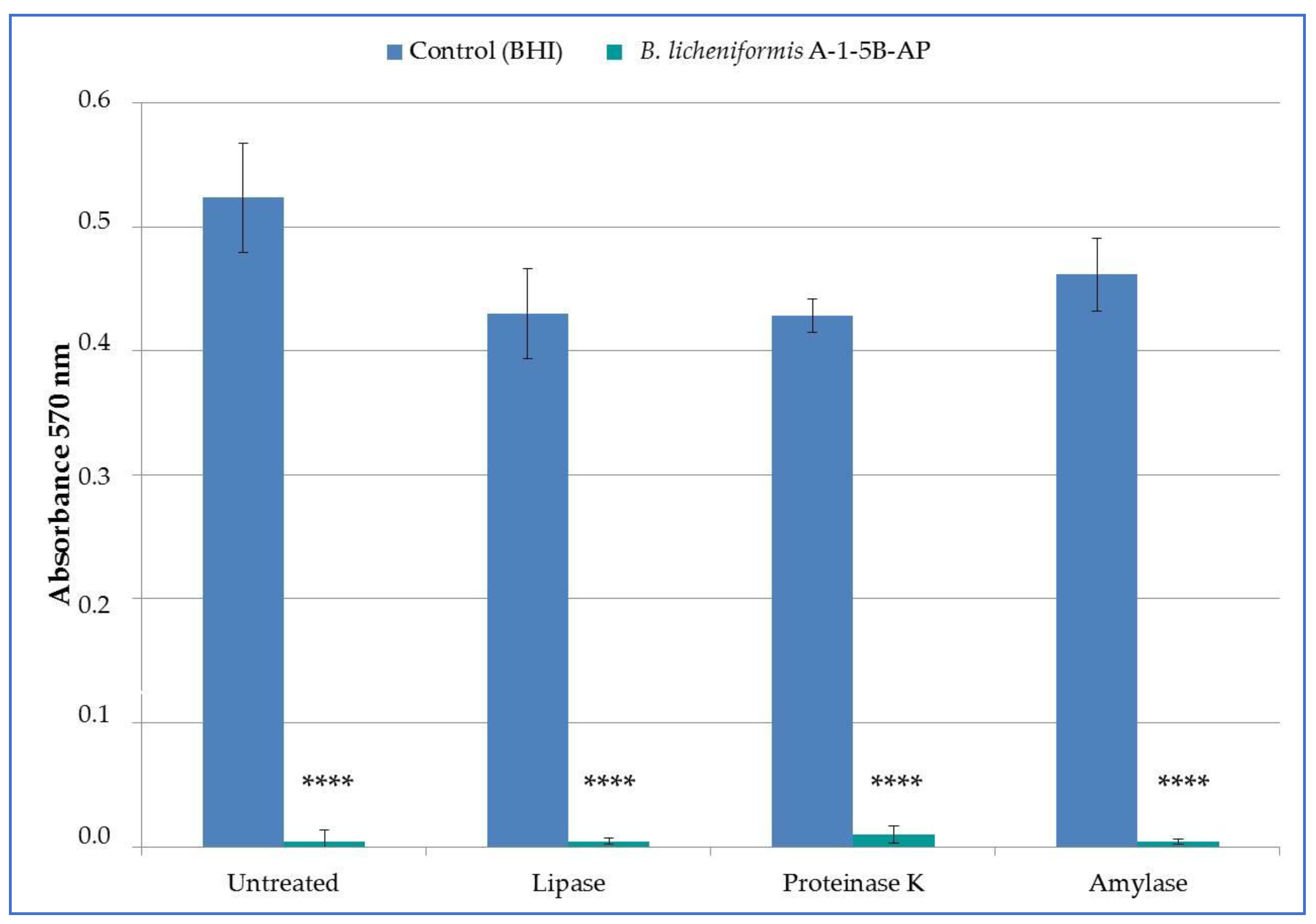
3.5. Effect of Enzymes on Antimicrobial Activity of nCFS B. licheniformis A-1-5B-AP
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zaura, E.; Twetman, S. Critical Appraisal of Oral Pre- and Probiotics for Caries Prevention and Care. Caries Res. 2019, 53, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Zupancic, K.; Kriksic, V.; Kovacevic, I.; Kovacevic, D. Influence of Oral Probiotic Streptococcus salivarius K12 on Ear and Oral Cavity Health in Humans: Systematic Review. Probiotics Antimicrob. Proteins 2017, 9, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Barzegari, A.; Kheyrolahzadeh, K.; Hosseiniyan Khatibi, S.M.; Sharifi, S.; Memar, M.Y.; Zununi Vahed, S. The Battle of Probiotics and Their Derivatives against Biofilms. Infect. Drug Resist. 2020, 13, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Kim, W.S.; Paik, H.D. Bacillus strains as human probiotics: Characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef]
- Romo-Barrera, C.M.; Castrillón-Rivera, L.E.; Palma-Ramos, A.; Castañeda-Sánchez, J.I.; Luna-Herrera, J. Bacillus licheniformis and Bacillus subtilis, Probiotics That Induce the Formation of Macrophage Extracellular Traps. Microorganisms 2021, 9, 2027. [Google Scholar] [CrossRef]
- Busarcevic, M.; Dalgalarrondo, M. Purification and genetic characterisation of the novel bacteriocin LS2 produced by the human oral strain Lactobacillus salivarius BGHO1. Int. J. Antimicrob. Agents 2012, 40, 127–134. [Google Scholar] [CrossRef]
- Yeak, K.Y.C.; Perko, M.; Staring, G.; Fernandez-Ciruelos, B.M.; Wells, J.M.; Abee, T.; Wells-Bennik, M. Lichenysin Production by Bacillus licheniformis Food Isolates and Toxicity to Human Cells. Front. Microbiol. 2022, 13, 831033. [Google Scholar] [CrossRef]
- Lee, C.; Kim, J.Y.; Song, H.S.; Kim, Y.B.; Choi, Y.E.; Yoon, C.; Nam, Y.D.; Roh, S.W. Genomic Analysis of Bacillus licheniformis CBA7126 Isolated from a Human Fecal Sample. Front. Pharmacol. 2017, 8, 724. [Google Scholar] [CrossRef]
- Nithya, V.; Muthukumar, S.P.; Halami, P.M. Safety assessment of Bacillus licheniformis Me1 isolated from milk for probiotic application. Int. J. Toxicol. 2012, 31, 228–237. [Google Scholar] [CrossRef]
- Rey, M.W.; Ramaiya, P.; Nelson, B.A.; Brody-Karpin, S.D.; Zaretsky, E.J.; Tang, M.; Lopez de Leon, A.; Xiang, H.; Gusti, V.; Clausen, I.G.; et al. Complete genome sequence of the industrial bacterium Bacillus licheniformis and comparisons with closely related Bacillus species. Genome Biol. 2004, 5, R77. [Google Scholar] [CrossRef]
- Rehman, H.U.; Siddique, N.N.; Aman, A.; Nawaz, M.A.; Baloch, A.H.; Qader, S.A.U. Morphological and molecular based identification of pectinase producing Bacillus licheniformis from rotten vegetable. J. Genet. Eng. Biotechnol. 2015, 13, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Veith, B.; Herzberg, C.; Steckel, S.; Feesche, J.; Maurer, K.H.; Ehrenreich, P.; Bäumer, S.; Henne, A.; Liesegang, H.; Merkl, R.; et al. The complete genome sequence of Bacillus licheniformis DSM13, an organism with great industrial potential. J. Mol. Microbiol. Biotechnol. 2004, 7, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Muras, A.; Romero, M.; Mayer, C.; Otero, A. Biotechnological applications of Bacillus licheniformis. Crit. Rev. Biotechnol. 2021, 41, 609–627. [Google Scholar] [CrossRef] [PubMed]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as Potential Probiotics: Status, Concerns, and Future Perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef]
- Kan, L.; Guo, F.; Liu, Y.; Pham, V.H.; Guo, Y.; Wang, Z. Probiotics Bacillus licheniformis Improves Intestinal Health of Subclinical Necrotic Enteritis-Challenged Broilers. Front. Microbiol. 2021, 12, 623739. [Google Scholar] [CrossRef]
- He, L.; Chen, W.; Liu, Y. Production and partial characterization of bacteriocin-like pepitdes by Bacillus licheniformis ZJU12. Microbiol. Res. 2006, 161, 321–326. [Google Scholar] [CrossRef]
- Stoica, R.M.; Moscovici, M.; Tomulescu, C.; Cășărică, A.; Băbeanu, N.; Popa, O.; Kahraman, H.A. Antimicrobial compounds of the genus Bacillus: A review. Rom. Biotechnol. Lett. 2019, 24, 1111–1119. [Google Scholar] [CrossRef]
- Begley, M.; Cotter, P.D.; Hill, C.; Ross, R.P. Identification of a novel two-peptide lantibiotic, lichenicidin, following rational genome mining for LanM proteins. Appl. Environ. Microbiol. 2009, 75, 5451–5460. [Google Scholar] [CrossRef]
- Petrova, P.; Arsov, A.; Ivanov, I.; Tsigoriyna, L.; Petrov, K. New Exopolysaccharides Produced by Bacillus licheniformis 24 Display Substrate-Dependent Content and Antioxidant Activity. Microorganisms 2021, 9, 2127. [Google Scholar] [CrossRef]
- Hertadi, R.; Umriani Permatasari, N.; Ratnaningsih, E. Box-Wilson Design for Optimization of in vitro Levan Production and Levan Application as Antioxidant and Antibacterial Agents. Iran. Biomed. J. 2021, 25, 202–212. [Google Scholar] [CrossRef]
- Permatasari, N.U.; Ratnaningsih, E.; Hertadi, R. Molecular cloning and expression of levansucrase gene from Bacillus licheniformis BK1 isolated from Bledug Kuwu Mud Crater. EurAsian J. Biosci. 2019, 13, 223–230. [Google Scholar]
- Nerurkar, A.S. Structural and molecular characteristics of lichenysin and its relationship with surface activity. In Biosurfactants, 1st ed.; Sen, R., Ed.; Springer: New York, NY, USA, 2010; Volume 1, pp. 304–315. [Google Scholar]
- Roy, A. A Review on the Biosurfactants: Properties, Types and its Applications. J. Fundam. Renew. Energy Appl. 2017, 8, 1000248. [Google Scholar] [CrossRef]
- Jiang, H.; Dong, H.; Zhang, G.; Yu, B.; Chapman, L.R.; Fields, M.W. Microbial diversity in water and sediment of Lake Chaka, an athalassohaline lake in northwestern China. Appl. Environ. Microbiol. 2006, 72, 3832–3845. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Chang, M.T.; Huang, L.; Chu, W.S. Development of a novel PCR assay based on the gyrase B gene for species identification of Bacillus licheniformis. Mol. Cell. Probes 2012, 26, 215–217. [Google Scholar] [CrossRef]
- Huang, Q.; Baum, L.; Fu, W.L. Simple and practical staining of DNA with GelRed in agarose gel electrophoresis. Clin. Lab. 2010, 56, 149–152. [Google Scholar]
- McGinnis, S.; Madden, T.L. BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 2004, 32, W20–W25. [Google Scholar] [CrossRef]
- Sadanand, R.L.N.; Raghunath, P. Prevalence and antibiotic susceptibility of Streptococcus pyogenes isolated from pyoderma in a tertiary care hospital, Hyderabad, South India. Indian J. Microbiol. Res. 2021, 8, 274–278. [Google Scholar] [CrossRef]
- Morikawa, M.; Hirata, Y.; Imanaka, T. A study on the structure-function relationship of lipopeptide biosurfactants. Biochim. Biophys. Acta. 2000, 1488, 211–218. [Google Scholar] [CrossRef]
- Nakapong, S.; Pichyangkura, R.; Ito, K.; Iizuka, M.; Pongsawasdi, P. High expression level of levansucrase from Bacillus licheniformis RN-01 and synthesis of levan nanoparticles. Int. J. Biol. Macromol. 2013, 54, 30–36. [Google Scholar] [CrossRef]
- Prieto, M.L.; O’Sullivan, L.; Tan, S.P.; McLoughlin, P.; Hughes, H.; O’Connor, P.M.; Cotter, P.D.; Lawlor, P.G.; Gardiner, G.E. Assessment of the bacteriocinogenic potential of marine bacteria reveals lichenicidin production by seaweed-derived Bacillus spp. Mar. Drugs 2012, 10, 2280–2299. [Google Scholar] [CrossRef]
- Bolocan, A.S.; Pennone, V.; O’Connor, P.M.; Coffey, A.; Nicolau, A.I.; McAuliffe, O.; Jordan, K. Inhibition of Listeria monocytogenes biofilms by bacteriocin-producing bacteria isolated from mushroom substrate. J. Appl. Microbiol. 2017, 122, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, T.; Rintaluoma, N.; Andersson, M.; Taimisto, A.M.; Ali-Vehmas, T.; Seppälä, A.; Priha, O.; Salkinoja-Salonen, M. Toxinogenic Bacillus pumilus and Bacillus licheniformis from mastitic milk. Vet. Microbiol. 2007, 124, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.M.; Kim, J.S.; Kim, H.S.; Kim, Y.Y.; Oh, J.K.; Jung, H.W.; Park, D.S.; Bae, K.H. Lactobacillus reuteri AN417 cell-free culture supernatant as a novel antibacterial agent targeting oral pathogenic bacteria. Sci. Rep. 2021, 11, 1631. [Google Scholar] [CrossRef] [PubMed]
- Özavci, V.; Erbas, G.; Parin, U.; Yüksel, H.T.; Kirkan, Ş. Molecular detection of feline and canine periodontal pathogens. Vet. Anim. Sci. 2019, 8, 100069. [Google Scholar] [CrossRef]
- Lin, Y.; Zhou, X.; Li, Y. Strategies for Streptococcus mutans biofilm dispersal through extracellular polymeric substances disruption. Mol. Oral Microbiol. 2022, 37, 1–8. [Google Scholar] [CrossRef]
- Messora, M.R.; Pereira, L.J.; Foureaux, R.; Oliveira, L.F.; Sordi, C.G.; Alves, A.J.; Napimoga, M.H.; Nagata, M.J.; Ervolino, E.; Furlaneto, F.A. Favourable effects of Bacillus subtilis and Bacillus licheniformis on experimental periodontitis in rats. Arch. Oral Biol. 2016, 66, 108–119. [Google Scholar] [CrossRef]
- Alkaya, B.; Laleman, I.; Keceli, S.; Ozcelik, O.; Cenk Haytac, M.; Teughels, W. Clinical effects of probiotics containing Bacillus species on gingivitis: A pilot randomized controlled trial. J. Periodontal Res. 2017, 52, 497–504. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, L.; Yue, L.; Ling, J.; Fan, M.; Yang, D.; Huang, Z.; Niu, Y.; Liu, J.; Zhao, J.; et al. Expert consensus on dental caries management. Int. J. Oral Sci. 2022, 14, 17. [Google Scholar] [CrossRef]
- Ratna Sudha, M.; Neelamraju, J.; Surendra Reddy, M.; Kumar, M. Evaluation of the Effect of Probiotic Bacillus coagulans Unique IS2 on Mutans Streptococci and Lactobacilli Levels in Saliva and Plaque: A Double-Blind, Randomized, Placebo-Controlled Study in Children. Int. J. Dent. 2020, 2020, 8891708. [Google Scholar] [CrossRef]
- Rivis, O.; Kryvtsova, M.; Nikolajchuk, V. Antagonistic activity of Bacillus probiotics against bacteria isolates of oral cavity of patients with periodontitis. Vìsn. Dnìpropetr. Unìv. Ser. Bìol. Med. 2013, 4, 10–13. [Google Scholar]
- Martins, K.S.; Magalhães, L.T.A.; de Almeida, J.G.; Pieri, F.A. Antagonism of Bacteria from Dog Dental Plaque against Human Cariogenic Bacteria. BioMed Res. Int. 2018, 2018, 2780948. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Tan, Z.; Wang, H.; Liu, W.; Miao, W. Antimicrobial effect of Bacillus licheniformis HN-5 bacitracin A on rice pathogen Pantoea ananatis. BioControl 2021, 66, 249–257. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, Q.; Xiao, T.; Zhu, Y.; Xiao, Y. Impact of Polymyxin Resistance on Virulence and Fitness among Clinically Important Gram-Negative Bacteria. Engineering 2022, 13, 178–185. [Google Scholar] [CrossRef]
- Tran, C.; Cock, I.E.; Chen, X.; Feng, Y. Antimicrobial Bacillus: Metabolites and Their Mode of Action. Antibiotics 2022, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Anumala, D.; Pasupuleti, M.K.; Nagireddy, R.R. Detection and Characterization of Prevotella Intermedia and Its In Vitro Susceptibility to Selected Antimicrobial Agents in Chronic Periodontitis and Acute Myocardial Infarction. Perio J. 2019, 3, 1–6. [Google Scholar] [CrossRef]
- Ahire, J.J.; Kashikar, M.S.; Lakshmi, S.G.; Madempudi, R. Identification and characterization of antimicrobial peptide produced by indigenously isolated Bacillus paralicheniformis UBBLi30 strain. 3 Biotech 2020, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, S.; Heidari, B.; Rassa, M. Antibacterial and hemolytic effects of aqueous and organic extracts from different tissues of sea urchin Echinometra mathaei on pathogenic streptococci. Int. Aquat. Res. 2016, 8, 299–308. [Google Scholar] [CrossRef]
- Nagasawa, R.; Sato, T.; Nomura, N.; Nakamura, T.; Senpuku, H. Potential Risk of Spreading Resistance Genes within Extracellular-DNA-Dependent Biofilms of Streptococcus mutans in Response to Cell Envelope Stress Induced by Sub-MICs of Bacitracin. Appl. Environ. Microbiol. 2020, 86, e00770-20. [Google Scholar] [CrossRef]
- Zaidi, S.; Bhardwaj, T.; Somvanshi, P.; Khan, A.U. Proteomic Characterization and Target Identification Against Streptococcus mutans Under Bacitracin Stress Conditions Using LC-MS and Subtractive Proteomics. Protein J. 2022, 41, 166–178. [Google Scholar] [CrossRef]
- Bahaddad, S.A.; Almalki, M.; Alghamdi, O.A.; Sohrab, S.S.; Yasir, M.; Azhar, E.I.; Chouayekh, H. Bacillus Species as Direct-Fed Microbial Antibiotic Alternatives for Monogastric Production. Probiotics Antimicrob. Proteins 2022, 1–16. [Google Scholar] [CrossRef]
- Sitthiyotha, T.; Pichyangkura, R.; Chunsrivirot, S. Molecular dynamics provides insight into how N251A and N251Y mutations in the active site of Bacillus licheniformis RN-01 levansucrase disrupt production of long-chain levan. PLoS ONE 2018, 13, e0204915. [Google Scholar] [CrossRef] [PubMed]
- Hundschell, C.S.; Braun, A.; Wefers, D.; Vogel, R.F.; Jakob, F. Size-Dependent Variability in Flow and Viscoelastic Behavior of Levan Produced by Gluconobacter albidus TMW 2.1191. Foods 2020, 9, 192. [Google Scholar] [CrossRef] [PubMed]
- Ağçeli, G.K.; Cihangir, N. Nano-sized biopolymer levan: Its antimicrobial, anti-biofilm and anti-cancer effects. Carbohydr. Res. 2020, 494, 108068. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E.Z. Antimicrobial Activity of a Biosurfactant Produced by Bacillus licheniformis Strain M104 Grown on Whey. Braz. Arch. Biol. Technol. 2013, 56, 259–268. [Google Scholar] [CrossRef]
- Dischinger, J.; Josten, M.; Szekat, C.; Sahl, H.G.; Bierbaum, G. Production of the Novel Two-Peptide Lantibiotic Lichenicidin by Bacillus licheniformis DSM 13. PLoS ONE 2009, 4, e6788. [Google Scholar] [CrossRef]
- Barbosa, J.C.; Silva, Í.C.; Caetano, T.; Mösker, E.; Seidel, M.; Lourenço, J.; Süssmuth, R.D.; Santos, N.C.; Gonçalves, S.; Mendo, S. Assessing the potential of the two-peptide lantibiotic lichenicidin as a new generation antimicrobial. World J. Microbiol. Biotechnol. 2022, 38, 18. [Google Scholar] [CrossRef]
- Abriouel, H.; Franz, C.M.; Ben Omar, N.; Gálvez, A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef]
- Karim, R.; Mahmud, M.; Hakim, M.A. Detection of bacteriocin like substances from normal skin microflora as alternative to conventional antibiotics. Asian J. Agric. Biol. 2019, 7, 531–537. [Google Scholar]
- Mercado, V.; Olmos, J. Bacteriocin Production by Bacillus Species: Isolation, Characterization, and Application. Probiotics Antimicrob. Proteins 2022, in press. [Google Scholar] [CrossRef]
- Yu, X.J.; Han, X.T.; Li, Y.Y.; Sun, Z.; Dong, C. Isolation, identification and prokaryotic expression of a bacteriocin-like substance from Bacillus licheniformis. Chin. J. Biotechnol. 2021, 37, 2453–2462. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, J.; Pan, L.; Dai, Z.; Liu, C.; Wang, J.; Zhou, H. Production of Bacteriocin-like Substances by Bacillus spp. JY-1 in Soy Whey. Adv. Biochem. 2019, 7, 65–70. [Google Scholar] [CrossRef]
- Shobharani, P.; Padmaja, R.J.; Halami, P.M. Diversity in the antibacterial potential of probiotic cultures Bacillus licheniformis MCC2514 and Bacillus licheniformis MCC2512. Res. Microbiol. 2015, 166, 546–554. [Google Scholar] [CrossRef]
- Zaya, N.E.; Vaughan, E.E.; Shah, S.K.; Castignetti, D. Bacitracin: Substantiation and elimination of contaminating proteolytic activity and use as an affinity chromatography ligand to purify a siderophore-degrading enzyme. Curr. Microbiol. 2002, 44, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Méndez, L.R.; Arrebola, Y.; Valdés-Tresanco, M.E.; Díaz-Guevara, L.; Bergado, G.; Sánchez, B.; Charli, J.L.; Pascual Alonso, I. Bestatin and bacitracin inhibit porcine kidney cortex dipeptidyl peptidase IV activity and reduce human melanoma MeWo cell viability. Int. J. Biol. Macromol. 2020, 164, 2944–2952. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Target Sequence (Gene) | Primer Sequence (5′ to 3′) | PCR Conditions | Product Size (bp) | Source |
|---|---|---|---|---|
| Levansucrase (lsRN) | TGCTCTAGACGATTCCCGCTTATACAGACTATAGAT | 95 °C 3 min, 24× [95 °C 1 min, 55 °C 30 s, 72 °C 4 min] 72 °C 10 min | 1793 | [30] |
| CGGGATCCTTATTTGTTTACCGTTAGTTCTCCC | ||||
| Lichenicidin (bli04127) | GGAAATGATTCTTTCATGG | 95 °C 5 min, 30× [95 °C 1 min, 55 °C 1 min, 72 °C 1 min] 72 °C 5 min | 215 | [31,32] |
| TTAGTTACAGCTTGGCATG | ||||
| Lichenysin synthetase (lchAA) | GTGCCTGATGTAACGAATG | 94 °C 2 min, 30× [94 °C 15 s, 60 °C 30 s, 72 °C 50 s] 72 °C 5 min | 735 | [33] |
| CACTTCCTGCCATATACC |
| Percentage of Growth Inhibition (%) | |||
|---|---|---|---|
| Bacterial Strains | B. licheniformis DSM 13 | B. licheniformis A-1-5B-AP | B. licheniformis A-2-11B-AP |
| P. gulae 3/H | 33.86 ± 4.12 | 53.25 ± 9.21 | 5.57 ± 2.78 |
| P. intermedia 1/P | 45.78 ± 1.38 | 83.29 ± 5.12 | 54.76 ± 2.78 |
| S. mutans ATCC 35668 | 97.48 ± 0.79 | 98.24 ± 0.82 | 98.30 ± 0.79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šurín Hudáková, N.; Kačírová, J.; Sondorová, M.; Šelianová, S.; Mucha, R.; Maďar, M. Inhibitory Effect of Bacillus licheniformis Strains Isolated from Canine Oral Cavity. Life 2022, 12, 1238. https://doi.org/10.3390/life12081238
Šurín Hudáková N, Kačírová J, Sondorová M, Šelianová S, Mucha R, Maďar M. Inhibitory Effect of Bacillus licheniformis Strains Isolated from Canine Oral Cavity. Life. 2022; 12(8):1238. https://doi.org/10.3390/life12081238
Chicago/Turabian StyleŠurín Hudáková, Natália, Jana Kačírová, Miriam Sondorová, Svetlana Šelianová, Rastislav Mucha, and Marián Maďar. 2022. "Inhibitory Effect of Bacillus licheniformis Strains Isolated from Canine Oral Cavity" Life 12, no. 8: 1238. https://doi.org/10.3390/life12081238