
Isogenic Japonica Rice Koshihikari Integrated with Late Flowering Gene Hd16 and Semidwarfing Gene sd1 to Prevent High Temperature Maturation and Lodging by Typhoon


Abstract
:1. Introduction
2. Materials and Methods
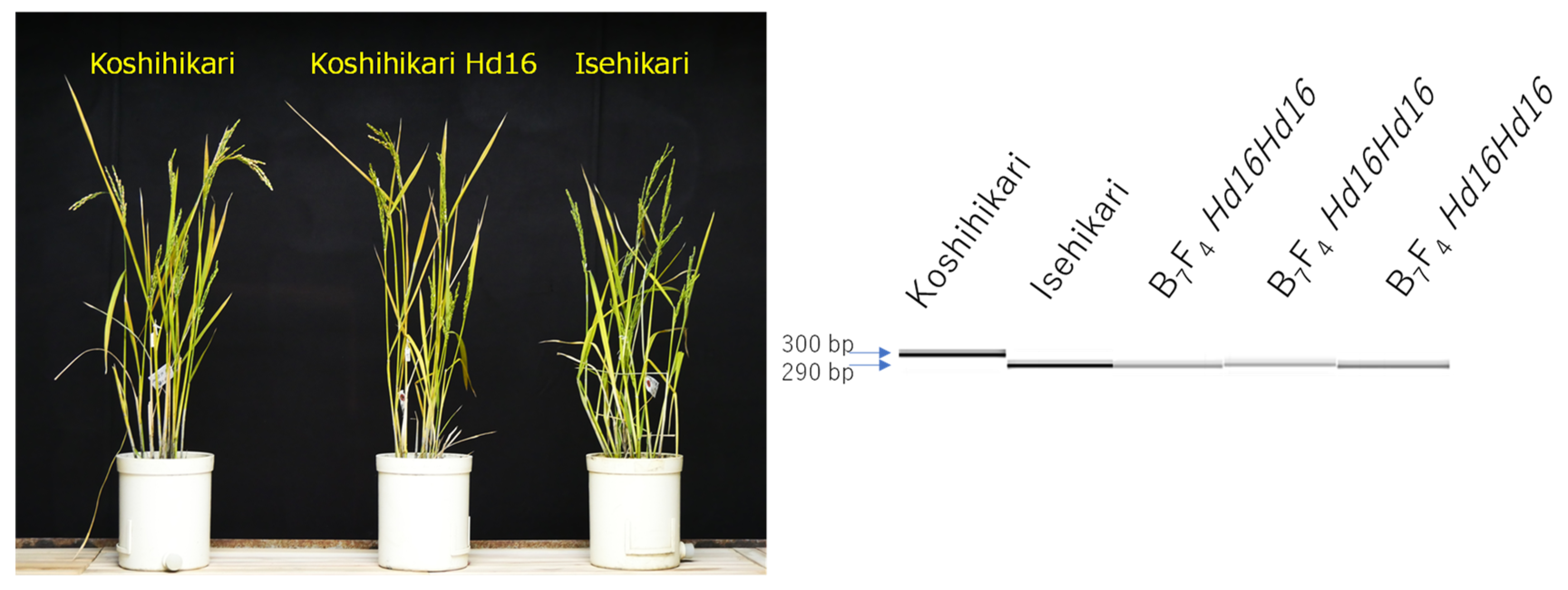
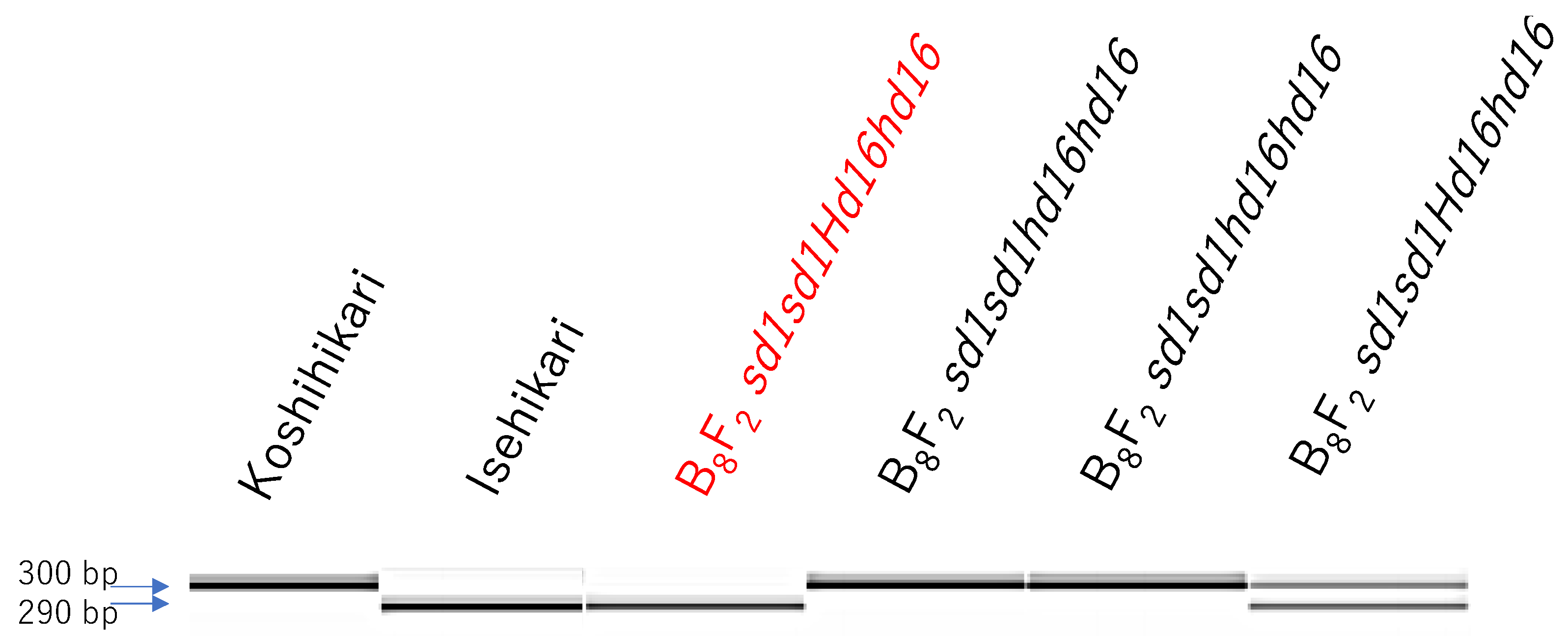
2.1. Development of Koshihikari sd1Hd16
2.2. Whole Genome Sequence Analysis
3. Results
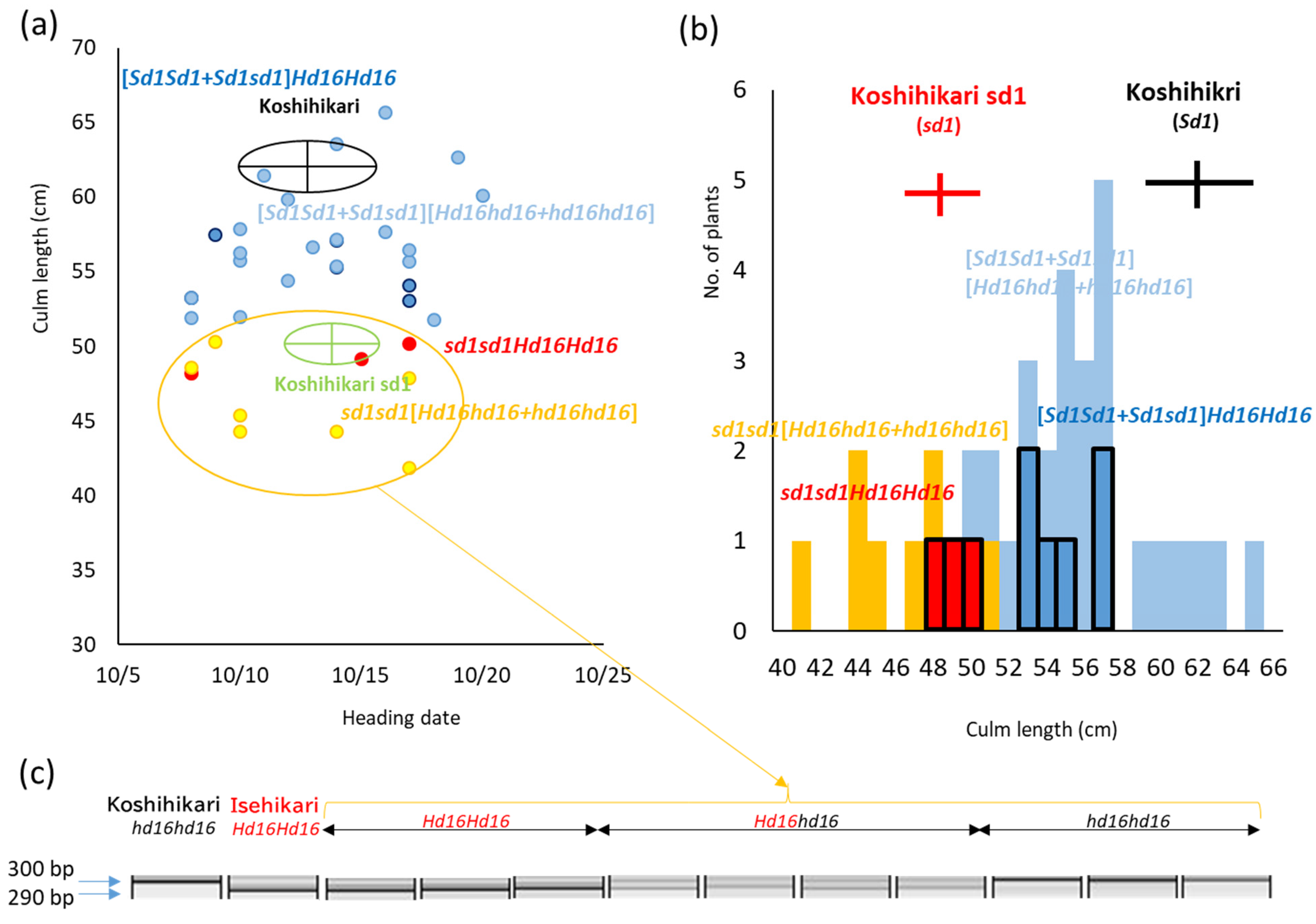
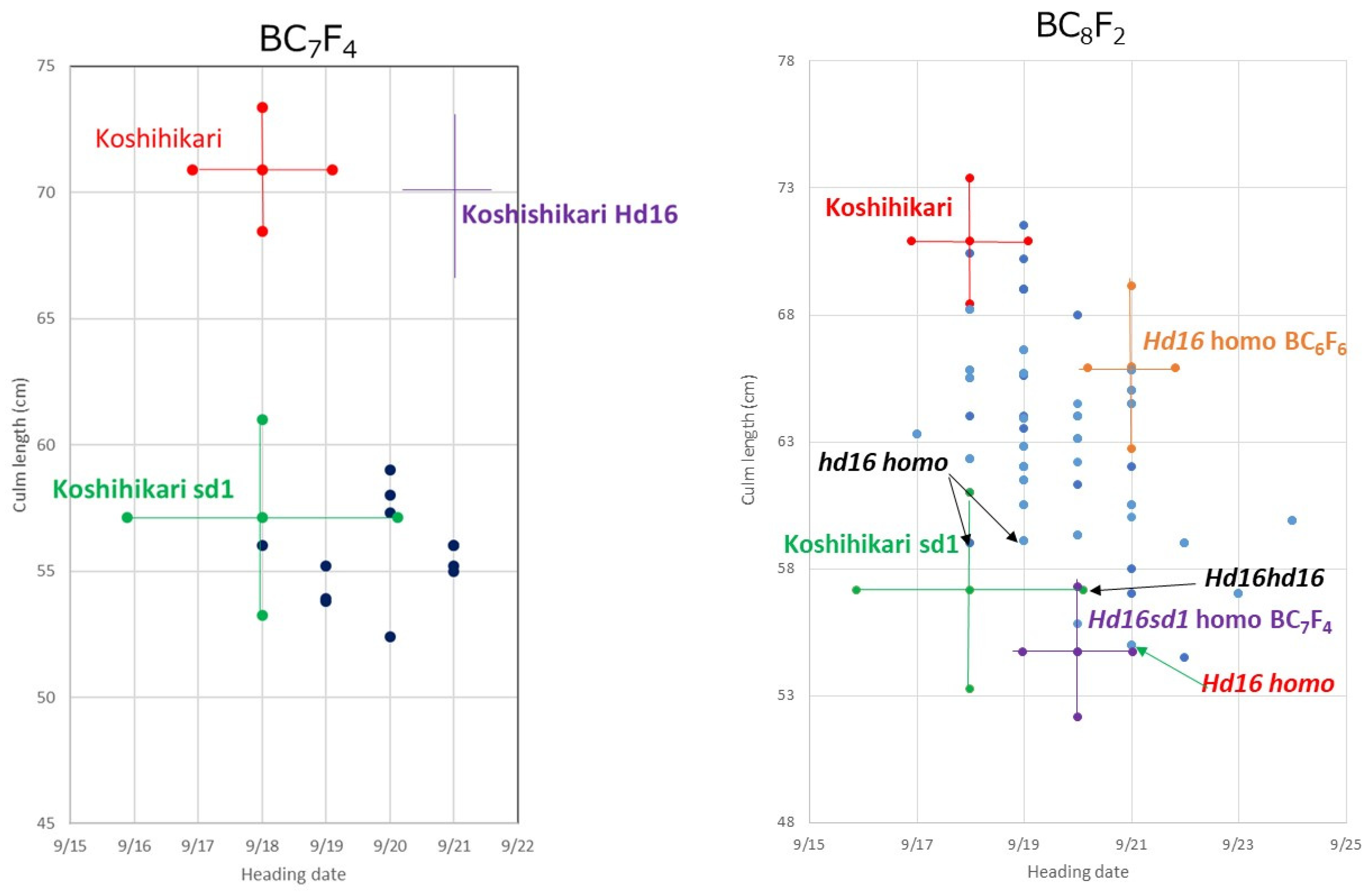
3.1. Development of Semidwarf Late-Maturing Koshihikari-Type Isogenic Line Koshihikari sd1Hd16
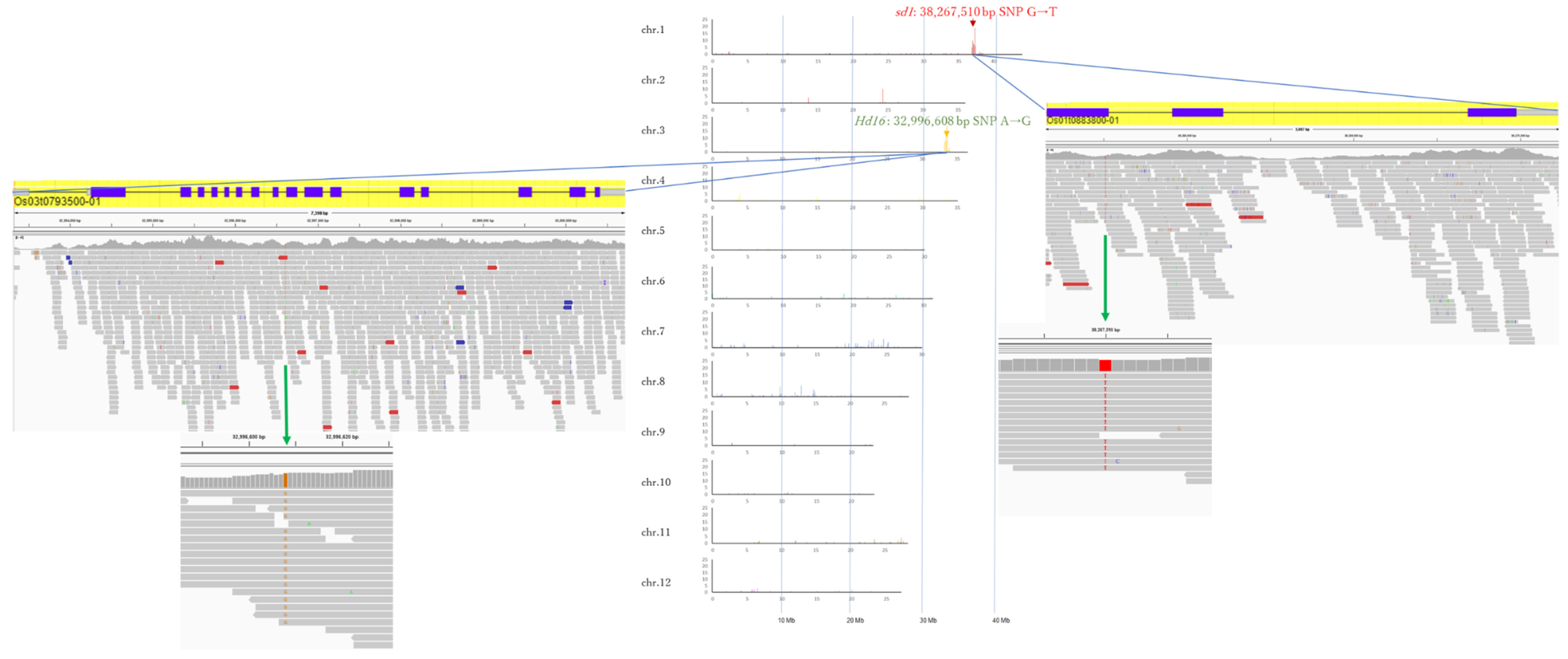
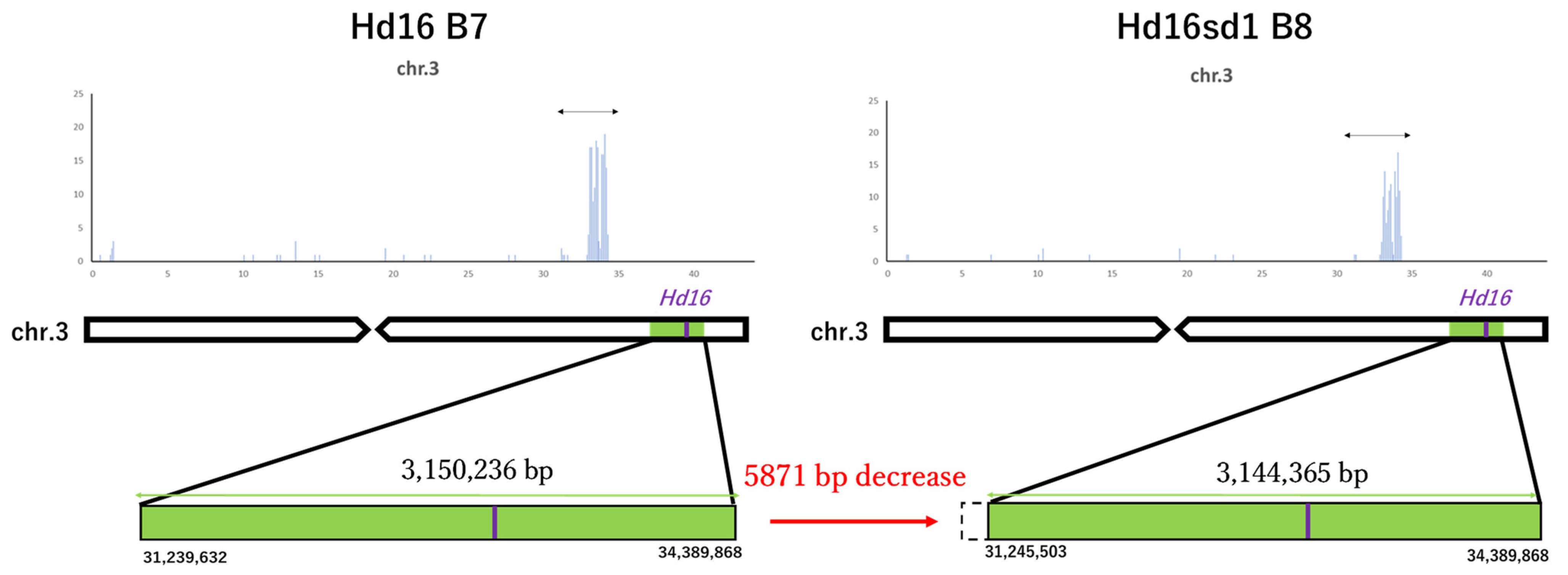
3.2. Whole Genome Sequencing of Koshihikari sd1Hd16
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Japan Meteorological Agency. 1.5 °C Global Warming: Summary of the IPCC Special Report for Policymakers (SPM) on the Impacts of Global Warming of 1.5 °C from Pre-Industrial Levels and Related Global Greenhouse Gas (GHG) Emissions in the Context of Strengthening Global Response to the Threat of Climate Change, Sustainable Development, and Poverty Eradication. 2018. Available online: http://www.env.go.jp/earth/ipcc/special_reports/sr1.5_spm.pdf (accessed on 28 July 2022).
- Dahiya, S.; Kumar, S.; Chaudhary, C. Lodging: Significance and preventive measures for increasing crop production. Int. J. Chem. Stud. 2018, 6, 700–705. [Google Scholar]
- Japan Meteorological Agency. Heisei 30 July 7 Heavy Rain and Floods. Available online: https://www.data.jma.go.jp/obd/stats/data/bosai/report/2018/20180713/20180713.html (accessed on 28 July 2022).
- Japan Meteorological Agency. 2018 (Heisei 30) Typhoons (Quick Report). Available online: https://www.jma.go.jp/jma/press/1812/21f/typhoon2018.pdf (accessed on 28 July 2022).
- Japan Fire and Disaster Management Agency. Firefighting White Paper, 2016 Edition, Volume 1. Disaster Conditions and Challenges, Section 5, Storm and Flood Damage Plan. Available online: http://www.fdma.go.jp/html/hakusho/h28/h28/pdf/part1_section5.pdf (accessed on 28 July 2022).
- Japan Meteorological Agency. Abnormal Weather Risk Map. Available online: http://www.data.jma.go.jp/cpdinfo/riskmap/heavyrain.html (accessed on 28 July 2022).
- Japan Meteorological Agency. Monthly Values from Observation Commencement. Available online: http://www.data.jma.go.jp/obd/stats/etrn/view/monthly_s3.php?prec_no=44&block_no=47662 (accessed on 28 July 2022).
- Japan Ministry of Agriculture, Forestry and Fisheries. Damage Situation by Heisei 30 July 7 Heavy Rain and Floods. Available online: http://www.maff.go.jp/j/saigai/ooame/20180628.html (accessed on 28 July 2022).
- Japan Meteorological Agency. Long-Term Changes in the Number and Frequency of Short-Term Heavy Rains Observed at AMeDAS. Available online: http://www.jma.go.jp/jma/kishou/info/heavyraintrend.html (accessed on 28 July 2022).
- Hergrove, T.; Co_man, W.R. Breeding History. In Rice that Changed the World: Cerebrating 50 Years of IR8; Rice Today: Laguna, Philippines, 2016; pp. 6–10. [Google Scholar]
- Khush, G.S. Green revolution: Preparing for the 21st century. Genome 1999, 42, 646–655. [Google Scholar] [CrossRef]
- Cho, Y.G.; Eun, M.Y.; Kim, Y.K.; Chung, T.Y.; Chae, Y.A. The semidwarf gene, sd-1, of rice (Oryza sativa L). 1. Linkage with the esterase locus, EstI-2. Theor. Appl. Genet. 1994, 89, 49–53. [Google Scholar] [CrossRef]
- Cho, Y.G.; Eun, M.Y.; McCouch, S.R.; Chae, Y.A. The semidwarf gene, sd-1, of rice (Oryza sativa L.) 2 Molecular mapping and marker-assisted selection. Theor. Appl. Genet. 1994, 89, 54–59. [Google Scholar] [CrossRef]
- Maeda, H.; Ishii, T.; Mori, H.; Kuroda, J.; Horimoto, M.; Takamure, I.; Kinoshita, T.; Kamijima, O. High density molecular map of semidwarfing gene, sd-1, in rice (Oryza sativa L.). Breed. Sci. 1997, 47, 317–320. [Google Scholar] [CrossRef]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. Green revolution: A mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef] [PubMed]
- Monna, L.; Kitazawa, N.; Yoshino, R.; Suzuki, J.; Masuda, H.; Maehara, Y.; Tanji, M.; Sato, M.; Nasu, S.; Minobe, Y. Positional cloning of rice semidwarfing gene, sd-1: Rice “Green revolution gene” encodes a mutant enzyme involved in gibberellin synthesis. DNA Res. 2002, 9, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Spielmeyer, W.; Ellis, M.H.; Chandler, P.M. Semidwarf (sd-1), “green revolution” rice, contains a defective gibberellin 20-oxidase gene. Proc. Natl. Acad. Sci. USA 2002, 99, 9043–9048. [Google Scholar] [CrossRef]
- Hedden, P. Constructing dwarf rice. Nat. Biotechnol. 2003, 21, 873–874. [Google Scholar] [CrossRef]
- Hedden, P. The genes of the Green Revolution. Trends Genet. 2003, 19, 5–9. [Google Scholar] [CrossRef]
- Tomita, M.; Ishii, K. Genetic performance of the semidwarfing allele sd1 derived from a Japonica rice cultivar and minimum requirements to detect its single-nucleotide polymorphism by MiSeq whole-genome sequencing. BioMed. Res. Int. 2018, 2018, 4241725. [Google Scholar] [CrossRef]
- Tomita, M. Introgression of Green Revolution sd1 gene into isogenic genome of rice super cultivar Koshihikari to create novel semidwarf cultivar ’Hikarishinseiki’ (Koshihikari-sd1). Field Crops Res. 2009, 114, 173–181. [Google Scholar] [CrossRef]
- Tomita, M.; Matsumoto, S. Transcription of rice green revolution gene sd1 is clarified by comparative RNA diagnosis using the isogenic background. Genom. Appl. Biol. 2011, 2, 29–35. [Google Scholar]
- MAFF. Hikarishinseiki varietal registration. In Official Gazette 8 Nov.Ministry of Agriculture, Forestry and Fisheries; MAFF: Tokyo, Japan, 2004; Volume 8. [Google Scholar]
- MAFF. Hikarishinseiki brand rice description in Okayama and Tottori prefectures. In Official Gazette 28 Mar. Ministry of Agriculture, Forestry and Fisheries; MAFF: Tokyo, Japan, 2007. [Google Scholar]
- Tomita, M. Hikarishinseiki. In Oryza sativa L. Plant Variety Protection Number: 201000072; USDA-AMS-ST-PVPO; USDA: Washington, DC, USA, 2013. Available online: http://www.ars-grin.gov/cgi-bin/npgs/pvp/showpvp.pl?pvpno=201000072 (accessed on 28 July 2022).
- Morita, S. Abortive white grains caused by high temperatures during the ripening period in rice paddies, insufficient completeness and decreased grain volume. Agric. Tech. 2005, 60, 443–446. [Google Scholar]
- Hirano, H.; Sano, Y. Comparison of waxy gene regulation in the endosperm and pollen in Oryza sativa L. Genes Genet. Syst. 2000, 75, 245–249. [Google Scholar] [CrossRef]
- Umemoto, T.; Terashima, K. Activity of granule-bound starch synthase is an important determinant of amylose content in rice endosperm. Funct. Plant Biol. 2002, 29, 1121–1124. [Google Scholar] [CrossRef]
- Yamakawa, H.; Hirose, T.; Kuroda, M.; Yamaguchi, T. Comprehensive expression profiling of rice gain filling-related genes under high temperature using DNA microarray. Plant Physiol. 2007, 144, 258–277. [Google Scholar] [CrossRef]
- Tashiro, T.; Wardlaw, I.F. The effect of high temperature on kernel dimensions and the type and occurrence of kernel damage in rice. Aust. J. Agric. Res. 1991, 42, 485–496. [Google Scholar] [CrossRef]
- Japan Ministry of Agriculture, Forestry and Fisheries. Damage Investigation. Available online: http://www.maff.go.jp/j/tokei/kouhyou/sakumotu/higai/index.html (accessed on 28 July 2022).
- Rice Stable Supply Organization. Rice Crops by Type. Available online: http://www.komenet.jp/pdf/H29sakutuke.pdf (accessed on 30 June 2022).
- Japan Ministry of Agriculture, Forestry and Fisheries. Rice product test results. Available online: http://www.maff.go.jp/j/seisan/syoryu/kensa/kome/ (accessed on 28 July 2022).
- Jinja Shimpo.“Continuation/Kuhimology” Food and Japanese People 16. Jinja Shimpo, 15 August 1996.
- Sentaku. The creation of the “Miracle Rice” at Ise Jingu Shrine’s Fields: When pale Koshihikari did not grow. Sentaku 1999, 25, 112–113. [Google Scholar]
- Jinja Shimpo. What Do People Believe? 22 Newly discovered genetic traits of Isehikari—The possibility of transposons is raising. Jinja Shimpo, 8 October 2001.
- Sentaku. The unknown “Next generation rice” Isehikari: More damage resistant and tastier than Koshihikari. Sentaku 2005, 31, 108–109. [Google Scholar]
- Saito, Y. Shinjo-sai Festival and Isehikari. 2006. Available online: http:/homepage.mac.com/saito_sy/isehikari/H151123ITniiname.html. (accessed on 30 June 2022).
- Tomita, M.; Tokuyama, R.; Matsumoto, S.; Ishii, K. Whole genome sequencing revealed a late-maturing isogenic rice Koshihikari integrated with Hd16 gene derived from an Ise Shrine mutant. Internat. J. Genom. 2022, 2022, 4565977. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler Transform. Bioinformatics 2014, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The sequence alignment/map (SAM) format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kemytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A MapReduce framework for analyzing nextgeneration DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Knutson, T.; Camargo, S.J.; Chan, J.C.L.; Emanuel, K.; Ho, C.H.; Kossin, J.; Mohapatra, M.; Satoh, M.; Sugi, M.; Walsh, K.; et al. Tropical cyclones and climate change assessment: Part I. Detection and attribution. Bull. Am. Meteorol. Soc. 2019, 100, 1987–2007. [Google Scholar] [CrossRef]
- Matsubara, K.; Hori, K.; Ogiso-Tanaka, E.; Yano, M. Cloning of quantitative trait genes from rice reveals conservation and divergence of photoperiod flowering pathways in Arabidopsis and rice. Front. Plant Sci. 2014, 15, 193. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Ogiso-Tanaka, E.; Matsubara, K.; Yamanouchi, U.; Ebana, H.; Yano, M. Hd16, a gene for casein kinase I, is involved in the control of rice flowering time by modulating the day-length response. Plant J. 2013, 76, 36–46. [Google Scholar] [CrossRef]
- Shibaya, T.; Nonoue, Y.; Ono, N.; Yamanouchi, U.; Hori, K.; Yano, M. Genetic interactions involved in the inhibition of heading by heading date QTL, Hd2 in rice under long-day conditions. Theor. Appl. Genet. 2011, 123, 1133. [Google Scholar] [CrossRef]
- Wei, X.; Xu, J.; Guo, H.; Jiang, L.; Chen, S.; Yu, C.; Zhou, Z.; Hu, P.; Zhai, H.; Wan, J. DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol. 2010, 153, 1747–1758. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Ebitani, T.; Yamamoto, T.; Sato, H.; Ohta, H.; Hirabayashi, H.; Kato, H.; Ando, I.; Nemoto, H.; Imbe, T. Development of isogenic lines of rice cultivar Koshihikari with early and late heading by marker-assisted selection. Breed. Sci. 2006, 56, 405–413. [Google Scholar] [CrossRef]
- Takeuchi, Y. Developing isogenic lines of Japanese rice cultivar ‘Koshihikari’ with early and late heading. Jpn. Agric. Res. Q. 2011, 45, 15–22. [Google Scholar] [CrossRef]
- Tanaka, N.; Saki Yoshida, S.; Takagi, H.; Terauchi, R.; Shimizu, A.; Fujiwara, T. Evidence for rice heading date 16 contribution to yield increase under low-nutrient conditions. Soil Sci. Plant Nutr. 2019, 65, 589–597. [Google Scholar] [CrossRef]
- Official Gazette No. 391, 18 March 2021, Ministry of Agriculture, Forestry and Fisheries, Plant varietal registration No. 28385, “Koshihaikri Suruga Hd16” Tokyo, Japan. 2021. Available online: http://www.hinshu2.maff.go.jp/vips/cmm/apCMM112.aspx?TOUROKU_NO=28385&LANGUAGE=Japanese. (accessed on 28 July 2022).
- Official Gazette No 1176, 26 July 2022, “Ministry of Agriculture, Forestry and Fisheries, Plant Varietal Registration No. 36171, “Koshihikari Suruga sd1Hd16” Tokyo, Japan. 2022. Available online: chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://www.maff.go.jp/j/shokusan/hinshu/gazette/syutugan/contents/314syutugan.pdf (accessed on 28 July 2022).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Days to Heading | Culm Length (cm) | Panicle Length (cm) | No. of Panicles (No./m2) | 1000-Grain Weight (g) | Grain Yield (kg/a) | Lodging Degree | Protein Content | Value of Taste |
|---|---|---|---|---|---|---|---|---|---|
| Koshihikari | 76.8 | 92.6 | 18.9 | 359 | 22.6 | 59.1 | 3.2 | 6.8 | 0.00 |
| Koshihikari Hd16 | 88.3 * | 109.0 * | 19.6 | 320 * | 22.6 | 61. | 3.5 | 6.8 | −0.13 |
| Koshihikari sd1 | 77.0 | 74.0 * | 18.8 | 375 * | 22.8 | 60.8 | 0.0 * | 7.1 | 0.14 |
| Koshihikari sd1Hd16 | 88.9 * | 78.4 * | 19.1 | 398 * | 23.0 | 63.2 * | 0.0 * | 7.0 | −0.06 |
| Position (bp) | Gene Name | Description |
|---|---|---|
| 33,625,002 | Os03g0805400 | Phosphatidic acid phosphatase type 2/haloperoxidase domain containing protein |
| 33,687,624 | Os03g0806400 | Elongation factor P family protein |
| 34,032,614 | Os03g0812200 | Zinc finger, RING/FYVE/PHD-type-domain containing protein. |
| 34,055,448 | Os03g0812800 | Calcium-binding allergen Ole e 8-like protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomita, M.; Tokuyama, R. Isogenic Japonica Rice Koshihikari Integrated with Late Flowering Gene Hd16 and Semidwarfing Gene sd1 to Prevent High Temperature Maturation and Lodging by Typhoon. Life 2022, 12, 1237. https://doi.org/10.3390/life12081237
Tomita M, Tokuyama R. Isogenic Japonica Rice Koshihikari Integrated with Late Flowering Gene Hd16 and Semidwarfing Gene sd1 to Prevent High Temperature Maturation and Lodging by Typhoon. Life. 2022; 12(8):1237. https://doi.org/10.3390/life12081237
Chicago/Turabian StyleTomita, Motonori, and Ryotaro Tokuyama. 2022. "Isogenic Japonica Rice Koshihikari Integrated with Late Flowering Gene Hd16 and Semidwarfing Gene sd1 to Prevent High Temperature Maturation and Lodging by Typhoon" Life 12, no. 8: 1237. https://doi.org/10.3390/life12081237