
Microbial Community Composition of the Antarctic Ecosystems: Review of the Bacteria, Fungi, and Archaea Identified through an NGS-Based Metagenomics Approach


Abstract
:1. Application of the Next-Generation Sequencing (NGS) Technology for Analysis of Antarctic Samples
1.1. Beginning of the Antarctic Environmental Biology
1.2. The Rise of Environmental Metagenomics
1.3. 16S Hypervariable Regions
1.4. Richness and Diversity Indices
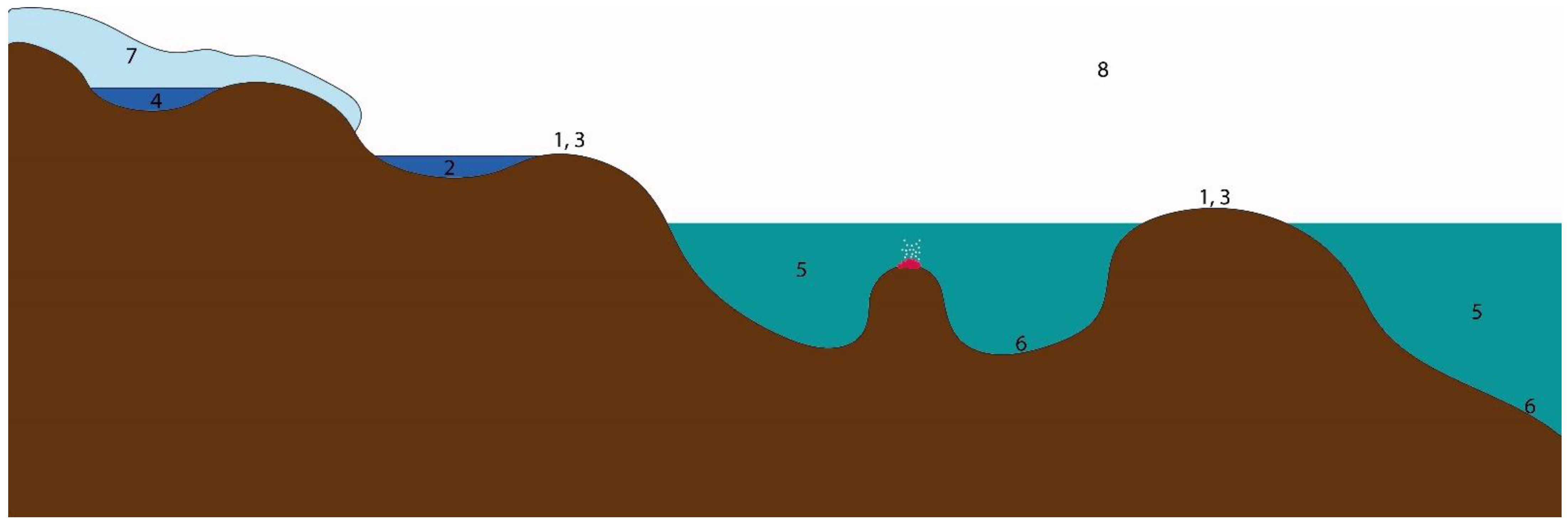
2. Community Composition of the Antarctic Ecosystems
2.1. General Trends in Antarctic Microorganism Distribution and Diversity
2.2. Sources of Microbial Life
2.3. Terrestrial Communities
2.3.1. Relationship between Geography and Microorganism Communities in Antarctic Soils
2.3.2. Relationships between Edaphic Factors and Microorganism Communities in Antarctic Soils
2.3.3. Roles of Different Microorganism Groups in Antarctic Soils
2.3.4. Lithobiont (Rock) Communities
2.4. Inland and Glacial Water Communities
2.4.1. Lake Communities
2.4.2. Meltwater Pond Communities
2.4.3. Subglacial Water Communities
2.5. Marine Communities
2.6. Microbial Mat and Sediment Communities
2.6.1. Community Composition
2.6.2. Factors Impacting the Mat Communities
2.7. Cryoconite Hole, Glacial Ice, and Snow Communities
2.7.1. Cryoconite Hole Communities
2.7.2. Ice and Glacial Microbial Communities
2.7.3. Snow Microbial Communities
2.8. Airborne Microorganisms
2.9. Impact of Macro Flora and Fauna on Microbial Communities
2.9.1. Impact of Higher Plants
2.9.2. Impact of Big Animals
2.9.3. Impact of Small Animals
2.10. Endemism
2.11. Impacts of Climatic Changes and Global Warming on Antarctic Microbial Communities
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franzmann, P.D. Examination of antarctic prokaryotic diversity through molecular comparisons. Biodivers. Conserv. 1996, 5, 1295–1305. [Google Scholar] [CrossRef]
- Lane, D.J.; Pace, B.; Olsen, G.J.; Stahl, D.A.; Sogin, M.L.; Pace, N.R. Rapid determination of 16s ribosomal rna sequences for phylogenetic analyses. Proc. Natl. Acad. Sci. USA 1985, 82, 6955–6959. [Google Scholar] [CrossRef] [Green Version]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16s ribosomal rna. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.R.; Macdonell, M.T.; Swartz, D. Identification of an antarctic endolithic microorganism by 5s rrna sequence analysis. Syst. Appl. Microbiol. 1989, 11, 182–186. [Google Scholar] [CrossRef]
- Franzmann, P.D.; Stackebrandt, E.; Sanderson, K.; Volkman, J.K.; Cameron, D.E.; Stevenson, P.L.; McMeekin, T.A.; Burton, H.R. Halobacterium lacusprofundi sp. Nov., a halophilic bacterium isolated from deep lake, antarctica. Syst. Appl. Microbiol. 1988, 11, 20–27. [Google Scholar] [CrossRef]
- Fox, E.J.; Reid-Bayliss, K.S.; Emond, M.J.; Loeb, L.A. Accuracy of next generation sequencing platforms. J. Next Gener. Seq. Appl. 2014, 1, 1000106. [Google Scholar] [CrossRef] [Green Version]
- Tytgat, B.; Verleyen, E.; Obbels, D.; Peeters, K.; De Wever, A.; D’hondt, S.; De Meyer, T.; Van Criekinge, W.; Vyverman, W.; Willems, A. Bacterial diversity assessment in antarctic terrestrial and aquatic microbial mats: A comparison between bidirectional pyrosequencing and cultivation. PLoS ONE 2014, 9, e97564. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Oh, H.-S.; Park, S.-C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16s rrna gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Porter, T.M.; Hajibabaei, M. Scaling up: A guide to high-throughput genomic approaches for biodiversity analysis. Mol. Ecol. 2018, 27, 313–338. [Google Scholar] [CrossRef] [Green Version]
- Campbell, I.B.; Claridge, G. Antarctica: Soils, Weathering Processes and Environment; Elsevier: Amsterdam, The Netherlands, 1987. [Google Scholar]
- Convey, P. Antarctic Ecosystems. 2017. Available online: https://nora.nerc.ac.uk/id/eprint/512275/ (accessed on 24 March 2022).
- Wynn-Williams, D.D. Ecological aspects of antarctic microbiology. In Advances in Microbial Ecology; Marshall, K.C., Ed.; Springer: Boston, MA, USA, 1990; pp. 71–146. [Google Scholar]
- Núñez-Montero, K.; Barrientos, L. Advances in antarctic research for antimicrobial discovery: A comprehensive narrative review of bacteria from antarctic environments as potential sources of novel antibiotic compounds against human pathogens and microorganisms of industrial importance. Antibiotics 2018, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Silva, T.R.e.; Silva, L.C.F.; de Queiroz, A.C.; Alexandre Moreira, M.S.; de Carvalho Fraga, C.A.; de Menezes, G.C.A.; Rosa, L.H.; Bicas, J.; de Oliveira, V.M.; Duarte, A.W.F. Pigments from antarctic bacteria and their biotechnological applications. Crit. Rev. Biotechnol. 2021, 41, 809–826. [Google Scholar] [CrossRef]
- Varrella, S.; Barone, G.; Tangherlini, M.; Rastelli, E.; Dell’Anno, A.; Corinaldesi, C. Diversity, ecological role and biotechnological potential of antarctic marine fungi. J. Fungi 2021, 7, 391. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; dos Santos, J.A.; Vianna, M.V.; Vieira, J.M.F.; Mallagutti, V.H.; Inforsato, F.J.; Wentzel, L.C.P.; Lario, L.D.; Rodrigues, A.; Pagnocca, F.C.; et al. Cold-adapted enzymes produced by fungi from terrestrial and marine antarctic environments. Crit. Rev. Biotechnol. 2018, 38, 600–619. [Google Scholar] [CrossRef] [Green Version]
- Peck, L.S.; Convey, P.; Barnes, D.K.A. Environmental constraints on life histories in antarctic ecosystems: Tempos, timings and predictability. Biol. Rev. 2006, 81, 75–109. [Google Scholar] [CrossRef]
- Gordon, D.A.; Priscu, J. Origin and phylogeny of microbes living in permanent antarctic lake ice. Microb. Ecol. 2000, 39, 197–202. [Google Scholar]
- Christner, B.C.; Kvitko, B.H.; Reeve, J.N. Molecular identification of bacteria and eukarya inhabiting an antarctic cryoconite hole. Extremophiles 2003, 7, 177–183. [Google Scholar] [CrossRef]
- Yergeau, E.; Newsham, K.K.; Pearce, D.A.; Kowalchuk, G.A. Patterns of bacterial diversity across a range of antarctic terrestrial habitats. Environ. Microbiol. 2007, 9, 2670–2682. [Google Scholar] [CrossRef]
- Yergeau, E.; Bokhorst, S.; Huiskes, A.H.L.; Boschker, H.T.S.; Aerts, R.; Kowalchuk, G.A. Size and structure of bacterial, fungal and nematode communities along an antarctic environmental gradient. FEMS Microbiol. Ecol. 2007, 59, 436–451. [Google Scholar] [CrossRef] [Green Version]
- Block, W.J.A.e.V. Terrestrial microbiology, invertebrates and ecosystems. Antarct. Ecol. 1984, 1, 163–236. [Google Scholar]
- Smith, R.I. Terrestrial Plant Biology of the Sub-Antarctic and Antarctic. 1984. Available online: https://nora.nerc.ac.uk/id/eprint/523706/ (accessed on 5 April 2022).
- Lawley, B.; Ripley, S.; Bridge, P.; Convey, P.J.A.; Microbiology, E. Molecular analysis of geographic patterns of eukaryotic diversity in antarctic soils. Appl. Environ. Microbiol. 2004, 70, 5963–5972. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, J.D.; Mohn, W.W. Unexpectedly high bacterial diversity in arctic tundra relative to boreal forest soils, revealed by serial analysis of ribosomal sequence tags. Appl. Environ. Microbiol. 2005, 71, 5710–5718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, K.A.; McCartney, H.A.; Lachlan-Cope, T.A.; Pearce, D.A. A preliminary study of airborne microbial biodiversity over peninsular antarctica. Cell Mol. Biol. 2004, 50, 537–542. [Google Scholar]
- Marshall, W.A. Biological particles over antarctica. Nature 1996, 383, 680. [Google Scholar] [CrossRef]
- Harris, J.M.; Tibbles, B.J. Factors affecting bacterial productivity in soils on isolated inland nunataks in continental antarctica. Microb. Ecol. 1997, 33, 106–123. [Google Scholar] [CrossRef]
- Kowalchuk, G.A.; Buma, D.S.; de Boer, W.; Klinkhamer, P.G.L.; van Veen, J.A. Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. Antonie Leeuwenhoek 2002, 81, 509. [Google Scholar] [CrossRef] [Green Version]
- Opelt, K.; Berg, G. Diversity and antagonistic potential of bacteria associated with bryophytes from nutrient-poor habitats of the baltic sea coast. Appl. Environ. Microbiol. 2004, 70, 6569–6579. [Google Scholar] [CrossRef] [Green Version]
- Smalla, K.; Wieland, G.; Buchner, A.; Zock, A.; Parzy, J.; Kaiser, S.; Roskot, N.; Heuer, H.; Berg, G. Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel electrophoresis: Plant-dependent enrichment and seasonal shifts revealed. Appl. Environ. Microbiol. 2001, 67, 4742–4751. [Google Scholar] [CrossRef] [Green Version]
- Simas, F.N.B.; Schaefer, C.E.G.R.; Melo, V.F.; Albuquerque-Filho, M.R.; Michel, R.F.M.; Pereira, V.V.; Gomes, M.R.M.; da Costa, L.M. Ornithogenic cryosols from maritime antarctica: Phosphatization as a soil forming process. Geoderma 2007, 138, 191–203. [Google Scholar] [CrossRef]
- Teixeira, L.C.R.S.; Peixoto, R.S.; Cury, J.C.; Sul, W.J.; Pellizari, V.H.; Tiedje, J.; Rosado, A.S. Bacterial diversity in rhizosphere soil from antarctic vascular plants of admiralty bay, maritime antarctica. ISME J. 2010, 4, 989–1001. [Google Scholar] [CrossRef] [Green Version]
- Aislabie, J.A.; Jordan, S.J.; Ayton, J.A.; Klassen, J.L.K.L.; Barker, G.M.B.M.; Turner, S.T. Bacterial diversity associated with ornithogenic soil of the ross sea region, antarcticathis article is one of a selection of papers in the special issue on polar and alpine microbiology. Can. J. Microbiol. 2009, 55, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.C.; Cary, S.C.; Hogg, I.D. The phylogeography of adelie penguin faecal flora. Environ. Microbiol. 2009, 11, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Ganzert, L.; Lipski, A.; Hubberten, H.-W.; Wagner, D. The impact of different soil parameters on the community structure of dominant bacteria from nine different soils located on livingston island, south shetland archipelago, antarctica. FEMS Microbiol. Ecol. 2011, 76, 476–491. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.K.; Barbier, B.A.; Bottos, E.M.; McDonald, I.R.; Cary, S.C. The inter-valley soil comparative survey: The ecology of dry valley edaphic microbial communities. ISME J. 2012, 6, 1046–1057. [Google Scholar] [CrossRef] [Green Version]
- Pearce, D.; Newsham, K.; Thorne, M.; Calvo-Bado, L.; Krsek, M.; Laskaris, P.; Hodson, A.; Wellington, E. Metagenomic analysis of a southern maritime antarctic soil. Front. Microbiol. 2012, 3, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pudasaini, S.; Wilson, J.; Ji, M.; van Dorst, J.; Snape, I.; Palmer, A.S.; Burns, B.P.; Ferrari, B.C. Microbial diversity of browning peninsula, eastern antarctica revealed using molecular and cultivation methods. Front. Microbiol. 2017, 8, 591. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.F.; Zhang, T.; Zhang, F.; Wang, E.T.; He, J.F.; Ding, H.; Zhang, B.T.; Liu, J.; Ran, X.B.; Zang, J.Y. Diversity and structure of soil bacterial communities in the fildes region (maritime antarctica) as revealed by 454 pyrosequencing. Front. Microbiol. 2015, 6, 1188. [Google Scholar] [CrossRef] [Green Version]
- Yergeau, E.; Bokhorst, S.; Kang, S.; Zhou, J.; Greer, C.W.; Aerts, R.; Kowalchuk, G.A. Shifts in soil microorganisms in response to warming are consistent across a range of antarctic environments. ISME J. 2012, 6, 692–702. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, B.C.; Bissett, A.; Snape, I.; van Dorst, J.; Palmer, A.S.; Ji, M.; Siciliano, S.D.; Stark, J.S.; Winsley, T.; Brown, M.V. Geological connectivity drives microbial community structure and connectivity in polar, terrestrial ecosystems. Environ. Microbiol. 2016, 18, 1834–1849. [Google Scholar] [CrossRef]
- Ji, M.; van Dorst, J.; Bissett, A.; Brown, M.V.; Palmer, A.S.; Snape, I.; Siciliano, S.D.; Ferrari, B.C. Microbial diversity at mitchell peninsula, eastern antarctica: A potential biodiversity “hotspot”. Polar Biol. 2016, 39, 237–249. [Google Scholar] [CrossRef]
- Chong, C.W.; Pearce, D.A.; Convey, P.; Yew, W.C.; Tan, I.K.P. Patterns in the distribution of soil bacterial 16s rrna gene sequences from different regions of antarctica. Geoderma 2012, 181–182, 45–55. [Google Scholar] [CrossRef]
- Kim, M.; Cho, A.; Lim, H.S.; Hong, S.G.; Kim, J.H.; Lee, J.; Choi, T.; Ahn, T.S.; Kim, O.-S. Highly heterogeneous soil bacterial communities around terra nova bay of northern victoria land, antarctica. PLoS ONE 2015, 10, e0119966. [Google Scholar] [CrossRef] [PubMed]
- Van Horn, D.J.; Okie, J.G.; Buelow, H.N.; Gooseff, M.N.; Barrett, J.E.; Takacs-Vesbach, C.D. Soil microbial responses to increased moisture and organic resources along a salinity gradient in a polar desert. Appl. Environ. Microbiol. 2014, 80, 3034–3043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Convey, P.; Smith, R.I.L. The terrestrial arthropod fauna and its habitats in northern marguerite bay and alexander island, maritime antarctic. Antarct. Sci. 1997, 9, 12–26. [Google Scholar] [CrossRef]
- Maslen, N.R.; Convey, P. Nematode diversity and distribution in the southern maritime antarctic—clues to history? Soil Biol. Biochem. 2006, 38, 3141–3151. [Google Scholar] [CrossRef]
- Greene, S.W.J.A.M.F.S.F. Terrestrial life of antarctica. J. Anim. Ecol. 1967, 5, 1–24. [Google Scholar]
- Romanovskaya, V.A.; Rokitko, P.V.; Mikheev, A.N.; Gushcha, N.I.; Malashenko, Y.R.; Chernaya, N.A. The effect of γ-radiation and desiccation on the viability of the soil bacteria isolated from the alienated zone around the chernobyl nuclear power plant. Microbiology 2002, 71, 608–613. [Google Scholar] [CrossRef]
- Bendia, A.G.; Signori, C.N.; Franco, D.C.; Duarte, R.T.D.; Bohannan, B.J.M.; Pellizari, V.H. A mosaic of geothermal and marine features shapes microbial community structure on deception island volcano, antarctica. Front. Microbiol. 2018, 9, 899. [Google Scholar] [CrossRef] [Green Version]
- Lezcano, M.Á.; Moreno-Paz, M.; Carrizo, D.; Prieto-Ballesteros, O.; Fernández-Martínez, M.Á.; Sánchez-García, L.; Blanco, Y.; Puente-Sánchez, F.; de Diego-Castilla, G.; García-Villadangos, M. Biomarker profiling of microbial mats in the geothermal band of cerro caliente, deception island (antarctica): Life at the edge of heat and cold. Astrobiology 2019, 19, 1490–1504. [Google Scholar] [CrossRef]
- Van Horn, D.J.; Van Horn, M.L.; Barrett, J.E.; Gooseff, M.N.; Altrichter, A.E.; Geyer, K.M.; Zeglin, L.H.; Takacs-Vesbach, C.D. Factors controlling soil microbial biomass and bacterial diversity and community composition in a cold desert ecosystem: Role of geographic scale. PLoS ONE 2013, 8, e66103. [Google Scholar] [CrossRef]
- Ugolini, F.C.; Bockheim, J.G. Antarctic soils and soil formation in a changing environment: A review. Geoderma 2008, 144, 1–8. [Google Scholar] [CrossRef]
- Cowan, D.A.; Makhalanyane, T.P.; Dennis, P.G.; Hopkins, D.W. Microbial ecology and biogeochemistry of continental antarctic soils. Front. Microbiol. 2014, 5, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onyenwoke, R.U.; Brill, J.A.; Farahi, K.; Wiegel, J. Sporulation genes in members of the low g+c gram-type-positive phylogenetic branch (firmicutes). Arch. Microbiol. 2004, 182, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Schimel, J.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef]
- Siciliano, S.D.; Palmer, A.S.; Winsley, T.; Lamb, E.; Bissett, A.; Brown, M.V.; van Dorst, J.; Ji, M.; Ferrari, B.C.; Grogan, P.; et al. Soil fertility is associated with fungal and bacterial richness, whereas ph is associated with community composition in polar soil microbial communities. Soil Biol. Biochem. 2014, 78, 10–20. [Google Scholar] [CrossRef]
- Niederberger, T.D.; Sohm, J.A.; Gunderson, T.E.; Parker, A.E.; Tirindelli, J.; Capone, D.G.; Carpenter, E.J.; Cary, S.C. Microbial community composition of transiently wetted antarctic dry valley soils. Front. Microbiol. 2015, 6, 9. [Google Scholar] [CrossRef]
- Tahon, G.; Tytgat, B.; Willems, A. Diversity of key genes for carbon and nitrogen fixation in soils from the sør rondane mountains, east antarctica. Polar Biol. 2018, 41, 2181–2198. [Google Scholar] [CrossRef]
- Richter, I.; Herbold, C.W.; Lee, C.K.; McDonald, I.R.; Barrett, J.E.; Cary, S.C. Influence of soil properties on archaeal diversity and distribution in the mcmurdo dry valleys, antarctica. FEMS Microbiol. Ecol. 2014, 89, 347–359. [Google Scholar] [CrossRef] [Green Version]
- Tiao, G.; Lee, C.K.; McDonald, I.R.; Cowan, D.A.; Cary, S.C. Rapid microbial response to the presence of an ancient relic in the antarctic dry valleys. Nat. Commun. 2012, 3, 660. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.-S.; Chae, N.; Lim, H.S.; Cho, A.; Kim, J.H.; Hong, S.G.; Oh, J. Bacterial diversity in ornithogenic soils compared to mineral soils on king george island, antarctica. J. Microbiol. 2012, 50, 1081–1085. [Google Scholar] [CrossRef]
- Cottrell, M.T.; Kirchman, D.L. Natural assemblages of marine proteobacteria and members of the cytophaga-flavobacter cluster consuming low- and high-molecular-weight dissolved organic matter. Appl. Environ. Microbiol. 2000, 66, 1692–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, D.; Yau, S.; Williams, T.J.; Allen, M.A.; Brown, M.V.; DeMaere, M.Z.; Lauro, F.M.; Cavicchioli, R. Key microbial drivers in antarctic aquatic environments. FEMS Microbiol. Rev. 2013, 37, 303–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cary, S.C.; McDonald, I.R.; Barrett, J.E.; Cowan, D.A. On the rocks: The microbiology of antarctic dry valley soils. Nat. Rev. Microbiol. 2010, 8, 129–138. [Google Scholar] [CrossRef]
- Pointing, S.B.; Chan, Y.; Lacap, D.C.; Lau, M.C.Y.; Jurgens, J.A.; Farrell, R.L. Highly specialized microbial diversity in hyper-arid polar desert. Proc. Natl. Acad. Sci. USA 2009, 106, 19964–19969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, S.A.; Rueckert, A.; Cowan, D.A.; Cary, S.C. Sources of edaphic cyanobacterial diversity in the dry valleys of eastern antarctica. ISME J. 2008, 2, 308–320. [Google Scholar] [CrossRef] [Green Version]
- Van Goethem, M.W.; Makhalanyane, T.P.; Valverde, A.; Cary, S.C.; Cowan, D.A. Characterization of bacterial communities in lithobionts and soil niches from victoria valley, antarctica. FEMS Microbiol. Ecol. 2016, 92, fiw051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.; Lacap, D.C.; Lau, M.C.Y.; Ha, K.Y.; Warren-Rhodes, K.A.; Cockell, C.S.; Cowan, D.A.; McKay, C.P.; Pointing, S.B. Hypolithic microbial communities: Between a rock and a hard place. Environ. Microbiol. 2012, 14, 2272–2282. [Google Scholar] [CrossRef]
- Warren-Rhodes, K.A.; Rhodes, K.L.; Pointing, S.B.; Ewing, S.A.; Lacap, D.C.; Gómez-Silva, B.; Amundson, R.; Friedmann, E.I.; McKay, C.P. Hypolithic cyanobacteria, dry limit of photosynthesis, and microbial ecology in the hyperarid atacama desert. Microb. Ecol. 2006, 52, 389–398. [Google Scholar] [CrossRef]
- Lacap-Bugler, D.C.; Lee, K.K.; Archer, S.; Gillman, L.N.; Lau, M.C.Y.; Leuzinger, S.; Lee, C.K.; Maki, T.; McKay, C.P.; Perrott, J.K.; et al. Global diversity of desert hypolithic cyanobacteria. Front. Microbiol. 2017, 8, 867. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.T.S.; Lacap-Bugler, D.C.; Lau, M.C.Y.; Caruso, T.; Rao, S.; de los Rios, A.; Archer, S.K.; Chiu, J.M.Y.; Higgins, C.; Van Nostrand, J.D.; et al. Taxonomic and functional diversity of soil and hypolithic microbial communities in miers valley, mcmurdo dry valleys, antarctica. Front. Microbiol. 2016, 7, 1642. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Tuffin, M.; Stafford, W.; Cary, C.; Lacap, D.C.; Pointing, S.B.; Cowan, D. Hypolithic microbial communities of quartz rocks from miers valley, mcmurdo dry valleys, antarctica. Polar Biol. 2011, 34, 1657. [Google Scholar] [CrossRef]
- Sutherland, D.L. Microbial mat communities in response to recent changes in the physiochemical environment of the meltwater ponds on the mcmurdo ice shelf, antarctica. Polar Biol. 2009, 32, 1023–1032. [Google Scholar] [CrossRef]
- Li, W.; Morgan-Kiss, R.M. Influence of environmental drivers and potential interactions on the distribution of microbial communities from three permanently stratified antarctic lakes. Front. Microbiol. 2019, 10, 1067. [Google Scholar] [CrossRef] [PubMed]
- Picazo, A.; Rochera, C.; Villaescusa, J.A.; Miralles-Lorenzo, J.; Velázquez, D.; Quesada, A.; Camacho, A. Bacterioplankton community composition along environmental gradients in lakes from byers peninsula (maritime antarctica) as determined by next-generation sequencing. Front. Microbiol. 2019, 10, 908. [Google Scholar] [CrossRef]
- Huang, J.P.; Swain, A.K.; Thacker, R.W.; Ravindra, R.; Andersen, D.T.; Bej, A.K. Bacterial diversity of the rock-water interface in an east antarctic freshwater ecosystem, lake tawani(p)†. Aquat. Biosyst. 2013, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Vick-Majors, T.J.; Priscu, J.C.; A Amaral-Zettler, L. Modular community structure suggests metabolic plasticity during the transition to polar night in ice-covered antarctic lakes. ISME J. 2014, 8, 778–789. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Sáez, L.; Sánchez, O.; Gasol, J.M.; Balagué, V.; Pedrós-Alio, C. Winter-to-summer changes in the composition and single-cell activity of near-surface arctic prokaryotes. Environ. Microbiol. 2008, 10, 2444–2454. [Google Scholar] [CrossRef]
- Foreman, C.M.; Dieser, M.; Greenwood, M.; Cory, R.M.; Laybourn-Parry, J.; Lisle, J.T.; Jaros, C.; Miller, P.L.; Chin, Y.-P.; McKnight, D.M. When a habitat freezes solid: Microorganisms over-winter within the ice column of a coastal antarctic lake. FEMS Microbiol. Ecol. 2011, 76, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Weisleitner, K.; Perras, A.; Moissl-Eichinger, C.; Andersen, D.T.; Sattler, B. Source environments of the microbiome in perennially ice-covered lake untersee, antarctica. Front. Microbiol. 2019, 10, 1019. [Google Scholar] [CrossRef]
- Rogers, S.O.; Shtarkman, Y.M.; Koçer, Z.A.; Edgar, R.; Veerapaneni, R.; D’Elia, T. Ecology of subglacial lake vostok (antarctica), based on metagenomic/metatranscriptomic analyses of accretion ice. Biology 2013, 2, 629–650. [Google Scholar] [CrossRef]
- Gura, C.; Rogers, S.O. Metatranscriptomic and metagenomic analysis of biological diversity in subglacial lake vostok (antarctica). Biology 2020, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Achberger, A.M.; Christner, B.C.; Michaud, A.B.; Priscu, J.C.; Skidmore, M.L.; Vick-Majors, T.J.; WISSARD Science Team. Microbial community structure of subglacial lake whillans, west antarctica. Front. Microbiol. 2016, 7, 1457. [Google Scholar] [CrossRef]
- Archer, S.D.J.; McDonald, I.R.; Herbold, C.W.; Cary, S.C. Characterisation of bacterioplankton communities in the meltwater ponds of bratina island, victoria land, antarctica. FEMS Microbiol. Ecol. 2014, 89, 451–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, S.D.J.; McDonald, I.R.; Herbold, C.W.; Lee, C.K.; Niederberger, T.S.; Cary, C. Temporal, regional and geochemical drivers of microbial community variation in the melt ponds of the ross sea region, antarctica. Polar Biol. 2016, 39, 267–282. [Google Scholar] [CrossRef]
- Campen, R.; Kowalski, J.; Lyons, W.B.; Tulaczyk, S.; Dachwald, B.; Pettit, E.; Welch, K.A.; Mikucki, J.A. Microbial diversity of an antarctic subglacial community and high-resolution replicate sampling inform hydrological connectivity in a polar desert. Environ. Microbiol. 2019, 21, 2290–2306. [Google Scholar] [CrossRef]
- Perini, L.; Gostinčar, C.; Gunde-Cimerman, N. Fungal and bacterial diversity of svalbard subglacial ice. Sci. Rep. 2019, 9, 20230. [Google Scholar] [CrossRef]
- Ghiglione, J.F.; Murray, A.E. Pronounced summer to winter differences and higher wintertime richness in coastal antarctic marine bacterioplankton. Environ. Microbiol. 2012, 14, 617–629. [Google Scholar] [CrossRef]
- Signori, C.N.; Thomas, F.; Enrich-Prast, A.; Pollery, R.C.G.; Sievert, S.M. Microbial diversity and community structure across environmental gradients in bransfield strait, western antarctic peninsula. Front. Microbiol. 2014, 5, 647. [Google Scholar] [CrossRef] [PubMed]
- Luria, C.M.; Ducklow, H.W.; Amaral-Zettler, L.A. Marine bacterial, archaeal and eukaryotic diversity and community structure on the continental shelf of the western antarctic peninsula. Aquat. Microb. Ecol. 2014, 73, 107–121. [Google Scholar] [CrossRef]
- Ghiglione, J.-F.; Galand, P.E.; Pommier, T.; Pedrós-Alió, C.; Maas, E.W.; Bakker, K.; Bertilson, S.; Kirchman, D.L.; Lovejoy, C.; Yager, P.L.; et al. Pole-to-pole biogeography of surface and deep marine bacterial communities. Proc. Natl. Acad. Sci. USA 2012, 109, 17633–17638. [Google Scholar] [CrossRef] [Green Version]
- Sadaiappan, B.; Kannan, S.; Palaniappan, S.; Manikkam, R.; Ramasamy, B.; Anilkumar, N.; Subramanian, M. Metagenomic 16s rdna amplicon data of microbial diversity and its predicted metabolic functions in the southern ocean (antarctic). Data Brief 2020, 28, 104876. [Google Scholar] [CrossRef]
- Signori, C.N.; Pellizari, V.H.; Enrich-Prast, A.; Sievert, S.M. Spatiotemporal dynamics of marine bacterial and archaeal communities in surface waters off the northern antarctic peninsula. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 149, 150–160. [Google Scholar] [CrossRef]
- Merbt, S.N.; Stahl, D.A.; Casamayor, E.O.; Martí, E.; Nicol, G.W.; Prosser, J.I. Differential photoinhibition of bacterial and archaeal ammonia oxidation. FEMS Microbiol. Lett. 2012, 327, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Sáez, L.; Andersson, A.; Heinrich, F.; Bertilsson, S. High archaeal diversity in antarctic circumpolar deep waters. Environ. Microbiol. Rep. 2011, 3, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Zhu, H.; Wang, J.; Li, T. Extracellular hydrolytic enzyme screening of culturable heterotrophic bacteria from deep-sea sediments of the southern okinawa trough. World J. Microbiol. Biotechnol. 2009, 25, 71–79. [Google Scholar] [CrossRef]
- Swan, B.K.; Martinez-Garcia, M.; Preston, C.M.; Sczyrba, A.; Woyke, T.; Lamy, D.; Reinthaler, T.; Poulton, N.J.; Masland, E.D.P.; Gomez, M.L.; et al. Potential for chemolithoautotrophy among ubiquitous bacteria lineages in the dark ocean. Science 2011, 333, 1296–1300. [Google Scholar] [CrossRef] [Green Version]
- Teeling, H.; Fuchs, B.M.; Becher, D.; Klockow, C.; Gardebrecht, A.; Bennke, C.M.; Kassabgy, M.; Huang, S.; Mann, A.J.; Waldmann, J.; et al. Substrate-controlled succession of marine bacterioplankton populations induced by a phytoplankton bloom. Science 2012, 336, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Grzymski, J.J.; Riesenfeld, C.S.; Williams, T.J.; Dussaq, A.M.; Ducklow, H.; Erickson, M.; Cavicchioli, R.; Murray, A.E. A metagenomic assessment of winter and summer bacterioplankton from antarctica peninsula coastal surface waters. ISME J. 2012, 6, 1901–1915. [Google Scholar] [CrossRef]
- Zeng, Y.-X.; Yu, Y.; Qiao, Z.-Y.; Jin, H.-Y.; Li, H.-R. Diversity of bacterioplankton in coastal seawaters of fildes peninsula, king george island, antarctica. Arch. Microbiol. 2014, 196, 137–147. [Google Scholar] [CrossRef]
- González, J.M.; Fernández-Gómez, B.; Fernàndez-Guerra, A.; Gómez-Consarnau, L.; Sánchez, O.; Coll-Lladó, M.; del Campo, J.; Escudero, L.; Rodríguez-Martínez, R.; Alonso-Sáez, L.; et al. Genome analysis of the proteorhodopsin-containing marine bacterium polaribacter sp. Med152 (flavobacteria). Proc. Natl. Acad. Sci. USA 2008, 105, 8724–8729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, M.V.; Lauro, F.M.; DeMaere, M.Z.; Muir, L.; Wilkins, D.; Thomas, T.; Riddle, M.J.; Fuhrman, J.A.; Andrews-Pfannkoch, C.; Hoffman, J.M.; et al. Global biogeography of sar11 marine bacteria. Mol. Syst. Biol. 2012, 8, 595. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.C.; Clarke, K.J. Fine structure of a terrestrial cyanobacterial mat from antarctica1. J. Phycol. 1992, 28, 199–202. [Google Scholar] [CrossRef]
- Jackson, E.E.; Hawes, I.; Jungblut, A.D. 16s rrna gene and 18s rrna gene diversity in microbial mat communities in meltwater ponds on the mcmurdo ice shelf, antarctica. Polar Biol. 2021, 44, 823–836. [Google Scholar] [CrossRef]
- Stanish, L.F.; O’Neill, S.P.; Gonzalez, A.; Legg, T.M.; Knelman, J.; McKnight, D.M.; Spaulding, S.; Nemergut, D.R. Bacteria and diatom co-occurrence patterns in microbial mats from polar desert streams. Environ. Microbiol. 2013, 15, 1115–1131. [Google Scholar] [CrossRef]
- Valdespino-Castillo, P.M.; Cerqueda-García, D.; Espinosa, A.C.; Batista, S.; Merino-Ibarra, M.; Taş, N.; Alcántara-Hernández, R.J.; Falcón, L.I. Microbial distribution and turnover in antarctic microbial mats highlight the relevance of heterotrophic bacteria in low-nutrient environments. FEMS Microbiol. Ecol. 2018, 94, fiy129. [Google Scholar] [CrossRef] [Green Version]
- Varin, T.; Lovejoy, C.; Jungblut, A.D.; Vincent, W.F.; Corbeil, J. Metagenomic analysis of stress genes in microbial mat communities from antarctica and the high arctic. Appl. Environ. Microbiol. 2012, 78, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Berg, K.A.; Lyra, C.; Sivonen, K.; Paulin, L.; Suomalainen, S.; Tuomi, P.; Rapala, J. High diversity of cultivable heterotrophic bacteria in association with cyanobacterial water blooms. ISME J. 2009, 3, 314–325. [Google Scholar] [CrossRef]
- Seckbach, J.; Oren, A. Microbial Mats: Modern and Ancient Microorganisms in Stratified Systems; Springer: Berlin/Heidelberg, Germany, 2010; Volume 14. [Google Scholar]
- Kleinteich, J.; Hildebrand, F.; Wood, S.A.; Ciŕs, S.; Agha, R.; Quesada, A.; Pearce, D.A.; Convey, P.; Kpper, F.C.; Dietrich, D.R. Diversity of toxin and non-toxin containing cyanobacterial mats of meltwater ponds on the antarctic peninsula: A pyrosequencing approach. Antarct. Sci. 2014, 26, 521–532. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.; Mojib, N.; Hakim, J.A.; Hawes, I.; Tanabe, Y.; Andersen, D.T.; Bej, A.K. Microbial communities and their predicted metabolic functions in growth laminae of a unique large conical mat from lake untersee, east antarctica. Front. Microbiol. 2017, 8, 1347. [Google Scholar] [CrossRef] [Green Version]
- Pessi, I.S.; Lara, Y.; Durieu, B.; Maalouf, P.d.C.; Verleyen, E.; Wilmotte, A. Community structure and distribution of benthic cyanobacteria in antarctic lacustrine microbial mats. FEMS Microbiol. Ecol. 2018, 94, fiy042. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, R.C.; Niederberger, T.D.; Greer, C.; Whyte, L.G. Microbial diversity of active layer and permafrost in an acidic wetland from the canadian high arctic. Can. J. Microbiol. 2011, 57, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Chin, K.-J.; Janssen, P.H. Propionate formation by opitutus terrae in pure culture and in mixed culture with a hydrogenotrophic methanogen and implications for carbon fluxes in anoxic rice paddy soil. Appl. Environ. Microbiol. 2002, 68, 2089–2092. [Google Scholar] [CrossRef] [Green Version]
- Larose, C.; Prestat, E.; Cecillon, S.; Berger, S.; Malandain, C.; Lyon, D.; Ferrari, C.; Schneider, D.; Dommergue, A.; Vogel, T.M. Interactions between snow chemistry, mercury inputs and microbial population dynamics in an arctic snowpack. PLoS ONE 2013, 8, e79972. [Google Scholar] [CrossRef] [Green Version]
- Zaikova, E.; Goerlitz, D.S.; Tighe, S.W.; Wagner, N.Y.; Bai, Y.; Hall, B.L.; Bevilacqua, J.G.; Weng, M.M.; Samuels-Fair, M.D.; Johnson, S.S. Antarctic relic microbial mat community revealed by metagenomics and metatranscriptomics. Front. Ecol. Evol. 2019, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Van Horn, D.J.; Wolf, C.R.; Colman, D.R.; Jiang, X.; Kohler, T.J.; McKnight, D.M.; Stanish, L.F.; Yazzie, T.; Takacs-Vesbach, C.D. Patterns of bacterial biodiversity in the glacial meltwater streams of the mcmurdo dry valleys, antarctica. FEMS Microbiol. Ecol. 2016, 92, fiw148. [Google Scholar] [CrossRef] [Green Version]
- Ogaki, M.B.; Pinto, O.H.B.; Vieira, R.; Neto, A.A.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Câmara, P.E.A.S.; Rosa, L.H. Fungi present in antarctic deep-sea sediments assessed using DNA metabarcoding. Microb. Ecol. 2021, 82, 157–164. [Google Scholar] [CrossRef]
- Emil Ruff, S.; Probandt, D.; Zinkann, A.-C.; Iversen, M.H.; Klaas, C.; Würzberg, L.; Krombholz, N.; Wolf-Gladrow, D.; Amann, R.; Knittel, K. Indications for algae-degrading benthic microbial communities in deep-sea sediments along the antarctic polar front. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 108, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Franco, D.C.; Signori, C.N.; Duarte, R.T.D.; Nakayama, C.R.; Campos, L.S.; Pellizari, V.H. High prevalence of gammaproteobacteria in the sediments of admiralty bay and north bransfield basin, northwestern antarctic peninsula. Front. Microbiol. 2017, 8, 153. [Google Scholar] [CrossRef] [Green Version]
- Learman, D.R.; Henson, M.W.; Thrash, J.C.; Temperton, B.; Brannock, P.M.; Santos, S.R.; Mahon, A.R.; Halanych, K.M. Biogeochemical and microbial variation across 5500 km of antarctic surface sediment implicates organic matter as a driver of benthic community structure. Front. Microbiol. 2016, 7, 284. [Google Scholar] [CrossRef]
- Carr, S.A.; Orcutt, B.N.; Mandernack, K.W.; Spear, J.R. Abundant atribacteria in deep marine sediment from the adélie basin, antarctica. Front. Microbiol. 2015, 6, 872. [Google Scholar] [CrossRef] [Green Version]
- Bahl, J.; Lau, M.C.Y.; Smith, G.J.D.; Vijaykrishna, D.; Cary, S.C.; Lacap, D.C.; Lee, C.K.; Papke, R.T.; Warren-Rhodes, K.A.; Wong, F.K.Y.; et al. Ancient origins determine global biogeography of hot and cold desert cyanobacteria. Nat. Commun. 2011, 2, 163. [Google Scholar] [CrossRef]
- Hodgson, D.A.; Convey, P.; Verleyen, E.; Vyverman, W.; McInnes, S.J.; Sands, C.J.; Fernández-Carazo, R.; Wilmotte, A.; De Wever, A.; Peeters, K.; et al. The limnology and biology of the dufek massif, transantarctic mountains 82° south. Polar Sci. 2010, 4, 197–214. [Google Scholar] [CrossRef]
- McKnight, D.M.; Tate, C.M.; Andrews, E.D.; Niyogi, D.K.; Cozzetto, K.; Welch, K.; Lyons, W.B.; Capone, D.G. Reactivation of a cryptobiotic stream ecosystem in the mcmurdo dry valleys, antarctica: A long-term geomorphological experiment. Geomorphology 2007, 89, 186–204. [Google Scholar] [CrossRef]
- de Menezes, G.C.A.; Câmara, P.E.A.S.; Pinto, O.H.B.; Convey, P.; Carvalho-Silva, M.; Simões, J.C.; Rosa, C.A.; Rosa, L.H. Fungi in the antarctic cryosphere: Using DNA metabarcoding to reveal fungal diversity in glacial ice from the antarctic peninsula region. Microb. Ecol. 2021, 83, 647–657. [Google Scholar] [CrossRef]
- Lutz, S.; Ziolkowski, L.A.; Benning, L.G. The biodiversity and geochemistry of cryoconite holes in queen maud land, east antarctica. Microorganisms 2019, 7, 160. [Google Scholar] [CrossRef] [Green Version]
- Au-Weisleitner, K.; Au-Hunger, L.; Au-Kohstall, C.; Au-Frisch, A.; Au-Storrie-Lombardi, M.C.; Au-Sattler, B. Laser-induced fluorescence emission (l.I.F.E.) as novel non-invasive tool for in-situ measurements of biomarkers in cryospheric habitats. JoVE 2019, 152, e60447. [Google Scholar]
- Price, P.B. Microbial life in glacial ice and implications for a cold origin of life. FEMS Microbiol. Ecol. 2007, 59, 217–231. [Google Scholar] [CrossRef] [Green Version]
- Bowman, J.S.; Rasmussen, S.; Blom, N.; Deming, J.W.; Rysgaard, S.; Sicheritz-Ponten, T. Microbial community structure of arctic multiyear sea ice and surface seawater by 454 sequencing of the 16s rna gene. ISME J. 2012, 6, 11–20. [Google Scholar] [CrossRef]
- Vishnivetskaya, T.A.; Erokhina, L.G.; Spirina, E.V.; Shatilovich, A.V.; Vorobyova, E.A.; Tsapin, A.I.; Gilichinsky, D.A. Viable Phototrophs: Cyanobacteria and Green Algae from the Permafrost Darkness; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- García-Lopez, E.; Serrano, S.; Calvo, M.A.; Peña Perez, S.; Sanchez-Casanova, S.; García-Descalzo, L.; Cid, C. Microbial community structure driven by a volcanic gradient in glaciers of the antarctic archipelago south shetland. Microorganisms 2021, 9, 392. [Google Scholar] [CrossRef]
- Martinez-Alonso, E.; Pena-Perez, S.; Serrano, S.; Garcia-Lopez, E.; Alcazar, A.; Cid, C. Taxonomic and functional characterization of a microbial community from a volcanic englacial ecosystem in deception island, antarctica. Sci. Rep. 2019, 9, 12158. [Google Scholar] [CrossRef]
- Rosa, L.H.; Pinto, O.H.B.; Šantl-Temkiv, T.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Câmara, P.E.A.S. DNA metabarcoding of fungal diversity in air and snow of livingston island, south shetland islands, antarctica. Sci. Rep. 2020, 10, 21793. [Google Scholar] [CrossRef]
- da Silva, T.H.; Câmara, P.E.A.S.; Pinto, O.H.B.; Carvalho-Silva, M.; Oliveira, F.S.; Convey, P.; Rosa, C.A.; Rosa, L.H. Diversity of fungi present in permafrost in the south shetland islands, maritime antarctic. Microb. Ecol. 2022, 83, 58–67. [Google Scholar] [CrossRef]
- Lopatina, A.; Medvedeva, S.; Shmakov, S.; Logacheva, M.D.; Krylenkov, V.; Severinov, K. Metagenomic analysis of bacterial communities of antarctic surface snow. Front. Microbiol. 2016, 7, 398. [Google Scholar] [CrossRef]
- Malard, L.A.; Šabacká, M.; Magiopoulos, I.; Mowlem, M.; Hodson, A.; Tranter, M.; Siegert, M.J.; Pearce, D.A. Spatial variability of antarctic surface snow bacterial communities. Front. Microbiol. 2019, 10, 461. [Google Scholar] [CrossRef]
- Michaud, L.; Lo Giudice, A.; Mysara, M.; Monsieurs, P.; Raffa, C.; Leys, N.; Amalfitano, S.; Van Houdt, R. Snow surface microbiome on the high antarctic plateau (dome c). PLoS ONE 2014, 9, e104505. [Google Scholar] [CrossRef]
- Bottos, E.M.; Woo, A.C.; Zawar-Reza, P.; Pointing, S.B.; Cary, S.C. Airborne bacterial populations above desert soils of the mcmurdo dry valleys, antarctica. Microb. Ecol. 2014, 67, 120–128. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Yu, X.; Ju, F.; Zhan, H.; Jiang, B.; Kang, H.; Xie, Z. Airborne bacterial community diversity, source and function along the antarctic coast. Sci. Total Environ. 2021, 765, 142700. [Google Scholar] [CrossRef]
- Roesch, L.F.W.; Fulthorpe, R.R.; Pereira, A.B.; Pereira, C.K.; Lemos, L.N.; Barbosa, A.D.; Suleiman, A.K.A.; Gerber, A.L.; Pereira, M.G.; Loss, A.; et al. Soil bacterial community abundance and diversity in ice-free areas of keller peninsula, antarctica. Appl. Soil Ecol. 2012, 61, 7–15. [Google Scholar] [CrossRef]
- Janssen, P.H. Identifying the dominant soil bacterial taxa in libraries of 16s rrna and 16s rrna genes. Appl. Environ. Microbiol. 2006, 72, 1719–1728. [Google Scholar] [CrossRef] [Green Version]
- Buyer, J.S.; Roberts, D.P.; Russek-Cohen, E. Microbial community structure and function in the spermosphere as affected by soil and seed type. Can. J. Microbiol. 1999, 45, 138–144. [Google Scholar] [CrossRef]
- Garbeva, P.; Veen, J.A.v.; Elsas, J.D.v. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef]
- Smith, J.J.; Tow, L.A.; Stafford, W.; Cary, C.; Cowan, D.A. Bacterial diversity in three different antarctic cold desert mineral soils. Microb. Ecol. 2006, 51, 413–421. [Google Scholar] [CrossRef]
- Zhu, R.; Shi, Y.; Ma, D.; Wang, C.; Xu, H.; Chu, H. Bacterial diversity is strongly associated with historical penguin activity in an antarctic lake sediment profile. Sci. Rep. 2015, 5, 17231. [Google Scholar] [CrossRef]
- Rodríguez-Marconi, S.; De la Iglesia, R.; Díez, B.; Fonseca, C.A.; Hajdu, E.; Trefault, N. Characterization of bacterial, archaeal and eukaryote symbionts from antarctic sponges reveals a high diversity at a three-domain level and a particular signature for this ecosystem. PLoS ONE 2015, 10, e0138837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergstrom, D.M.; Convey, P.; Huiskes, A.H. Trends in Antarctic Terrestrial and Limnetic Ecosystems; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Rosa, L.H.; Zani, C.L.; Cantrell, C.L.; Duke, S.O.; Van Dijck, P.; Desideri, A.; Rosa, C.A. Fungi in antarctica: Diversity, ecology, effects of climate change, and bioprospection for bioactive compounds. In Fungi of Antarctica: Diversity, Ecology and Biotechnological Applications; Rosa, L.H., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–17. [Google Scholar]
- Constable, A.J.; Meredith, M.P.; Ducklow, H.W.; Murphy, E.J.; Linse, K.; Kawaguchi, S. Impacts and effects of ocean warming on antarctic ecosystems and species. In Explaining Ocean Warming: Causes, Scale, Effects and Consequences; Laffoley, D., Baxter, J.M., Eds.; IUCN: Gland, Switzerland, 2016; p. 337. [Google Scholar]
- Danovaro, R.; Rastelli, E.; Corinaldesi, C.; Tangherlini, M.; Dell’Anno, A. Marine archaea and archaeal viruses under global change [version 1; peer review: 2 approved]. F1000Research 2017, 6, 1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danovaro, R.; Corinaldesi, C.; Dell’Anno, A.; Rastelli, E. Potential impact of global climate change on benthic deep-sea microbes. FEMS Microbiol. Lett. 2017, 364, fnx214. [Google Scholar] [CrossRef]
- Evans, C.; Brandsma, J.; Meredith, M.P.; Thomas, D.N.; Venables, H.J.; Pond, D.W.; Brussaard, C.P.D. Shift from carbon flow through the microbial loop to the viral shunt in coastal antarctic waters during austral summer. Microorganisms 2021, 9, 460. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.R.; Raymond, B.; Bracegirdle, T.J.; Chadès, I.; Fuller, R.A.; Shaw, J.D.; Terauds, A. Climate change drives expansion of antarctic ice-free habitat. Nature 2017, 547, 49–54. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Amplicon Sequencing | Shotgun Sequencing | |
|---|---|---|
| Advantages | – Can work with small sample volumes | – Very high resolution (up to SNPs) |
| – Results are focused on a single kingdom | – Results for all DNA in the sample | |
| – Cheap | ||
| Disadvantages | – Smaller taxonomic resolution | – Requires larger sample volumes |
| – Results restricted to a single kingdom | – Results for a single kingdom are harder to distinguish | |
| – Variations in the number of amplicons as a result of PCR amplification or simply the nature of the organism | – Expensive for the time being |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doytchinov, V.V.; Dimov, S.G. Microbial Community Composition of the Antarctic Ecosystems: Review of the Bacteria, Fungi, and Archaea Identified through an NGS-Based Metagenomics Approach. Life 2022, 12, 916. https://doi.org/10.3390/life12060916
Doytchinov VV, Dimov SG. Microbial Community Composition of the Antarctic Ecosystems: Review of the Bacteria, Fungi, and Archaea Identified through an NGS-Based Metagenomics Approach. Life. 2022; 12(6):916. https://doi.org/10.3390/life12060916
Chicago/Turabian StyleDoytchinov, Vesselin V., and Svetoslav G. Dimov. 2022. "Microbial Community Composition of the Antarctic Ecosystems: Review of the Bacteria, Fungi, and Archaea Identified through an NGS-Based Metagenomics Approach" Life 12, no. 6: 916. https://doi.org/10.3390/life12060916