
Potential Effect of DIMBOA (2,4-Dihydroxy-7-methoxy-1,4-benzoxazin-3-one) on Alleviating the Autotoxic Coumarin Stress in Alfalfa (Medicago sativa) Seedlings
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Coumarin and DIMBOA Treatment Solution
2.2. Effects of Coumarin and DIMBOA Treatment on Seed Germination
2.3. Effects of Coumarin and DIMBOA Treatment on Photosynthetic Physiology, Root Morphology, andAntioxidant Physiology of Seedlings
2.4. Measurement Indicators and Methods
2.4.1. Measurement of Photosynthetic Parameters
2.4.2. Measurement of Chlorophyll
2.4.3. Measurement of Root Morphological Characteristics
2.4.4. Measurement of MDA and ROS (H2O2, OH• and O2•−) Content
2.4.5. Measurement of Antioxidant Enzymes Activities and Antioxidant Content
2.4.6. Measurement of Soluble Protein and Soluble Sugar Content
3. Statistical Analysis
4. Results
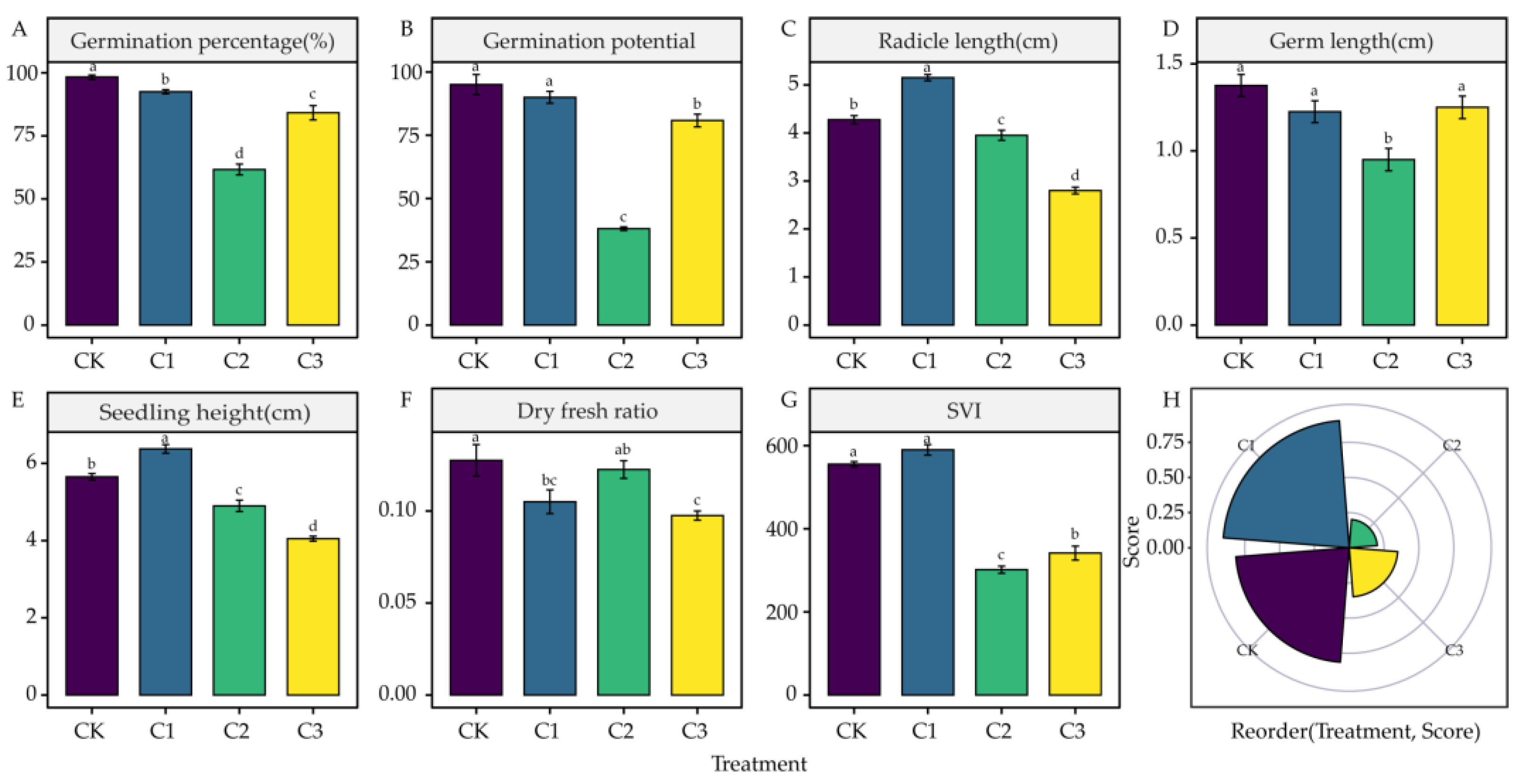
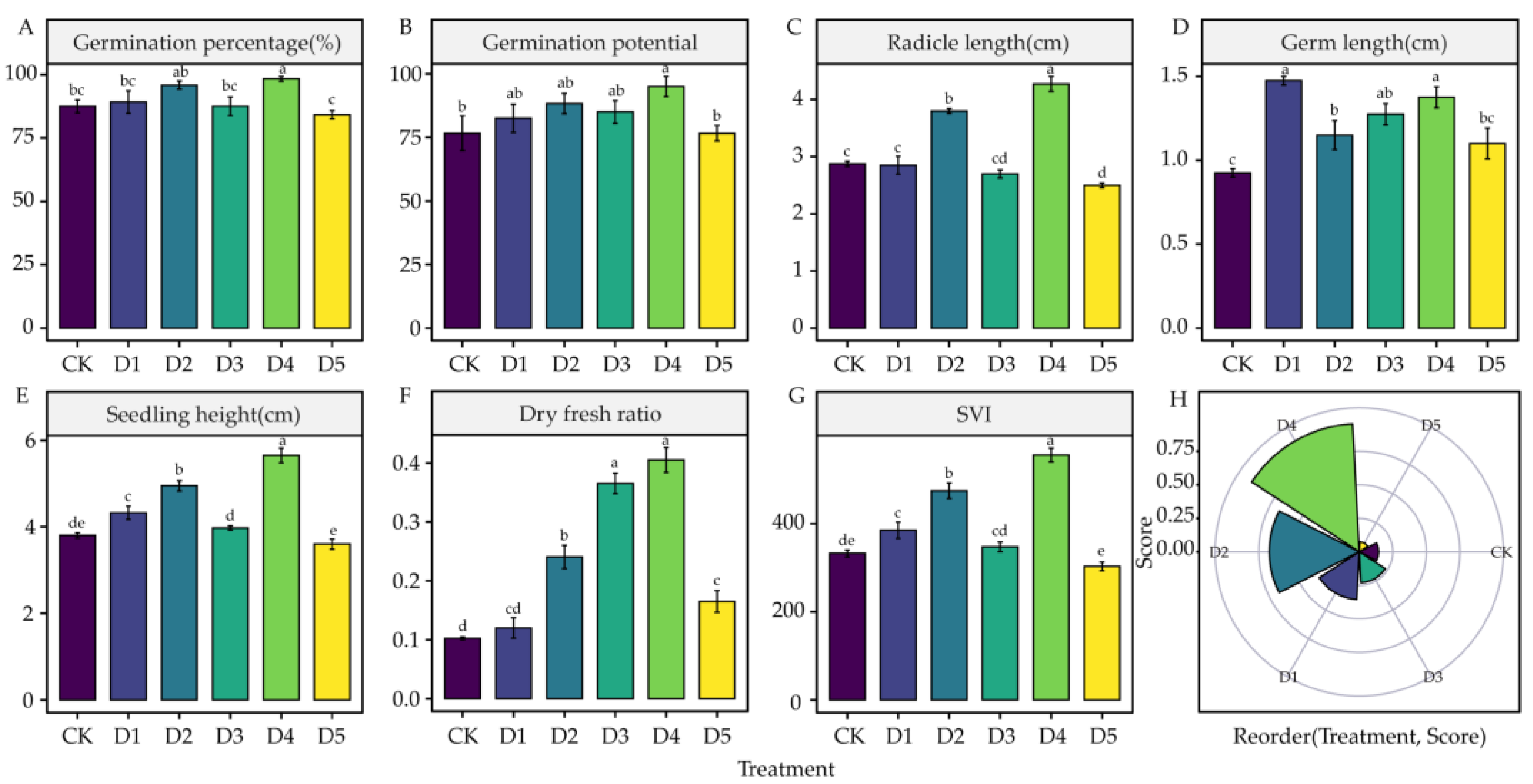
4.1. Effects of DIMBOA and Coumarin on Alfalfa Seed Germination
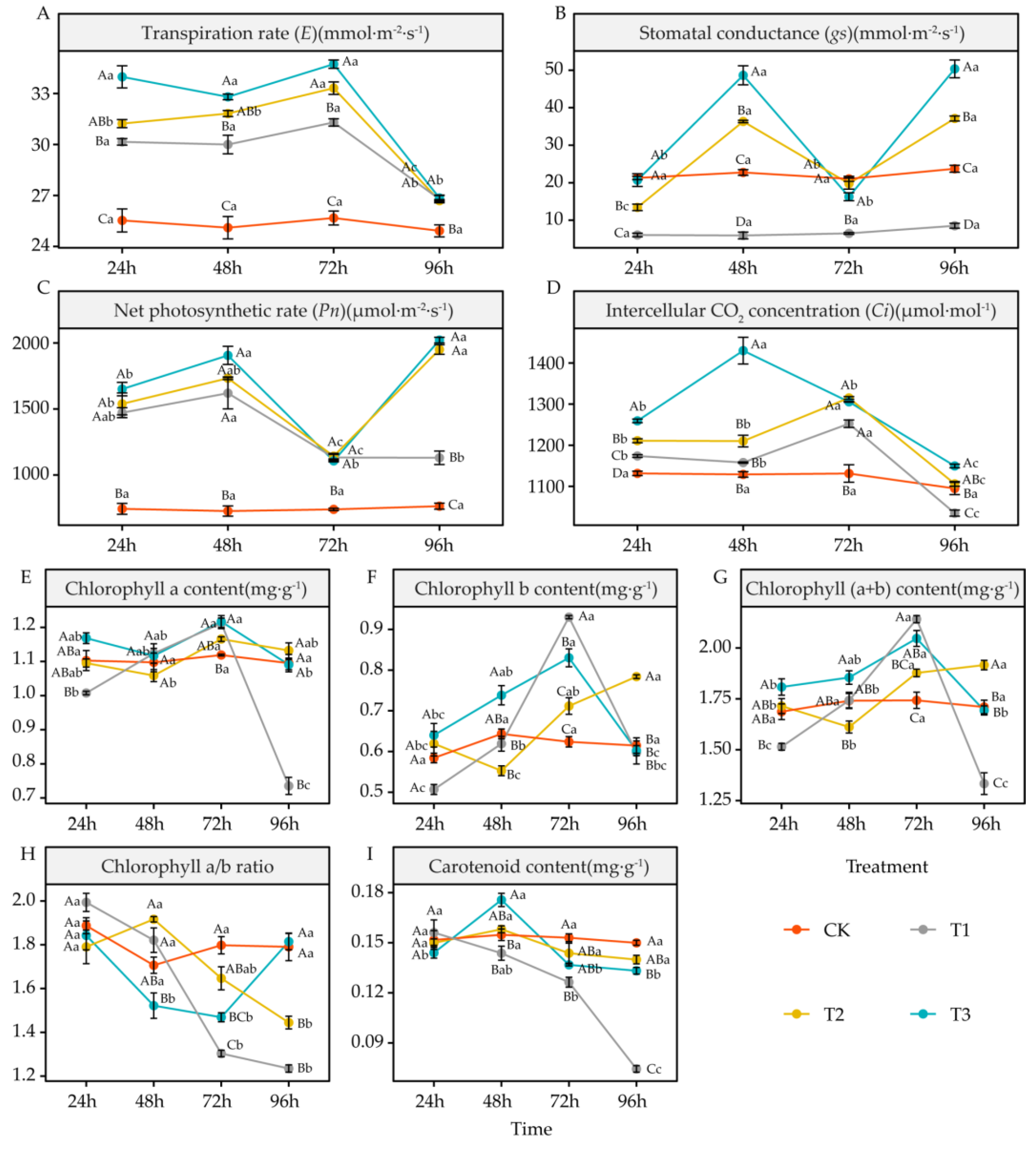
4.2. Effects of DIMBOA and Coumarin on Photosynthetic Parameter Activities of Alfalfa
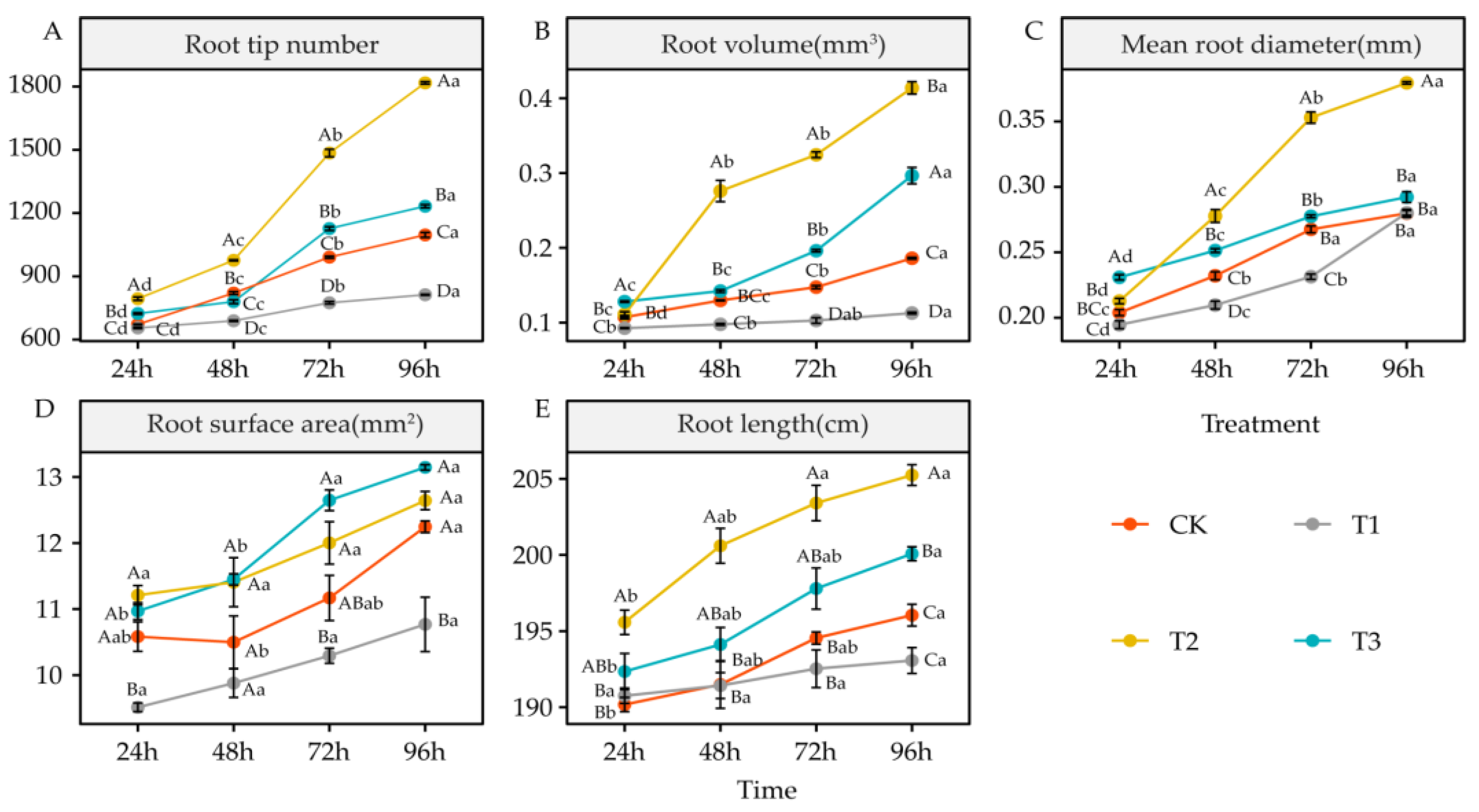
4.3. Effects of DIMBOA and Coumarin on Root Morphological Characteristics of Alfalfa
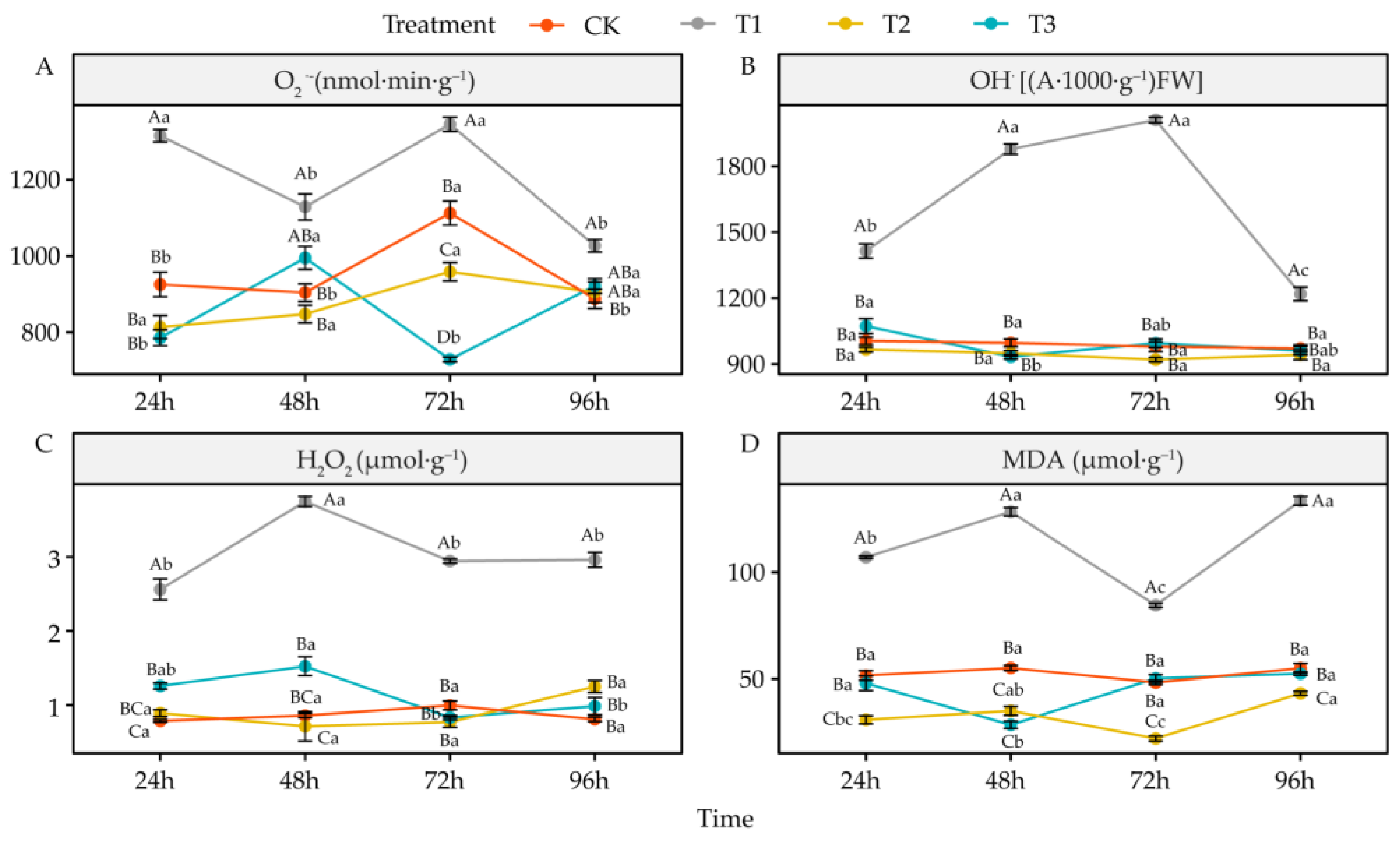
4.4. Effects of DIMBOA and Coumarin on ROS and Malondialdehyde of Alfalfa
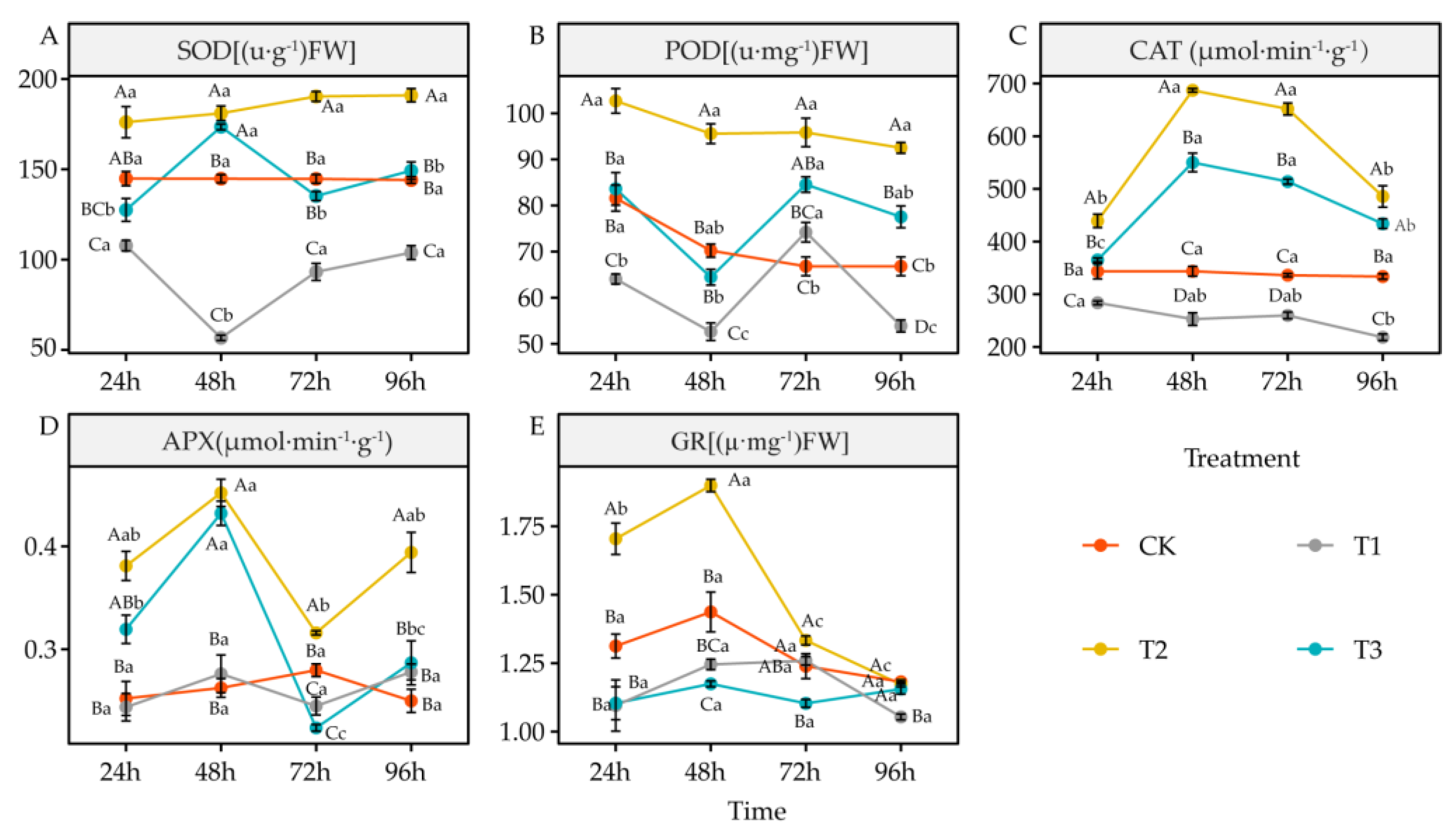
4.5. Effects of DIMBOA and Coumarin on Antioxidant Enzymes Activities of Alfalfa
4.6. Effects of DIMBOA and Coumarin on Antioxidants of Alfalfa
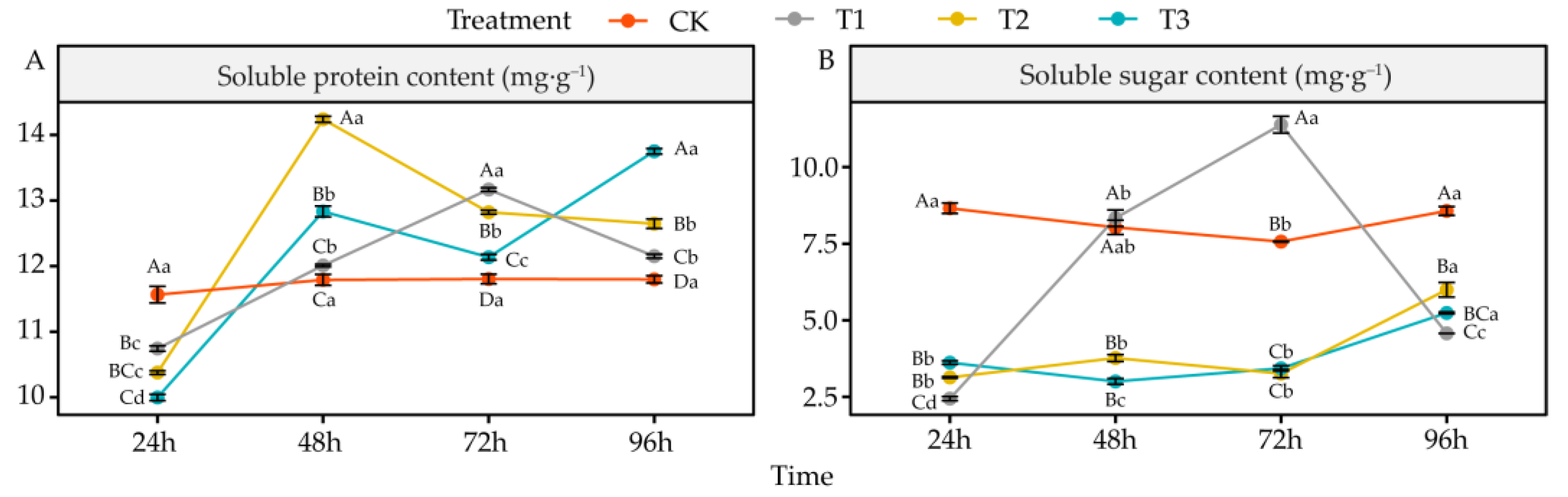
4.7. Effects of DIMBOA and Coumarin on Soluble Protein and Soluble Sugar Content of Alfalfa
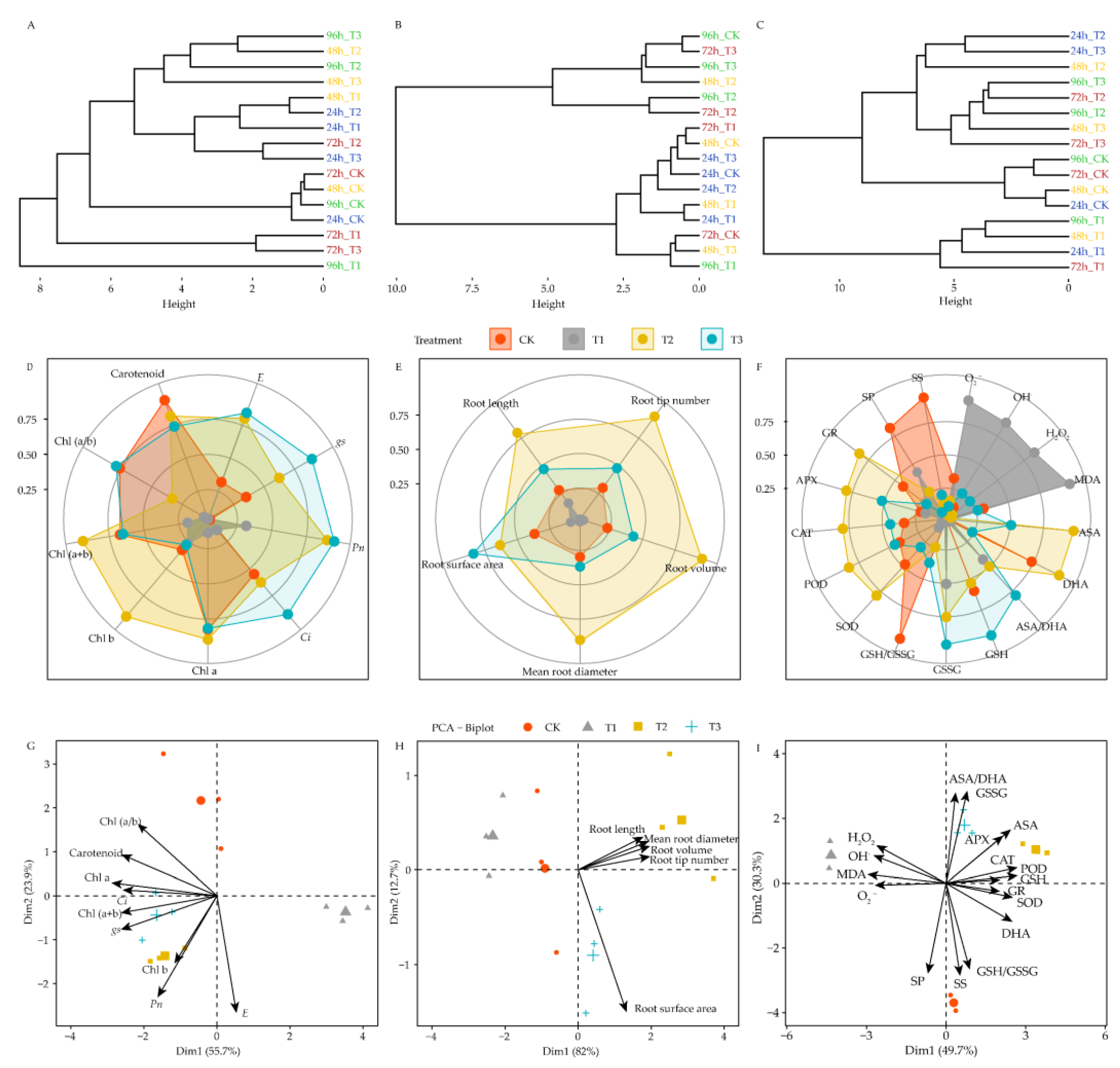
4.8. Effects of DIMBOA and Coumarin on Photosynthetic, Root Characteristics, and Physiological Indicators of Stress Resistance of Alfalfa
5. Discussion
5.1. Effects of DIMBOA and Coumarin on Alfalfa Seed Germination
5.2. Effects of DIMBOA and Coumarin on Photosynthetic Parameters of Alfalfa
5.3. Effects of DIMBOA and Coumarin on Root Characteristics of Alfalfa
5.4. Alteration of Oxidative Damage in Alfalfa under DIMBOA and Coumarin
5.5. Alteration of Antioxidant Enzymes Activities and Antioxidant in Alfalfa under DIMBOA and Coumarin
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chen, H.T.; Zeng, Y.; Yang, Y.Z.; Huang, L.L.; Tang, B.L.; Zhang, H.; Hao, F.; Liu, W.; Li, Y.H.; Liu, Y.B.; et al. Allele-aware chromosome-level genome assembly and efficient transgene-free genome editing for the autotetraploid cultivated alfalfa. Nat. Commun. 2020, 11, 2494. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, T.; Ma, L.; Zhao, Z.; Zhao, P.; Nan, Z.; Wang, Y. Global transcriptome sequencing using the Illumina platform and the development of EST-SSR markers in autotetraploid alfalfa. PLoS ONE 2013, 8, e83549. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, B.K.; Ghimire, B.; Yu, C.Y.; Chung, I.M. Allelopathic and Autotoxic Effects of Medicago sativa-Derived Allelochemicals. Plants 2019, 8, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennings, J.A.; Nelson, C.J. Zone of autotoxic influence around established alfalfa plants. Agron. J. 2002, 70, 21–26. [Google Scholar] [CrossRef]
- Abdulrahman, A.A.; Habib, S.A. Allelopathic effect of alfalfa (Medicago sativa) on bladygrass (Imperata cylindrica). J. Chem. Ecol. 1989, 15, 2289–2300. [Google Scholar] [CrossRef]
- Razavi, S.M. Plant coumarins as allelopathic agents. Int. J. Biol. Chem. 2011, 5, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Hedge, R.S.; Miller, D.A. Concentration dependency and stage of crop growth in alfalfa autotoxicity. Agron. J. 1992, 84, 940–946. [Google Scholar]
- Chon, S.U.; Choi, S.K.; Jung, S.; Jang, H.G.; Pyo, B.S.; Kim, S.M. Effects of alfalfa leaf extracts and phenolic allelochemicals on early seedling growth and root morphology of alfalfa and barnyard grass. Crop Prot. 2002, 21, 1077–1082. [Google Scholar] [CrossRef]
- Aliotta, G.; Cafeoro, G.; Fiorentino, A.; Strumia, S. Inhibition of radish germination and root growth by Coumarin and Phe-nylpropanoids. J. Chem. Ecol. 1993, 19, 175–183. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; De, S.C.; Sidari, M.; Sorgonà, A.; Badiani, M.; Cacco, G. Influence of coumarin on the net nitrate uptake in durum wheat. New Phytol. 2001, 150, 227–237. [Google Scholar] [CrossRef]
- EL-Shahawy, T.A.; Abdelhamid, M.T. Potential allelopathic effect of six phaseolus vulgaris recombinant inbred lines for weed control. AJBAS 2013, 7, 462–467. [Google Scholar]
- Bais, H.P.; Prithiviraj, B.; Jha, A.K.; Ausubel, F.M.; Vivanco, J.M. Mediation of pathogen resistance by exudation of antimicrobials from roots. Nature 2005, 434, 217–221. [Google Scholar] [CrossRef]
- Sun, P.; Yuuko, T.; Hideyuki, M.; Yoshihara, T. Antifungal Compounds from the Root and Root Exudate of Zea mays. Biosci. Biotechnol. Biochem. 2004, 68, 1366–1368. [Google Scholar]
- Miller, D.A. Allelopathy in forage crop system. Agron. J. 1996, 88, 854–859. [Google Scholar] [CrossRef]
- Niemeyer, H.M. Hydroxamic acids (4-Hydroxy-1,4-benzoxazin-3-ones) defence chemicals in the gramineae. Phytochemistry 1988, 27, 3349–3358. [Google Scholar] [CrossRef]
- Richardson, M.D.; Bacon, C.W. Catabolism of 6-methoxy-benzoxazolinone and benzoxazolinone by Fusarium moniliforme. Mycologia 1995, 87, 510–517. [Google Scholar] [CrossRef]
- Feng, M.Y.; Chong, R.X.; Song, G.L.; Lin, C.S.; Li, J.H. Effects of DIMBOA on several enzymatic systems in Asian corn borer, Ostrinia furnacalis (Guenee). J Chem. Ecol. 1995, 21, 2047–2056. [Google Scholar]
- Thackray, D.J.; Wratien, S.D.; Edwards, P.J.; Niemeyer, H.M. Resistance to the aphids Sitobion avenae and Rhopalosiphum padi in Gramineae in relation to hydroxamic levels. Ann. Appl. Biol. 1990, 116, 573–582. [Google Scholar] [CrossRef]
- Schulz, M.; Marocco, A.; Tabaglio, V.; Macias, F.A.; Molinillo, J. Benzoxazinoids in Rye Allelopathy From Discovery to Ap-plication in Sustainable Weed Control and Organic Farming. J. Chem. Ecol. 2013, 39, 154–174. [Google Scholar] [CrossRef]
- Villagrasa, M.; Eljarrat, E.; Barcelὁ, D. Analysis of benzoxazinone derivatives in plant tissues and their degradation products in agricultural soils. Trends Analyt. Chem. 2009, 28, 1103–1114. [Google Scholar] [CrossRef]
- Wu, H.W.; Haig, T.; Pratley, J.; Lemerle, D.; An, M. Distribution and exudation of allelochemocals in wheat Triticum aestivum. J. Chem. Ecol. 2000, 26, 2141–2154. [Google Scholar] [CrossRef]
- Perez, F.J. Allelopathic effect of hydroxamic acids from cereals on Avena sativa and A. Fatua. Phytochemistry 1990, 29, 773–776. [Google Scholar] [CrossRef]
- Chen, K.J.; Zheng, Y.Q.; Kong, C.H.; Zhang, S.Z.; Li, J.; Liu, X.G. 2,4-Dihydroxy-7-methoxy-1,4-benzoxazin-3-one(DIMBOA) and 6- methoxy-benzoxazolin-2-one(MBOA) levels in the wheat rhizosphere and their effect on the soil microbial community structure. J. Agric. Food Chem. 2010, 58, 12710–12716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Shi, S.L.; Li, X.L.; Li, C.N.; Zhang, C.M.; Yun, A.; Kang, W.J.; Yin, G.L. Effects of Autotoxicity on Alfalfa (Medicago sativa): Seed Germination, Oxidative Damage and Lipid Peroxidation of Seedlings. Agronomy 2021, 11, 1027. [Google Scholar] [CrossRef]
- International Seed Testing Association (ISTA). International Rules for Seed Testing; ISTA Press: Basserdorf, Switzerland, 2012. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 357–359. [Google Scholar]
- Pallavi, S.; Bhushan, J.A.; Dubey, R.S.; Shanker, D.R.; Mohammad, P. Reactive oxygen species, oxidative damage, and anti-oxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar]
- von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid peroxidation. Meth. Enzymol. 1990, 186, 421–431. [Google Scholar]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Montagu, M.V.; Inzé, D.; Camp, W.V. Catalase is a sink for H2O2 and is indispensable for stress de-fence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Zhao, Z.G.; Si, J.; Di, C.X.; Han, J.; An, L.Z. Brassinosteroids alleviate chilling-induced oxidative damage by en-hancing anti-oxidant defense system in suspension cultured cells of Chorispora bungeana. Plant Growth Regul. 2009, 59, 207–214. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Formation of hydrogen peroxide by isolated cell walls from horseradish (Armoracia lapathifolia Gilib.). Planta 1976, 130, 175–180. [Google Scholar] [CrossRef]
- De Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; Medeiros, J.R.; Gomes-Filho, E. Hydrogen peroxide pre-treatment induces salt-stress acclimation in maize plants. J. Plant Physiol. 2005, 162, 1114–1122. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Havir, E.A.; Mchale, N.A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Murshed, R.; Lopezlauri, F.; Sallanon, H. Microplate quantification of enzymes of the plant ascorbate-glutathione cycle. Anal. Biochem. 2008, 383, 320–322. [Google Scholar] [CrossRef]
- Murshed, R.; Lopezlauri, F.; Sallanon, H. Effect of water stress on antioxidant systems and oxidative parameters in fruits of tomato (Solanum lycopersicon L. cv. Microtom). Physiol. Mol. Biol. Plants 2013, 19, 363–378. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Buysse, J.; Merckx, R. An improved colorimetric method to quantify sugar content of plant tissue. J. Exp. Bot. 1993, 44, 1627–1629. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Miller, D.A. Allelopathic influence of nine for age grass extracts on germination and seedling growth of alfalfa. Agron. J. 1995, 87, 767–772. [Google Scholar] [CrossRef]
- Hall, M.H.; Henderlong, P.R. Alfalfa autotoxic fraction characterization and initial separation. Crop Sci. 1989, 29, 425–428. [Google Scholar] [CrossRef]
- Dornbos, D.L.; Spencer, G.F.; Miller, R.W. Medicarpin delays alfalfa seed germination and seedling growth. Crop Sci. 1990, 30, 162–166. [Google Scholar] [CrossRef]
- Liu, J.P.; Zhang, X.Q.; Luo, B.; You, M.H. The Effect of Lixivium of Limpograss on Growth of Legumina Forages. Seed 2005, 24, 5–8. [Google Scholar]
- Miller, D.A. Allelopathie effects of alfalfa. Chem. Ecol. 1983, 9, 1059–1072. [Google Scholar] [CrossRef]
- Wang, J.C.; Pan, K.W.; Wu, N.; Luo, P.; Li, F.H. The study on varietal differences in allelopathic potential of Chinese prickly ash (Zanthoxylum piperitum). Acta Ecol. Sin. 2005, 25, 1591–1598. [Google Scholar]
- Mao, M.H.; Yu, F.L.; Kong, L.S. Allelopathy effects of Artemisia ordosica. Acta Ecol. Sin. 1999, 19, 670–676. [Google Scholar]
- Yan, F.; Liang, X.; Zhu, X. The role of DIMBOA on the feeding of Asian corn borer, Ostrinia furnacalis (Guenee) (Lep. Pyralidae). J. Appl. Entomol. 2010, 123, 49–53. [Google Scholar] [CrossRef]
- Calabrese, R.M.; Baldwin, L.A. Hormesis: The dose-response revolution. Annu. Rev. Pharmacol. 2003, 42, 175–197. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.M.; Kim, K.H.; Ahn, J.K.; Chun, S.C.; Kim, C.S.; Kim, J.T.; Kim, S.H. Screening of allelochemicals on barnyard grass (Echinochloa crusgalli) and identification of potentially allelopathic compounds from rice (Oryza sativa) variety hull extracts. Crop Prot. 2002, 21, 913–920. [Google Scholar] [CrossRef]
- Rokem, J.S.; Lantz, A.E.; Nielsen, J. Systems biology of antibiotic production by microorganisms. Nat. Prod. Rep. 2007, 24, 1262–1287. [Google Scholar] [CrossRef]
- Debabrata, P.; Sharma, S.G.; Sarkar, R.K. Chlorophy II fluorescence parameters, CO2 photosynthetic rate and regeneration capacity as a result of complete submergence and subsequent re-emergence in rice (Oryza sativa L.). Aquat. Bot. 2008, 88, 127–133. [Google Scholar]
- Paridaak; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Hao, S.X.; Cao, H.X.; Wang, H.B.; Pan, X.Y. The physiological responses of tomato to water stress and re-water in different growth periods. Sci. Hortic. 2019, 249, 143–154. [Google Scholar] [CrossRef]
- Zhang, B.B.; Zhang, H.; Jing, Q.; Wang, J.X. Light pollution on the growth, physiology and chlorophyll fluorescence response of landscape plant perennial ryegrass (Lolium perenne L.). Ecol. Indic. 2020, 115, 106448. [Google Scholar] [CrossRef]
- Eunbhlling, F.A.; Rasmussen, J.A.; Schon, M.K. Effects of three phenolic acids on chlorophyll content and growth of soybean and grain sorghum seedlings. J. Chem. Ecol. 1979, 5, 815–824. [Google Scholar]
- Cheng, Z.H. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2015, 6, 1020. [Google Scholar]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils: A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Zhou, B.; Kong, C.H.; Li, Y.H. Chemical constituents of wild oat and crabgrass essential oils and their effects on the growth and allelochemical production of wheat. Weed Biol. Manag. 2013, 13, 62–69. [Google Scholar] [CrossRef]
- Li, W.R.; Zhang, S.Q.; Lun, S.; Eneji, A. Changes in root characteristics, gas exchange and water use efficiency following water stress and rehydration of alfalfa and sorghum. Aust. J. Crop Sci. 2011, 5, 1521–1532. [Google Scholar]
- Vaughan, D.; Ord, B. Influence of phenolic acids on morphological changes in roots of Pisum sativum. J. Sci. Food Agric. 2010, 52, 289–299. [Google Scholar] [CrossRef]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Sun, Y.; Xiao, C.L.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Physiological basis of different allelopathic reactions of cucumber and figleaf gourd plants to cinnamic acid. J. Exp. Bot. 2007, 58, 3765–3773. [Google Scholar] [CrossRef] [PubMed]
- Weismann, D.; Hartvigsen, K.; Lauer, N.; Bennett, K.L.; Scholl, H.P.N.; Charbel, P.I.; Cano, M.; Brandstätter, H.; Tsimikas, S.; Skerka, C.; et al. Complement factor H binds malondialdehyde epitopes and protects from oxidative stress. Nature 2011, 478, 76–81. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Shulaev, V.; Breusegem, F.V. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascor-bate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, A.; Davis, T.D.; Walser, R.H. Uniconazole-induced alleviation of low-temperature damage in relation to antiox-idant activity. Hortscience 1989, 24, 955–957. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Setia, N.; Kaur, S.; Kohli, R.K. 2-Benzoxazolinone (BOA) induced oxidative stress, lipid and perox-idation and changes in some antioxidant enzyme activities in mung bean (Phaseolus aureus). Plant Physiol. Biochem. 2006, 44, 819–827. [Google Scholar] [CrossRef]
- Hong, Y.; Hu, H.Y.; Xie, X.; Li, F.M. Responses of enzymatic antioxidants and non-enzymatic antioxidantsin the cyanobacte-rium Microcystis aeruginosa to the allelochemical ethyl 2-methyl acetoacetate (EMA) isolated from reed (Phragmites communis). J. Plant Physiol. 2008, 165, 1264–1273. [Google Scholar] [CrossRef]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef]
- Paradies, G.; Petrosillo, G.; Paradies, V.; Reiter, R.J.; Ruggiero, F.M. Melatonin, cardiolipin and mitochondrial bioenergetics in health and disease. J. Pineal Res. 2010, 48, 297–310. [Google Scholar] [CrossRef]
- Spencer, C.M.; Cai, Y.; Martin, R.; Gaffney, S.H.; Goulding, P.N.; Magnolato, D.; Lilley, T.H.; Haslam, E. Polyphenol complexation: Some thoughts and observations. Phytochemistry 1988, 27, 2397–2409. [Google Scholar] [CrossRef]
- Han, Z.P.; Guo, S.R.; Jiao, Y.S. Effect of NaCl stress on growth and photosynthetic gas exchange of watermelon seedlings. Acta Bot. Boreali-Occident. Sin. 2008, 28, 745–751. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Information | Coumarin | DIMBOA |
|---|---|---|
| Catalog No. | ZC-53633 | ZTR-D459950 |
| CAS No. | 91-64-5 | 15893-52-4 |
| Molecular Formula | C9H6O2 | C9H9NO5 |
| Molecular Weight | 146.14 | 211.17 |
| Storage | 0–8 °C | −20 °C |
| Purity | 99.97% (HPLC) | 95% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Shi, S.; Zhang, X.; Li, C.; Wang, H.; Kang, W.; Yin, G. Potential Effect of DIMBOA (2,4-Dihydroxy-7-methoxy-1,4-benzoxazin-3-one) on Alleviating the Autotoxic Coumarin Stress in Alfalfa (Medicago sativa) Seedlings. Life 2022, 12, 2140. https://doi.org/10.3390/life12122140
Li X, Shi S, Zhang X, Li C, Wang H, Kang W, Yin G. Potential Effect of DIMBOA (2,4-Dihydroxy-7-methoxy-1,4-benzoxazin-3-one) on Alleviating the Autotoxic Coumarin Stress in Alfalfa (Medicago sativa) Seedlings. Life. 2022; 12(12):2140. https://doi.org/10.3390/life12122140
Chicago/Turabian StyleLi, Xiaolong, Shangli Shi, Xiaoyan Zhang, Changning Li, Huning Wang, Wenjuan Kang, and Guoli Yin. 2022. "Potential Effect of DIMBOA (2,4-Dihydroxy-7-methoxy-1,4-benzoxazin-3-one) on Alleviating the Autotoxic Coumarin Stress in Alfalfa (Medicago sativa) Seedlings" Life 12, no. 12: 2140. https://doi.org/10.3390/life12122140