
Emergence and Dissemination of Extraintestinal Pathogenic High-Risk International Clones of Escherichia coli


Abstract
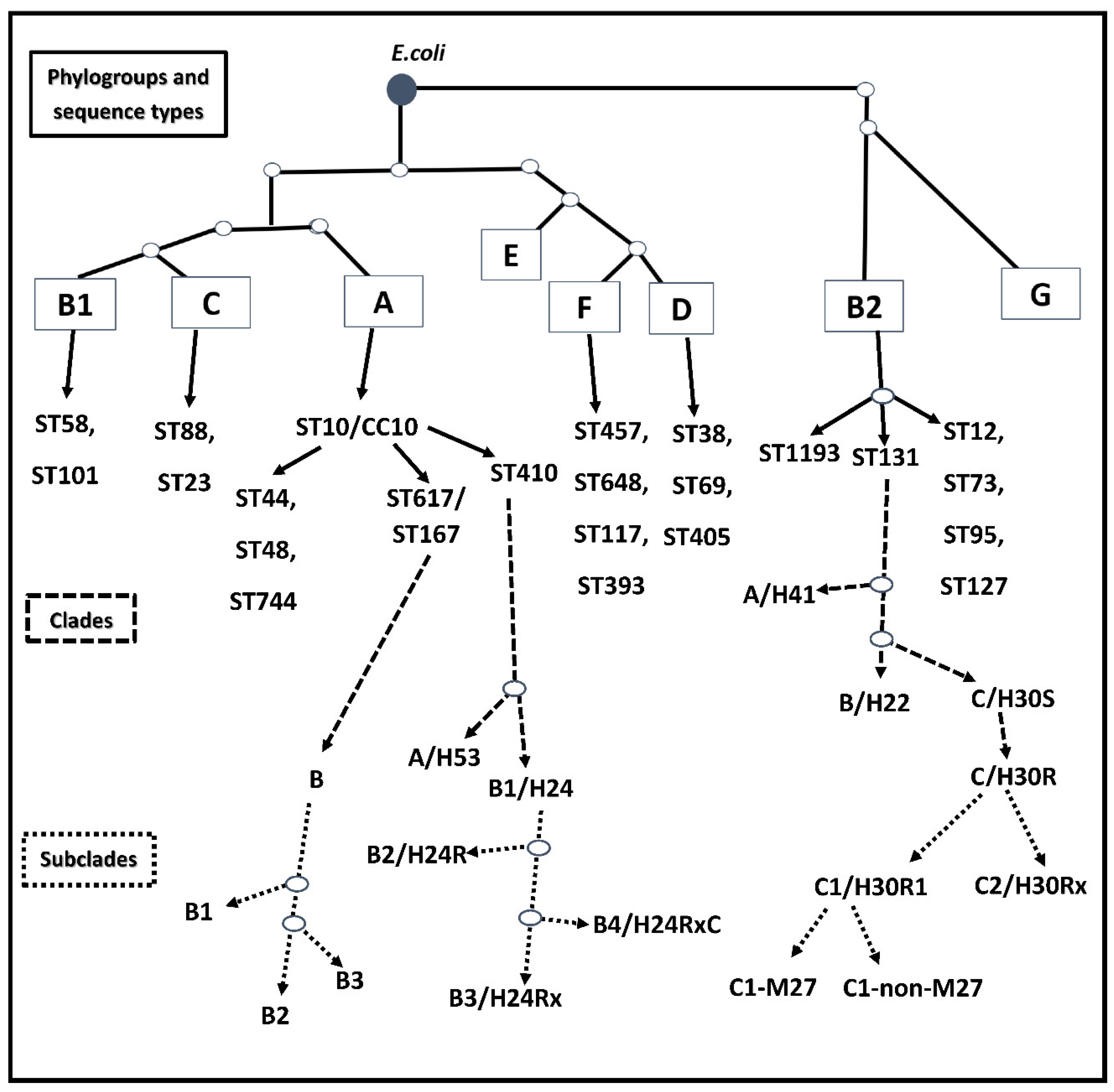
:1. Introduction
2. CC/ST131, the Worldwide Predominant High-Risk Clone
3. ST1193, a Recently Emerging Pandemic MDR Clone from Phylogroup B2
4. ST69 and CC10, the Second and Third Most Common High-Risk Clones
5. ST405 and High-Risk CC/ST38 Clones from Phylogenetic Group D
6. ST457, a Novel Emerging Clone from Phylogroup F
7. Further Potential ExPEC High-Risk Clones
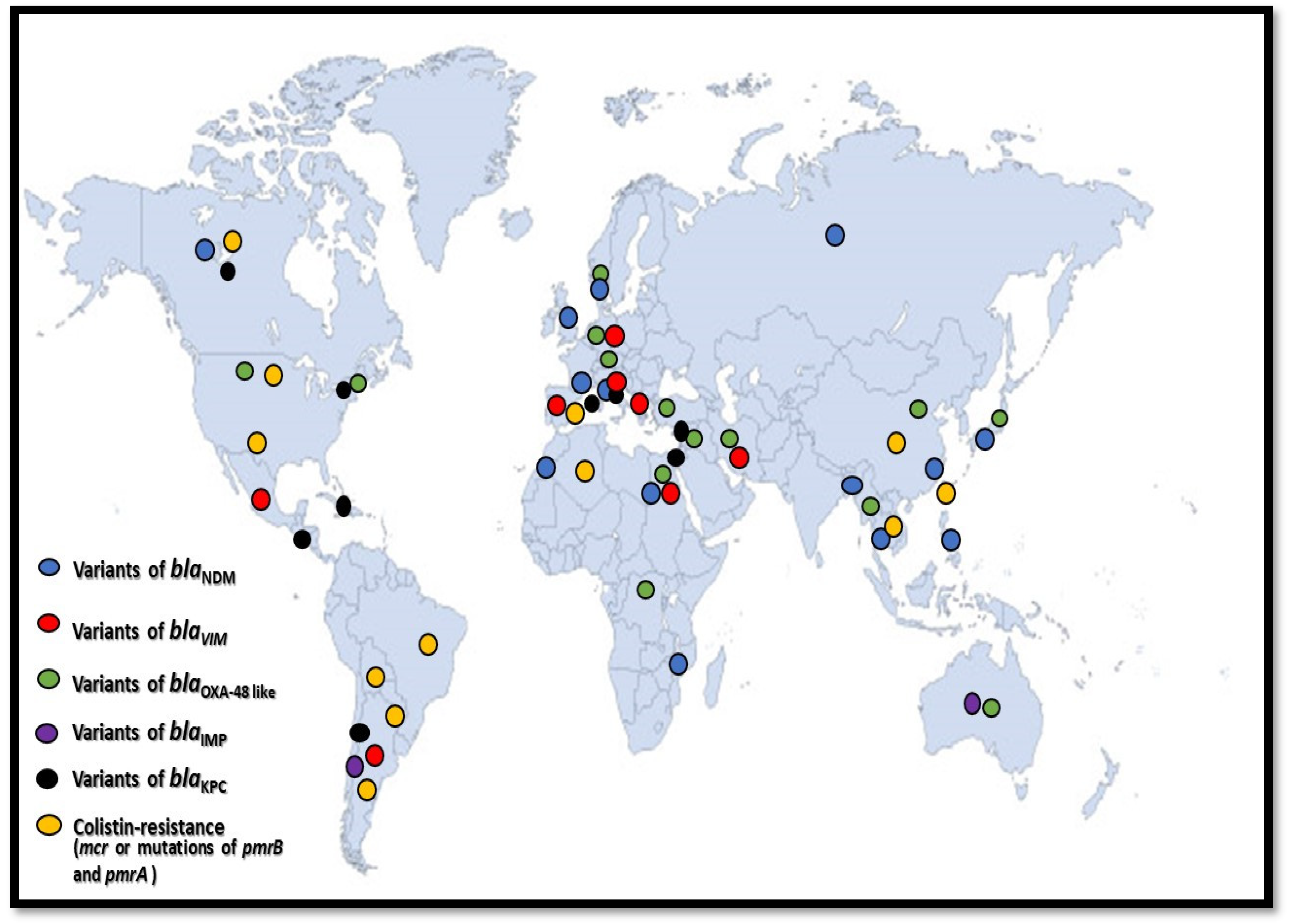
8. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pitout, J.D.D. Extraintestinal pathogenic Escherichia coli: An update on antimicrobial resistance, laboratory diagnosis and treatment. Expert Rev. Anti-Infect. Ther. 2012, 10, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M.; Kárpáti, K.; Nagy, Á.L.; Gugolya, M.; Stájer, A.; Burián, K. Association between biofilm-production and antibiotic resistance in Escherichia coli isolates: A laboratory-based case study and a literature review. Acta Microbiol. Immunol. Hung. 2021, 68, 217–226. [Google Scholar] [CrossRef]
- Livermore, D.M. Has the era of untreatable infections arrived? J. Antimicrob. Chemother. 2009, 64 (Suppl. 1), i29–i36. [Google Scholar] [CrossRef] [Green Version]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Cummins, E.A.; Snaith, A.E.; McNally, A.; Hall, R.J. The role of potentiating mutations in the evolution of pandemic Escherichia coli clones. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mathers, A.J.; Peirano, G.; Pitout, J.D.D. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Zhu, J.; Gong, D.; Wu, L.; Zhu, Y.; Hu, L. Whole genome sequence of EC16, a blaNDM-5-, blaCTX-M-55-, and fosA3-coproducing Escherichia coli ST167 clinical isolat from China. J. Glob. Antimicrob. Resist. 2022, 29, 296–298. [Google Scholar] [CrossRef]
- Baquero, F.; Tedim, A.-S.P.; Coque, T.M. Antibiotic resistance shaping multi-level population biology of bacteria. Front. Microbiol. 2013, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Doi, Y. Treatment options for carbapenem-resistant gram-negative bacterial infections. Clin. Infect. Dis. 2019, 69, S565–S575. [Google Scholar] [CrossRef] [Green Version]
- Mazzariol, A.; Bazaj, A.; Cornaglia, G. Multi-drug-resistant Gram-negative bacteria causing urinary tract infections: A review. J. Chemother. 2017, 29, 2–9. [Google Scholar] [CrossRef]
- Adekanmbi, A.O.; Usidamen, S.; Onilude, A.A. Molecular characterization of ESBL- producing uropathogenic Escherichia coli recovered from urine samples of patients attending a University Teaching hospital in Nigeria. Acta Microbiol. Immunol. Hung. 2021, 68, 169–176. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L. The difficult-to-control spread of carbapenemase producers among Enterobacteriaceae worldwide. Clin. Microbiol. Infect. 2014, 20, 821–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordmann, P.; Poirel, L. Epidemiology and diagnostics of carbapenem resistance in Gram-negative bacteria. Clin. Infect. Dis. 2019, 69, S521–S528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Pitout, J.D.D.; DeVinney, R. Escherichia coli ST131: A multidrug-resistant clone primed for global domination. F1000Research 2017, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Van der Bij, A.K.; Peirano, G.; Pitondo-Silva, A.; Pitout, J.D. The presence of genes encoding for different virulence factors in clonally related Escherichia coli that produce CTX-Ms. Diagn. Microbiol. Infect. Dis. 2012, 72, 297–302. [Google Scholar] [CrossRef]
- Johnston, B.D.; Thuras, P.; Porter, S.B.; Anacker, M.; VonBank, B.; Vagnone, P.S.; Witwer, M.; Castanheira, M.; Johnson, J.R. Global molecular epidemiology of carbapenem-resistant Escherichia coli (2002–2017). Eur. J. Clin. Microbiol. 2021, 1–13. [Google Scholar] [CrossRef]
- Liu, C.M.; Stegger, M.; Aziz, M.; Johnson, T.J.; Waits, K.; Nordstrom, L.; Gauld, L.; Weaver, B.; Rolland, D.; Statham, S.; et al. Escherichia coli ST131-H22 as a foodborne uropathogen. mBio 2018, 9, e00470-18. [Google Scholar] [CrossRef] [Green Version]
- Nesporova, K.; Wyrsch, E.R.; Valcek, A.; Bitar, I.; Chaw, K.; Harris, P.; Hrabak, J.; Literak, I.; Djordjevic, S.P.; Dolejska, M. Escherichia coli Sequence Type 457 Is an Emerging Extended-Spectrum-β-Lactam-Resistant Lineage with Reservoirs in Wildlife and Food-Producing Animals. Antimicrob. Agents Chemother. 2020, 65, e01118-20. [Google Scholar] [CrossRef]
- Sanz, M.B.; De Belder, D.; de Mendieta, J.M.; Faccone, D.; Poklepovich, T.; Lucero, C.; Rapoport, M.; Campos, J.; Tuduri, E.; Saavedra, M.O.; et al. Carbapenemase-Producing Extraintestinal Pathogenic Escherichia coli From Argentina: Clonal Diversity and Predominance of Hyperepidemic Clones CC10 and CC131. Front. Microbiol. 2022, 13, 830209. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S.; Paterson, D.L. Antibiotics in the clinical pipeline in October 2019. J. Antibiot. 2020, 73, 329–364. [Google Scholar] [CrossRef] [PubMed]
- Yahav, D.; Giske, C.G.; Grāmatniece, A.; Abodakpi, H.; Tam, V.H.; Leibovici, L. New β-Lactam-β-Lactamase Inhibitor Combinations. Clin. Microbiol. Rev. 2020, 34, e00115-20. [Google Scholar] [CrossRef]
- Kocsis, B.; Gulyás, D.; Szabó, D. Delafloxacin, Finafloxacin, and Zabofloxacin: Novel Fluoroquinolones in the Antibiotic Pipeline. Antibiotics 2021, 10, 1506. [Google Scholar] [CrossRef] [PubMed]
- Scudeller, L.; Righi, E.; Chiamenti, M.; Bragantini, D.; Menchinelli, G.; Cattaneo, P.; Giske, C.G.; Lodise, T.; Sanguinetti, M.; Piddock, L.J.V.; et al. Systematic review and meta-analysis of in vitro efficacy of antibiotic combination therapy against carbapenem-resistant Gram-negative bacilli. Int. J. Antimicrob. Agents 2021, 57, 106344. [Google Scholar] [CrossRef] [PubMed]
- Carrara, E.; Savoldi, A.; Piddock, L.J.; Franceschi, F.; Ellis, S.; Sharland, M.; Brink, A.J.; Harris, P.N.; Levy-Hara, G.; Rohit, A.; et al. Clinical management of severe infections caused by carbapenem-resistant gram-negative bacteria: A worldwide cross-sectional survey addressing the use of antibiotic combinations. Clin. Microbiol. Infect. 2022, 28, 66–72. [Google Scholar] [CrossRef]
- Roer, L.; Overballe-Petersen, S.; Hansen, F.; Johannesen, T.B.; Stegger, M.; Bortolaia, V.; Leekitcharoenphon, P.; Korsgaard, H.B.; Seyfarth, A.M.; Mossong, J.; et al. ST131 fimH22 Escherichia coli isolate with a blaCMY-2/IncI1/ST12 plasmid obtained from a patient with bloodstream infection: Highly similar to E. coli isolates of broiler origin. J. Antimicrob. Chemother. 2019, 74, 557–560. [Google Scholar] [CrossRef]
- Léger, A.; Lambraki, I.; Graells, T.; Cousins, M.; Henriksson, P.J.G.; Harbarth, S.; Carson, C.A.; Majowicz, S.E.; Troell, M.; Parmley, E.J.; et al. Characterizing social-ecological context and success factors of antimicrobial resistance interventions across the One Health spectrum: Analysis of 42 interventions targeting E. coli. BMC Infect. Dis. 2021, 21, 873. [Google Scholar] [CrossRef]
- Price, L.B.; Johnson, J.R.; Aziz, M. The epidemic of extended spectrum- β -lactamase-producing Escherichia coli ST131 is driven by a single highly pathogenic subclone. H 30-Rx. mBio 2013, 4, e00377-13. [Google Scholar] [CrossRef] [Green Version]
- Füzi, M. Dissimilar fitness associated with resistance to fluoroquinolones influences clonal dynamics of various multiresistant bacteria. Front. Microbiol. 2016, 7, 1071. [Google Scholar] [CrossRef] [PubMed]
- Johansson, V.; Nykäsenoja, S.; Myllyniemi, A.L.; Rossow, H.; Heikinheimo, A. Genomic characterization of ESBL/AmpC-producing and high-risk clonal lineages of Escherichia coli and Klebsiella pneumoniae in imported dogs with shelter and stray background. J. Glob. Antimicrob. Resist. 2022, 30, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Gladstone, R.A.; McNally, A.; Pöntinen, A.K.; Tonkin-Hill, G.; Lees, J.A.; Skytén, K.; Cléon, F.; Christensen, M.O.K.; Haldorsen, B.C.; Bye, K.K.; et al. Emergence and dissemination of antimicrobial resistance in Escherichia coli causing bloodstream infections in Norway in 2002-17: A nationwide, longitudinal, microbial population genomic study. Lancet Microbe 2021, 2, e331–e341. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, R.R.; Henderson, I.R. The evolution of the Escherichia coli phylogeny. Infect. Genet. Evol. 2012, 12, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walk, S.T.; Alm, E.W.; Gordon, D.M.; Ram, J.L.; Toranzos, G.A.; Tiedje, J.M.; Whittam, T.S. Cryptic lineages of the genus Escherichia. Appl. Environ. Microbiol. 2009, 75, 6534–6544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beghain, J.; Bridier-Nahmias, A.; Le Nagard, H.; Denamur, E.; Clermont, O. ClermonTyping: An easy-to-use and accurate in silico method for Escherichia genus strain phylotyping. Microb. Genom. 2018, 4, e000192. [Google Scholar] [CrossRef]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef] [PubMed]
- Shaik, S.; Ranjan, A.; Tiwari, S.K.; Hussain, A.; Nandanwar, N.; Kumar, N.; Jadhav, S.; Semmler, T.; Baddam, R.; Islam, M.A.; et al. Comparative genomic analysis of globally dominant ST131 clone with other epidemiologically successful extraintestinal pathogenic Escherichia coli (ExPEC) lineages. mBio 2017, 8, e01596-17. [Google Scholar] [CrossRef] [Green Version]
- Mazumder, R.; Hussain, A.; Abdullah, A.; Islam, M.N.; Sadique, M.T.; Muniruzzaman, S.M.; Tabassum, A.; Halim, F.; Akter, N.; Ahmed, D.; et al. International High-Risk Clones Among Extended-Spectrum β-Lactamase-Producing Escherichia coli in Dhaka, Bangladesh. Front. Microbiol. 2021, 12, 736464. [Google Scholar] [CrossRef]
- Peirano, G.; Chen, L.; Nobrega, D.; Finn, T.J.; Kreiswirth, B.N.; DeVinney, R.; Pitout, J.D.D. Genomic Epidemiology of Global Carbapenemase-Producing Escherichia coli, 2015–2017. Emerg. Infect. Dis. 2022, 28, 924–931. [Google Scholar] [CrossRef]
- Riley, L.W. Distinguishing Pathovars from Nonpathovars: Escherichia coli. Microbiol. Spectr. 2020, 8, AME-0014-2020. [Google Scholar] [CrossRef] [PubMed]
- Lindblom, A.; Kiszakiewicz, C.; Kristiansson, E.; Yazdanshenas, S.; Kamenska, N.; Karami, N.; Åhrén, C. The impact of the ST131 clone on recurrent ESBL-producing E. coli urinary tract infection: A prospective comparative study. Sci. Rep. 2022, 12, 10048. [Google Scholar] [CrossRef] [PubMed]
- Nicolas-Chanoine, M.H.; Blanco, J.; Leflon-Guibout, V.; Demarty, R.; Alonso, M.P.; Caniça, M.M.; Park, Y.J.; Lavigne, J.P.; Pitout, J.; Johnson, J.R. Intercontinental emergence of Escherichia coli clone O25:H4-ST131 producing CTX-M-15. J. Antimicrob. Chemother. 2008, 61, 273–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coque, T.M.; Novais, A.; Carattoli, A.; Poirel, L.; Pitout, J.; Peixe, L.; Baquero, F.; Cantón, R.; Nordmann, P. Dissemination of clonally related Escherichia coli strains expressing extended-spectrum beta-lactamase CTX-M-15. Emerg. Infect. Dis. 2008, 14, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Johnson, J.R. A new clone sweeps clean: The enigmatic emergence of Escherichia coli sequence type 131. Antimicrob. Agents Chemother. 2014, 58, 4997–5004. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, X.; Yang, A.W.; Tang, B.; Jian, Z.J.; Zhong, Y.M.; Li, H.L.; Li, Y.M.; Yan, Q.; Liang, X.H.; et al. Community Fecal Carriage and Molecular Epidemiology of Extended-Spectrum β-Lactamase- and Carbapenemase-Producing Escherichia coli from Healthy Children in the Central South China. Infect. Drug Resist. 2022, 15, 1601–1611. [Google Scholar] [CrossRef]
- Bueris, V.; Sellera, F.P.; Fuga, B.; Sano, E.; Carvalho, M.P.N.; Couto, S.C.F.; Moura, Q.; Lincopan, N. Convergence of virulence and resistance in international clones of WHO critical priority enterobacterales isolated from Marine Bivalves. Sci. Rep. 2022, 12, 5707. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, T.; Clermont, O.; Gouriou, S.; Picard, B.; Nassif, X.; Denamur, E.; Tenaillon, O. Extraintestinal virulence is a coincidental by-product of commensalism in B2 phylogenetic group Escherichia coli strains. Mol. Biol. Evol. 2007, 24, 2373–2384. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Dhanji, H.; Upton, M.; Gibreel, T.; Fox, A.; Boyd, D.; Mulvey, M.R.; Nordmann, P.; Ruppe, E.; Sarthou, J.L.; et al. Rapid detection of the O25b-ST131 clone of Escherichia coli encompassing the CTX-M-15-producing strains. J. Antimicrob. Chemother. 2009, 64, 274–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croxall, G.; Hale, J.; Weston, V.; Manning, G.; Cheetham, P.; Achtman, M.; McNally, A. Molecular epidemiology of extraintestinal pathogenic Escherichia coli isolates from a regional cohort of elderly patients highlights the prevalence of ST131 strains with increased antimicrobial resistance in both community and hospital care settings. J. Antimicrob. Chemother. 2011, 66, 2501–2508. [Google Scholar] [CrossRef]
- Russo, T.A.; Johnson, J.R. Proposal for a new inclusive designation for extraintestinal pathogenic isolates of Escherichia coli: ExPEC. J. Infect. Dis. 2000, 181, 1753–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas-Chanoine, M.H.; Bertrand, X.; Madec, J.Y. Escherichia coli ST131, an intriguing clonal group. Clin. Microbiol. Rev. 2014, 27, 543–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Zakour, N.L.; Alsheikh-Hussain, A.S.; Ashcroft, M.M.; Khanh Nhu, N.T.; Roberts, L.W.; Stanton-Cook, M.; Schembri, M.A.; Beatson, S.A. Sequential Acquisition of Virulence and Fluoroquinolone Resistance Has Shaped the Evolution of Escherichia coli ST131. mBio 2016, 7, e00347-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, P.N.A.; Ben Zakour, N.L.; Roberts, L.W.; Wailan, A.M.; Zowawi, H.M.; Tambyah, P.A.; Lye, D.; Jureen, R.; Lee, T.H.; Yin, M.; et al. Whole genome analysis of cephalosporin-resistant Escherichia coli from bloodstream infections in Australia, New Zealand and Singapore: High prevalence of CMY-2 producers and ST131 carrying blaCTX-M-15 and blaCTX-M-27. J. Antimicrob. Chemother. 2018, 73, 634–642. [Google Scholar] [CrossRef]
- Zhong, Y.; Guo, S.; Seow, K.L.G.; Ming, G.O.H.; Schlundt, J. Characterization of extended-spectrum beta-lactamase-producing Escherichia coli isolates from Jurong Lake, Singapore, with whole-genome sequencing. Int. J. Environ. Res. Public Health 2021, 18, 937. [Google Scholar] [CrossRef]
- Lopes, R.; Furlan, J.P.R.; Dos Santos, L.D.R.; Gallo, I.F.L.; Stehling, E.G. Colistin-Resistant mcr-1-Positive Escherichia coli ST131-H22 Carrying blaCTX-M-15 and qnrB19 in Agricultural Soil. Front. Microbiol. 2021, 12, 659900. [Google Scholar] [CrossRef]
- Saidenberg, A.B.S.; Stegger, M.; Price, L.B.; Johannesen, T.B.; Aziz, M.; Cunha, M.P.V.; Moreno, A.M.; Knöbl, T. mcr-Positive Escherichia coli ST131-H22 from Poultry in Brazil. Emerg. Infect. Dis. 2020, 26, 1951–1954. [Google Scholar] [CrossRef]
- Pajand, O.; Rahimi, H.; Darabi, N.; Roudi, S.; Ghassemi, K.; Aarestrup, F.M.; Leekitcharoenphon, P. Arrangements of Mobile Genetic Elements among Virotype E Subpopulation of Escherichia coli Sequence Type 131 Strains with High Antimicrobial Resistance and Virulence Gene Content. mSphere 2021, 6, e0055021. [Google Scholar] [CrossRef]
- Nielsen, D.W.; Ricker, N.; Barbieri, N.L.; Wynn, J.L.; Gomez-Duarte, O.G.; Iqbal, J.; Nolan, L.K.; Allen, H.K.; Logue, C.M. Complete genome sequence of the multidrug-resistant neonatal meningitis Escherichia coli serotype O75:H5:K1 strain mcjchv-1 (NMEC-O75). Microbiol. Resour. Announc. 2018, 7, e01043-18. [Google Scholar] [CrossRef] [Green Version]
- Massella, E.; Giacometti, F.; Bonilauri, P.; Reid, C.; Djordjevic, S.; Merialdi, G.; Bacci, C.; Fiorentini, L.; Massi, P.; Bardasi, L.; et al. Antimicrobial Resistance Profile and ExPEC Virulence Potential in Commensal Escherichia coli of Multiple Sources. Antibiotics 2021, 10, 351. [Google Scholar] [CrossRef]
- Monte, D.F.; Sellera, F.P.; Fernandes, M.R.; Moura, Q.; Landgraf, M.; Lincopan, N. Genome Sequencing of an Escherichia coli Sequence Type 617 Strain Isolated from Beach Ghost Shrimp (Callichirus major) from a Heavily Polluted Ecosystem Reveals a Wider Resistome against Heavy Metals and Antibiotics. Microbiol. Resour. Announc. 2019, 8, e01471-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Xu, J.; He, F. Draft genome sequence of an NDM-5, CTX-M-15 and OXA-1 co-producing Escherichia coli ST167 clinical strain isolated from a urine sample. J. Glob. Antimicrob. Resist. 2018, 14, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Takayama, Y.; Sekizuka, T.; Matsui, H.; Adachi, Y.; Eda, R.; Nihonyanagi, S.; Wada, T.; Matsui, M.; Suzuki, S.; Takaso, M.; et al. Characterization of the IncFII-IncFIB(pB171) Plasmid Carrying blaNDM-5 in Escherichia coli ST405 Clinical Isolate in Japan. Infect. Drug Resist. 2020, 13, 561–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumbana, J.J.; Santona, A.; Fiamma, M.; Taviani, E.; Deligios, M.; Zimba, T.; Lucas, G.; Sacarlal, J.; Rubino, S.; Paglietti, B. Extraintestinal Pathogenic Escherichia coli ST405 Isolate Coharboring blaNDM-5 and blaCTXM-15: A New Threat in Mozambique. Microb. Drug Resist. 2021, 27, 1633–1640. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Feng, Y.; Zhou, W.; McNally, A.; Zong, Z. Cryptic transmission of ST405 Escherichia coli carrying blaNDM-4 in hospital. Sci. Rep. 2018, 8, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, P.R.; McKinnon, J.; Liu, M.; Djordjevic, S.P. Multidrug Resistant Uropathogenic Escherichia coli ST405 With a Novel, Composite IS26 Transposon in a Unique Chromosomal Location. Front. Microbiol. 2019, 9, 3212. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, M.R.; Sellera, F.P.; Cunha, M.P.V.; Lopes, R.; Cerdeira, L.; Lincopan, N. Emergence of CTX-M-27-producing Escherichia coli of ST131 and clade C1-M27 in an impacted ecosystem with international maritime traffic in South America. J. Antimicrob. Chemother. 2020, 75, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.; Mangat, R.; Mostafa, H.H.; Taffner, S.; Wang, J.; Dumyati, G.; Stanton, R.A.; Daniels, J.B.; Campbell, D.; Lutgring, J.D.; et al. Detection of CTX-M-27 β-Lactamase Genes on Two Distinct Plasmid Types in ST38 Escherichia coli from Three, U.S. States. Antimicrob. Agents Chemother. 2021, 65, e0082521. [Google Scholar] [CrossRef]
- Mostafa, H.H.; Cameron, A.; Taffner, S.M.; Wang, J.; Malek, A.; Dumyati, G.; Hardy, D.J.; Pecora, N.D. Genomic Surveillance of Ceftriaxone-Resistant Escherichia coli in Western New York Suggests the Extended-Spectrum β-Lactamase blaCTX-M-27 Is Emerging on Distinct Plasmids in ST38. Front. Microbiol. 2020, 11, 1747. [Google Scholar] [CrossRef]
- Sellera, F.P.; Fernandes, M.R.; Moura, Q.; Lopes, R.B.; Souza, T.A.; Cerdeira, L.; Lincopan, N. Draft genome sequence of a blaCMY-2/IncI1-harbouring Escherichia coli D:ST457 isolated from coastal benthic organisms. J. Glob. Antimicrob. Resist. 2018, 14, 83–84. [Google Scholar] [CrossRef]
- Sartori, L.; Fernandes, M.R.; Ienne, S.; de Souza, T.A.; Gregory, L.; Cerdeira, L.; Lincopan, N. Draft genome sequences of two fluoroquinolone-resistant CTX-M-15-producing Escherichia coli ST90 (ST23 complex) isolated from a calf and a dairy cow in South America. J. Glob. Antimicrob. Resist. 2017, 11, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Stephens, C.M.; Skerker, J.M.; Sekhon, M.S.; Arkin, A.P.; Riley, L.W. Complete genome sequences of four Escherichia coli ST95 isolates from bloodstream infections. Genome Announc. 2015, 3, e01241-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlan, J.P.R.; Gonzalez, I.H.L.; Ramos, P.L.; Stehling, E.G. International high-risk clone of multidrug-resistant CTX-M-8-producing Escherichia coli C-ST410 infecting an elephant (Loxodonta africana) in a zoo. J. Glob. Antimicrob. Resist. 2020, 22, 643–645. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.R.; Sellera, F.P.; Moura, Q.; Gaspar, V.C.; Cerdeira, L.; Lincopan, N. International high-risk clonal lineages of CTX-M-producing Escherichia coli F-ST648 in free-roaming cats, South America. Infect. Genet. Evol. 2018, 66, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.A.; Zaccariotto, T.R.; Piveta, C.; Hofling, C.C.; Resende, M.R.; Levy, C.E.; Esposito, F.; Fernandes, M.R.; Cerdeira, L.; Lincopan, N. MCR-1-positive colistin-resistant Escherichia coli in immunocompromised hospitalised patients. Int. J. Antimicrob. Agents 2018, 52, 438–440. [Google Scholar] [CrossRef]
- Reid, C.J.; Cummins, M.L.; Börjesson, S.; Brouwer, M.S.M.; Hasman, H.; Hammerum, A.M.; Roer, L.; Hess, S.; Berendonk, T.; Nešporová, K.; et al. A role for ColV plasmids in the evolution of pathogenic Escherichia coli ST58. Nat. Commun. 2022, 13, 683. [Google Scholar] [CrossRef]
- Santos, A.C.M.; Silva, R.M.; Valiatti, T.B.; Santos, F.F.; Santos-Neto, J.F.; Cayô, R.; Streling, A.P.; Nodari, C.S.; Gales, A.C.; Nishiyama-Jr, M.Y.; et al. Virulence Potential of a Multidrug-Resistant Escherichia coli Strain Belonging to the Emerging Clonal Group ST101-B1 Isolated from Bloodstream Infection. Microorganisms 2020, 8, 827. [Google Scholar] [CrossRef]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Danzeisen, J.L.; Youmans, B.; Case, K.; Llop, K.; Munoz-Aguayo, J.; Flores-Figueroa, C.; Aziz, M.; Stoesser, N.; Sokurenko, E.; et al. Separate F-Type Plasmids Have Shaped the Evolution of the H30 Subclone of Escherichia coli Sequence Type 131. mSphere 2016, 1, e00121-16. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, Y.; Pitout, J.D.; Gomi, R.; Matsuda, T.; Noguchi, T.; Yamamoto, M.; Peirano, G.; DeVinney, R.; Bradford, P.A.; Motyl, M.R.; et al. Global Escherichia coli sequence type 131 clade with blaCTX-M-27 gene. Emerg. Infect. Dis. 2016, 22, 1900–1907. [Google Scholar] [CrossRef]
- Merino, I.; Hernández-García, M.; Turrientes, M.C.; Pérez-Viso, B.; López-Fresneña, N.; Diaz-Agero, C.; Maechler, F.; Fankhauser-Rodriguez, C.; Kola, A.; Schrenzel, J.; et al. Emergence of ESBL-producing Escherichia coli ST131-C1-M27 clade colonizing patients in Europe. J. Antimicrob. Chemother. 2018, 73, 2973–2980. [Google Scholar] [CrossRef] [PubMed]
- Peirano, G.; Lynch, T.; Matsumara, Y.; Nobrega, D.; Finn, T.J.; DeVinney, R.; Pitout, J.D. Trends in population dynamics of Escherichia coli sequence type 131, Calgary, Alberta, Canada, 2006–2016. Emerg. Infect. Dis. 2020, 26, 2907–2915. [Google Scholar] [CrossRef] [PubMed]
- Hojabri, Z.; Darabi, N.; Arab, M.; Saffari, F.; Pajand, O. Clonal diversity, virulence genes content and subclone status of Escherichia coli sequence type 131: Comparative analysis of E. coli ST131 and non-ST131 isolates from Iran. BMC Microbiol. 2019, 19, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, K.; Tóth, Á.; Kamotsay, K.; Németh, V.; Szabó, D. Population snapshot of the extended-spectrum β-lactamase-producing Escherichia coli invasive strains isolated from a Hungarian hospital. Ann. Clin. Microbiol. Antimicrob. 2022, 21, 3. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chavda, K.D.; Al Laham, N.; Melano, R.G.; Jacobs, M.R.; Bonomo, R.A.; Kreiswirth, B.N. Complete nucleotide sequence of a blaKPC-harboring IncI2 plasmid and its dissemination in New Jersey and New York hospitals. Antimicrob. Agents Chemother. 2013, 57, 5019–5025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Chavda, K.D.; Melano, R.G.; Hong, T.; Rojtman, A.D.; Jacobs, M.R.; Bonomo, R.A.; Kreiswirth, B.N. Molecular survey of the dissemination of two blaKPC-harboring IncFIA plasmids in New Jersey and New York hospitals. Antimicrob. Agents Chemother. 2014, 58, 2289–2294. [Google Scholar] [CrossRef] [Green Version]
- Strydom, K.A.; Chen, L.; Kock, M.; Stoltz, A.C.; Peirano, G.; Nobrega, D.B.; Lowe, M.; Ehlers, M.M.; Mbelle, N.M.; Kreiswirth, B.N.; et al. Klebsiella pneumoniae ST307 with OXA-181: Threat of a high-risk clone and promiscuous plasmid in a resource-constrained healthcare setting. J. Antimicrob. Chemother. 2020, 75, 896–902. [Google Scholar] [CrossRef]
- Johnson, J.R.; Clermont, O.; Johnston, B.; Clabots, C.; Tchesnokova, V.; Sokurenko, E.; Junka, A.F.; Maczynska, B.; Denamur, E. Rapid and specific detection, molecular epidemiology, and experimental virulence of the O16 subgroup within Escherichia coli sequence type 131. J. Clin. Microbiol. 2014, 52, 1358–1365. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, Q.; Huang, J.; Cao, Y.; Zhao, Z.; Li, B. Epidemic potential of Escherichia coli O16: H41-ST131:compared with pandemic O25b: H30-ST131 Lineage. Infect. Drug Resist. 2021, 14, 2625–2632. [Google Scholar] [CrossRef]
- Petty, N.K.; Ben Zakour, N.L.; Stanton-Cook, M.; Skippington, E.; Totsika, M.; Forde, B.M.; Phan, M.D.; Gomes Moriel, D.; Peters, K.M.; Davies, M.; et al. Global dissemination of a multidrug-resistant Escherichia coli clone. Proc. Natl. Acad. Sci. USA 2014, 111, 5694–5699. [Google Scholar] [CrossRef]
- Chen, S.L.; Ding, Y.; Apisarnthanarak, A.; Kalimuddin, S.; Archuleta, S.; Omar, S.F.S.; De, P.P.; Koh, T.H.; Chew, K.L.; Atiya, N.; et al. The higher prevalence of extended spectrum beta-lactamases among Escherichia coli ST131 in Southeast Asia is driven by expansion of a single, locally prevalent subclone. Sci. Rep. 2019, 9, 13245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moulin-Schouleur, M.; Répérant, M.; Laurent, S.; Brée, A.; Mignon-Grasteau, S.; Germon, P. Extraintestinal pathogenic Escherichia coli strains of avian and human origin: Link between phylogenetic relationships and common virulence patterns. J. Clin. Microbiol. 2007, 45, 3366–7336. [Google Scholar] [CrossRef] [Green Version]
- Mora, A.; Herrera, A.; Mamani, R.; López, C.; Alonso, M.P.; Blanco, J.E.; Blanco, M.; Dahbi, G.; García-Garrote, F.; Pita, J.M.; et al. Recent emergence of clonal group O25b:K1:H4-B2-ST131 ibeA strains among Escherichia coli poultry isolates, including CTX-M-9-producing strains, and comparison with clinical human isolates. Appl. Environ. Microbiol. 2010, 76, 6991–6997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, C.; Boerlin, P.; Daignault, D.; Dozois, C.M.; Dutil, L.; Galanakis, C.; Reid-Smith, R.J.; Tellier, P.-P.; Tellis, P.A.; Ziebell, K.; et al. Food reservoir for Escherichia coli causing urinary tract infections. Emerg. Infect. Dis. 2010, 16, 88–95. [Google Scholar] [CrossRef]
- Heiden, S.E.; Kurz, M.S.E.; Bohnert, J.; Bayingana, C.; Ndoli, J.M.; Sendegeya, A.; Gahutu, J.B.; Eger, E.; Mockenhaupt, F.P.; Schaufler, K. Flies from a tertiary hospital in Rwanda carry multidrug-resistant Gram-negative pathogens including extended-spectrum beta-lactamase-producing E. coli sequence type 131. Antimicrob. Resist. Infect. Control 2020, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Eger, E.; Heiden, S.E.; Korolew, K.; Bayingana, C.; Ndoli, J.M.; Sendegeya, A.; Gahutu, J.B.; Kurz, M.S.E.; Mockenhaupt, F.P.; Müller, J.; et al. Circulation of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli of Pandemic Sequence Types 131, 648, and 410 Among Hospitalized Patients, Caregivers, and the Community in Rwanda. Front. Microbiol. 2021, 12, 662575. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Elnekave, E.; Miller, E.A.; Munoz-Aguayo, J.; Figueroa, C.F.; Johnston, B.; Nielson, D.W.; Logue, C.M.; Johnson, J.R. Phylogenomic analysis of extraintestinal pathogenic Escherichia coli sequence type 1193, an emerging multidrug-resistant clonal group. Antimicrob. Agents Chemother. 2018, 63, e01913-18. [Google Scholar] [CrossRef] [Green Version]
- Peirano, G.; Matsumara, Y.; Nobrega, D.; DeVinney, R.; Pitout, J. Population-based epidemiology of Escherichia coli ST1193 causing blood stream infections in a centralized Canadian region. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 1–7. [Google Scholar] [CrossRef]
- Pitout, J.D.D.; Peirano, G.; Chen, L.; DeVinney, R.; Matsumura, Y. Escherichia coli ST1193: Following in the Footsteps of E. coli ST131. Antimicrob. Agents Chemother. 2022, 66, e0051122. [Google Scholar] [CrossRef]
- Birgy, A.; Madhi, F.; Jung, C.; Levy, C.; Cointe, A.; Bidet, P.; Hobson, C.A.; Bechet, S.; Sobral, E.; Vuthien, H.; et al. Diversity and trends in population structure of ESBL-producing Enterobacteriaceae in febrile urinary tract infections in children in France from 2014 to 2017. J. Antimicrob. Chemother. 2020, 75, 96–105. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, J.; Yao, K.; Gao, W.; Wang, Y. Molecular characteristics of the new emerging global clone ST1193 among clinical isolates of Escherichia coli from neonatal invasive infections in China. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 833–840. [Google Scholar] [CrossRef]
- Wang, C.H.; Siu, L.K.; Chang, F.Y.; Chiu, S.K.; Lin, J.C. A Resistance Mechanism in Non-mcr Colistin-Resistant Escherichia coli in Taiwan: R81H Substitution in PmrA Is an Independent Factor Contributing to Colistin Resistance. Microbiol. Spectr. 2021, 9, e0002221. [Google Scholar] [CrossRef] [PubMed]
- Riley, L.W. Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin. Microbiol. Infect. 2014, 20, 380–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallonen, T.; Brodrick, H.J.; Harris, S.R.; Corander, J.; Brown, N.M.; Martin, V.; Peacock, S.J.; Parkhill, J. Systematic longitudinal survey of invasive Escherichia coli in England demonstrates a stable population structure only transiently disturbed by the emergence of ST131. Genome Res. 2017, 27, 1437–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Kawahara, R.; Harada, K.; Teruya, S.; Nakayama, T.; Motooka, D.; Nakamura, S.; Nguyen, P.D.; Kumeda, Y.; Van-Dang, C. The presence of colistin resistance gene mcr-1 and -3 in ESBL producing Escherichia coli isolated from food in Ho Chi Minh City Vietnam. FEMS Microbiol. Lett. 2018, 365, fny100. [Google Scholar] [CrossRef] [PubMed]
- Hameed, M.F.; Chen, Y.; Bilal, H.; Khan, S.; Ge, H.; Xiaofang, C.; Gu, P. The Co-occurrence of mcr-3 and fosA3 in IncP plasmid in ST131 Escherichia coli: A novel case. J. Infect. Dev. Ctries. 2022, 16, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Nüesch-Inderbinen, M.T.; Baschera, M.; Zurfluh, K.; Hächler, H.; Nüesch, H.; Stephan, R. Clonal Diversity, Virulence Potential and Antimicrobial Resistance of Escherichia coli Causing Community Acquired Urinary Tract Infection in Switzerland. Front. Microbiol. 2017, 8, 2334. [Google Scholar] [CrossRef] [Green Version]
- Massella, E.; Reid, C.J.; Cummins, M.L.; Anantanawat, K.; Zingali, T.; Serraino, A.; Piva, S.; Giacometti, F.; Djordjevic, S.P. Snapshot Study of Whole Genome Sequences of Escherichia coli from Healthy Companion Animals, Livestock, Wildlife, Humans and Food in Italy. Antibiotics 2020, 9, 782. [Google Scholar] [CrossRef]
- Nascimento, T.; Cantamessa, R.; Melo, L.; Fernandes, M.R.; Fraga, E.; Dropa, M.; Sato, M.I.; Cerdeira, L.; Lincopan, N. International high-risk clones of Klebsiella pneumoniae KPC-2/CC258 and Escherichia coli CTX-M-15/CC10 in urban lake waters. Sci. Total Environ. 2017, 598, 910–915. [Google Scholar] [CrossRef]
- Reid, C.J.; DeMaere, M.Z.; Djordjevic, S.P. Australian porcine clonal complex 10 (CC10) Escherichia coli belong to multiple sublineages of a highly diverse global CC10 phylogeny. Microb. Genom. 2019, 5, e000225. [Google Scholar] [CrossRef] [PubMed]
- Skarżyńska, M.; Zaja̧c, M.; Bomba, A.; Bocian, Ł.; Kozdruń, W.; Polak, M.; Wia̧cek, J.; Wasyl, D. Antimicrobial Resistance Glides in the Sky-Free-Living Birds as a Reservoir of Resistant Escherichia coli With Zoonotic Potential. Front. Microbiol. 2021, 12, 656223. [Google Scholar] [CrossRef] [PubMed]
- Fuga, B.; Sellera, F.P.; Cerdeira, L.; Esposito, F.; Cardoso, B.; Fontana, H.; Moura, Q.; Cardenas-Arias, A.; Sano, E.; Ribas, R.M.; et al. WHO Critical Priority Escherichia coli as One Health Challenge for a Post-Pandemic Scenario: Genomic Surveillance and Analysis of Current Trends in Brazil. Microbiol. Spectr. 2022, 10, e0125621. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ye, K.; Li, X.; Ye, L.; Guo, L.; Wang, L.; Yang, J. Genetic characterization of Carbapenem-Resistant Escherichia coli from China, 2015–2017. BMC Microbiol. 2021, 21, 248. [Google Scholar] [CrossRef]
- Schaufler, K.; Semmler, T.; Wieler, L.H.; Wöhrmann, M.; Baddam, R.; Ahmed, N.; Müller, K.; Kola, A.; Fruth, A.; Ewers, C.; et al. Clonal spread and interspecies transmission of clinically relevant ESBL-producing Escherichia coli of ST410—Another successful pandemic clone? FEMS Microbiol. Ecol. 2016, 92, fiv155. [Google Scholar] [CrossRef] [Green Version]
- Falgenhauer, L.; Imirzalioglu, C.; Ghosh, H.; Gwozdzinski, K.; Schmiedel, J.; Gentil, K.; Bauerfeind, R.; Kämpfer, P.; Seifert, H.; Michael, G.B.; et al. Circulation of clonal populations of fluoroquinolone-resistant CTX-M-15- producing Escherichia coli ST410 in humans and animals in Germany. Int. J. Antimicrob. Agents 2016, 47, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, Y.; Wu, W.; Xie, Y.; Wang, X.; Zhang, X.; Chen, X.; Zong, Z. First report of OXA-181-producing Escherichia coli in China and characterization of the isolate using whole-genome sequencing. Antimicrob. Agents Chemother. 2015, 59, 5022–5025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piazza, A.; Comandatore, F.; Romeri, F.; Pagani, C.; Floriano, A.M.; Ridolfo, A.; Antona, C.; Brilli, M.; Mattioni Marchetti, V.; Bandi, C.; et al. First report of an ST410 OXA-181 and CTX-M-15 coproducing Escherichia coli clone in Italy: A whole-genome sequence characterization. Microb. Drug Resist. 2018, 24, 1207–1209. [Google Scholar] [CrossRef]
- Roer, L.; Overballe-Petersen, S.; Hansen, F.; Schønning, K.; Wang, M.; Røder, B.L.; Hansen, D.S.; Justesen, U.S.; Andersen, L.P.; Fulgsang-Damgaard, D.; et al. Escherichia coli Sequence Type 410 Is Causing New International High-Risk Clones. mSphere 2018, 3, e00337-18. [Google Scholar] [CrossRef] [Green Version]
- Overballe-Petersen, S.; Roer, L.; Ng, K.; Hansen, F.; Justesen, U.S.; Andersen, L.P.; Stegger, M.; Hammerum, A.M.; Hasman, H. Complete nucleotide sequence of an Escherichia coli sequence type 410 strain carrying blaNDM-5 on an IncF multidrug resistance plasmid and bla OXA-181 on an IncX3 plasmid. Genome Announc. 2018, 6, e01542-17. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Zheng, X.; Sun, Y.; Fang, R.; Zhang, S.; Zhang, X.; Lin, J.; Cao, J.; Zhou, T. Molecular mechanisms and epidemiology of carbapenem-resistant Escherichia coli isolated from Chinese patients during 2002–2017. Infect. Drug Resist. 2020, 13, 501–512. [Google Scholar] [CrossRef]
- Peri, A.M.; Piazza, A.; De Zan, V.; Carugati, M.; Muscatello, A.; Comandatore, F.; De Lorenzis, E.; Pluderi, M.; Arghittu, M.; Cariani, L.; et al. Autochthonous ST405 NDM-5 producing Escherichia coli causing fatal sepsis in Northern Italy. Int. J. Antimicrob. Agents 2020, 55, 105953. [Google Scholar] [CrossRef]
- Touati, M.; Hadjadj, L.; Berrazeg, M.; Baron, S.A.; Rolain, J.M. Emergence of Escherichia coli harbouring mcr-1 and mcr-3 genes in North West Algerian farmlands. J. Glob. Antimicrob. Resist. 2020, 21, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, G.; Wiltshire, C.; Macaskill, S.; Tournu, H.; Budge, S.; Brown, A.J. Emergence and spread of B2-ST131-O25b, B2-ST131-O16 and D-ST405 clonal groups among extended-spectrum-beta-lactamase-producing Escherichia coli in Japan. J. Antimicrob. Chemother. 2012, 67, 2612–2620. [Google Scholar] [CrossRef] [Green Version]
- Chattaway, M.A.; Jenkins, C.; Ciesielczuk, H.; Day, M.; DoNascimento, V.; Day, M.; Rodríguez, I.; van Essen-Zandbergen, A.; Schink, A.K.; Wu, G.; et al. Evidence of evolving extraintestinal enteroaggregative Escherichia coli ST38 clone. Emerg. Infect. Dis. 2014, 20, 1935–1937. [Google Scholar] [CrossRef]
- Poirel, L.; Bernabeu, S.; Fortineau, N.; Podglajen, I.; Lawrence, C.; Nordmann, P. Emergence of OXA-48-producing Escherichia coli clone ST38 in France. Antimicrob. Agents Chemother. 2011, 55, 4937–4938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turton, J.F.; Doumith, M.; Hopkins, K.L.; Perry, C.; Meunier, D.; Woodford, N. Clonal expansion of Escherichia coli ST38 carrying a chromosomally integrated OXA-48 carbapenemase gene. J. Med. Microbiol. 2016, 65, 538–546. [Google Scholar] [CrossRef]
- Gauthier, L.; Dortet, L.; Cotellon, G.; Creton, E.; Cuzon, G.; Ponties, V.; Bonnin, R.A.; Naas, T. Diversity of Carbapenemase-Producing Escherichia coli Isolates in France in 2012–2013. Antimicrob. Agents Chemother. 2018, 62, e00266-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falgenhauer, L.; Nordmann, P.; Imirzalioglu, C.; Yao, Y.; Falgenhauer, J.; Hauri, A.M.; Heinmüller, P.; Chakraborty, T. Cross-border emergence of clonal lineages of ST38 Escherichia coli producing the OXA-48-like carbapenemase OXA-244 in Germany and Switzerland. Int. J. Antimicrob. Agents 2020, 56, 106157. [Google Scholar] [CrossRef]
- Kremer, K.; Kramer, R.; Neumann, B.; Haller, S.; Pfennigwerth, N.; Werner, G.; Gatermann, S.; Schroten, H.; Eckmanns, T.; Hans, J.B. Rapid spread of OXA-244-producing Escherichia coli ST38 in Germany: Insights from an integrated molecular surveillance approach; 2017 to January 2020. Eurosurveillance 2020, 25, 2000923. [Google Scholar] [CrossRef]
- Nesporova, K.; Jamborova, I.; Valcek, A.; Medvecky, M.; Literak, I.; Dolejska, M. Various conjugative plasmids carrying the mcr-5 gene in Escherichia coli isolates from healthy chickens in Paraguay. J. Antimicrob. Chemother. 2019, 74, 3394–3397. [Google Scholar] [CrossRef]
- Sekizuka, T.; Matsui, M.; Yamane, K.; Takeuchi, F.; Ohnishi, M.; Hishinuma, A.; Arakawa, Y.; Kuroda, M. Complete sequencing of the bla(NDM-1)-positive IncA/C plasmid from Escherichia coli ST38 isolate suggests a possible origin from plant pathogens. PLoS ONE 2011, 6, e25334. [Google Scholar] [CrossRef]
- Guenther, S.; Semmler, T.; Stubbe, A.; Stubbe, M.; Wieler, L.H.; Schaufler, K. Chromosomally encoded ESBL genes in Escherichia coli of ST38 from Mongolian wild birds. J. Antimicrob. Chemother. 2017, 72, 1310–1313. [Google Scholar] [CrossRef]
- Mora, A.; García-Peña, F.J.; Alonso, M.P.; Pedraza-Diaz, S.; Ortega-Mora, L.M.; Garcia-Parraga, D.; López, C.; Viso, S.; Dahbi, G.; Marzoa, J.; et al. Impact of human-associated Escherichia coli clonal groups in Antarctic pinnipeds: Presence of ST73, ST95, ST141 and ST131. Sci. Rep. 2018, 8, 4678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Accogli, M.; Giani, T.; Monaco, M.; Giufrè, M.; García-Fernández, A.; Conte, V.; D’Ancona, F.; Pantosti, A.; Rossolini, G.M.; Cerquetti, M. Emergence of Escherichia coli ST131 sub-clone H30 producing VIM-1 and KPC-3 carbapenemases, Italy. J. Antimicrob. Chemother. 2014, 69, 2293–2296. [Google Scholar] [CrossRef] [Green Version]
- Chavda, K.D.; Chen, L.; Jacobs, M.R.; Bonomo, R.A.; Kreiswirth, B.N. Molecular diversity and plasmid analysis of KPC-producing Escherichia coli. Antimicrob. Agents Chemother. 2016, 60, 4073–4081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aquino-Andrade, A.; Merida-Vieyra, J.; Arias de la Garza, E.; Arzate-Barbosa, P.; De Colsa Ranero, A. Carbapenemase-producing Enterobacteriaceae in Mexico: Report of seven non-clonal cases in a pediatric hospital. BMC Microbiol. 2018, 18, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, F.; Tian, D.; Wang, B.; Zhao, W.; Qin, H.; Zhang, T.; Zhang, H. Fecal carriage and molecular epidemiology of carbapenem-resistant Enterobacteriaceae from outpatient children in Shanghai. BMC Infect. Dis. 2019, 19, 678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGann, P.; Snesrud, E.; Maybank, R.; Corey, B.; Ong, A.C.; Clifford, R.; Hinkle, M.; Whitman, T.; Lesho, E.; Schaecher, K.E. Escherichia coli harboring mcr-1 and blaCTX-M on a novel IncF plasmid: First report of mcr-1 in the United States. Antimicrob. Agents Chemother. 2016, 60, 4420–4421. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Qu, F.; Shan, B.; Huang, B.; Jia, W.; Chen, C.; Li, A.; Miao, M.; Zhang, X.; Bao, C.; et al. Detection of the mcr-1 colistin resistance gene in carbapenem-resistant Enterobacteriaceae from different hospitals in China. Antimicrob. Agents Chemother. 2016, 60, 5033–5035. [Google Scholar] [CrossRef] [Green Version]
- Tada, T.; Nhung, P.H.; Shimada, K.; Tsuchiya, M.; Phuong, D.M.; Anh, N.Q.; Ohmagari, N.; Kirikae, T. Emergence of colistin-resistant Escherichia coli clinical isolates harboring mcr-1 in Vietnam. Int. J. Infect. Dis. 2017, 63, 72–73. [Google Scholar] [CrossRef]
- Vieyra, J.; De Colsa-Ranero, A.; Arzate-Barbosa, P.; Arias-de la Garza, E.; Méndez-Tenorio, A.; Murcia-Garzón, J.; Aquino-Andrade, A. First clinical isolate of Escherichia coli harboring mcr-1 gene in Mexico. PLoS ONE 2019, 14, e0214648. [Google Scholar] [CrossRef]
- Wang, X.; Zhai, W.; Li, J.; Liu, D.; Zhang, Q.; Shen, Z.; Wang, S.; Wang, Y. Presence of an mcr-3 variant in Aeromonas caviae, Proteus mirabilis, and Escherichia coli from one domestic duck. Antimicrob. Agents Chemother. 2017, 62, e02106-17. [Google Scholar] [CrossRef] [Green Version]
- Melendez, D.; Roberts, M.C.; Greninger, A.L.; Weissman, S.; No, D.; Rabinowitz, P.; Wasser, S. Whole-genome analysis of extraintestinal pathogenic Escherichia coli (ExPEC) MDR ST73 and ST127 isolated from endangered southern resident killer whales (Orcinus orca). J. Antimicrob. Chemother. 2019, 74, 2176–2180. [Google Scholar] [CrossRef] [PubMed]
- Cummins, M.L.; Reid, C.J.; Djordjevic, S.P. F Plasmid Lineages in Escherichia coli ST95: Implications for Host Range, Antibiotic Resistance, and Zoonoses. mSystems 2022, 7, e0121221. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G. The Challenge to Control Emergence of Antibiotic Resistance in Virulent Escherichia coli Isolates in Latin America. Microbiol. Spectr. 2022, 10, e01506-22. [Google Scholar] [CrossRef]
- Kadar, B.; Kocsis, B.; Nagy, K.; Szabo, D. The renaissance of polymyxins. Curr. Med. Chem. 2013, 20, 3759–3773. [Google Scholar] [CrossRef]
- Roer, L.; Hansen, F.; Thomsen, M.C.F.; Knudsen, J.D.; Hansen, D.S.; Wang, M.; Samulioniené, J.; Justesen, U.S.; Rřder, B.L.; Schumacher, H.; et al. WGS-based surveillance of third-generation cephalosporin-resistant Escherichia coli from bloodstream infections in Denmark. J. Antimicrob. Chemother. 2017, 72, 1922–1929. [Google Scholar] [CrossRef]
- Luo, C.; Walk, S.T.; Gordon, D.M.; Feldgarden, M.; Tiedje, J.M.; Konstantinidis, K.T. Genome sequencing of environmental Escherichia coli expands understanding of the ecology and speciation of the model bacterial species. Proc. Natl. Acad. Sci. USA 2011, 108, 7200–7205. [Google Scholar] [CrossRef] [Green Version]
- Tchesnokova, V.; Billig, M.; Chattopadhyay, S.; Linardopoulou, E.; Aprikian, P.; Roberts, P.L.; Skrivankova, V.; Johnston, B.; Gileva, A.; Igusheva, I.; et al. Predictive diagnostics for Escherichia coli infections based on the clonal association of antimicrobial resistance and clinical outcome. J. Clin. Microbiol. 2013, 51, 2991–2999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denamur, E.; Clermont, O.; Bonacorsi, S.; Gordon, D. The population genetics of pathogenic Escherichia coli. Nat. Rev. Microbiol. 2021, 19, 37–54. [Google Scholar] [CrossRef] [PubMed]
- Flament-Simon, S.C.; Nicolas-Chanoine, M.H.; García, V.; Duprilot, M.; Mayer, N.; Alonso, M.P.; García-Meniño, I.; Blanco, J.E.; Blanco, M.; Blanco, J. Clonal Structure, Virulence Factor-encoding Genes and Antibiotic Resistance of Escherichia coli, Causing Urinary Tract Infections and Other Extraintestinal Infections in Humans in Spain and France during 2016. Antibiotics 2020, 9, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flament-Simon, S.-C.; de Toro, M.; Mora, A.; García, V.; García-Meniño, I.; Díaz-Jiménez, D.; Herrera, A.; Blanco, J. Whole Genome Sequencing and Characteristics of mcr-1–Harboring Plasmids of Porcine Escherichia coli Isolates Belonging to the High-Risk Clone O25b:H4-ST131 Clade, B. Front. Microbiol. 2020, 11, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Meniño, I.; Lumbreras, P.; Lestón, L.; Álvarez-Álvarez, M.; García, V.; Hammerl, J.A.; Fernández, J.; Mora, A. Occurrence and Genomic Characterization of Clone ST1193 Clonotype 14-64 in Uncomplicated Urinary Tract Infections Caused by Escherichia coli in Spain. Microbiol. Spectr. 2022, 10, 0004122. [Google Scholar] [CrossRef] [PubMed]
- Horesh, G.; Blackwell, G.A.; Tonkin-Hill, G.; Corander, J.; Heinz, E.; Thomson, N.R. A comprehensive and high-quality collection of Escherichia coli genomes and their genes. Microb. Genom. 2021, 7, 000499. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sequence Type | Accession Numbers (Bioproject, Biosample, SRA, European Nucleotide Archive, EMBL-EBI Database Project) | Reference |
|---|---|---|
| ST131-B | JAENHI000000000.1 (E. coli strain S802) | [56] |
| VEWQ00000000 (E. coli strain UPEC U34) | [57] | |
| ST131-C | JSXN00000000 (E. coli strain NA101) JSXO00000000 (E. coli strain NA112) | [38] |
| JACHQR000000000.1 (E. coli strain LMLEEc001) JACHQP000000000.1 (E. coli strain LMLEEc003) JACHQO000000000.1 (E. coli strain LMLEEc010) JACHQK000000000.1 (E. coli strain LMLEEc025) | [39] | |
| JAJPAL000000000.1 (E. coli strain ECO112) | [20] | |
| NCVZ00000000.1 (E. coli strain EcMO) | [47] | |
| PRJEB46895 (E. coli strain EC-119) | [58] | |
| ST1193 | CP030111 (E. coli strain MCJCHV-1) | [59] |
| JACHQB000000000.1 (E. coli strain LMLEEc041) | [39] | |
| ST69 | SAMN11246379 (E. coli strain EC81) SAMN11246556 (E. coli strain EC369) SAMN11246590 (E. coli strain EC801) SAMN11246609 (E. coli strain EC820) | [60] |
| CC10 (ST617) | PIZJ00000000 (E. coli strain ECCO2) | [61] |
| CC10 (ST167) | CP074120 (E. coli strain EC16) | [7] |
| QLNK00000000 (E. coli strain ECWJ1) | [62] | |
| ST405 | AP019803 (E. coli strain KY1497) | [63] |
| ERS4552076 (E. coli strain SSM100) | [64] | |
| NGUU00000000 (E. coli strain WCHEC96200) | [65] | |
| NXEQ00000000 (E. coli strain 2009-30) NXER00000000 (E. coli strain 2009-27) | [66] | |
| ST38 | NCWA00000000.1 (E. coli strain Ec6M) | [67] |
| SAMN17315482 (strain URMC_401_E_coli) | [68] | |
| RZGB00000000 (E. coli strain URMC_9) RZBC00000000 (E. coli strain URMC_13) | [69] | |
| NCWA00000000.1 (E. coli strain Ec6M) | [47] | |
| MVIO00000000 (E. coli strain NA090) | [38] | |
| ST457 | NDBC00000000 (E. coli strain EM1CRO) | [70] |
| CC23 | NBCL00000000 (E. coli strain 13B) LYPE00000000 (E. coli strain Ec47VL) | [71] |
| ST95 | CP012625 (E. coli strain SF-468) CP012631 (E. coli strain SF-173) CP012633 (E. coli strain SF-166) CP012635 (E. coli strain SF-88) | [72] |
| ST73 | JACHPD000000000.1 (E. coli strain LMLEEc115) | [39] |
| ST410 | VFBH01000000 (E. coli strain A240) | [73] |
| ST648 | PEDQ00000000.1 (E. coli strain ICBECG2) PEDR00000000.1 (E. coli strain ICBECG4) | [74] |
| ST393 | QGIF00000000 (E. coli strain 77H) QHCX00000000 (E. coli strain 51H) | [75] |
| ST58 | SRX10825685 (E. coli strain IBIS_39) | [76] |
| ST101 | VYQD00000000 (E. coli strain EC121) | [77] |
| Resistance Markers of ExPEC High-Risk Clones | ||||||
|---|---|---|---|---|---|---|
| Sequence Type (ST) [References] | Beta-Lactamases | Resistance to Fluoroquinolones | Resistance to Colistin | Inc Plasmid Types | ||
| ESBL | AmpC | Carbapenemases (Ambler Class A, B, D) | ||||
| ST-131-A [46,54,55] | blaCTX-M-14, blaCTX-M-15, blaCTX-M-27+, | blaDHA-1 | blaKPC-3, blaOXA-48 | mutations in gyrA, parC, parE, double alteration of gyrA +, qnrB4, qnrB66, aac(6′)Ib-cr | No data available | FI, FII, A1, B1, B10 |
| ST-131-B (*) + [38,46,56,57] | blaCTX-M-15+ | No data available | qnrA1, * qnrB19+ | mcr-1,(*) mcr-3,(*) mcr-5, * mcr-9* | F-ColV like lineage, FIB-like, FII, HI2, HI2A | |
| ST-131-C1-M27 [20,46,83] | blaCTX-M-27, blaCTX-M−2, blaTEM | No data available | blaNDM-1, blaOXA-232 | No data available | FI:A2:B20 | |
| ST-131-C1-non-M27 [58] | blaCTX-M-14,
blaTEM | No data available | blaKPC-2, blaKPC-3, and blaKPC-18 | No data available | ||
| ST-131-C2 (*) [40,58] | blaCTX-M-15, (*) blaCTX-M-G-1, blaCTX-M-G-2, blaCTX-M-G-8, blaCTX-M-G-9, blaCTX-M-G-25, blaTEM (*) blaSHV-12 | blaFOX−5, | blaNDM-1, blaNDM-5, blaVIM-1, blaKPC-2, blaKPC-3, blaKPC-18, blaOXA-48, blaOXA-181, blaOXA-232, co-expression of blaNDM-1 and blaVIM-1 | qnrB19, (*) qnrS1, mutations in gyrA, parC, parE | mutations in pmrA and pmrB, mcr-3 | FII:A1:B-, F36:A4:B1 |
| ST1193 [39,40,59,84,102] | blaCTX-M-9, blaCTX-M-15, blaCTX-M-27, blaCTX-M-55, blaTEM-1B | blaCMY-2 | blaNDM-1, blaKPC-2 | mutations in gyrA, parC or parE, aac(6′)Ib-cr | mutations in pmrA and pmrB | FII, FIB, I1 |
| ST69 (*) [20,40,60,100,105] | blaCTX-M-1, blaCTX-M-14, blaCTX-M-15, blaCTX-M-27, blaTEM-1B, (*) blaTEM-1C | blaCMY-6 (co-carriage with blaNDM-1) | blaNDM-1, blaKPC-2 | mutations in gyrA and parC (*) | mcr-1,+ mcr-3+ | FIIB |
| CC10 (containing ST10, ST44, ST48, ST167 and ST617) (+) [7,61,62,112,113] | blaCTX-M-14, blaCTX-M-15, blaCTX-M-27, blaCTX-M-55, blaTEM-1B, blaTEM-169, blaTEM-206, blaTEM-214 blaSHV-12 | blaCMY-42, | blaNDM-1, blaNDM-5, blaNDM-9, blaIMP-8, blaKPC-2, blaKPC-3, blaOXA-181, (co-expressed with blaNDM-5) blaOXA-48 (co-expressed with blaNDM-5) | mutations in gyrA and parC, qnrS1, (*) qnrB19, qepA4, aac(6′)-Ib-cr | mcr-1 (co-expressed with blaNDM-1) | IncFIA, Col-like, I1 |
| ST405 (+) [63,64,65,66] | blaCTX-M-14, blaCTX-M-15, blaCTX-M-88, blaTEM-1, blaTEM-12 | No data available | blaNDM-4, blaNDM-5, blaKPC-2, blaOXA-48 | mutations in gyrA, parC, and parE, qnrS1, aac(6′)-Ib-cr | mcr-1+ | FIB, FII, F1A, Col-like, F1:A1:B49 |
| ST38 (*) [38,40,67,68,69,70] | blaCTX-M-2, blaCTX-M-3, blaCTX-M -9, blaCTX-M -14, (*) blaCTX-M-15, (*) blaCTX-M-27, blaTEM-1B | blaCMY-12, blaDHA-1 | blaNDM-1, blaNDM-6, blaOXA-48, blaOXA-244 | mutations in gyrA, parC | mcr-5 (*) | FII:A-:B10, FI:A2:B20 |
| ST457 (*) [19,70] | blaCTX-M-1, blaCTX-M-2, blaCTX-M-3, blaCTX-M-8; blaCTX-M-12, blaCTX-M-14, blaCTX-M-15, blaCTX-M-27, blaCTX-M-55, blaTEM-1 | blaCMY-2 | blaNDM-5, blaNDM-9, blaIMP-4, blaKPC-2, blaKPC-3, blaOXA-23 | mutations in gyrA and parC, qnrB19 * | mcr-1 (*), mcr-3 (*), mcr-5 (*) | F64:A:B:27, I1 |
| Linkage between ExPEC Clones and Colistin, Fosfomycin, Nitrofurantoin Resistance Determinants | ||||||
|---|---|---|---|---|---|---|
| Colistin Resistance | Fosfomycin Resistance | Nitrofurantion Resistance | ||||
| pmrA and pmrB Mutations | mcr-1 | mcr-3 | mcr-5 | mcr-9 | fosA3 Enzymatic Activity | Nitroreductase (nfsA, nfsB) Enzymatic Activity |
| ST131 ST1193 | ST131 ST69 CC10 ST405 ST457 ST410 ST48 ST58 ST88 ST57 ST156 ST224 ST345 ST354 ST393 ST744 ST850 ST3024 ST8900 ST12657 | ST69 ST457 ST101 ST155 ST206 ST443 ST1081 ST1638 ST5038 | ST131-B ST38 ST457 ST57 ST93 ST113 ST165 ST189 ST224 ST366 ST580 ST641 ST752 ST2705 ST6853 ST8061 | ST131-B | ST131 ST1193 ST69 CC10 ST38 ST457 ST95 ST12 ST117 ST307 ST648 ST744 ST746 ST1730 ST2646 | ST131 ST38 |
| Resistance Genes and Mechanisms | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Clones | blaCTX-M-14-Variants (e.g., CTX-M-2) | blaCTX-M-15 | blaCTX-M-27 | blaKPC-Variants | blaOXA-48-Variants (e.g., OXA-244) | blaIMP-Variants | blaNDM-Variants | blaVIM-Variants | Fluoro-Quinolones | Colistin |
| ST131 (including A, B, C1, C2 clades) | 2004 | 2005 | 2004 | 2011 | 2012 | 2010 | 2015-2017 (2022) 1 | 2004 | 2014 | |
| ST1193 | 2013 | 2017 | 2022 | 2022 | 2012 | 2021 | ||||
| ST69 | 2014–2017 (2020) 2 | 2014–2017 (2020) 2 | 2014–2017 (2020) 2 | 2015–2017 (2022) 1 | 2022 | 2017 | 2018 | |||
| CC10 (ST10 with its variants, namely ST167, ST617, ST410) | 2015 | 2015 | 2017 | 2016 | 2015 | 2022 | 2015 | 2017 | 2022 | |
| ST405 | 2012 | 2012 | 2016 | 2016 | 2013 (2020), 2016 3 | 2012 | 2020 | |||
| ST38 | 2011 | 2020 | 2020 | 2011 | 2011 | 2013 (2015) 4 | 2019 | |||
| ST457 | 2016 | 2016 | 2012 | 2014 | 2020 | 2019 | 2014 | 2008 | 2012 (2020) 5 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocsis, B.; Gulyás, D.; Szabó, D. Emergence and Dissemination of Extraintestinal Pathogenic High-Risk International Clones of Escherichia coli. Life 2022, 12, 2077. https://doi.org/10.3390/life12122077
Kocsis B, Gulyás D, Szabó D. Emergence and Dissemination of Extraintestinal Pathogenic High-Risk International Clones of Escherichia coli. Life. 2022; 12(12):2077. https://doi.org/10.3390/life12122077
Chicago/Turabian StyleKocsis, Béla, Dániel Gulyás, and Dóra Szabó. 2022. "Emergence and Dissemination of Extraintestinal Pathogenic High-Risk International Clones of Escherichia coli" Life 12, no. 12: 2077. https://doi.org/10.3390/life12122077