
Effects of Gibberellin Pre-Treatment on Seed Germination and Seedling Physiology Characteristics in Industrial Hemp under Drought Stress Condition

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Germination Experiment
2.3. Determination of Germination and Morphological Traits
2.4. Determination of Physiological and Biochemical Indices
2.5. Data Analyses
3. Results
3.1. Effects of GA3 Pre-Treatment on Germination Indexes of Hempseeds under Drought Stress
3.1.1. ANOVA Analysis of the Effect of Seed Pre-Treatment by GA3 on Germination Indexes of Hempseeds under Drought Stress
3.1.2. Effect of GA3 Pre-Treatment on Germination Rate and Germination Potential of Hempseeds under Drought Stress
3.1.3. Effects of GA3 Pre-Treatment on Hypocotyl and Radicle Growth of Hempseed under Drought Stress
3.2. Effects of Seeds Pre-Treatment by GA3 on Physiology Characteristics in Hemp Seedlings under Drought Stress
3.2.1. Effects of Seeds Pre-Treatment by GA3 on Osmotic Regulation Substances in Hemp Seedlings under Drought Stress
3.2.2. Effects of Seed Pre-Treatment by GA3 on Lipid Peroxidation in Hemp Seedlings under Drought Stress
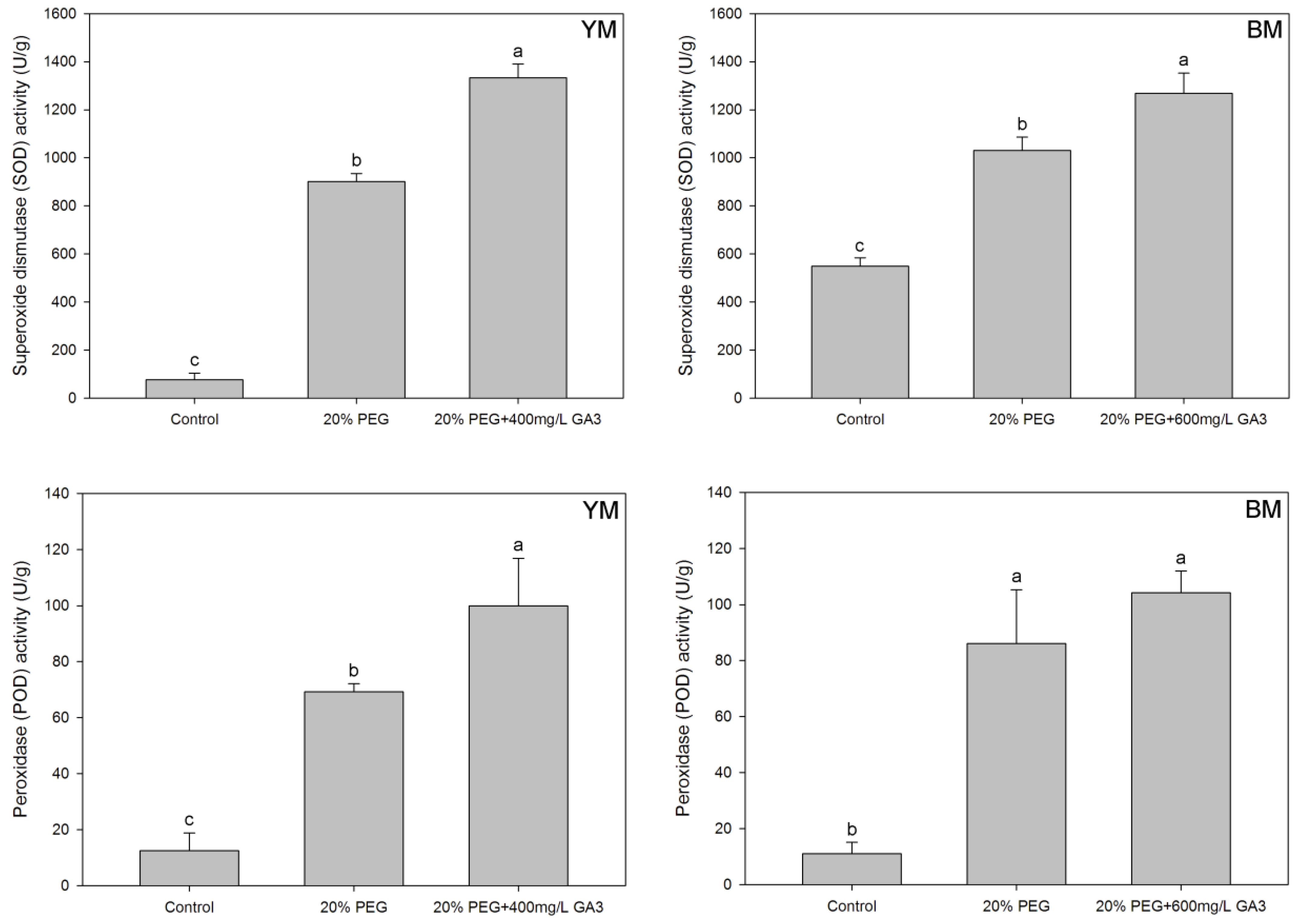
3.2.3. Effects of Seed Pre-Treatment by GA3 on Antioxidant Enzymes Activity in Hemp Seedlings under Drought Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bouloc, P.; Allegret, S.; Arnaud, L. Hemp: Industrial Production and Uses; CABI: Wallingford, UK, 2013. [Google Scholar]
- Chandra, S.; Hemant, L.; Mahmoud, A.E. Cannabis sativa L.-Botany and Biotechnology; Springer International Publishing: Berlin, Germany, 2017. [Google Scholar]
- Dai, A. Drought under global warming: A review. WIRES Clim. Chang. 2011, 2, 45–65. [Google Scholar] [CrossRef] [Green Version]
- Fernando, N.; Humphries, T.; Florentine, S.K.; Chauhan, B.S. Factors affecting seed germination of feather fingergrass (Chloris virgata). Weed Sci. 2016, 64, 605–612. [Google Scholar] [CrossRef]
- Liu, F.; Yang, M.; Du, G.; Long, B.; He, F.; Liu, H.; Yang, Y. Foundation and Application of Industrial Hemp; Science Press: Beijing, China, 2015. [Google Scholar]
- Amaducci, S.; Scordia, D.; Liu, F.H.; Zhang, Q.; Guo, H.; Testa, G.; Cosentino, S.L. Key cultivation techniques for hemp in Europe and China. Ind. Crops Prod. 2015, 68, 2–16. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Kamiya, Y. Gibberellins and light-stimulated seed germination. J. Plant Growth Regul. 2001, 20, 369–376. [Google Scholar] [CrossRef]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Kaur, S.; Gupta, A.K.; Kaur, N. Gibberellic acid and kinetin partially reverse the effect of water stress on germination and seedling growth in chickpea. Plant Growth Regul. 1998, 25, 29–33. [Google Scholar] [CrossRef]
- Kaur, S.; Gupta, A.K.; Kaur, N. Effect of GA3, kinetin and indole acetic acid on carbohydrate metabolism in chickpea seedlings germinating under water stress. Plant Growth Regul. 2000, 30, 61–70. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Q.; Wang, G.X.; Wang, X.D.; Guo, J.Y. Germination, osmotic adjustment, and antioxidant enzyme activities of gibberellin-pretreated Picea asperata seeds under water stress. New For. 2010, 39, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Ansari, O.; Azadi, M.S.; Sharif-Zadeh, F.; Sharif-Zadeh, F.; Younesi, E. Effect of hormone priming on germination characteristics and enzyme activity of mountain rye (Secale montanum) seeds under drought stress conditions. J. Stress Physiol. Biochem. 2013, 9, 61–71. [Google Scholar]
- Shariatmadari, M.H.; Parsa, M.; Nezami, A.; Kafi, M. The effects of hormonal priming on emergence, growth and yield of chickpea under drought stress in glasshouse and field. Biosci. Res. 2017, 14, 34–41. [Google Scholar]
- Li, G.; Wang, Q.; Li, X.; Zhang, Y.; Zhu, J.; Du, G.; Liu, F. Effects of gibberellin and Vc soaking seeds on the seedling physiology of hemp seeds at the initial stage of germination under drought stress. Seed 2018, 37, 67–71. [Google Scholar]
- Du, G.; Zhou, B.; Yang, Y.; Deng, G.; Liu, F. Evaluation of drought stress of hemp at seed germination stage under PEG simulation. Chin. Agric. Sci. Bull. 2015, 31, 147–153. [Google Scholar]
- Blandinières, H.; Leoni, M.; Ferrarini, A.; Amaducci, S. Ranking 26 European hemp (Cannabis sativa L.) cultivars for osmotic stress tolerance and transpiration efficiency. Ind. Crops Prod. 2021, 170, 113774. [Google Scholar] [CrossRef]
- Hu, H.; Liu, H.; Liu, F. Seed germination of hemp (Cannabis sativa L.) cultivars responds differently to the stress of salt type and concentration. Ind. Crops Prod. 2018, 123, 254–261. [Google Scholar] [CrossRef]
- Wang, X.K. Experimental Principle and Technology of Plant Physiology and Biochemistry, 3rd ed.; Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Hegarty, T.W. Seed activation and seed germination under moisture stress. New Phytol. 1977, 78, 349–359. [Google Scholar] [CrossRef]
- Florido, M.; Bao, L.; Lara, R.M.; Castro, Y.; Acosta, R.; Álvarez, M. Effect of water stress simulated with peg 6000 on tomato seed germination (Solanum section Lycopersicon). Cultiv. Trop. 2018, 39, 87–92. [Google Scholar]
- Partheeban, C.; Chandrasekhar, C.N.; Jeyakumar, P.; Ravikesavan, R.; Gnanam, R. Effect of PEG induced drought stress on seed germination and seedling characters of maize (Zea mays L.) genotypes. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1095–1104. [Google Scholar] [CrossRef] [Green Version]
- Channaoui, S.; El Kahkahi, R.; Charafi, J.; Mazouz, H.; El Fechtali, M.; Nabloussi, A. Germination and seedling growth of a set of rapeseed (Brassica napus) varieties under drought stress conditions. Int. J. Environ. Agric. Biotechnol. 2017, 2, 487–494. [Google Scholar] [CrossRef]
- Karssen, C.M.; Zagorski, S.; Kepczynski, J.; Groot, S.P.C. Key role for endogenous gibberellins in the control of seed germination. Ann. Bot.-Lond. 1989, 63, 71–80. [Google Scholar] [CrossRef]
- Heikal, M.M.; Shaddad, M.A.; Ahmed, A.M. Effect of water stress and gibberellic acid on germination of flax, sesame and onion seeds. Biol. Plant. 1982, 24, 124–129. [Google Scholar] [CrossRef]
- Edelstein, M.; Ben Tal, Y.; Wodner, M.; Kigel, J. Role of endogenous gibberellins in germination of melon (Cucumis melo) seeds. Physiol. Plant. 1995, 95, 113–119. [Google Scholar] [CrossRef]
- Jones, R.L.; Carbonell, J. Regulation of the synthesis of barley aleurone α-amylase by gibberellic acid and calcium ions. Plant Physiol. 1984, 76, 213–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, C.; Wang, Y. Exogenous salicylic acid-induced nitric oxide regulates leaf water condition through root osmoregulation of maize seedlings under drought stress. Braz. J. Bot. 2017, 40, 591–597. [Google Scholar] [CrossRef]
- Ai, L.; Li, Z.H.; Xie, Z.X.; Tian, X.L.; Eneji, A.E.; Duan, L.S. Coronatine alleviates polyethylene glycol-induced water stress in two rice (Oryza sativa L.) cultivars. J. Agron. Crop Sci. 2008, 194, 360–368. [Google Scholar] [CrossRef]
- McDonald, M.B. Seed deterioration: Physiology, repair and assessment. Seed Sci. Technol. 1999, 27, 177–237. [Google Scholar]
- Hu, H.; Liu, H.; Du, G.; Yang, F.; Deng, G.; Yang, Y.; Liu, F. Fiber and seed type of hemp (Cannabis sativa L.) responded differently to salt-alkali stress in seedling growth and physiological indices. Ind. Crops Prod. 2019, 129, 624–630. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Effect | Cultivar | Treatment | Cultivar × Treatment | |||
|---|---|---|---|---|---|---|
| Parameter | df | F-Value | df | F-Value | df | F-Value |
| Germination potential | 1 | 75.294 *** | 6 | 37.353 *** | 6 | 4.718 ** |
| Germination rate | 1 | 75.409 *** | 6 | 36.828 *** | 6 | 4.094 ** |
| Hypocotyl length | 1 | 36.973 *** | 6 | 114.749 *** | 6 | 14.576 *** |
| Radicle length | 1 | 65.094 *** | 6 | 29.768 *** | 6 | 3.317 * |
| Cultivar | Treatment | Germination Potential (%) | Germination Rate (%) | Hypocotyl Length (cm) | Radicle Length (cm) |
|---|---|---|---|---|---|
| YM | Control | 61.11 + 3.85 a | 86.67 + 3.33 a | 5.31 + 0.41 a | 5.26 + 0.32 a |
| 20% PEG | 16.67 + 3.33 d | 35.56 + 1.93 e | 1.09 + 0.23 e | 3.42 + 0.59 d | |
| 20% PEG + 0 mg/L GA3 | 37.78 + 3.85 c | 53.33 + 5.77 d | 1.65 + 0.14 d | 3.52 + 0.31 d | |
| 20% PEG + 200 mg/L GA3 | 42.22 + 8.39 c | 70.00 + 6.67 bc | 1.83 + 0.22 d | 3.87 + 0.11 cd | |
| 20% PEG + 400 mg/L GA3 | 52.22 + 1.93 ab | 78.89 + 5.09 ab | 2.89 + 0.12 b | 4.79 + 0.04 ab | |
| 20% PEG + 600 mg/L GA3 | 46.67 + 5.77 bc | 71.11 + 1.93 bc | 2.51 + 0.19 bc | 4.35 + 0.41 bc | |
| 20% PEG + 800 mg/L GA3 | 43.33 + 6.67 bc | 68.89 + 6.94 c | 2.41 + 0.34 c | 3.94 + 0.17 cd | |
| BM | Control | 51.11 + 5.09 a | 80.00 + 6.67 a | 3.33 + 0.41 a | 5.42 + 0.89 a |
| 20% PEG | 18.89 + 3.85 e | 31.11 + 6.94 d | 1.35 + 0.06 d | 1.99 + 0.37 d | |
| 20% PEG + 0 mg/L GA3 | 21.11 + 3.85 de | 40.00 + 11.55 cd | 1.44 + 0.22 d | 2.18 + 0.61 d | |
| 20% PEG + 200 mg/L GA3 | 24.44 + 1.93 cde | 44.44 + 7.70 cd | 1.58 + 0.11 d | 2.70 + 0.26 cd | |
| 20% PEG + 400 mg/L GA3 | 27.78 + 5.09 cd | 46.67 + 5.77 bc | 2.28 + 0.23 bc | 3.25 + 0.24 bc | |
| 20% PEG + 600 mg/L GA3 | 37.78 + 5.09 b | 58.89 + 3.85 b | 2.54 + 0.14 b | 3.78 + 0.24 b | |
| 20% PEG + 800 mg/L GA3 | 30.00 + 3.33 c | 45.56 + 6.94 bc | 2.11 + 0.10 c | 2.74 + 0.29 cd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, G.; Zhang, H.; Yang, Y.; Zhao, Y.; Tang, K.; Liu, F. Effects of Gibberellin Pre-Treatment on Seed Germination and Seedling Physiology Characteristics in Industrial Hemp under Drought Stress Condition. Life 2022, 12, 1907. https://doi.org/10.3390/life12111907
Du G, Zhang H, Yang Y, Zhao Y, Tang K, Liu F. Effects of Gibberellin Pre-Treatment on Seed Germination and Seedling Physiology Characteristics in Industrial Hemp under Drought Stress Condition. Life. 2022; 12(11):1907. https://doi.org/10.3390/life12111907
Chicago/Turabian StyleDu, Guanghui, Hanxue Zhang, Yang Yang, Yinhong Zhao, Kailei Tang, and Feihu Liu. 2022. "Effects of Gibberellin Pre-Treatment on Seed Germination and Seedling Physiology Characteristics in Industrial Hemp under Drought Stress Condition" Life 12, no. 11: 1907. https://doi.org/10.3390/life12111907