
Major Surgical Trauma Impairs the Function of Natural Killer Cells but Does Not Affect Monocyte Cytokine Synthesis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Study Design
2.2. Isolation and Culture of Mononuclear Cells
2.3. Flow Cytometry
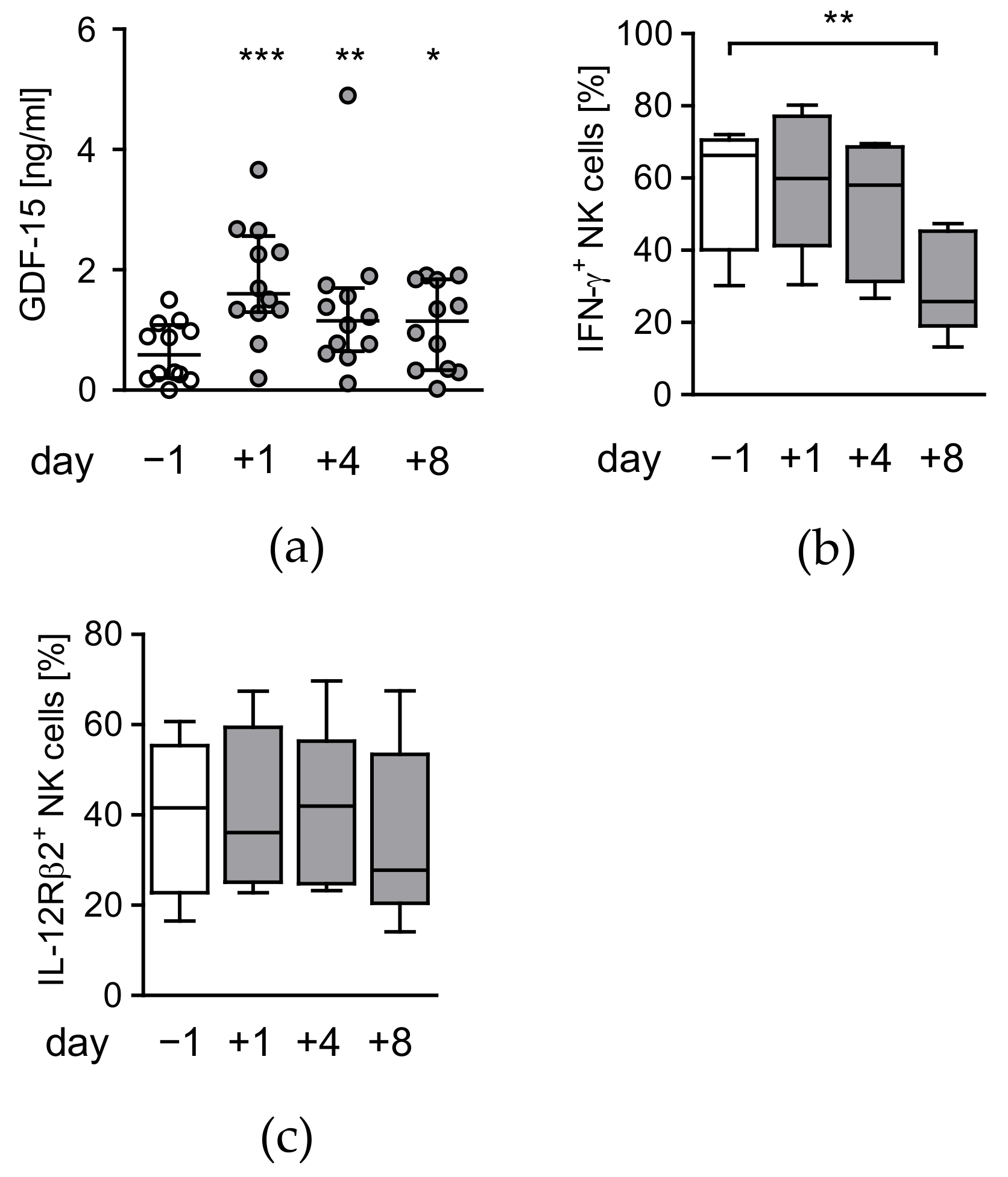
2.4. Quantification of GDF-15
2.5. Statistical Analyses
3. Results
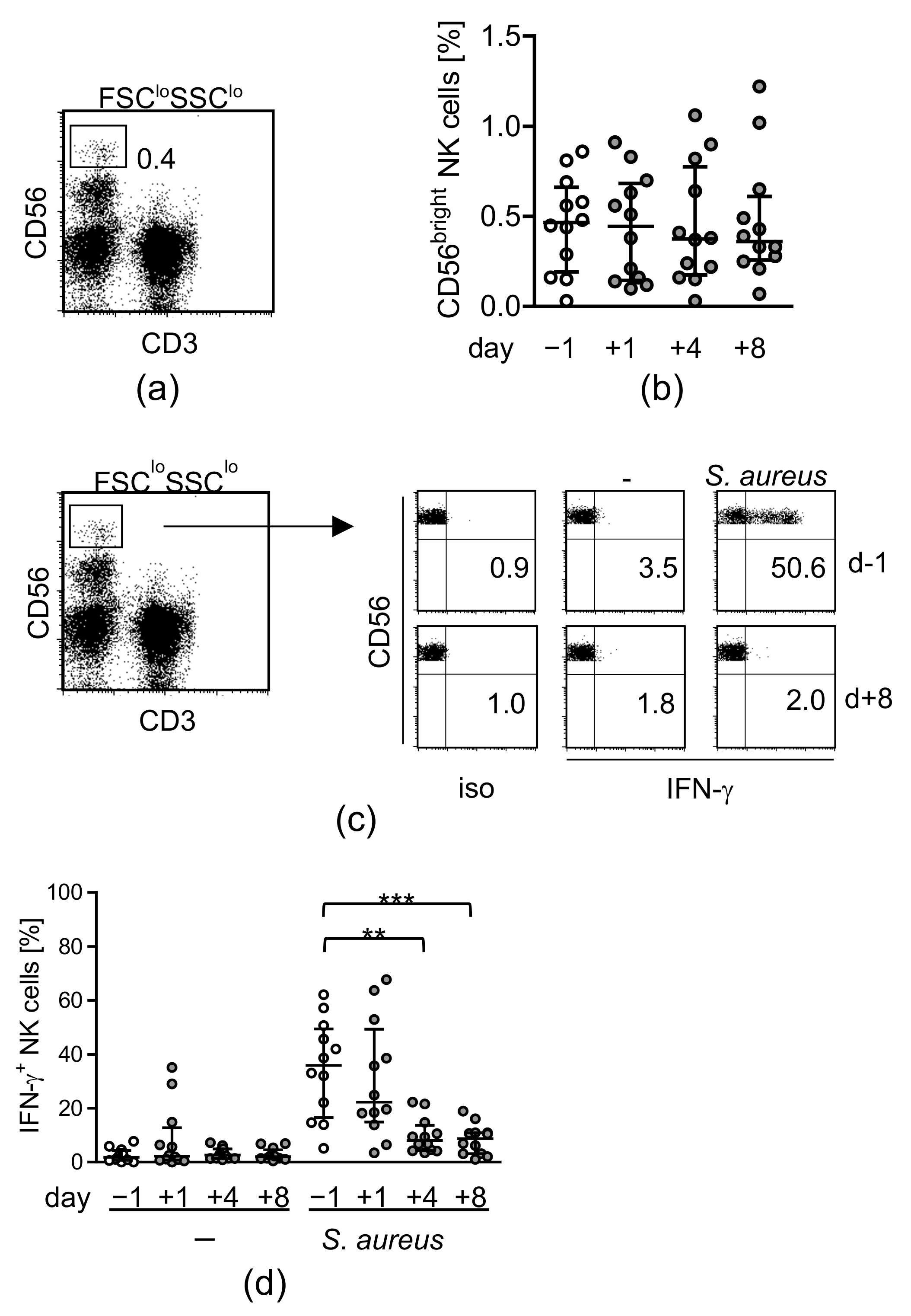
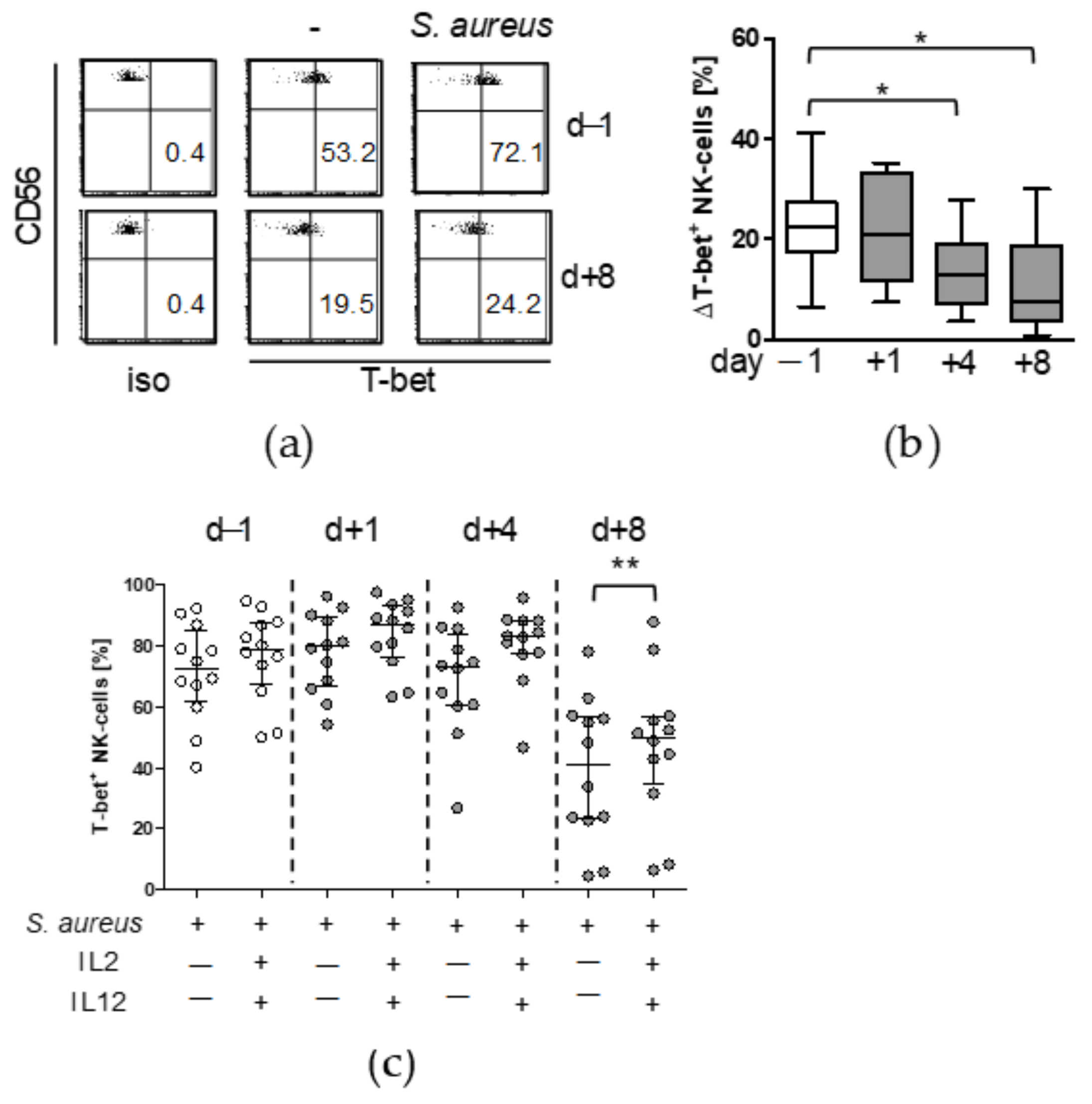
3.1. Major Spine Surgery Impairs the NK Cell Response to S. aureus
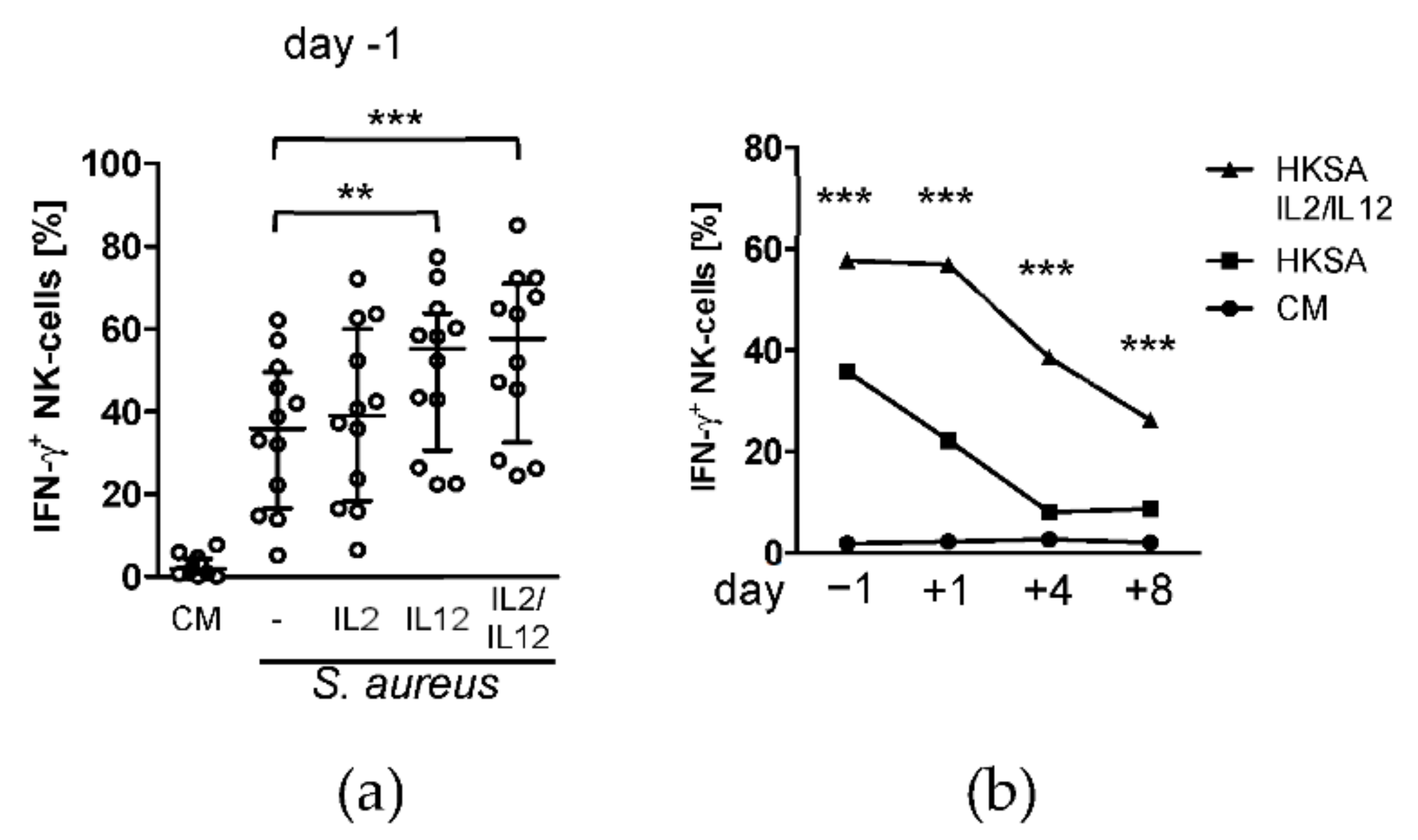
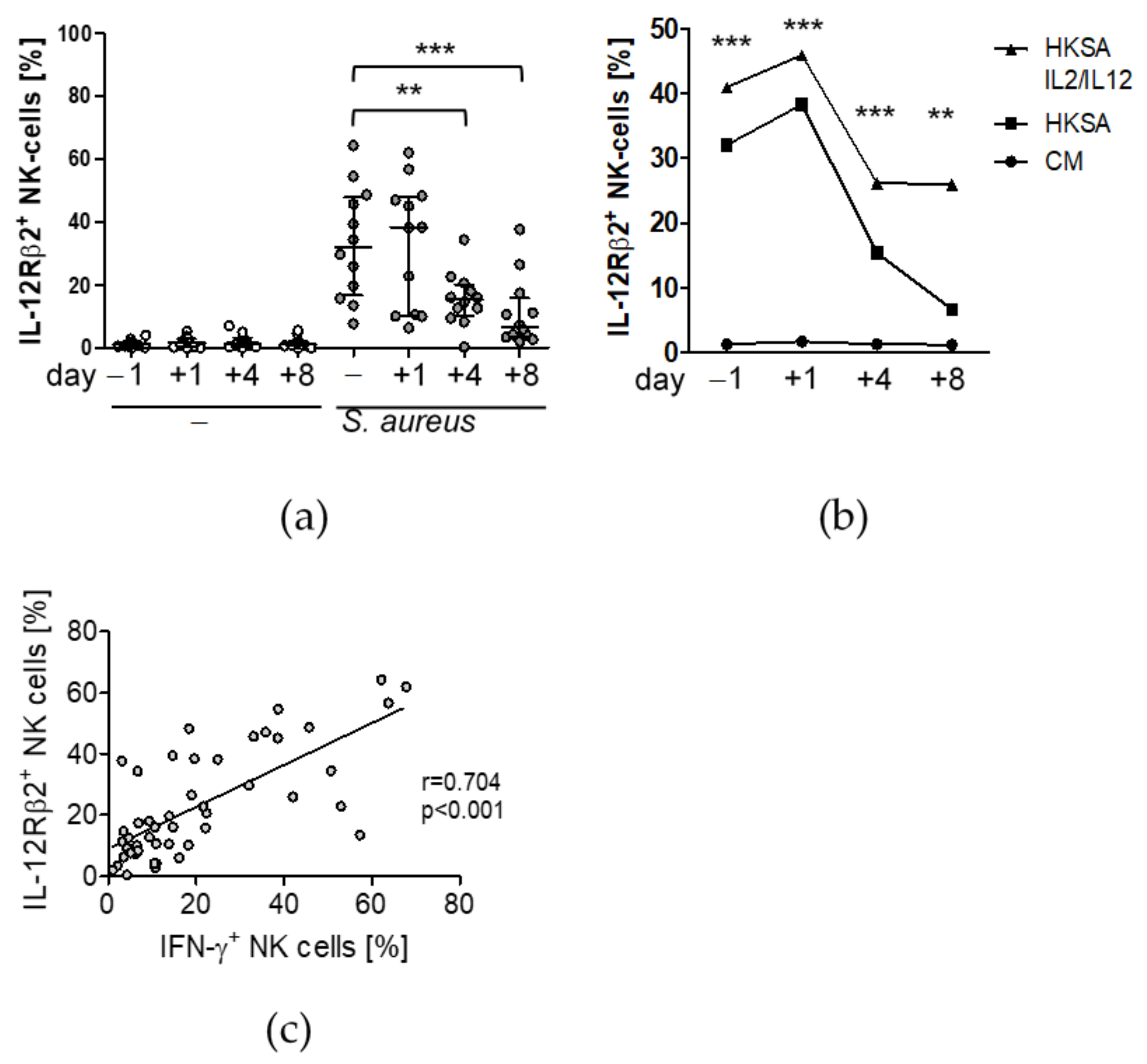
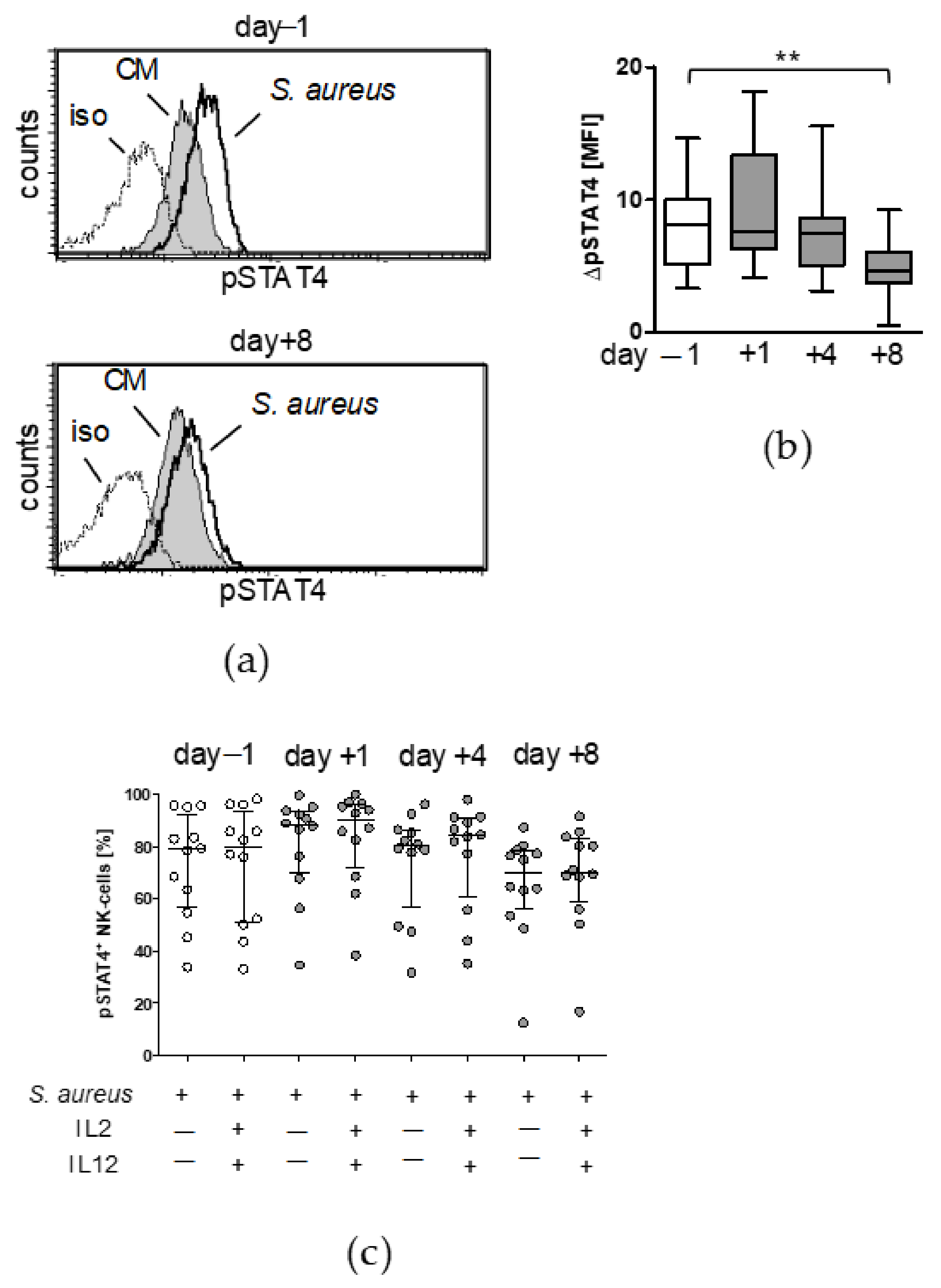
3.2. Treatment with IL-12 Rescues NK Cell Activity after Surgery
3.3. Circulating Factors Contribute to Late NK Cell Suppression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lord, J.M.; Midwinter, M.J.; Chen, Y.F.; Belli, A.; Brohi, K.; Kovacs, E.J.; Koenderman, L.; Kubes, P.; Lilford, R.J. The systemic immune response to trauma: An overview of pathophysiology and treatment. Lancet 2014, 384, 1455–1465. [Google Scholar] [CrossRef] [Green Version]
- Caligiuri, M.A. Human natural killer cells. Blood 2008, 112, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Michel, T.; Theresine, M.; Andres, E.; Hentges, F.; Zimmer, J. CD56bright natural killer (NK) cells: An important NK cell subset. Immunology 2009, 126, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Lodoen, M.B.; Lanier, L.L. Natural killer cells as an initial defense against pathogens. Curr. Opin. Immunol. 2006, 18, 391–398. [Google Scholar] [CrossRef]
- Michel, T.; Hentges, F.; Zimmer, J. Consequences of the crosstalk between monocytes/macrophages and natural killer cells. Front. Immunol. 2013, 3, 403. [Google Scholar] [CrossRef] [Green Version]
- Thomas, R.; Yang, X. NK-DC Crosstalk in Immunity to Microbial Infection. J. Immunol. Res. 2016, 2016, 6374379. [Google Scholar] [CrossRef] [Green Version]
- Thierfelder, W.E.; van Deursen, J.M.; Yamamoto, K.; Tripp, R.A.; Sarawar, S.R.; Carson, R.T.; Sangster, M.Y.; Vignali, D.A.; Doherty, P.C.; Grosveld, G.C.; et al. Requirement for Stat4 in interleukin-12-mediated responses of natural killer and T cells. Nature 1996, 382, 171–174. [Google Scholar] [CrossRef]
- Jacobson, N.G.; Szabo, S.J.; Weber-Nordt, R.M.; Zhong, Z.; Schreiber, R.D.; Darnell, J.E., Jr.; Murphy, K.M. Interleukin 12 signaling in T helper type 1 (Th1) cells involves tyrosine phosphorylation of signal transducer and activator of transcription (Stat)3 and Stat4. J. Exp. Med. 1995, 181, 1755–1762. [Google Scholar] [CrossRef]
- Afkarian, M.; Sedy, J.R.; Yang, J.; Jacobson, N.G.; Cereb, N.; Yang, S.Y.; Murphy, T.L.; Murphy, K.M. T-bet is a STAT1-induced regulator of IL-12R expression in naive CD4+ T cells. Nat. Immunol. 2002, 3, 549–557. [Google Scholar] [CrossRef]
- Wang, K.S.; Frank, D.A.; Ritz, J. Interleukin-2 enhances the response of natural killer cells to interleukin-12 through up-regulation of the interleukin-12 receptor and STAT4. Blood 2000, 95, 3183–3190. [Google Scholar] [CrossRef]
- Lucas, M.; Schachterle, W.; Oberle, K.; Aichele, P.; Diefenbach, A. Dendritic cells prime natural killer cells by trans-presenting interleukin 15. Immunity 2007, 26, 503–517. [Google Scholar] [CrossRef] [Green Version]
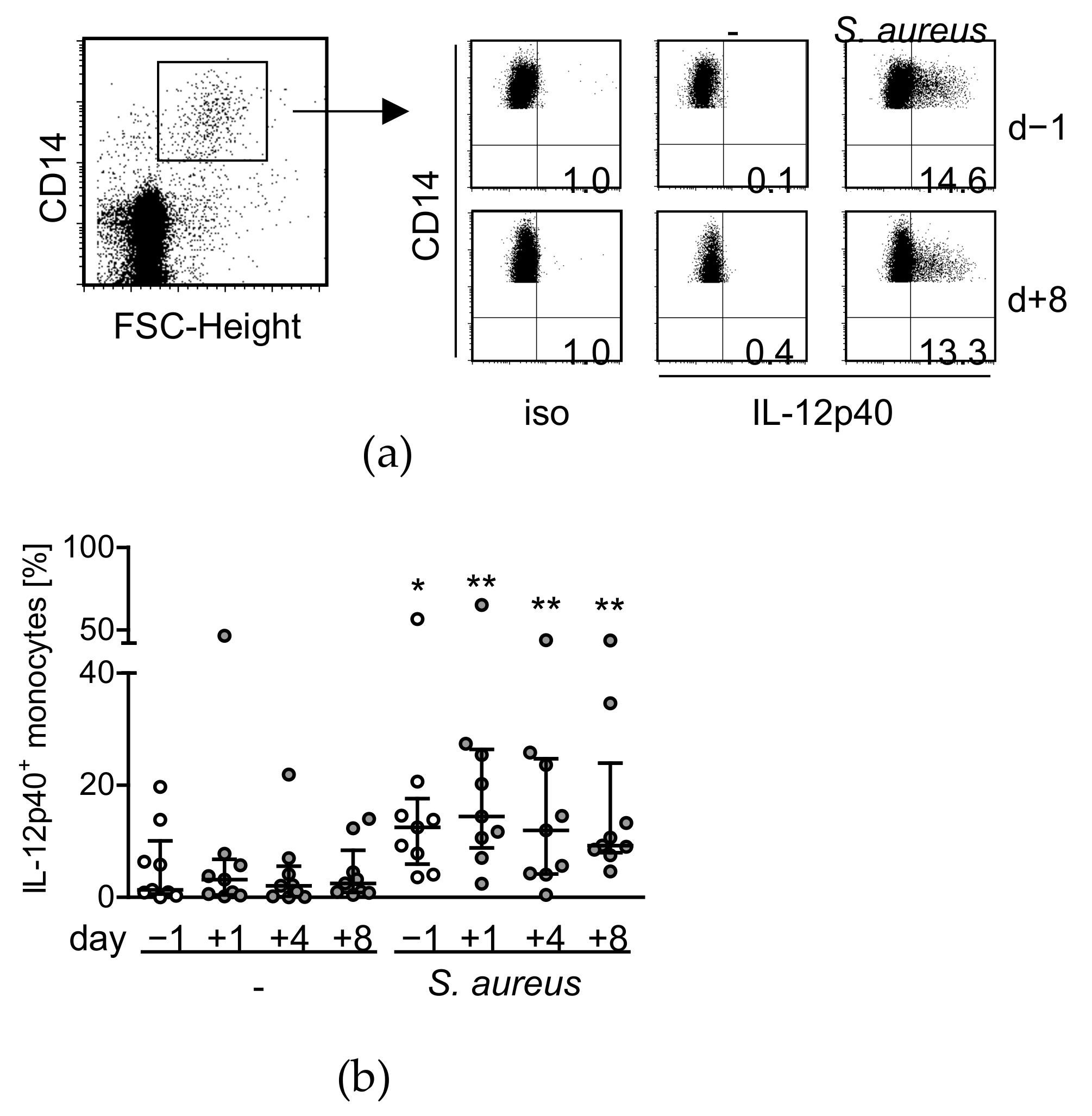
- Spolarics, Z.; Siddiqi, M.; Siegel, J.H.; Garcia, Z.C.; Stein, D.S.; Denny, T.; Deitch, E.A. Depressed interleukin-12-producing activity by monocytes correlates with adverse clinical course and a shift toward Th2-type lymphocyte pattern in severely injured male trauma patients. Crit. Care Med. 2003, 31, 1722–1729. [Google Scholar] [CrossRef]
- Kleinertz, H.; Hepner-Schefczyk, M.; Ehnert, S.; Claus, M.; Halbgebauer, R.; Boller, L.; Huber-Lang, M.; Cinelli, P.; Kirschning, C.; Flohé, S.; et al. Circulating growth/differentiation factor 15 is associated with human CD56 (bright) natural killer cell dysfunction and nosocomial infection in severe systemic inflammation. EBioMedicine 2019, 43, 380–391. [Google Scholar] [CrossRef] [Green Version]
- Pull ter Gunne, A.F.; Cohen, D.B. Incidence, prevalence, and analysis of risk factors for surgical site infection following adult spinal surgery. Spine 2009, 34, 1422–1428. [Google Scholar] [CrossRef]
- Street, J.T.; Lenehan, B.J.; DiPaola, C.P.; Boyd, M.D.; Kwon, B.K.; Paquette, S.J.; Dvorak, M.F.; Rampersaud, Y.R.; Fisher, C.G. Morbidity and mortality of major adult spinal surgery. A prospective cohort analysis of 942 consecutive patients. Spine J. 2012, 12, 22–34. [Google Scholar] [CrossRef]
- Reinhardt, R.; Pohlmann, S.; Kleinertz, H.; Hepner-Schefczyk, M.; Paul, A.; Flohé, S.B. Invasive Surgery Impairs the Regulatory Function of Human CD56 bright Natural Killer Cells in Response to Staphylococcus aureus. Suppression of Interferon-gamma Synthesis. PLoS ONE 2015, 10, e0130155. [Google Scholar] [CrossRef]
- Cheng, M.; Nguyen, M.H.; Fantuzzi, G.; Koh, T.J. Endogenous interferon-gamma is required for efficient skeletal muscle regeneration. Am. J. Physiol. Cell Physiol. 2008, 294, C1183–C1191. [Google Scholar] [CrossRef] [Green Version]
- Panduro, M.; Benoist, C.; Mathis, D. Treg cells limit IFN-gamma production to control macrophage accrual and phenotype during skeletal muscle regeneration. Proc. Natl. Acad. Sci. USA 2018, 115, E2585–E2593. [Google Scholar] [CrossRef] [Green Version]
- Kimura, F.; Shimizu, H.; Yoshidome, H.; Ohtsuka, M.; Miyazaki, M. Immunosuppression following surgical and traumatic injury. Surg. Today 2010, 40, 793–808. [Google Scholar] [CrossRef]
- Hensler, T.; Heidecke, C.D.; Hecker, H.; Heeg, K.; Bartels, H.; Zantl, N.; Wagner, H.; Siewert, J.R.; Holzmann, B. Increased susceptibility to postoperative sepsis in patients with impaired monocyte IL-12 production. J. Immunol. 1998, 161, 2655–2659. [Google Scholar]
- Carson, W.E.; Yu, H.; Dierksheide, J.; Pfeffer, K.; Bouchard, P.; Clark, R.; Durbin, J.; Baldwin, A.S.; Peschon, J.; Johnson, P.R.; et al. A fatal cytokine-induced systemic inflammatory response reveals a critical role for NK cells. J. Immunol. 1999, 162, 4943–4951. [Google Scholar]
- Cheron, A.; Floccard, B.; Allaouchiche, B.; Guignant, C.; Poitevin, F.; Malcus, C.; Crozon, J.; Faure, A.; Guillaume, C.; Marcotte, G.; et al. Lack of recovery in monocyte human leukocyte antigen-DR expression is independently associated with the development of sepsis after major trauma. Crit. Care 2010, 14, R208. [Google Scholar] [CrossRef] [Green Version]
- Gouel-Chéron, A.; Allaouchiche, B.; Guignant, C.; Davin, F.; Floccard, B.; Monneret, G. Early interleukin-6 and slope of monocyte human leukocyte antigen-DR: A powerful association to predict the development of sepsis after major trauma. PLoS ONE 2012, 7, e33095. [Google Scholar] [CrossRef] [Green Version]
- Galbraith, N.; Walker, S.; Carter, J.; Polk, H.C., Jr. Past, Present, and Future of Augmentation of Monocyte Function in the Surgical Patient. Surg. Infect. 2016, 17, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Heringlake, M.; Charitos, E.I.; Erber, K.; Berggreen, A.E.; Heinze, H.; Paarmann, H. Preoperative plasma growth-differentiation factor-15 for prediction of acute kidney injury in patients undergoing cardiac surgery. Crit. Care 2016, 20, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients (n = 12) | |
|---|---|
| age (y) | 40 (19–67) 1 |
| sex (m/f) | 2/10 |
| no. of fused segments | 7 (3–11) |
| co-morbidities | |
| diabetes (n, %) | 1/8.3 |
| cardiovascular (n, %) | 3/25 |
| autoimmunity | 2/16.6 |
| smoker | 0/0 |
| duration of surgery (h) | 6 (5–7) |
| blood transfusion (mL) | 500 (0–1750) |
| ICU length of stay (d) | 1 (1–2) |
| Patients (n = 12) | |
|---|---|
| Leukocytes (x103/µL) | |
| d − 1 | 7.9 (6.3–10.4) 1 |
| d + 1 | 10.4 (7.5–12.5) *** |
| CRP (ng/dL) | |
| d + 1 | 4.4 (4.1–5.7) ### |
| d + 2 | 18.4 (14.6–22.0) ### |
| d + 4 | 9.1 (5.3–17.7) ### |
| IL-6 (d + 1) (pg/dL) | 212 (118–273) ### |
| myoglobin (d + 1) (ng/mL) | 928 (557–1693) ### |
| Correlation of GDF-15 on d − 1 with | Spearman r | p-Value |
|---|---|---|
| IFN-γ d − 1 | −0.7622 | 0.006 |
| IFN-γ d + 1 | −0.5455 | 0.071 |
| IFN-γ d + 4 | −0.6573 | 0.024 |
| IFN-γ d + 8 | −0.2727 | 0.391 |
| LOS1 | 0.6679 | 0.021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller-Heck, R.M.; Bösken, B.; Michiels, I.; Dudda, M.; Jäger, M.; Flohé, S.B. Major Surgical Trauma Impairs the Function of Natural Killer Cells but Does Not Affect Monocyte Cytokine Synthesis. Life 2022, 12, 13. https://doi.org/10.3390/life12010013
Müller-Heck RM, Bösken B, Michiels I, Dudda M, Jäger M, Flohé SB. Major Surgical Trauma Impairs the Function of Natural Killer Cells but Does Not Affect Monocyte Cytokine Synthesis. Life. 2022; 12(1):13. https://doi.org/10.3390/life12010013
Chicago/Turabian StyleMüller-Heck, Roman M., Björn Bösken, Ivo Michiels, Marcel Dudda, Marcus Jäger, and Stefanie B. Flohé. 2022. "Major Surgical Trauma Impairs the Function of Natural Killer Cells but Does Not Affect Monocyte Cytokine Synthesis" Life 12, no. 1: 13. https://doi.org/10.3390/life12010013