
Molecular Analysis of DPY19L2, PICK1 and SPATA16 in Italian Unrelated Globozoospermic Men

 , , ,
, , ,
Abstract
:1. Introduction
- PICK1 (protein interacting with C kinase 1) gene is located on chromosome 22q13.1. It encodes for a membrane protein paramount for protein and vesicle trafficking. Deletion of this gene leads to round-headed spermatozoa and oligozoospermia. Liu et al. [17] discovered a homozygous missense mutation (G198A) in exon 13 of the PICK1 gene in a Chinese family. The family members affected by this homozygous missense mutation showed infertility caused by the absence of the acrosome.
- SPATA16 (spermatogenesis associated 16) gene is located on chromosome 3q26.31. The encoded protein is involved in acrosome biogenesis during proacrosomal vesicle transport. A homozygous mutation in this spermatogenesis-specific gene was identified in a consanguineous family [18].
- DPY19L2 (dpy-19-like 2 (C. elegans)) gene is located on chromosome 12q14.2 and it is considered the main gene involved in the etiopathogenesis of human globozoospermia [19,20,21]. It encodes for a protein involved in anchoring the acrosome to the spermatozoa nucleus [22]. Patients without DPY19L2 have normal or subnormal sperm concentration, indicating that this gene plays a role in spermiogenesis but not in germ cell proliferation or meiosis [20,22,23]. Furthermore, a correlation exists between the severity of the phenotype and oocyte fertilization and the type of DPY19L2 mutation [16,20,24,25].
2. Materials and Methods
2.1. Patients
2.2. Semen Analysis
2.3. DNA Extraction
2.4. PCR and Sequencing
- DPY19L2: we studied exons 1, 10, 11, 12, 20 and 22. Analysis was carried out with qualitative PCR followed by electrophoresis on 2% agarose gel.
- SPATA16: we analyzed exon 4. To detect any mutations, amplified samples were purified and underwent automated sequencing based on the Sanger method by using 3500 Genetic Analyser (Applied Biosystem, Waltham, MA, USA). Purification of samples was carried on with ®PureLink PCR purification kit (Invitrogen, Life Tecnologies, Waltham, MA, USA). Amplification reaction occurred in a volume of 13 µL containing 1 µL of ®Big Dye (Applied Biosystems, Waltham, MA, USA), 2 µL of buffer ®Big Dye Terminator (Applied Biosystems, Waltham, MA, USA), 1 µL of each primer, 18 ng of DNA and the amount of DNAse free water (Ambion®, Waltham, MA, USA) needed to reach the final reaction volume. Raw data from the capillary electrophoresis were analyzed by Sequencing Analysis (Applied Biosystems, Waltham, MA, USA). Subsequently, multiple alignment of the sequences versus the template was carried out to identify point mutations using the software Geneious Prime 2020.2 (Biomatters, Ltd. L2, Auckland, New Zealand).
- PICK1: we investigated exon 13 by sequencing as described above.
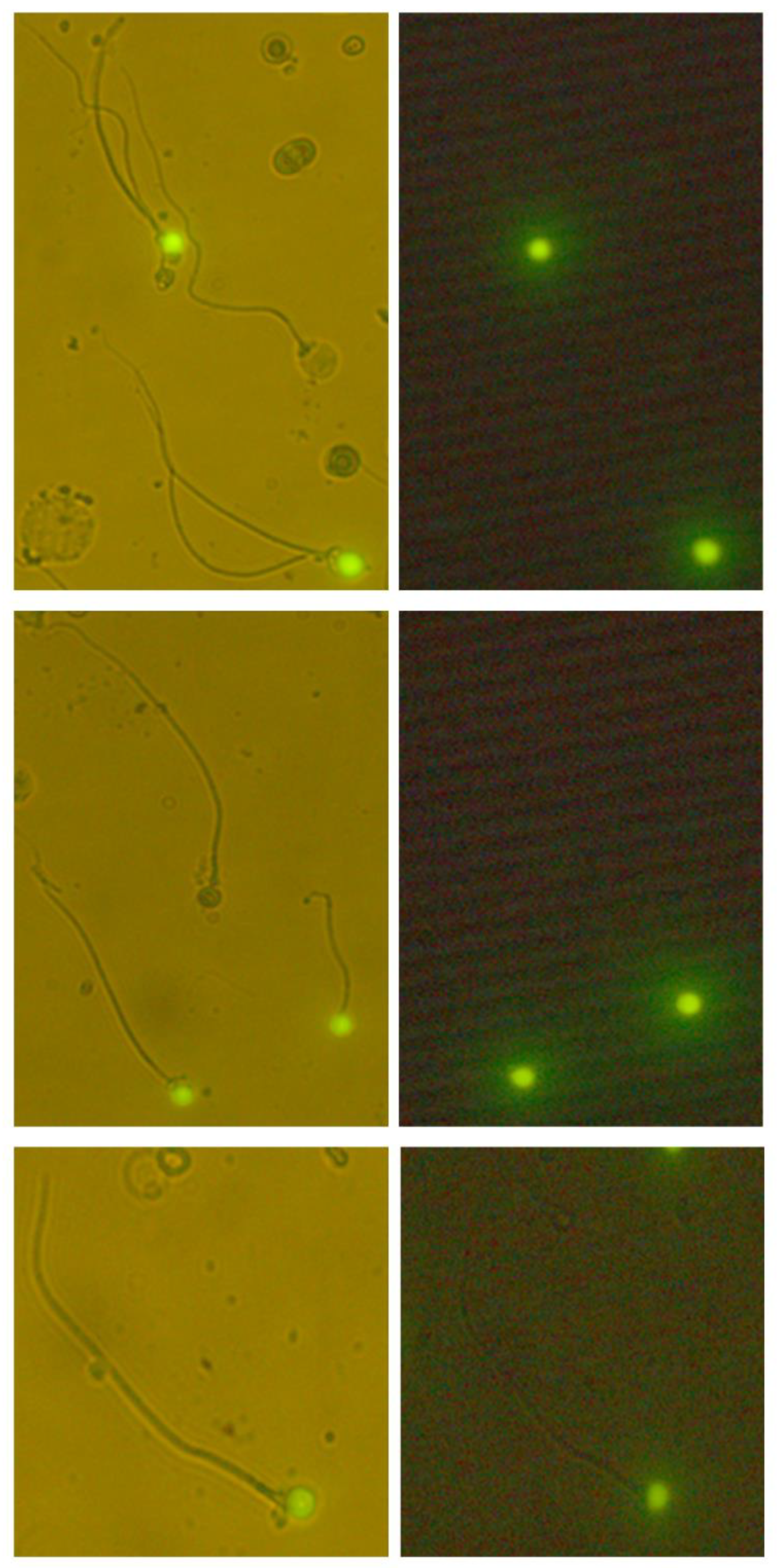
2.5. Sperm DNA Fragmentation
2.6. Statistical Analysis
3. Results
3.1. Semen Analysis
3.2. Genetic Analysis
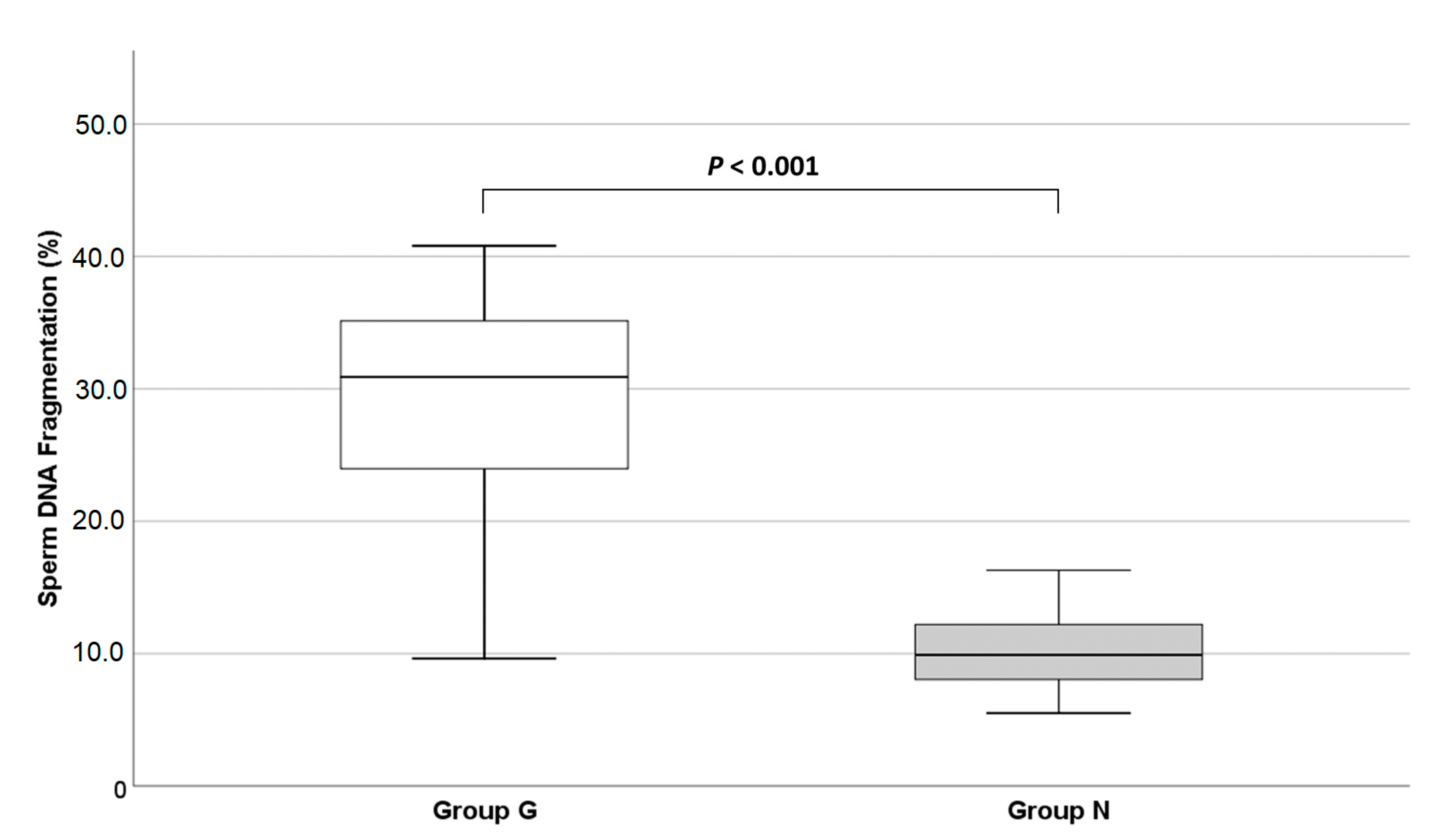
3.3. Sperm Chromatin Integrity
4. Discussion
4.1. Sperm Parameters
4.2. Genetic Analysis

{kind=link}
{kind=link}
| Gene | Exons Examined | Method to Identify Mutations | N° Globozoospermic Patients | N Patients Carrying Mutation | Reference |
|---|---|---|---|---|---|
| PICK1 | exon 13 | PCR and Sequencing | 3 members of a Chinese family | 1/3 homozygous mutated (c.1567G>A) | [17] |
| all exons | PCR and Sequencing | 1 Macedonian man | 0/1 | [50] | |
| exon 13 | qPCR | 27 Iranian men (of which 17 unrelated) | 0/27 | [48] | |
| all exons | PCR and Sequencing | 4 unrelated Tunisian men (with no DPY19L2 mutations) | 0/4 | [53] | |
| exon 13 | Sequencing | 18 unrelated Italian men | 0/18 | Present study | |
| SPATA16 | exon 4 | Genomewide scan analysis using a 10K SNP array | 3 brothers of a consanguineous Ashkenazi Jewish family | 3/3 homozygous mutated (c.848G>A) | [18] |
| all exons | PCR and Sequencing | 1 Macedonian man | 1/1 two polymorphisms (rs115897458 and rs508508) | [50] | |
| exon 2 | PCR and Sequencing | 19 (DPY19L2 undeleted) unrelated men originating from France, Italy, Tunisia, Turkey, Libya and Morocco | 2/19 (unrelated Tunisian men) deleted | [46] | |
| exon 4 | qPCR | 27 Iranian men (of which 17 unrelated) | 0/27 | [48] | |
| all exons | PCR and Sequencing | 4 unrelated Tunisian men (with no DPY19L2 mutations) | 0/4 | [53] | |
| exon 2 | PCR and Sequencing | 2 unrelated Tunisian men | 2/2 deleted | [47] | |
| exon 4 | Sequencing | 18 unrelated Italian men | 0/18 | Present study | |
| DPY19L2 | all exons | Whole genome SNP scan | 20 men (15 from Tunisia, 1 from Algeria, 2 from Morocco, 1 from Turkey and 1 from Slovenia), most of them first cousins | 15/20 homozygous deleted | [19] |
| exons 2, 7, 9, 10, 13, 17, 21 | Genome-wide scan analysis using 10K SNP arrays | 28 men (4 brothers from a Jordanian consanguineous family, 11 from France, 2 brothers from Algeria, 1 from Iran, 4 from Tunisia, 1 from Lybia, 1 from Italy, 1 from Morocco and 3 of undetermined origin) | 4 Jordanian brothers: homozygous deleted for all the exons examined 4 unrelated subjects deleted | [20] | |
| all exons | Multiplex Ligation-dependent Probe Amplification (MLPA) and Sequencing | 34 men from France and Tunisia (including 20 men described in Harbuz et al. 2011) | 23/34 (67.6%) homozygous deleted 2/34 (5.9%) heterozygous deleted 9/34 (26.4%) non-deleted Point mutations identified: - exon 8: heterozygous missense mutation (c.869G>A) - exon 9: heterozygous nonsense mutation (c.1024C>T) - exon 10: homozygous missense mutation (c.1073T>A) | [21] | |
| exons 4, 5, 6, 7, 8, 9, 10, 11, 15, 16, 21 | PCR | 54 genetically independent men for all types of mutations (from 13 different countries including Iran, France, Algeria, Turkey, Morocco, Belgium, USA, Italy) | 36/54 mutated (69.4%: homozygous deleted; 19.4%: heterozygous composite; 11.1%: homozygous point mutated) Point mutations identified: - exon 8: missense mutation (c.869G>A), non-synonymous mutation (c.892C>T) - exon 9: premature stop codon (c.1033C>T) - exon 15: non-synonymous mutation (c.1478C>G) - exon 21: premature stop codon (c.2038A>T) - exon 11: premature stop codon (c.1183delT) | [49] | |
| exons 1, 11, 22 | PCR and Sequencing | 2 Macedonian men | 2/2 homozygous deleted | [50] | |
| all exons | PCR and Sequencing | 15 unrelated Chinese men | 4/15 homozygous deleted | [51] | |
| all exons | qPCR | 9 men (7 from Italy and 2 from Spain) | 3/9 deleted (2 homozygous, 1 heterozygous) 1/9 wild-type 5/9 point mutated (4 missense, 3 intronic and 2 synonymous) | [24] | |
| exons 1, 17, 22 | PCR and Sequencing | 5 men from Algeria (of which 3 brothers) | 5/5 homozygous deleted | [52] | |
| exons 1, 5, 6, 7, 11, 22 | qPCR | 27 Iranian men (of which 17 unrelated) | 20/27 deleted | [48] | |
| all exons | PCR and Sequencing | 18 unrelated Tunisian men | 11/18: homozygous deleted in exon 10 2/18: homozygous for non-synonymous mutation (c.892C>T) in exon 8 1/18: homozygous for a new splice-site mutation at the junction exon–intron 16 | [53] | |
| exon 10 | PCR and Sequencing | 2 unrelated Tunisian men | 0/2 | [47] | |
| all exons | Whole-exome sequencing | 9 unrelated Chinese men | 5/9 deleted 4/9 with novel point mutations | [54] | |
| exons 1, 10, 11, 12, 20, 22 | PCR | 18 unrelated Italian men | 6/18 deleted (1/18 in exon 11; 1/18 in exon 22; 4/18 in exons 10, 12 and 22) | Present study |
4.3. Sperm Chromatin Integrity
4.4. Clinical Implications for Assisted Reproduction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pallotti, F.; Paoli, D.; Carlini, T.; Vestri, A.R.; Martino, G.; Lenzi, A.; Lombardo, F. Varicocele and semen quality: A retrospective case-control study of 4230 patients from a single centre. J. Endocrinol. Investig. 2018, 41, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Corona, G.; Sansone, A.; Pallotti, F.; Ferlin, A.; Pivonello, R.; Isidori, A.M.; Maggi, M.; Jannini, E.A. People smoke for nicotine, but lose sexual and reproductive health for tar: A narrative review on the effect of cigarette smoking on male sexuality and reproduction. J. Endocrinol. Investig. 2020, 43, 1391–1408. [Google Scholar] [CrossRef]
- Holstein, A.F.; Schirren, C.; Schirren, C.G. Human spermatids and spermatozoa lacking acrosomes. J. Reprod. Fertil. 1973, 35, 489–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dam, A.H.D.M.; Feenstra, I.; Westphal, J.R.; Ramos, L.; van Golde, R.J.; Kremer, J.A. Globozoospermia revisited. Hum. Reprod. Update 2007, 13, 63–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubino, P.; Viganò, P.; Luddi, A.; Piomboni, P. The ICSI procedure from past to future: A systematic review of the more controversial aspects. Hum. Reprod. Update 2016, 22, 194–227. [Google Scholar] [CrossRef] [Green Version]
- Chansel-Debordeaux, L.; Dandieu, S.; Bechoua, S.; Jimenez, C. Reproductive outcome in globozoospermic men: Update and prospects. Andrology 2015, 3, 1022–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirren, C.G.; Holstein, A.F.; Schirren, C. Über die Morphogenese rund-köpfiger Spermatozoen des Menschen. Andrologia 1971, 3, 117–125. [Google Scholar]
- Singh, G. Ultrastructural features of round-headed human spermatozoa. Int. J. Fertil. 1992, 37, 99–102. [Google Scholar]
- Dam, A.H.D.M.; Ramos, L.; Dijkman, H.B.; Woestenenk, R.; Robben, H.; van den Hoven, L.; Kremer, J.A. Morphology of partial globozoospermia. J. Androl. 2011, 32, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, Y.; Lescoat, D.; Le Lannou, D. Anomalous distribution of nuclear basic proteins in round-headed human spermatozoa. Andrologia 1990, 22, 549–555. [Google Scholar] [CrossRef]
- Yassine, S.; Escoffier, J.; Martinez, G.; Coutton, C.; Karaouzène, T.; Zouari, R.; Ravanat, J.-L.; Metzler-Guillemain, C.; Lee, H.C.; Fissore, R.; et al. Dpy19 l2-deficient globozoospermic sperm display altered genome packaging and DNA damage that compromises the initiation of embryo development. Mol. Hum. Reprod. 2015, 21, 169–185. [Google Scholar] [CrossRef] [Green Version]
- Vicari, E.; Perdichizzi, A.; De Palma, A.; Burrello, N.; D’Agata, R.; Calogero, A.E. Globozoospermia is associated with chromatin structure abnormalities: Case report. Hum. Reprod. 2002, 17, 2128–2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatimel, N.; Leandri, R.D.; Foliguet, B.; Bujan, L.; Parinaud, J. Sperm cephalic vacuoles: New arguments for their non acrosomal origin in two cases of total globozoospermia. Andrology 2013, 1, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Vozdova, M.; Rybar, R.; Kloudova, S.; Prinosilova, P.; Texl, P.; Rubes, J. Total globozoospermia associated with increased frequency of immature spermatozoa with chromatin defects and aneuploidy: A case report. Andrologia 2014, 46, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Perrin, A.; Coat, C.; Nguyen, M.H.; Talagas, M.; Morel, F.; Amice, J.; De Braekeleer, M. Molecular cytogenetic and genetic aspects of globozoospermia: A review. Andrologia 2013, 45, 1–9. [Google Scholar] [CrossRef] [PubMed]
- De Braekeleer, M.; Nguyen, M.H.; Morel, F.; Perrin, A. Genetic aspects of monomorphic teratozoospermia: A review. J. Assist. Reprod. Genet. 2015, 32, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Shi, Q.-W.; Lu, G.-X. A newly discovered mutation in PICK1 in a human with globozoospermia. Asian J. Androl. 2010, 12, 556–560. [Google Scholar] [CrossRef]
- Dam, A.H.D.M.; Koscinski, I.; Kremer, J.A.M.; Moutou, C.; Jaeger, A.-S.; Oudakker, A.R.; Tournaye, H.; Charlet, N.; Lagier-Tourenne, C.; van Bokhoven, H.; et al. Homozygous mutation in SPATA16 is associated with male infertility in human globozoospermia. Am. J. Hum. Genet. 2007, 81, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Harbuz, R.; Zouari, R.; Pierre, V.; Ben Khelifa, M.; Kharouf, M.; Coutton, C.; Merdassi, G.; Abada, F.; Escoffier, J.; Nikas, Y.; et al. A recurrent deletion of DPY19L2 causes infertility in man by blocking sperm head elongation and acrosome formation. Am. J. Hum. Genet. 2011, 88, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Koscinski, I.; Elinati, E.; Fossard, C.; Redin, C.; Muller, J.; Velez de la Calle, J.; Schmitt, F.; Ben Khelifa, M.; Ray, P.F.; Ray, P.; et al. DPY19L2 deletion as a major cause of globozoospermia. Am. J. Hum. Genet. 2011, 88, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Coutton, C.; Zouari, R.; Abada, F.; Ben Khelifa, M.; Merdassi, G.; Triki, C.; Escalier, D.; Hesters, L.; Mitchell, V.; Levy, R.; et al. MLPA and sequence analysis of DPY19L2 reveals point mutations causing globozoospermia. Hum. Reprod. 2012, 27, 2549–2558. [Google Scholar] [CrossRef] [Green Version]
- Pierre, V.; Martinez, G.; Coutton, C.; Delaroche, J.; Yassine, S.; Novella, C.; Pernet-Gallay, K.; Hennebicq, S.; Ray, P.F.; Arnoult, C. Absence of Dpy19l2, a new inner nuclear membrane protein, causes globozoospermia in mice by preventing the anchoring of the acrosome to the nucleus. Development 2012, 139, 2955–2965. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Li, L.; Tang, J.; Li, Y.; Lin, W.Y.; Martin, F.; Grant, D.; Solloway, M.; Parker, L.; Ye, W.; et al. A mouse knockout library for secreted and transmembrane proteins. Nat. Biotechnol. 2010, 28, 749–755. [Google Scholar] [CrossRef]
- Chianese, C.; Fino, M.G.; Riera Escamilla, A.; Lòpez Rodrigo, O.; Vinci, S.; Guarducci, E.; Daguin, F.; Muratori, M.; Tamburrino, L.; Lo Giacco, D.; et al. Comprehensive investigation in patients affected by sperm macrocephaly and globozoospermia. Andrology 2015, 3, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutton, C.; Escoffier, J.; Martinez, G.; Arnoult, C.; Ray, P.F. Teratozoospermia: Spotlight on the main genetic actors in the human. Hum. Reprod. Update 2015, 21, 455–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Carlini, T.; Paoli, D.; Pelloni, M.; Faja, F.; Dal Lago, A.; Lombardo, F.; Lenzi, A.; Gandini, L. Sperm DNA fragmentation in Italian couples with recurrent pregnancy loss. Reprod. Biomed. Online 2017, 34, 58–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoli, D.; Pecora, G.; Pallotti, F.; Faja, F.; Pelloni, M.; Lenzi, A.; Lombardo, F. Cytological and molecular aspects of the ageing sperm. Hum. Reprod. 2019, 34, 218–227. [Google Scholar] [CrossRef]
- Kang-Decker, N.; Mantchev, G.T.; Juneja, S.C.; McNiven, M.A.; van Deursen, J.M. Lack of acrosome formation in Hrb-deficient mice. Science 2001, 294, 1531–1533. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Ito, C.; Natsume, Y.; Sugitani, Y.; Yamanaka, H.; Kuretake, S.; Yanagida, K.; Sato, A.; Toshimori, K.; Noda, T. Lack of acrosome formation in mice lacking a Golgi protein, GOPC. Proc. Natl. Acad. Sci. USA 2002, 99, 11211–11216. [Google Scholar] [CrossRef] [Green Version]
- Xiao, N.; Kam, C.; Shen, C.; Jin, W.; Wang, J.; Lee, K.M.; Jiang, L.; Xia, J. PICK1 deficiency causes male infertility in mice by disrupting acrosome formation. J. Clin. Investig. 2009, 119, 802–812. [Google Scholar] [CrossRef] [Green Version]
- Fesahat, F.; Henkel, R.; Agarwal, A. Globozoospermia syndrome: An update. Andrologia 2020, 52, e13459. [Google Scholar] [CrossRef]
- Larson, K.L.; Brannian, J.D.; Singh, N.P.; Burbach, J.A.; Jost, L.K.; Hansen, K.P.; Kreger, D.O.; Evenson, D.P. Chromatin structure in globozoospermia: A case report. J. Androl. 2001, 22, 424–431. [Google Scholar] [PubMed]
- Ghasemzadeh, J.; Talebi, A.R.; Khalili, M.A.; Fesahat, F.; Halvaei, I.; Nabi, A.; Ashourzadeh, S. Sperm parameters, protamine deficiency, and apoptosis in total globozoospermia. Iran. J. Reprod. Med. 2015, 13, 495–502. [Google Scholar] [PubMed]
- Hosseinifar, H.; Yazdanikhah, S.; Modarresi, T.; Totonchi, M.; Sadighi Gilani, M.; Sabbaghian, M. Correlation between sperm DNA fragmentation index and CMA 3 positive spermatozoa in globozoospermic patients. Andrology 2015, 3, 526–531. [Google Scholar] [CrossRef] [Green Version]
- Eskandari, N.; Tavalaee, M.; Zohrabi, D.; Nasr-Esfahani, M.H. Association between total globozoospermia and sperm chromatin defects. Andrologia 2018, 50, e12843. [Google Scholar] [CrossRef]
- Talebi, A.R.; Ghasemzadeh, J.; Khalili, M.A.; Halvaei, I.; Fesahat, F. Sperm chromatin quality and DNA integrity in partial versus total globozoospermia. Andrologia 2018, 50, e12823. [Google Scholar] [CrossRef] [PubMed]
- Tavalaee, M.; Nomikos, M.; Lai, F.A.; Nasr-Esfahani, M.H. Expression of sperm PLCζ and clinical outcomes of ICSI-AOA in men affected by globozoospermia due to DPY19L2 deletion. Reprod. Biomed. Online 2018, 36, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Florke-Gerloff, S.; Topfer-Petersen, E.; Muller-Esterl, W.; Mansouri, A.; Schatz, R.; Schirren, C.; Schill, W.; Engelal, W. Biochemical and genetic investigation of round-headed spermatozoa in infertile men including two brothers and their father. Andrologia 1984, 16, 187–202. [Google Scholar] [CrossRef]
- Carrell, D.T.; Emery, B.R.; Liu, L. Characterization of aneuploidy rates, protamine levels, ultrastructure, and functional ability of roundheaded sperm from two siblings and implications for intracytoplasmic sperm injection. Fertil. Steril. 1999, 71, 511–516. [Google Scholar] [CrossRef]
- Carrell, D.T.; Wilcox, A.L.; Udoff, L.C.; Thorp, C.; Campbell, B. Chromosome 15 aneuploidy in the sperm and conceptus of a sibling with variable familial expression of round-headed sperm syndrome. Fertil. Steril. 2001, 76, 1258–1260. [Google Scholar] [CrossRef]
- Kilani, Z.; Ismail, R.; Ghunaim, S.; Mohamed, H.; Hughes, D.; Brewis, I.; Barratt, C.L.R. Evaluation and treatment of familial globozoospermia in five brothers. Fertil. Steril. 2004, 82, 1436–1439. [Google Scholar] [CrossRef]
- Dirican, E.K.; Isik, A.; Vicdan, K.; Sozen, E.; Suludere, Z. Clinical pregnancies and livebirths achieved by intracytoplasmic injection of round headed acrosomeless spermatozoa with and without oocyte activation in familial globozoospermia: Case report. Asian J. Androl. 2008, 10, 332–336. [Google Scholar] [CrossRef]
- Lu, L.; Lin, M.; Xu, M.; Zhou, Z.M.; Sha, J.H. Gene functional research using polyethylenimine-mediated in vivo gene transfection into mouse spermatogenic cells. Asian J. Androl. 2006, 8, 53–59. [Google Scholar] [CrossRef]
- Fujihara, Y.; Oji, A.; Larasati, T.; Kojima-Kita, K.; Ikawa, M. Human Globozoospermia-Related Gene Spata16 Is Required for Sperm Formation Revealed by CRISPR/Cas9-Mediated Mouse Models. Int. J. Mol. Sci. 2017, 18, 2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellnati, E.; Fossard, C.; Okutman, O.; Ghédir, H.; Ibala-Romdhane, S.; Ray, P.F.; Saad, A.; Hennebicq, S.; Viville, S. A new mutation identified in SPATA16 in two globozoospermic patients. J. Assist. Reprod. Genet. 2016, 33, 815–820. [Google Scholar] [CrossRef] [Green Version]
- Ghédir, H.; Braham, A.; Viville, S.; Saad, A.; Ibala-Romdhane, S. Comparison of sperm morphology and nuclear sperm quality in SPATA16- and DPY19L2-mutated globozoospermic patients. Andrologia 2019, 51, e13277. [Google Scholar] [CrossRef] [PubMed]
- Modarres, P.; Tanhaei, S.; Tavalaee, M.; Ghaedi, K.; Deemeh, M.R.; Nasr-Esfahani, M.H. Assessment of DPY19L2 Deletion in Familial and Non-Familial Individuals with Globozoospermia and DPY19L2 Genotyping. Int. J. Fertil. Steril. 2016, 10, 196–207. [Google Scholar] [PubMed]
- Ellnati, E.; Kuentz, P.; Redin, C.; Jaber, S.; Vanden Meerschaut, F.; Makarian, J.; Koscinski, I.; Nasr-Esfahani, M.H.; Demirol, A.; Gurgan, T.; et al. Globozoospermia is mainly due to DPY19L2 deletion via non-allelic homologous recombination involving two recombination hotspots. Hum. Mol. Genet. 2012, 21, 3695–3702. [Google Scholar] [CrossRef] [PubMed]
- Noveski, P.; Madjunkova, S.; Maleva, I.; Sotiroska, V.; Petanovski, Z.; Plaseska-Karanfilska, D. A Homozygous Deletion of the DPY19l2 Gene is a Cause of Globozoospermia in Men from the Republic of Macedonia. Balkan J. Med. Genet. 2013, 16, 73–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Gong, F.; Lin, G.; Lu, G. DPY19L2 gene mutations are a major cause of globozoospermia: Identification of three novel point mutations. Mol. Hum. Reprod. 2013, 19, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Ounis, L.; Zoghmar, A.; Coutton, C.; Rouabah, L.; Hachemi, M.; Martinez, D.; Martinez, G.; Bellil, I.; Khelifi, D.; Arnoult, C.; et al. Mutations of the aurora kinase C gene causing macrozoospermia are the most frequent genetic cause of male infertility in Algerian men. Asian J. Androl. 2015, 17, 68–73. [Google Scholar] [PubMed]
- Ghédir, H.; Ibala-Romdhane, S.; Okutman, O.; Viot, G.; Saad, A.; Viville, S. Identification of a new DPY19L2 mutation and a better definition of DPY19L2 deletion breakpoints leading to globozoospermia. Mol. Hum. Reprod. 2016, 22, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.L.; Zhu, F.X.; Yan, J.; Chen, L.; Tang, W.H.; Xiao, S.; Mo, W.K.; Zhang, Z.G.; He, X.J.; Qiao, J.; et al. Novel DPY19L2 variants in globozoospermic patients and the overcoming this male infertility. Asian J. Androl. 2019, 21, 183–189. [Google Scholar]
- Baccetti, B.; Collodel, G.; Piomboni, P. Apoptosis in human ejaculated sperm cells. J. Submicrosc. Cytol. Pathol. 1996, 28, 587–596. [Google Scholar] [PubMed]
- Tejera, A.; Molla, M.; Muriel, L.; Remohi, J.; Pellicer, A.; De Pablo, J.L. Successful pregnancy and childbirth after intracytoplasmic sperm injection with calcium ionophore oocyte activation in a globozoospermic patient. Fertil. Steril. 2008, 90, 1202–1205. [Google Scholar] [CrossRef]
- Egashira, A.; Murakami, M.; Haigo, K.; Horiuchi, T.; Kuramoto, T. A successful pregnancy and live birth after intracytoplasmic sperm injection with globozoospermic sperm and electrical oocyte activation. Fertil. Steril. 2009, 92, e5–e9. [Google Scholar] [CrossRef]
- Taylor, S.L.; Yoon, S.Y.; Morshedi, M.S.; Lacey, D.R.; Jellerette, T.; Fissore, R.A.; Oehninger, S. Complete globozoospermia associated with PLCzeta deficiency treated with calcium ionophore and ICSI results in pregnancy. Reprod. Biomed. Online 2010, 20, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Brahem, S.; Mehdi, M.; Elghezal, H.; Saad, A. Analysis of sperm aneuploidies and DNA fragmentation in patients with globozoospermia or with abnormal acrosomes. Urology 2011, 77, 1343–1348. [Google Scholar] [CrossRef]
- Perrin, A.; Louanjli, N.; Ziane, Z.; Louanjli, T.; Le Roy, C.; Guéganic, N.; Amice, V.; De Braekeleer, M.; Morel, F. Study of aneuploidy and DNA fragmentation in gametes of patients with severe teratozoospermia. Reprod. Biomed. Online 2011, 22, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Zhioua, A.; Merdassi, G.; Bhouri, R.; Ferfouri, F.; Ben Ammar, A.; Amouri, A.; Vialard, F.; Zhioua, F. Apport de l’exploration cytogénétique et ultrastructurale dans le pronostic de fertilité des sujets globozoospermiques. Andrologie 2011, 21, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Haghighat, S.; Tavalaee, M.; Kouhkan, A.; Zakeri, Z.; Noureddini, M.; Shahverdi, A.H.; Nasr Esfahani, M.H. Reduction of truncated Kit Expression in men with abnormal semen parameters, globozoospermia and history of low or fertilization failure. Cell, J. 2019, 21, 314–321. [Google Scholar]
- Moretti, E.; Collodel, G.; Salvatici, M.C.; Belmonte, G.; Signorini, C. New insights into sperm with total globozoospermia: Increased fatty acid oxidation and centrin1 alteration. Syst. Biol. Reprod. Med. 2019, 65, 390–399. [Google Scholar] [CrossRef]
- Sermondade, N.; Hafhouf, E.; Dupont, C.; Bechoua, S.; Palacios, C.; Eustache, F.; Poncelet, C.; Benzacken, B.; Lévy, R.; Sifer, C. Successful childbirth after intracytoplasmic morphologically selected sperm injection without assisted oocyte activation in a patient with globozoospermia. Hum. Reprod. 2011, 26, 2944–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grassetti, D.; Paoli, D.; Gallo, M.; D’Ambrosio, A.; Lombardo, F.; Lenzi, A.; Gandini, L. Protamine-1 and -2 polymorphisms and gene expression in male infertility: An Italian study. J. Endocrinol. Investig. 2012, 35, 882–888. [Google Scholar]
- Lundin, K.; Sjögren, A.; Nilsson, L.; Hamberger, L. Fertilization and pregnancy after intracytoplasmic microinjection of acrosomeless spermatozoa. Fertil. Steril. 1994, 62, 1266–1267. [Google Scholar] [CrossRef]
- Stone, S.; O’Mahony, F.; Khalaf, Y.; Taylor, A.; Braude, P. A normal live birth after intracytoplasmic sperm injection for globozoospermia without assisted oocytes activation: Case report. Hum. Reprod. 2000, 15, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Zeyneloglu, H.B.; Baltaci, V.; Duran, H.E.; Erdemli, E.; Batioglu, S. Achievement of pregnancy in globozoospermia with Y chromosome microdeletion after ICSI. Hum. Reprod. 2002, 17, 1833–1836. [Google Scholar] [CrossRef] [Green Version]
- Battaglia, D.E.; Koehler, J.K.; Klein, N.A.; Tucker, M.J. Failure of oocyte activation after intracytoplasmic sperm injection using round-headed sperm. Fertil. Steril. 1997, 68, 118–122. [Google Scholar] [CrossRef]
- Sfontouris, I.A.; Nastri, C.O.; Lima, M.L.; Tahmasbpourmarzouni, E.; Raine-Fenning, N.; Martins, W.P. Artificial oocyte activation to improve reproductive outcomes in women with previous fertilization failure: A systematic review and meta-analysis of RCTs. Hum. Reprod. 2015, 30, 1831–1841. [Google Scholar] [CrossRef] [Green Version]
- Fawzy, M.; Emad, M.; Mahran, A.; Sabry, M.; Fetih, A.N.; Abdelghafar, H.; Rasheed, S. Artificial oocyte activation with SrCl2 or calcimycin after ICSI improves clinical and embryological outcomes compared with ICSI alone: Results of a randomized clinical trial. Hum. Reprod. 2018, 33, 1636–1644. [Google Scholar] [CrossRef]
- Murugesu, S.; Saso, S.; Jones, B.P.; Bracewell-Milnes, T.; Athanasiou, T.; Mania, A.; Serhal, P.; Ben-Nagi, J. Does the use of calcium ionophore during artificial oocyte activation demonstrate an effect on pregnancy rate? A meta-analysis. Fertil. Steril. 2017, 108, 468–482. [Google Scholar] [CrossRef] [Green Version]
- Dam, A.H.; Pijnenburg, A.J.; Hendriks, J.C.; Westphal, H.; Ramos, L.; Kremer, J.A. Intracytoplasmic sperm injection in partial globozoospermia. Fertil. Steril. 2012, 97, 60–66. [Google Scholar] [CrossRef]
- Kuentz, P.; Vanden Meerschaut, F.; Elinati, E.; Nasr-Esfahani, M.H.; Gurgan, T.; Iqbal, N.; Carré-Pigeon, F.; Brugnon, F.; Gitlin, S.A.; Velez de la Calle, J.; et al. Assisted oocyte activation overcomes fertilization failure in globozoospermic patients regardless of the DPY19L2 status. Hum. Reprod. 2013, 28, 1054–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaca, N.; Yilmaz, R.; Kanten, G.E.; Kervancioglu, E.; Solakoglu, S.; Kervancioglu, M.E. First successful pregnancy in a globozoospermic patient having homozygous mutation in SPATA16. Fertil. Steril. 2014, 102, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Canepa, P.; Casciano, I.; De Leo, C.; Massarotti, C.; Anserini, P.; Remorgida, V.; Scaruffi, P. A successful healthy childbirth and an ongoing evolutive pregnancy in a case of partial globozoospermia by hyaluronic acid sperm selection. Andrologia 2019, 51, e13178. [Google Scholar] [CrossRef] [PubMed]
- Kochhar, P.K.; Ghosh, P. Intracytoplasmic Sperm Injection with Assisted Oocyte Activation Resulting in Successful Pregnancies and Live Birth in Couples with Globozoospermia: A Report of Two Cases. J. Hum. Reprod. Sci. 2018, 11, 72–74. [Google Scholar] [CrossRef]
- Niu, X.; Ruan, Q.; Witz, C.A.; Wang, W. Comparison of Human Oocyte Activation Between Round-Headed Sperm Injection Followed by Calcium Ionophore Treatment and Normal Sperm Injection in a Patient With Globozoospermia. Front. Endocrinol. 2020, 11, 183. [Google Scholar] [CrossRef]
- Ghazavi, F.; Peymani, M.; Hashemi, M.S.; Ghaedi, K.; Nasr-Esfahani, M.H. Embryos derived from couples with consanguineous marriages with globozoospermia should be screened for gender or DPY19L2 deletion. Andrologia 2019, 51, e13221. [Google Scholar] [CrossRef]


| Semen Volume (ml) | Sperm Concentration (106/mL) | Total Sperm Number (106/ejaculate) | Progressive Motility (%) | Abnormal Forms (%) | |
|---|---|---|---|---|---|
| Group G | 2.5 ± 1.3 (2.5) | 75.1 ± 69.7 (59.0) | 162.6 ± 128.3 (124.5) | 36.4 ± 10.8 (40.0) | 94.6 ± 7.1 (100.0) |
| Group N | 3.3 ± 1.2 (3.0) | 84.6 ± 51.9 (75.0) | 273.1 ± 188.2 (244.0) | 51.5 ± 6.4 (55.0) | 88.0 ± 2.9 (87.0) |
| p value | 0.07 | 0.254 | 0.018 | <0.001 | <0.001 |
| Reference | N Patients (Case vs. Control) | Method to Evaluate SDF | DNA Fragmentation Index | |
|---|---|---|---|---|
| Case | Control | |||
| [55] | NA | Hoechst 33258 DNA staining TUNEL Electron Microscopy | 10% | 0.1% |
| [33] | 1 vs. 2 | TEM SCSA COMET | TEM: no elevated levels of apoptotic bodies SCSA and COMET: 13% | NA |
| [12] | 1 vs. 4 | TUNEL | 37.0 ± 1.7% | 22.5 ± 1.2% |
| [56] | 1 (case report) | SCD | 45.7% (threshold value: 30%) | / |
| [57] | 1 vs. 1 | AO | 97.1% | 41.3% |
| [58] | 1 vs. 1 | TUNEL | 80% | 27 ± 13% |
| [59] | 2 vs. 20 | TUNEL | Patient 1: 40% Patient 2: 80% | 12 ± 2.12% |
| [60] | 1 vs. unspecified | TUNEL | 9.6% | 1.20 ± 0.95% |
| [64] | 1 (case report) | TUNEL | 6% | / |
| [61] | 6 vs. unspecified | TUNEL | 15.5 ± 9,09% | <13% |
| [14] | 1 (case report) | SCSA TB CMA3 AB | SCSA: 35.3% (threshold value 30%) TB: 36.6% (threshold value 35%) CMA3: 47.7% AB: 56.3% | / |
| [24] | 8 vs. 90 | TUNEL | 46.92 ± 4.20% | 34.04 ± 1.53% |
| [34] | 15 vs. 21 | TUNEL CMA3 | TUNEL: 17.60 ± 10.72% CMA3: 65.93 ± 11.77% | TUNEL: 5.95 ± 3.02% CMA3: 21.24 ± 7.37% |
| [35] | 20 vs. 40 | SCSA CMA3 | SCSA: 33.83 ± 3.64% CMA3: 49.70 ± 4.88% | SCSA: 16.31 ± 1.58% CMA3: 30.97 ± 1.71% |
| [36] | 30 vs. 22 | TUNEL | 19.21 ± 3.75% | 8.58 ± 1.12% |
| [37] | 27 vs. 30 | SCD TUNEL AB TB CMA3 AO | SCD: partial globo 61.7 ± 13.2; total globo 57.8 ± 11.4 TUNEL: partial globo 12.3 ± 9.2; total globo 18.3 ± 10.1 AB: partial globo 81.3 ± 10.6; total globo 79.8 ± 12.9 TB: partial globo 73.1 ± 16; total globo 86.3 ± 9.1 CMA3: partial globo 60.1 ± 13.9; total globo 68.6 ± 11 AO: partial globo 33.2 ± 26.4; total globo 30.1 ± 18.4 | SCD: 11.5 ± 6.2 TUNEL: 5.9 ± 3 AB: 24.2 ± 16.2 TB: 32.8 ± 18.6 CMA3: 26.1 ± 11.6 AO: 11.5 ± 7.5 |
| [47] | 8 vs. 25 | TUNEL | 41.58 ± 10.99% | 10.25 ± 3.83% |
| [62] | 10 vs. 30 | TUNEL | 23.6 ± 5.67% | 5.24 ± 1.23% |
| [63] | 1 vs. 3 | AB AO TEM | AB: 78 ± 2.65% AO: 22.33 ± 2.52% TEM: chromatin compromised | NA |
| Present study | 18 vs. 31 | TUNEL | 29.7 ± 8.2% | 10.0 ± 2.8% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faja, F.; Pallotti, F.; Cargnelutti, F.; Senofonte, G.; Carlini, T.; Lenzi, A.; Lombardo, F.; Paoli, D. Molecular Analysis of DPY19L2, PICK1 and SPATA16 in Italian Unrelated Globozoospermic Men. Life 2021, 11, 641. https://doi.org/10.3390/life11070641
Faja F, Pallotti F, Cargnelutti F, Senofonte G, Carlini T, Lenzi A, Lombardo F, Paoli D. Molecular Analysis of DPY19L2, PICK1 and SPATA16 in Italian Unrelated Globozoospermic Men. Life. 2021; 11(7):641. https://doi.org/10.3390/life11070641
Chicago/Turabian StyleFaja, Fabiana, Francesco Pallotti, Francesco Cargnelutti, Giulia Senofonte, Tania Carlini, Andrea Lenzi, Francesco Lombardo, and Donatella Paoli. 2021. "Molecular Analysis of DPY19L2, PICK1 and SPATA16 in Italian Unrelated Globozoospermic Men" Life 11, no. 7: 641. https://doi.org/10.3390/life11070641