
Double Recombinant Vaccinia Virus: A Candidate Drug against Human Glioblastoma

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Virus
2.3. Animals
2.4. Cell Viability Assay
2.5. Flow Cytometry
2.6. VV-GMCSF-Lact Biodistribution in SCID Mice with Orthotopically Transplanted U87 MG Tumors and in Healthy SCID Mice
- Forward 5′-TACCATCGGCGGATCACATC-3′;
- Reversed 5′-AGCACAATACCGGGAGATGG-3′;
- Probe 5′-FAM-TCAGAAAACCCAAACACTACAACGTCA-BHQ-1-3′;
- Forward 5′-GATCAGACAAGGCAAAGCGAAG-3′;
- Reversed 5′-GACCCCAGTTTTAGCCAACATG-3′;
- Probe 5′-ROX-CTGTCCAGCTTTGAGGAAATC–BHQ2-3′.
2.7. VV-GMCSF-Lact Antitumor Efficacy against Subcutaneously Transplanted Human Glioblastoma Cells into SCID Mice
2.8. Ethical Statement
2.9. Statistics
3. Results
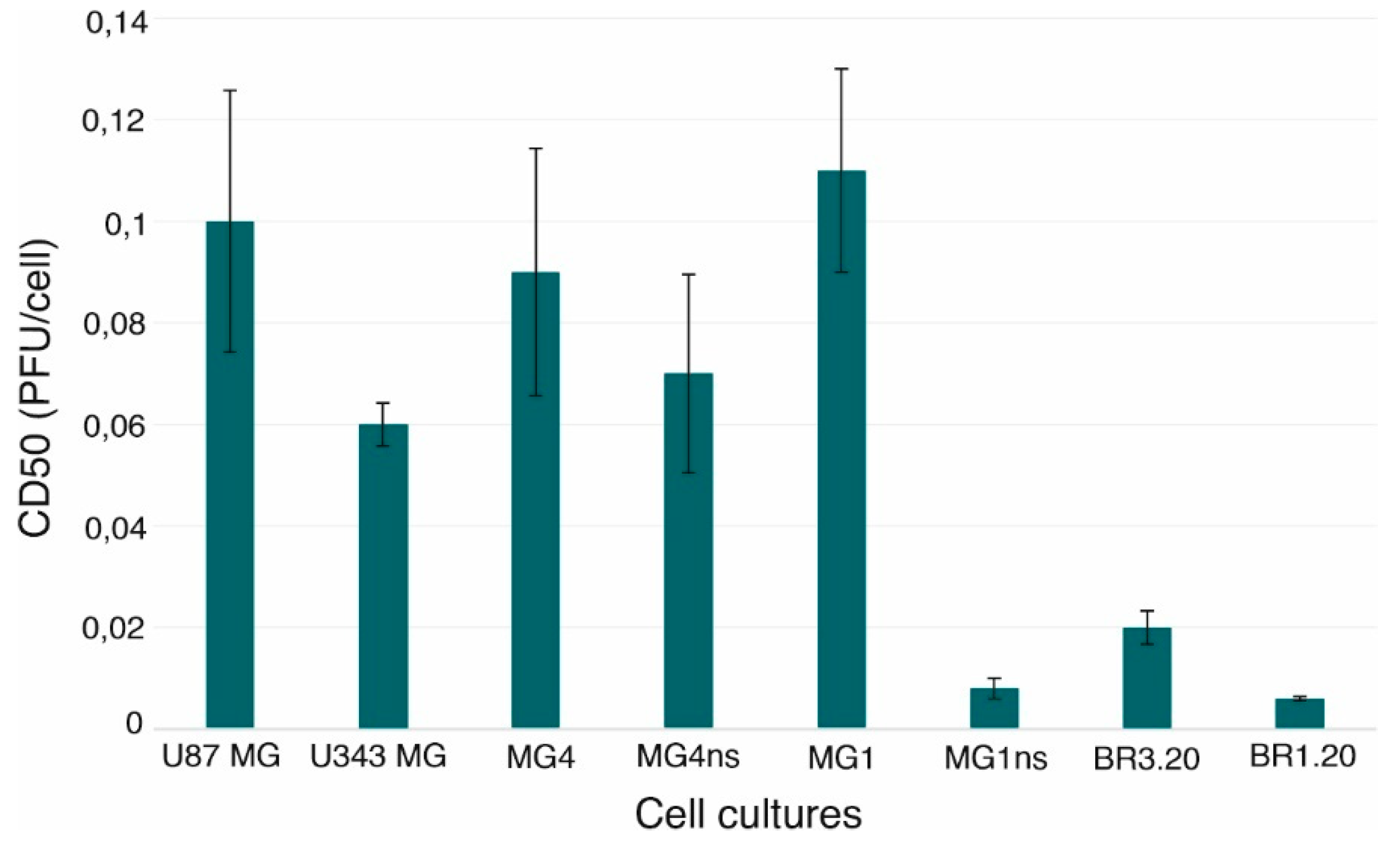
3.1. VV-GMCSF-Lact Cytotoxic Activity against Human Glioblastoma Cells
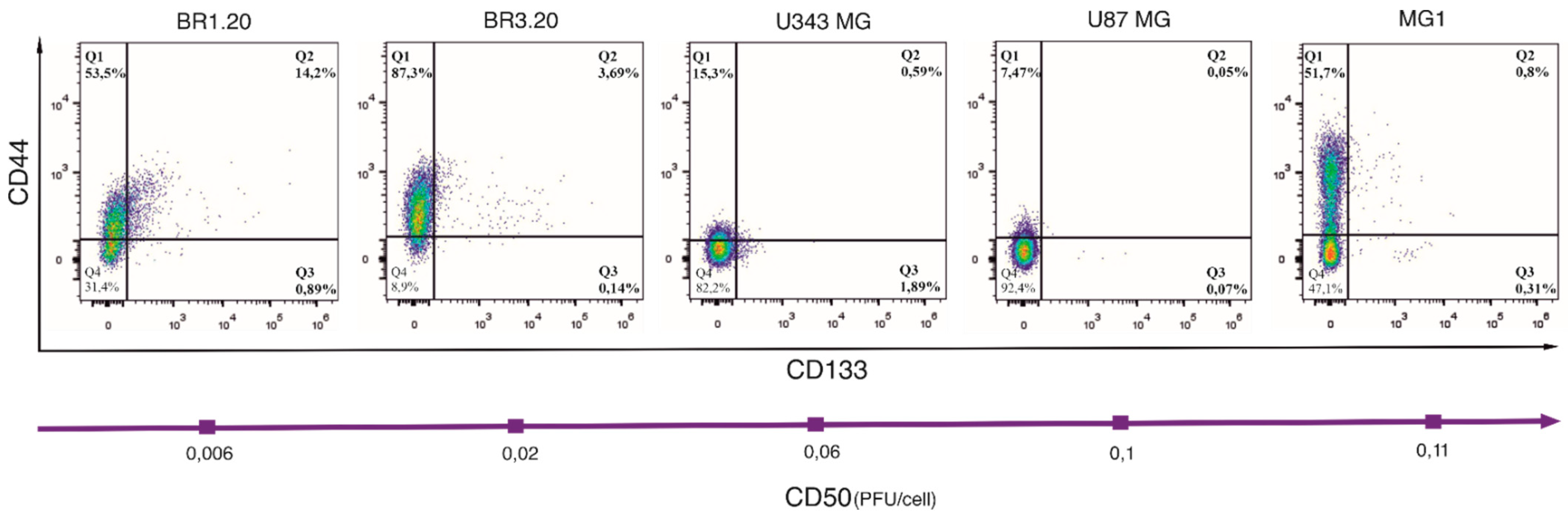
3.2. CD133 and CD44 Expression in Cells with Different VV-GMCSF-Lact Sensitivity
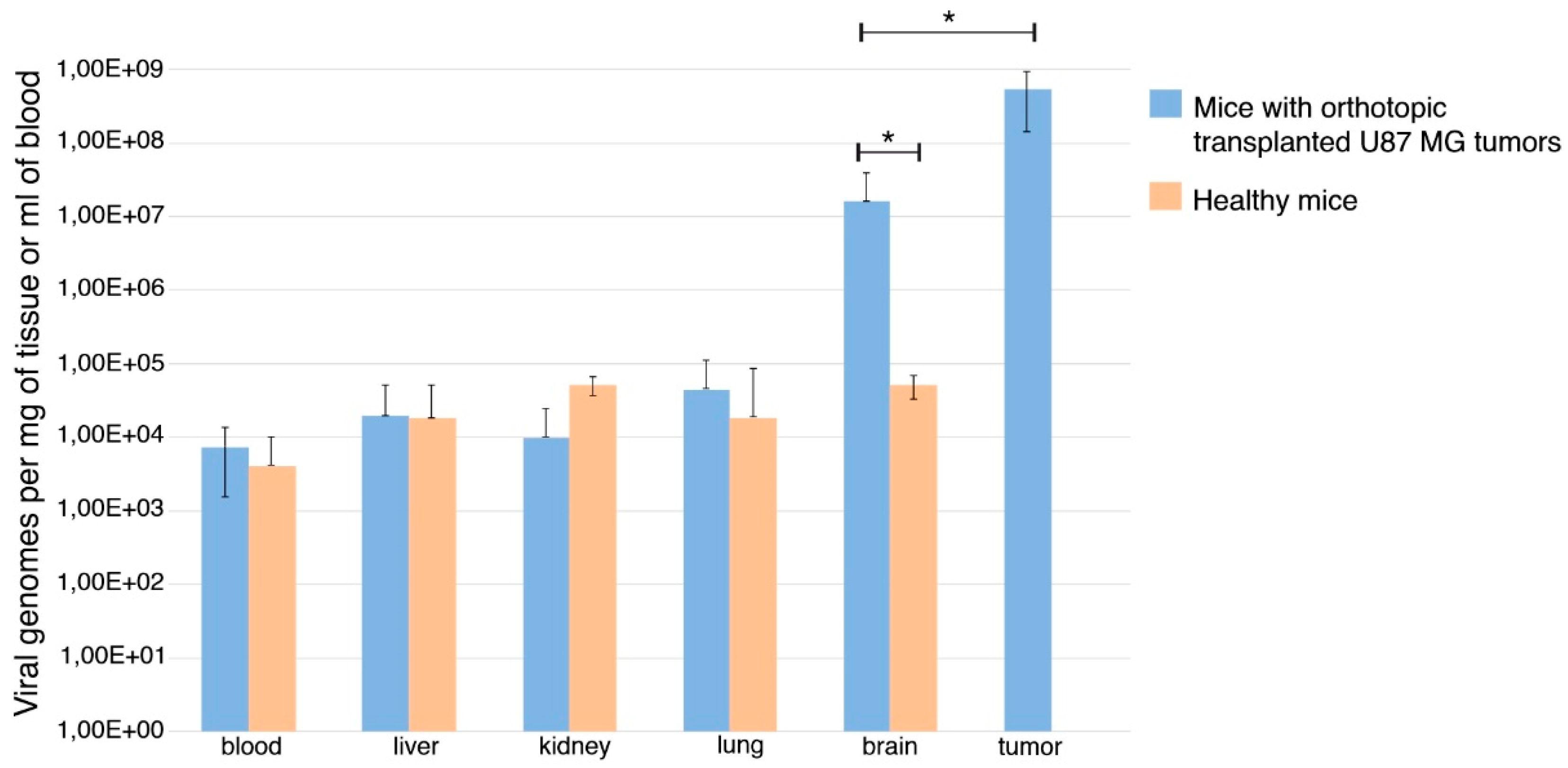
3.3. VV-GMCSF-Lact Ability to Cross the Blood–Brain Barrier
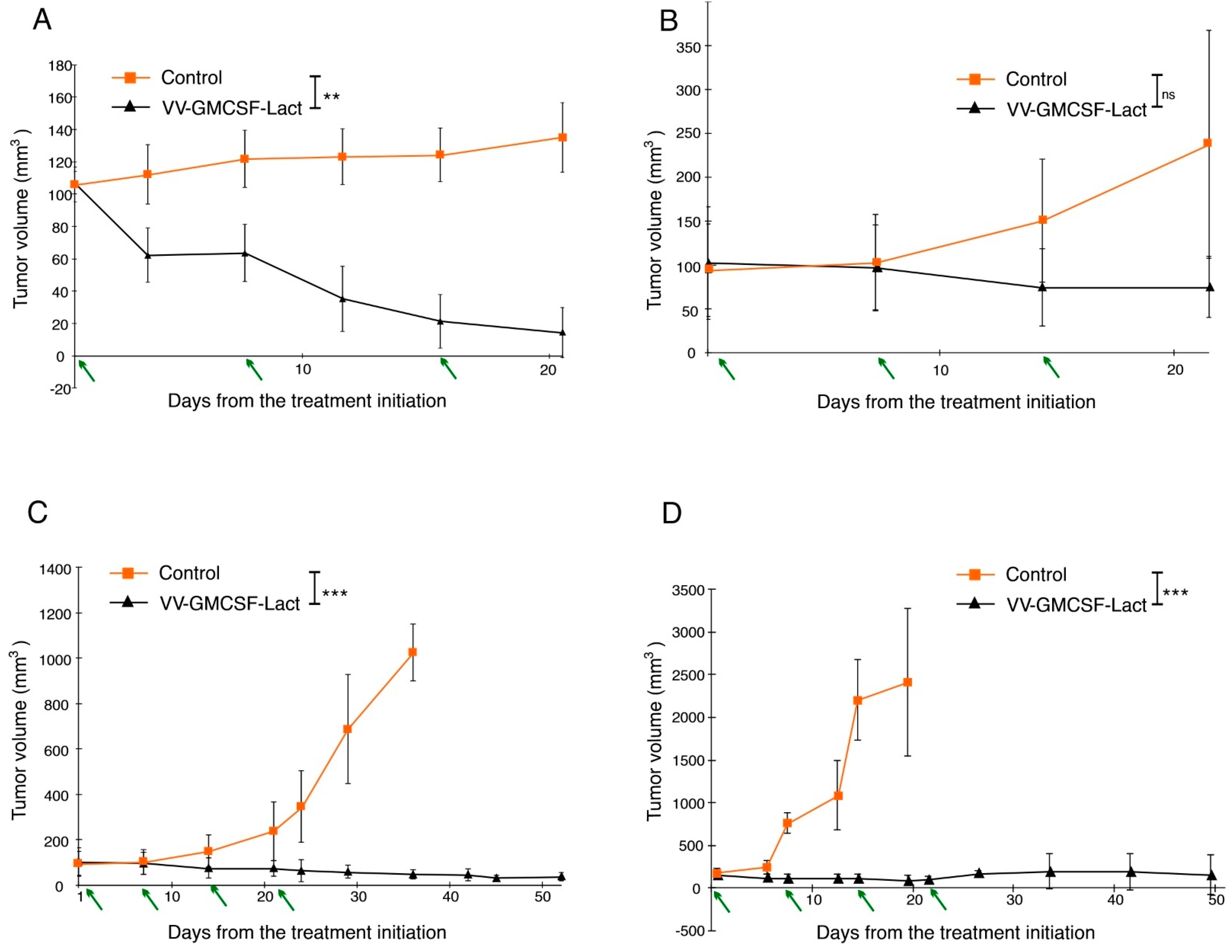
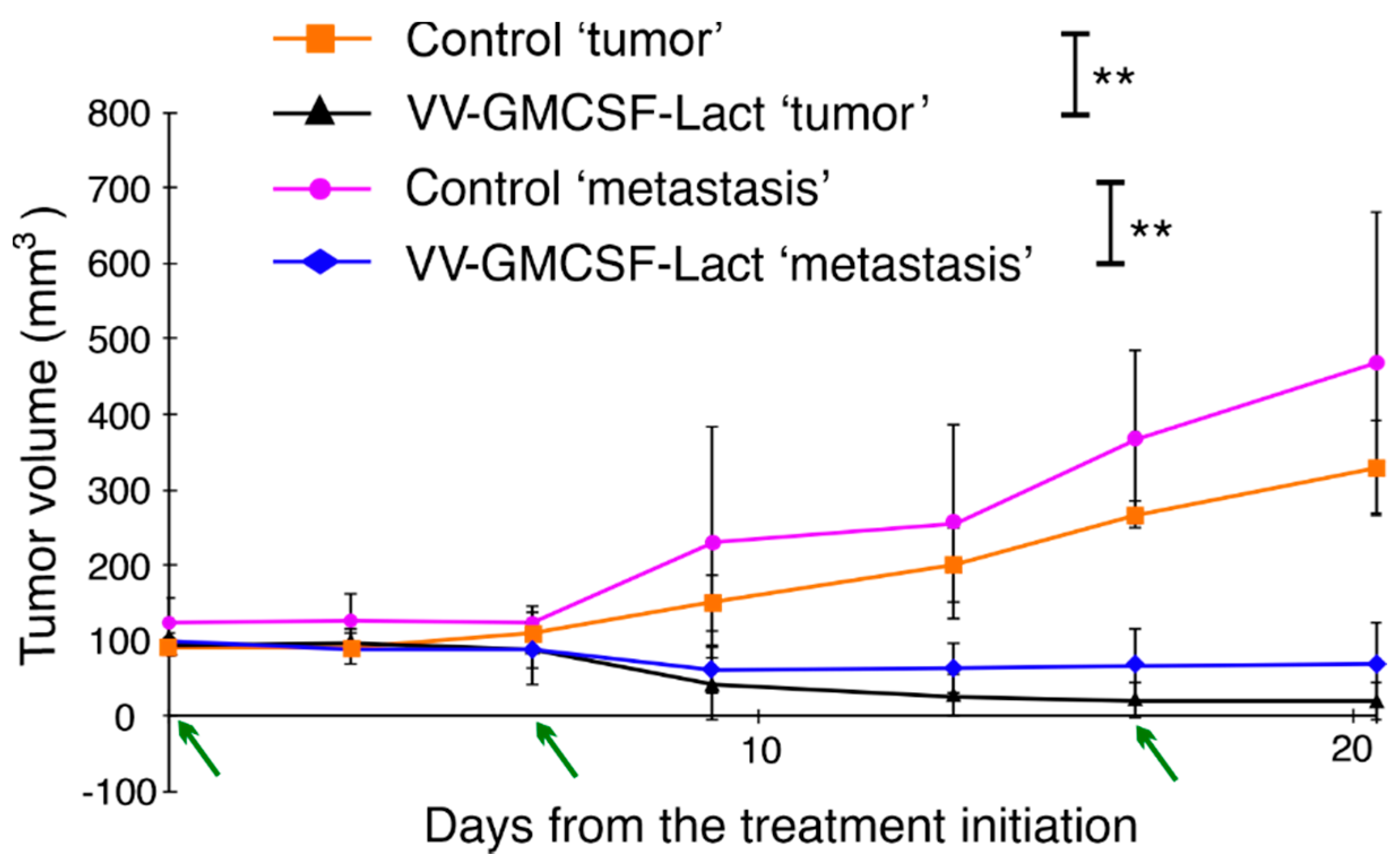
3.4. VV-GMCSF-Lact Antitumor and Antimetastatic Efficacy against Human Glioblastoma
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Culture | U87 MG | U343 MG | MG4 | MG4ns | MG1 | MG1ns | BR3.20 | BR1.20 |
|---|---|---|---|---|---|---|---|---|
| U87 MG | × | * | ns | ns | ns | ** | ** | ** |
| U343 MG | * | × | ns | ns | * | ** | ** | ** |
| MG4 | ns | ns | × | ns | ns | ** | ** | ** |
| MG4ns | ns | ns | ns | × | ns | ** | ** | ** |
| MG1 | ns | * | ns | ns | × | ** | ** | ** |
| MG1ns | ** | ** | ** | ** | ** | × | * | ns |
| BR3.20 | ** | ** | ** | ** | ** | * | × | * |
| BR1.20 | ** | ** | ** | ** | ** | ns | * | × |
References
- Yang, M.; Oh, I.Y.; Mahanty, A.; Jin, W.-L.; Yoo, J.S. Immunotherapy for Glioblastoma: Current State, Challenges, and Future Perspectives. Cancers 2020, 12, 2334. [Google Scholar] [CrossRef] [PubMed]
- Birzu, C.; French, P.; Caccese, M.; Cerretti, G.; Idbaih, A.; Zagonel, V.; Lombardi, G. Recurrent Glioblastoma: From Molecular Landscape to New Treatment Perspectives. Cancers 2020, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Lawler, S.E.; Speranza, M.-C.; Cho, C.-F.; Chiocca, E.A. Oncolytic Viruses in Cancer Treatment: A Review. JAMA Oncol. 2017, 3, 841–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, F.; Tran, N.; Puduvalli, V.; Elder, J.; Fink, K.; Conrad, C.; Yung, W.; Penas-Prado, M.; Gomez-Manzano, C.; Peterkin, J.; et al. Phase 1b Open-Label Randomized Study of the Oncolytic Adenovirus DNX-2401 Administered with or without Interferon Gamma for Recurrent Glioblastoma. J. Clin. Oncol. 2017, 35, 2002. [Google Scholar] [CrossRef]
- Todo, T. ATIM-14. Results of Phase II Clinical Trial of Oncolytic Herpes Virus g47δ in Patients with Glioblastoma. Neuro-Oncology 2019, 21, vi4. [Google Scholar] [CrossRef]
- Foreman, P.M.; Friedman, G.K.; Cassady, K.A.; Markert, J.M. Oncolytic Virotherapy for the Treatment of Malignant Glioma. Neurotherapeutics 2017, 14, 333–344. [Google Scholar] [CrossRef] [Green Version]
- Niemann, J.; Woller, N.; Brooks, J.; Fleischmann-Mundt, B.; Martin, N.T.; Kloos, A.; Knocke, S.; Ernst, A.M.; Manns, M.P.; Kubicka, S.; et al. Molecular Retargeting of Antibodies Converts Immune Defense against Oncolytic Viruses into Cancer Immunotherapy. Nat. Commun 2019, 10, 3236. [Google Scholar] [CrossRef] [Green Version]
- Koval, O.A.; Fomin, A.S.; Kaledin, V.I.; Semenov, D.V.; Potapenko, M.O.; Kuligina, E.V.; Nikolin, V.P.; Nikitenko, E.V.; Richter, V.A. A Novel Pro-Apoptotic Effector Lactaptin Inhibits Tumor Growth in Mice Models. Biochimie 2012, 94, 2467–2474. [Google Scholar] [CrossRef]
- Kochneva, G.; Sivolobova, G.; Tkacheva, A.; Grazhdantseva, A.; Troitskaya, O.; Nushtaeva, A.; Tkachenko, A.; Kuligina, E.; Richter, V.; Koval, O. Engineering of Double Recombinant Vaccinia Virus with Enhanced Oncolytic Potential for Solid Tumor Virotherapy. Oncotarget 2016, 7, 74171–74188. [Google Scholar] [CrossRef] [Green Version]
- Da Hora, C.C.; Schweiger, M.W.; Wurdinger, T.; Tannous, B.A. Patient-Derived Glioma Models: From Patients to Dish to Animals. Cells 2019, 8, 1177. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.-M.; Zhang, J.-G.; Zhang, X.; Li, Q. Targeting Cancer Stem Cells for Reversing Therapy Resistance: Mechanism, Signaling, and Prospective Agents. Sig Transduct Target. 2021, 6, 62. [Google Scholar] [CrossRef]
- Hassn Mesrati, M.; Behrooz, A.B.; Abuhamad, A.Y.; Syahir, A. Understanding Glioblastoma Biomarkers: Knocking a Mountain with a Hammer. Cells 2020, 9, 1236. [Google Scholar] [CrossRef]
- Kochneva, G.V.; Grazhdantseva, A.A.; Sivolobova, G.F.; Tkacheva, A.V.; Shvalov, A.N.; Unusova, A.U.; Ryabchikova, E.I.; Netesov, S.V. A model of the artificial metastasis of human epidermoid carcinoma A431 in nude mice for examination of the oncolytic activity of vaccinia virus. Vavilovskii Zhurnal Genet. Sel.—Vavilov J. Genet. Breed. 2015, 19, 480–486. [Google Scholar] [CrossRef] [Green Version]
- Lu, V.M.; Shah, A.H.; Vallejo, F.A.; Eichberg, D.G.; Luther, E.M.; Shah, S.S.; Komotar, R.J.; Ivan, M.E. Clinical Trials Using Oncolytic Viral Therapy to Treat Adult Glioblastoma: A Progress Report. Neurosurg. Focus 2021, 50, E3. [Google Scholar] [CrossRef]
- Zonov, E.; Kochneva, G.; Yunusova, A.; Grazhdantseva, A.; Richter, V.; Ryabchikova, E. Features of the Antitumor Effect of Vaccinia Virus Lister Strain. Viruses 2016, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.; Wang, P.; Alusi, G.; Shi, H.; Chu, Y.; Wang, J.; Bhakta, V.; McNeish, I.; McCart, A.; Lemoine, N.R.; et al. Lister Strain Vaccinia Virus with Thymidine Kinase Gene Deletion Is a Tractable Platform for Development of a New Generation of Oncolytic Virus. Gene Ther. 2015, 22, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Semenov, D.V.; Fomin, A.S.; Kuligina, E.V.; Koval, O.A.; Matveeva, V.A.; Babkina, I.N.; Tikunova, N.V.; Richter, V.A. Recombinant Analogs of a Novel Milk Pro-Apoptotic Peptide, Lactaptin, and Their Effect on Cultured Human Cells. Protein J. 2010, 29, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Rius-Rocabert, S.; García-Romero, N.; García, A.; Ayuso-Sacido, A.; Nistal-Villan, E. Oncolytic Virotherapy in Glioma Tumors. Int. J. Mol. Sci. 2020, 21, 7604. [Google Scholar] [CrossRef] [PubMed]
- de Sostoa, J.; Dutoit, V.; Migliorini, D. Oncolytic Viruses as a Platform for the Treatment of Malignant Brain Tumors. Int. J. Mol. Sci. 2020, 21, 7449. [Google Scholar] [CrossRef]
- Skaga, E.; Kulesskiy, E.; Brynjulvsen, M.; Sandberg, C.J.; Potdar, S.; Langmoen, I.A.; Laakso, A.; Gaál-Paavola, E.; Perola, M.; Wennerberg, K.; et al. Feasibility Study of Using High-throughput Drug Sensitivity Testing to Target Recurrent Glioblastoma Stem Cells for Individualized Treatment. Clin. Transl. Med. 2019, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Soubéran, A.; Tchoghandjian, A. Practical Review on Preclinical Human 3D Glioblastoma Models: Advances and Challenges for Clinical Translation. Cancers 2020, 12, 2347. [Google Scholar] [CrossRef]
- Luger, A.-L.; Lorenz, N.I.; Urban, H.; Divé, I.; Engel, A.L.; Strassheimer, F.; Dettmer, K.; Zeiner, P.S.; Shaid, S.; Struve, N.; et al. Activation of Epidermal Growth Factor Receptor Sensitizes Glioblastoma Cells to Hypoxia-Induced Cell Death. Cancers 2020, 12, 2144. [Google Scholar] [CrossRef] [PubMed]
- William, D.; Mokri, P.; Lamp, N.; Linnebacher, M.; Classen, C.F.; Erbersdobler, A.; Schneider, B. Amplification of the EGFR Gene Can Be Maintained and Modulated by Variation of EGF Concentrations in in Vitro Models of Glioblastoma Multiforme. PLoS ONE 2017, 12, e0185208. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-C.; Wei, X.-T.; Guan, J.-H.; Shu, H.; Chen, D. EGF Stimulates Glioblastoma Metastasis by Induction of Matrix Metalloproteinase-9 in an EGFR-Dependent Mechanism. Oncotarget 2017, 8, 65969–65982. [Google Scholar] [CrossRef] [Green Version]
- Meshki, J.; Douglas, S.D.; Hu, M.; Leeman, S.E.; Tuluc, F. Substance P Induces Rapid and Transient Membrane Blebbing in U373MG Cells in a P21-Activated Kinase-Dependent Manner. PLoS ONE 2011, 6, e25332. [Google Scholar] [CrossRef] [Green Version]
- Mercer, J.; Helenius, A. Vaccinia Virus Uses Macropinocytosis and Apoptotic Mimicry to Enter Host Cells. Science 2008, 320, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Won, C.; Kim, B.; Yi, E.H.; Choi, K.; Kim, E.; Jeong, J.; Lee, J.; Jang, J.; Yoon, J.; Jeong, W.; et al. Signal Transducer and Activator of Transcription 3-mediated CD133 Up-regulation Contributes to Promotion of Hepatocellular Carcinoma. Hepatology 2015, 62, 1160–1173. [Google Scholar] [CrossRef] [Green Version]
- Tang, K.H.; Ma, S.; Lee, T.K.; Chan, Y.P.; Kwan, P.S.; Tong, C.M.; Ng, I.O.; Man, K.; To, K.-F.; Lai, P.B.; et al. CD133+ Liver Tumor-Initiating Cells Promote Tumor Angiogenesis, Growth, and Self-Renewal through Neurotensin/Interleukin-8/CXCL1 Signaling. Hepatology 2012, 55, 807–820. [Google Scholar] [CrossRef]
- Wei, Y.; Jiang, Y.; Zou, F.; Liu, Y.; Wang, S.; Xu, N.; Xu, W.; Cui, C.; Xing, Y.; Liu, Y.; et al. Activation of PI3K/Akt Pathway by CD133-P85 Interaction Promotes Tumorigenic Capacity of Glioma Stem Cells. Proc. Natl. Acad. Sci. USA 2013, 110, 6829–6834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoranjan, B.; Chokshi, C.; Venugopal, C.; Subapanditha, M.; Savage, N.; Tatari, N.; Provias, J.P.; Murty, N.K.; Moffat, J.; Doble, B.W.; et al. A CD133-AKT-Wnt Signaling Axis Drives Glioblastoma Brain Tumor-Initiating Cells. Oncogene 2020, 39, 1590–1599. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Garcia, H.; Alvarado-Estrada, K.; Krishnan, S.; Quinones-Hinojosa, A.; Trifiletti, D.M. Nanoparticles for Stem Cell Therapy Bioengineering in Glioma. Front. Bioeng. Biotechnol. 2020, 8, 558375. [Google Scholar] [CrossRef]
- Drumm, M.R.; Dixit, K.S.; Grimm, S.; Kumthekar, P.; Lukas, R.V.; Raizer, J.J.; Stupp, R.; Chheda, M.G.; Kam, K.-L.; McCord, M.; et al. Extensive Brainstem Infiltration, Not Mass Effect, Is a Common Feature of End-Stage Cerebral Glioblastomas. Neuro-Oncology 2020, 22, 470–479. [Google Scholar] [CrossRef]
- Chiocca, E.A.; Nakashima, H.; Kasai, K.; Fernandez, S.A.; Oglesbee, M. Preclinical Toxicology of RQNestin34.5v.2: An Oncolytic Herpes Virus with Transcriptional Regulation of the ICP34.5 Neurovirulence Gene. Mol. Ther.—Methods Clin. Dev. 2020, 17, 871–893. [Google Scholar] [CrossRef] [PubMed]
- Zadeh, G.; Lang, F.; Daras, M.; Cloughesy, T.; Colman, H.; Ong, S.; Ramakrishna, R.; Vogelbaum, M.; Groves, M.; Nassiri, F.; et al. ATIM-24. Interim Results of a Phase II Multicenter Study of the Conditionally Replicative Oncolytic Adenovirus DNX-2401 with Pembrolizumab (Keytruda) for Recurrent Glioblastoma; Captive Study (Keynote-192). Neuro-Oncology 2018, 20, vi6. [Google Scholar] [CrossRef] [Green Version]
- Oberoi, R.K.; Parrish, K.E.; Sio, T.T.; Mittapalli, R.K.; Elmquist, W.F.; Sarkaria, J.N. Strategies to Improve Delivery of Anticancer Drugs across the Blood–Brain Barrier to Treat Glioblastoma. Neuro-Oncology 2016, 18, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganipineni, L.P.; Danhier, F.; Préat, V. Drug Delivery Challenges and Future of Chemotherapeutic Nanomedicine for Glioblastoma Treatment. J. Control. Release 2018, 281, 42–57. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C.; Lu, W. The Blood-Brain/Tumor Barriers: Challenges and Chances for Malignant Gliomas Targeted Drug Delivery. Curr. Pharm. Biotechnol. 2012, 13, 2380–2387. [Google Scholar] [CrossRef]
- Cha, G.D.; Kang, T.; Baik, S.; Kim, D.; Choi, S.H.; Hyeon, T.; Kim, D.-H. Advances in Drug Delivery Technology for the Treatment of Glioblastoma Multiforme. J. Control. Release 2020, 328, 350–367. [Google Scholar] [CrossRef]
- Bahar, M.W.; Graham, S.C.; Chen, R.A.-J.; Cooray, S.; Smith, G.L.; Stuart, D.I.; Grimes, J.M. How Vaccinia Virus Has Evolved to Subvert the Host Immune Response. J. Struct. Biol. 2011, 175, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Lauer, U.M.; Schell, M.; Beil, J.; Berchtold, S.; Koppenhöfer, U.; Glatzle, J.; Königsrainer, A.; Möhle, R.; Nann, D.; Fend, F.; et al. Phase I Study of Oncolytic Vaccinia Virus GL-ONC1 in Patients with Peritoneal Carcinomatosis. Clin. Cancer Res. 2018, 24, 4388–4398. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Breitbach, C.J.; Lee, J.; Park, J.O.; Lim, H.Y.; Kang, W.K.; Moon, A.; Mun, J.-H.; Sommermann, E.M.; Maruri Avidal, L.; et al. Phase 1b Trial of Biweekly Intravenous Pexa-Vec (JX-594), an Oncolytic and Immunotherapeutic Vaccinia Virus in Colorectal Cancer. Mol. Ther. 2015, 23, 1532–1540. [Google Scholar] [CrossRef]
- Downs-Canner, S.; Guo, Z.S.; Ravindranathan, R.; Breitbach, C.J.; O’Malley, M.E.; Jones, H.L.; Moon, A.; McCart, J.A.; Shuai, Y.; Zeh, H.J.; et al. Phase 1 Study of Intravenous Oncolytic Poxvirus (VvDD) in Patients With Advanced Solid Cancers. Mol. Ther. 2016, 24, 1492–1501. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.; Reid, T.; Ruo, L.; Breitbach, C.J.; Rose, S.; Bloomston, M.; Cho, M.; Lim, H.Y.; Chung, H.C.; Kim, C.W.; et al. Randomized Dose-Finding Clinical Trial of Oncolytic Immunotherapeutic Vaccinia JX-594 in Liver Cancer. Nat. Med. 2013, 19, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, N.; Poirier, J.T.; Burga, L.N.; Bostina, M. Virus–Receptor Interactions and Virus Neutralization: Insights for Oncolytic Virus Development. Oncolytic Virotherapy 2020, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco-Luzón, L.; González-Murillo, Á.; Alcántara-Sánchez, C.; García-García, L.; Tabasi, M.; Huertas, A.L.; Chesler, L.; Ramírez, M. Systemic Oncolytic Adenovirus Delivered in Mesenchymal Carrier Cells Modulate Tumor Infiltrating Immune Cells and Tumor Microenvironment in Mice with Neuroblastoma. Oncotarget 2020, 11, 347–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kijima, N.; Kanemura, Y. Mouse Models of Glioblastoma. In Glioblastoma; De Vleeschouwer, S., Ed.; Codon Publications: Brisbane, Australia, 2017; ISBN 978-0-9944381-2-6. [Google Scholar]
- Hicks, W.H.; Bird, C.E.; Traylor, J.I.; Shi, D.D.; El Ahmadieh, T.Y.; Richardson, T.E.; McBrayer, S.K.; Abdullah, K.G. Contemporary Mouse Models in Glioma Research. Cells 2021, 10, 712. [Google Scholar] [CrossRef]
- Huszthy, P.C.; Daphu, I.; Niclou, S.P.; Stieber, D.; Nigro, J.M.; Sakariassen, P.Ø.; Miletic, H.; Thorsen, F.; Bjerkvig, R. In Vivo Models of Primary Brain Tumors: Pitfalls and Perspectives. Neuro-Oncology 2012, 14, 979–993. [Google Scholar] [CrossRef] [Green Version]
- Almendro, V.; Marusyk, A.; Polyak, K. Cellular Heterogeneity and Molecular Evolution in Cancer. Annu. Rev. Pathol. 2013, 8, 277–302. [Google Scholar] [CrossRef] [Green Version]
- Ernst, A.; Hofmann, S.; Ahmadi, R.; Becker, N.; Korshunov, A.; Engel, F.; Hartmann, C.; Felsberg, J.; Sabel, M.; Peterziel, H.; et al. Genomic and Expression Profiling of Glioblastoma Stem Cell-like Spheroid Cultures Identifies Novel Tumor-Relevant Genes Associated with Survival. Clin. Cancer Res. 2009, 15, 6541–6550. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.J.; Homer, N.; O’Connor, B.D.; Chen, Z.; Eskin, A.; Lee, H.; Merriman, B.; Nelson, S.F. U87MG Decoded: The Genomic Sequence of a Cytogenetically Aberrant Human Cancer Cell Line. PLoS Genet. 2010, 6, e1000832. [Google Scholar] [CrossRef]
- Kerstetter-Fogle, A.E.; Harris, P.L.R.; Brady-Kalnay, S.M.; Sloan, A.E. Generation of Glioblastoma Patient-Derived Intracranial Xenografts for Preclinical Studies. Int. J. Mol. Sci. 2020, 21, 5113. [Google Scholar] [CrossRef] [PubMed]
- Anghileri, E.; Elena, A.; Castiglione, M.; Melina, C.; Nunziata, R.; Raffaele, N.; Boffano, C.; Carlo, B.; Nazzi, V.; Vittoria, N.; et al. Extraneural Metastases in Glioblastoma Patients: Two Cases with YKL-40-Positive Glioblastomas and a Meta-Analysis of the Literature. Neurosurg. Rev. 2016, 39, 37–45. [Google Scholar] [CrossRef]
- Sun, Q.; Xu, R.; Xu, H.; Wang, G.; Shen, X.; Jiang, H. Extracranial Metastases of High-Grade Glioma: The Clinical Characteristics and Mechanism. World, J. Surg. Oncol. 2017, 15, 181. [Google Scholar] [CrossRef] [Green Version]
- Rosen, J.; Blau, T.; Grau, S.J.; Barbe, M.T.; Fink, G.R.; Galldiks, N. Extracranial Metastases of a Cerebral Glioblastoma: A Case Report and Review of the Literature. Case Rep. Oncol. 2018, 11, 591–600. [Google Scholar] [CrossRef] [Green Version]
- Vollmann-Zwerenz, A.; Leidgens, V.; Feliciello, G.; Klein, C.A.; Hau, P. Tumor Cell Invasion in Glioblastoma. Int. J. Mol. Sci. 2020, 21, 1932. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.J.; Cho, W.H.; Kang, D.W.; Cha, S.H. Extraneural Metastasis of Glioblastoma Multiforme Presenting as an Unusual Neck Mass. J. Korean Neurosurg Soc. 2012, 51, 147. [Google Scholar] [CrossRef]
- Lah, T.T.; Novak, M.; Breznik, B. Brain Malignancies: Glioblastoma and Brain Metastases. Semin. Cancer Biol. 2020, 60, 262–273. [Google Scholar] [CrossRef]
- Piccirilli, M.; Brunetto, G.M.F.; Rocchi, G.; Giangaspero, F.; Salvati, M. Extra Central Nervous System Metastases from Cerebral Glioblastoma Multiforme in Elderly Patients. Clinico-Pathological Remarks on Our Series of Seven Cases and Critical Review of the Literature. Tumori J. 2008, 94, 40–51. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasileva, N.; Ageenko, A.; Dmitrieva, M.; Nushtaeva, A.; Mishinov, S.; Kochneva, G.; Richter, V.; Kuligina, E. Double Recombinant Vaccinia Virus: A Candidate Drug against Human Glioblastoma. Life 2021, 11, 1084. https://doi.org/10.3390/life11101084
Vasileva N, Ageenko A, Dmitrieva M, Nushtaeva A, Mishinov S, Kochneva G, Richter V, Kuligina E. Double Recombinant Vaccinia Virus: A Candidate Drug against Human Glioblastoma. Life. 2021; 11(10):1084. https://doi.org/10.3390/life11101084
Chicago/Turabian StyleVasileva, Natalia, Alisa Ageenko, Maria Dmitrieva, Anna Nushtaeva, Sergey Mishinov, Galina Kochneva, Vladimir Richter, and Elena Kuligina. 2021. "Double Recombinant Vaccinia Virus: A Candidate Drug against Human Glioblastoma" Life 11, no. 10: 1084. https://doi.org/10.3390/life11101084