Loganic Acid, an Iridoid Glycoside Extracted from Cornus mas L. Fruits, Reduces of Carbonyl/Oxidative Stress Biomarkers in Plasma and Restores Antioxidant Balance in Leukocytes of Rats with Streptozotocin-Induced Diabetes Mellitus

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Materials and Sample Preparation of Loganic Acid
2.3. Identification of Compounds by Liquid Chromatography-Mass Spectrometry (LC-MS)
2.4. Determination of Compounds by HPLC
2.5. Animal Experiments
2.5.1. Animal Study
2.5.2. Induction of DM in Rats
2.5.3. Experimental Design
2.6. Measurements of Body Weight and Weight Gain
2.7. Fasting Blood Glucose and Oral Glucose Tolerance Test (OGTT)
2.8. Blood Collection
2.9. Isolation of Blood Leukocytes
2.10. Blood Parameters
2.10.1. Parameters Related to Erythrocytes
2.10.2. Leukocyte Related Parameters
2.11. Carbonyl/Oxidative Stress Markers in Plasma and Leukocytes
2.11.1. Quantification of Reactive Oxygen Species (ROS) in Leukocytes
2.11.2. Reduced Glutathione (GSH)
2.11.3. Lipid Peroxidation (LPO)
2.11.4. Assay of Oxidatively Modified Proteins (OMPs)
2.11.5. Assay of Advanced Oxidation Protein Products (AOPPs)
2.11.6. Assay of Advanced Glycation End-Products (AGEs)
2.12. Antioxidant Enzymes Assays
2.13. Statistical Analysis
3. Results
3.1. Qualitative and Quantitative Analysis of Compounds of the Purified Loganic Acid Extract
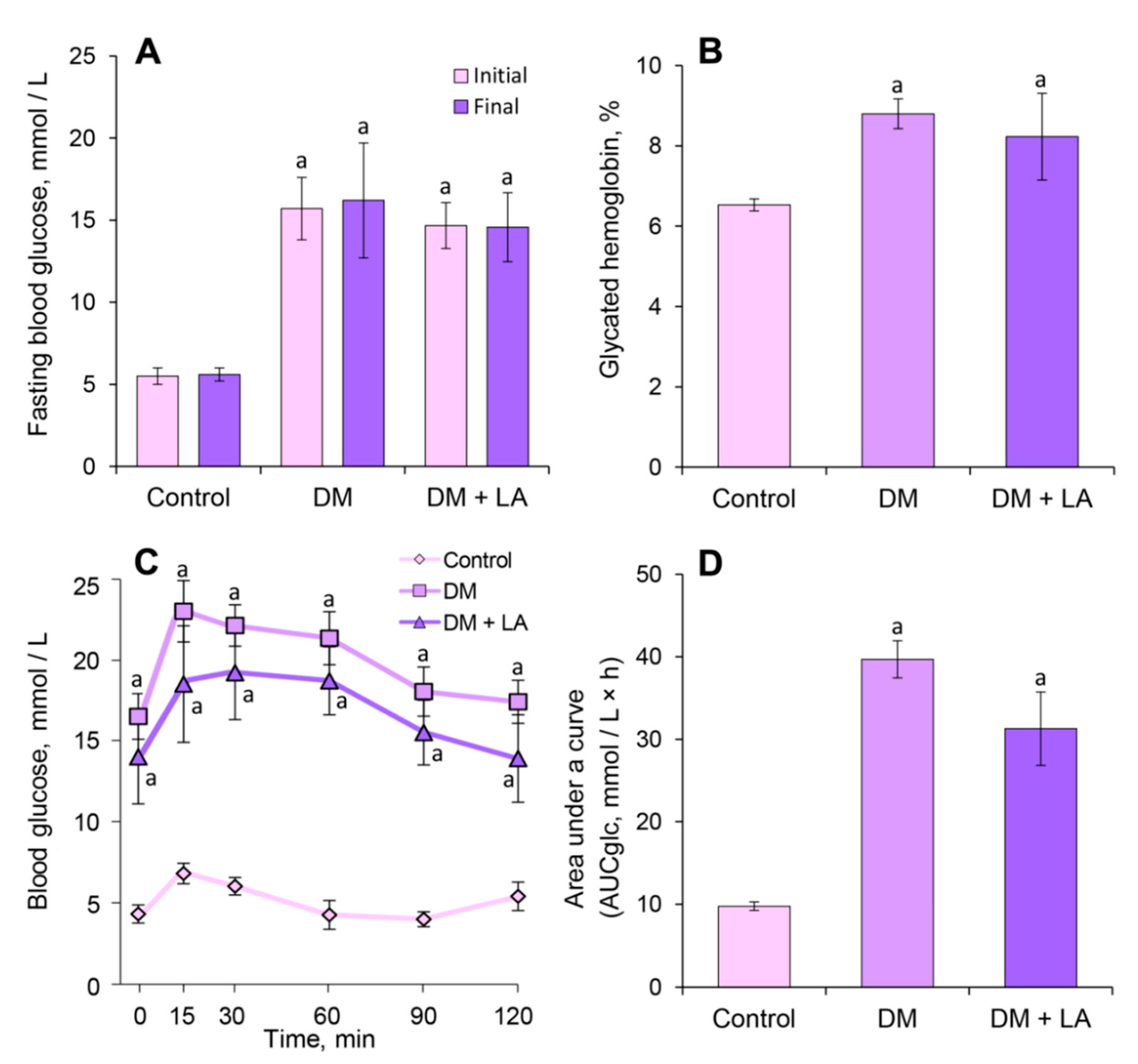
3.2. Loganic Acid and Antidiabetic Effects
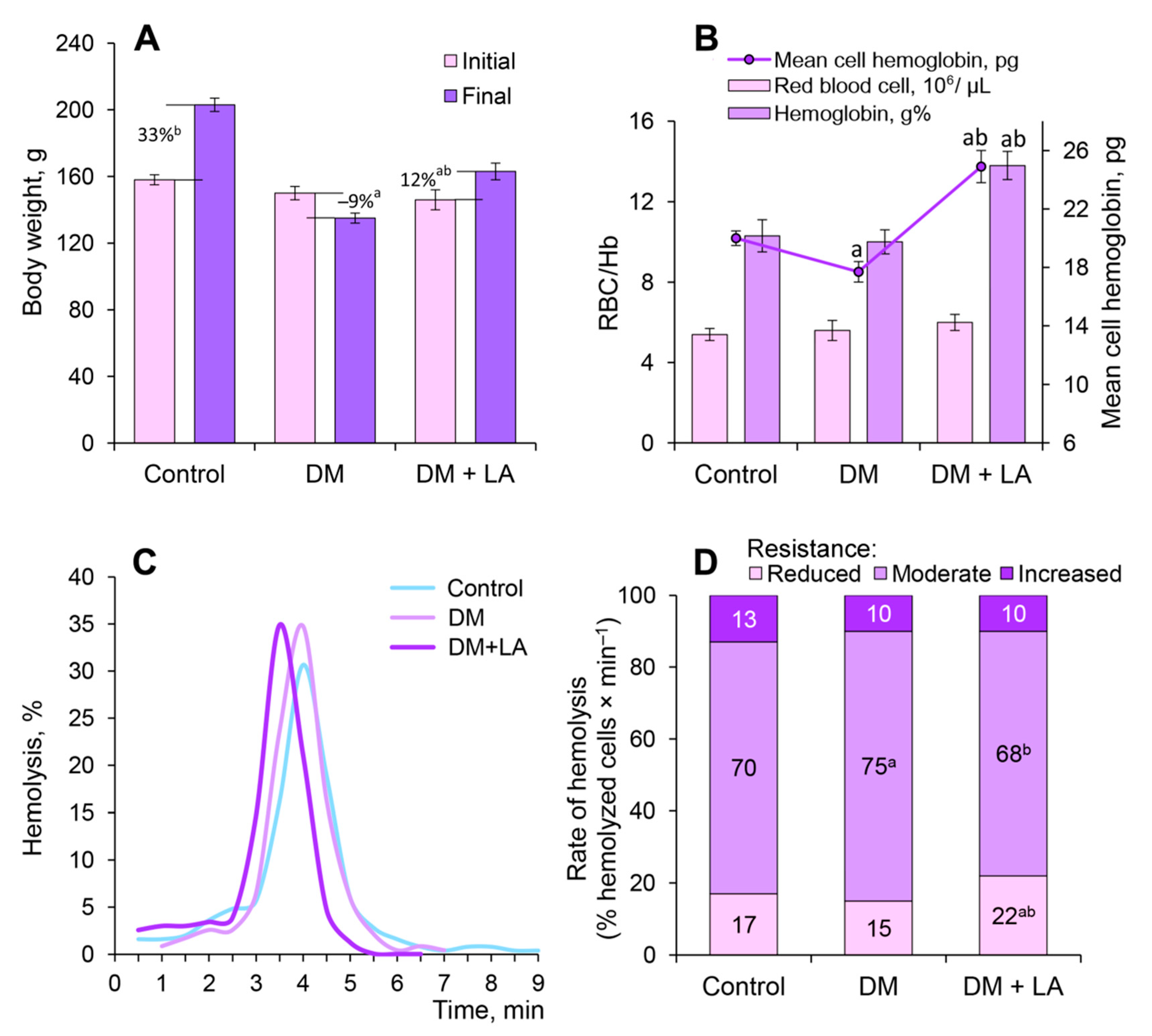
3.3. Loganic Acid Alter Body Weight and Erythrocyte-Related Parameters
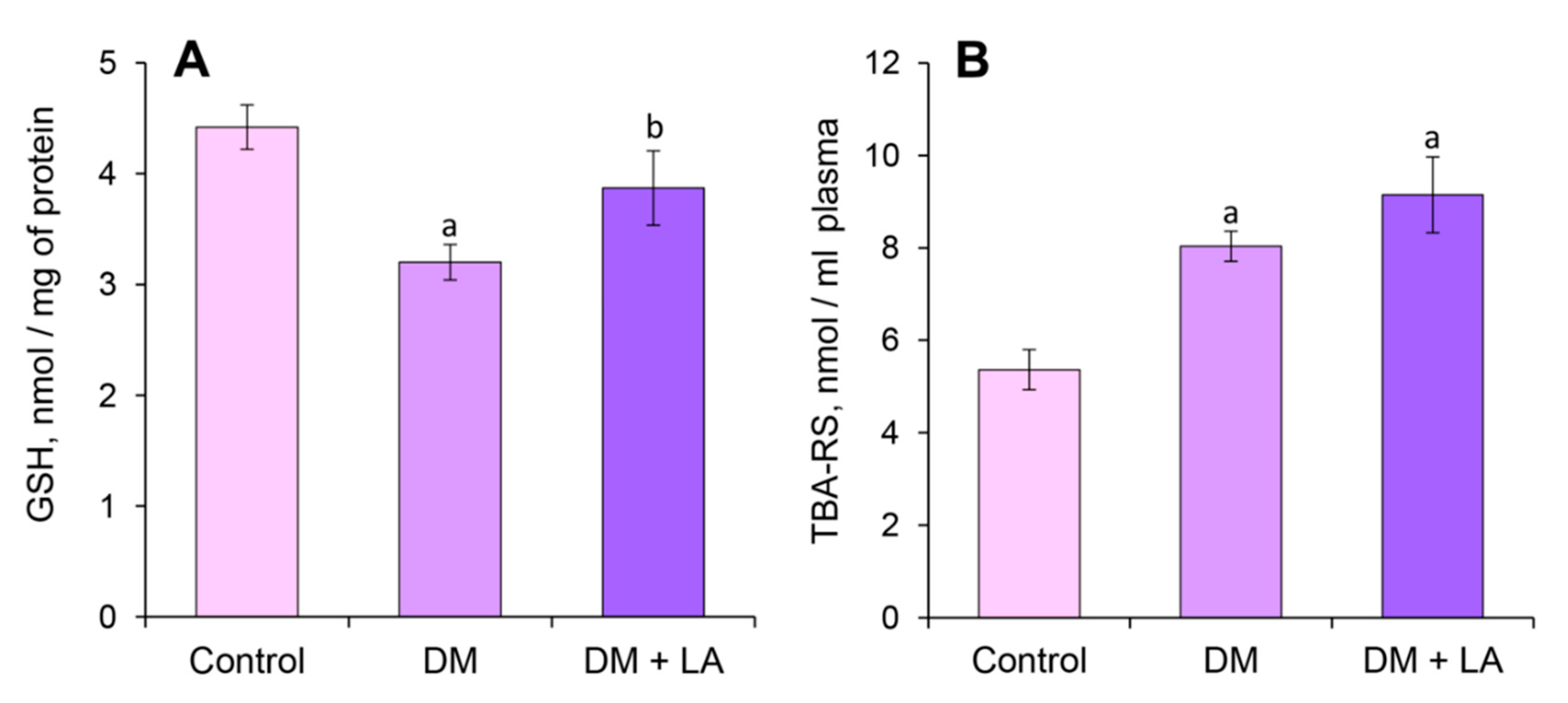
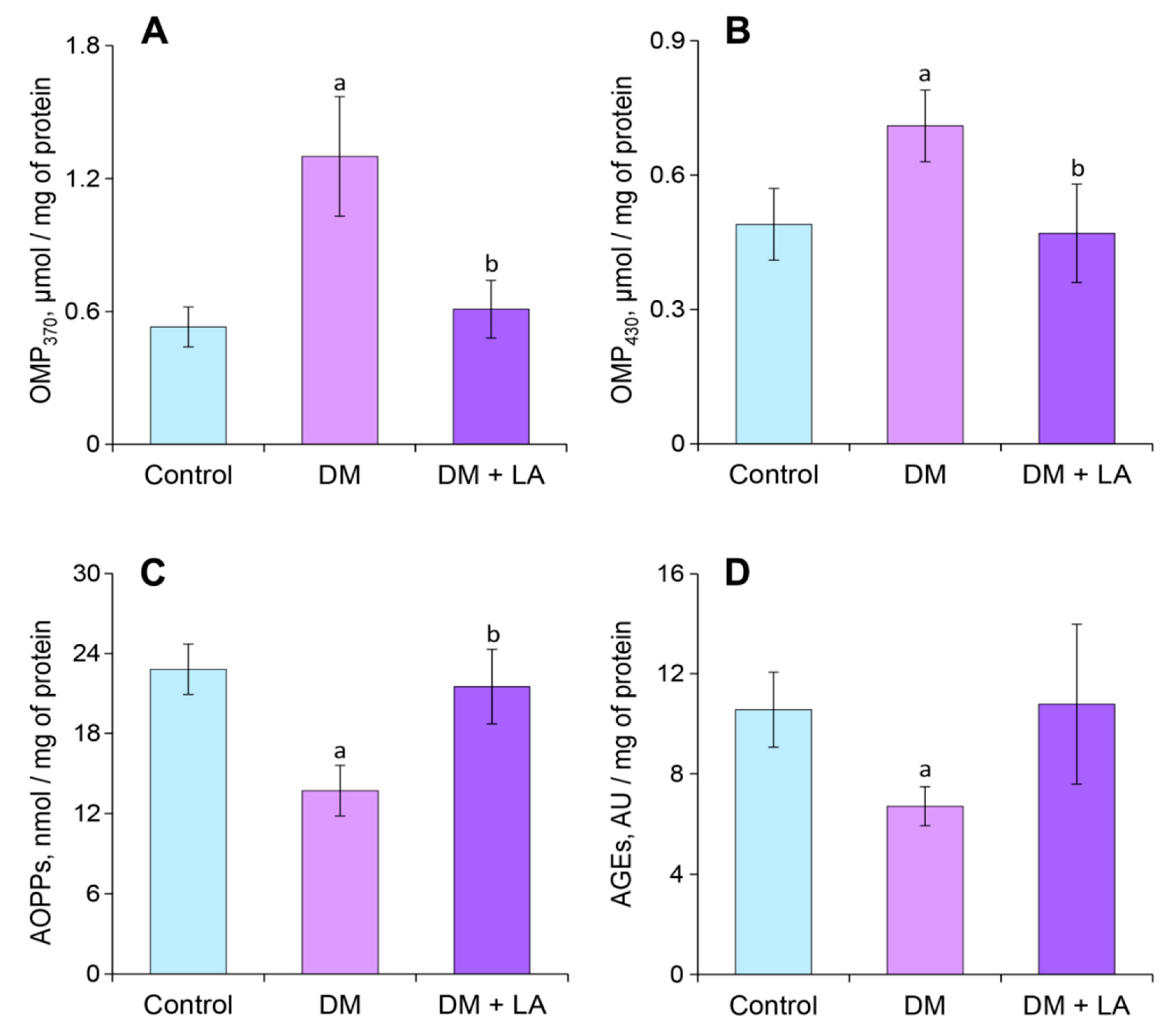
3.4. Loganic Acid Changes GSH Content and OMPs in Plasma
3.5. Loganic Acid Enhances an Antioxidant Defense System of Leukocytes
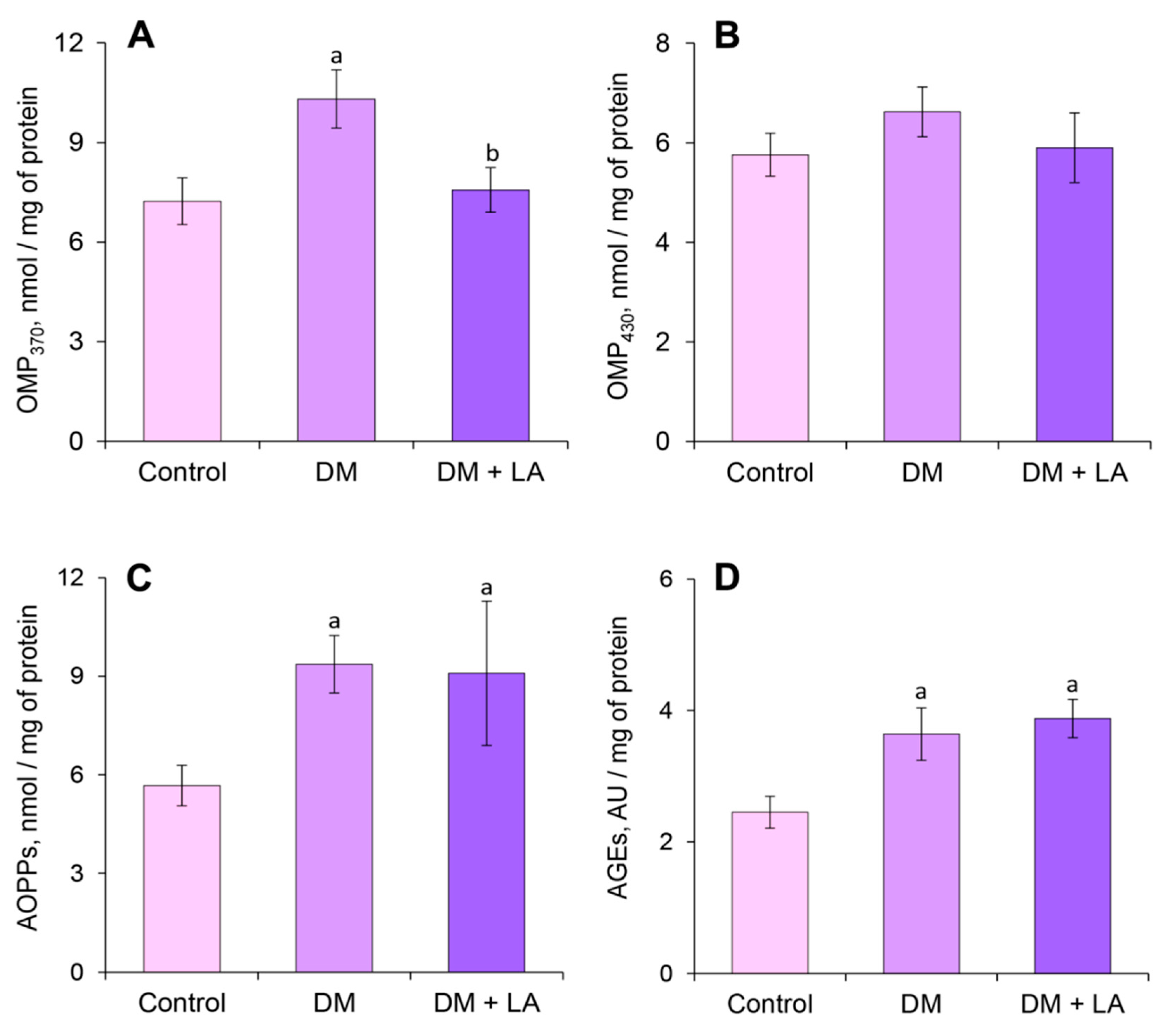
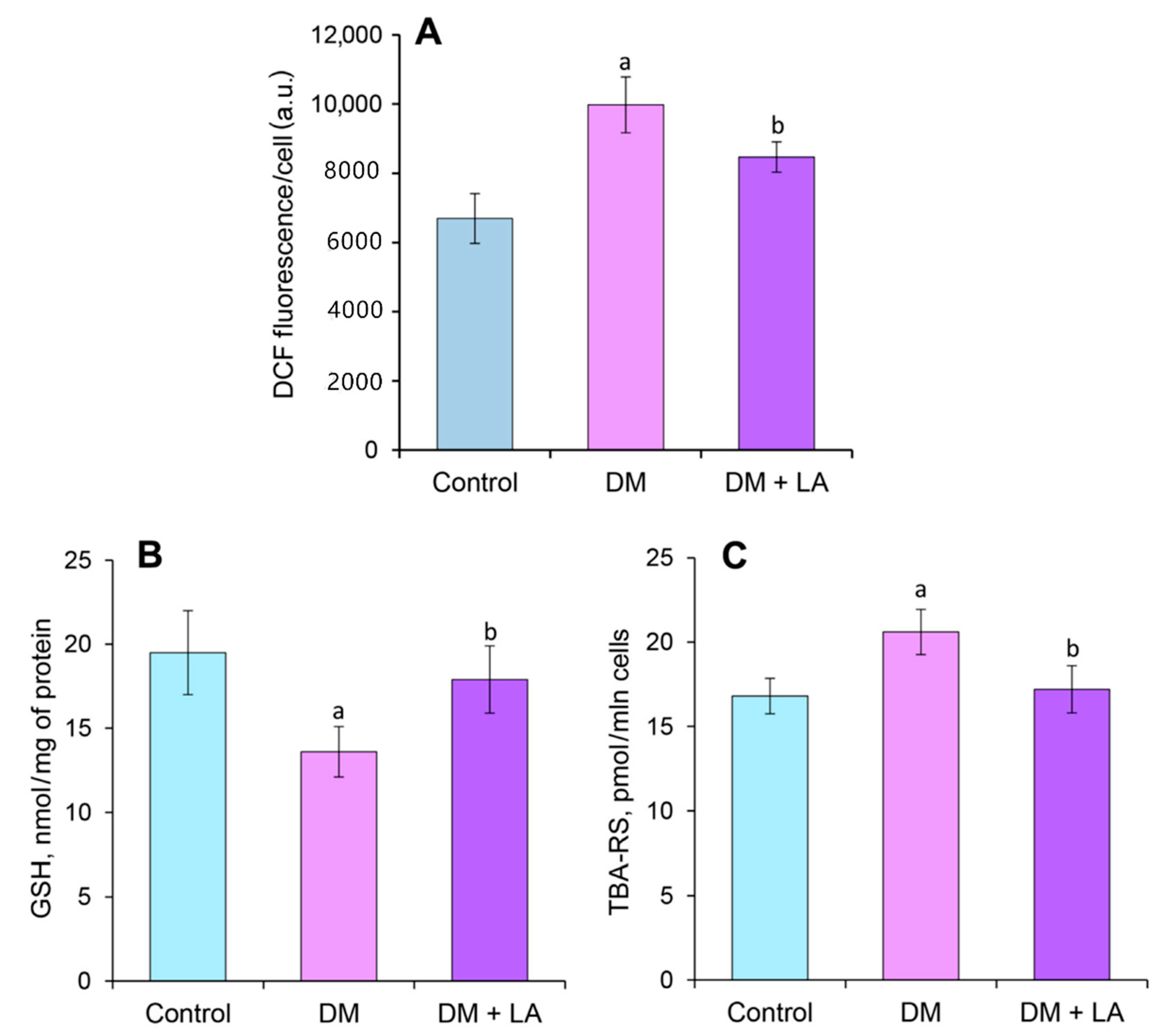
3.6. Loganic Acid Alleviates Content of Carbonyl-Oxidative Stress Metabolites in Leukocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Herold, K.C.; Vignali, D.A.; Cooke, A.; Bluestone, J.A. Type 1 diabetes: Translating mechanistic observations into effective clinical outcomes. Nat. Rev. Immunol. 2013, 13, 243–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, S.N.; Reddy, N.M.; Patil, K.R.; Nakhate, K.T.; Ojha, S.; Patil, C.R.; Agrawal, Y.O. Challenges and issues with streptozotocin-induced diabetes—A clinically relevant animal model to understand the diabetes pathogenesis and evaluate therapeutics. Chem. Biol. Interact. 2016, 244, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric Oxide and Peroxynitrite in Health and Disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yang, L.; Lei, L.; Chen, R.; Chen, H.; Zhang, H. Glucagon-like peptide-1 attenuates advanced oxidation protein product-mediated damage in islet microvascular endothelial cells partly through the RAGE pathway. Int. J. Mol. Med. 2016, 38, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Chaimum-aom, A.; Chomko, S.; Talubmook, C. Toxicology and Oral Glucose Tolerance Test (OGTT) of Thai Medicinal Plant Used for Diabetes controls, Phyllanthus acidus L. (EUPHORBIACEAE). Phcog. J. 2017, 9, 58–61. [Google Scholar] [CrossRef] [Green Version]
- Dzydzan, O.; Bila, I.; Kucharska, A.Z.; Brodyak, I.; Sybirna, N. Antidiabetic effects of extracts of red and yellow fruits of cornelian cherries (Cornus mas L.) on rats with streptozotocin-induced diabetes mellitus. Food Funct. 2019, 10, 6459–6472. [Google Scholar] [CrossRef] [Green Version]
- Jia, N.; Chu, W.; Li, Y.; Ding, L.; Duan, J.; Cui, J.; Cao, S.; Zhao, C.; Wu, Y.; Wen, A. Iridoid glycosides from the flowers of Gentiana macrophylla Pall. ameliorate collagen-induced arthritis in rats. J. Ethnopharmacol. 2016, 189, 1–9. [Google Scholar] [CrossRef]
- Recio, M.C.; Giner, R.M.; Manez, S.; Rios, J.L. Structural considerations on the iridoids as anti-inflammatory agents. Planta Med. 1994, 60, 232–234. [Google Scholar] [CrossRef]
- Ji, D.; Zhang, C.; Li, J.; Yang, H.; Shen, J.; Yang, Z. A new iridoid glycoside from the roots of Dipsacus asper. Molecules 2012, 17, 1419–1424. [Google Scholar] [CrossRef] [PubMed]
- Sozanski, T.; Kucharska, A.Z.; Rapak, A.; Szumny, D.; Trocha, M.; Merwid-Lad, A.; Dzimira, S.; Piasecki, T.; Piórecki, N.; Magdalan, J.; et al. Iridoid-loganic acid versus anthocyanins from the Cornus mas fruits (cornelian cherry): Common and different effects on diet-induced atherosclerosis, PPARs expression and inflammation. Atherosclerosis 2016, 254, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Kim, J.; Yeo, S.; Kim, G.; Ko, E.-H.; Lee, S.W.; Li, W.Y.; Choi, C.W.; Jeong, S.Y. Antiadipogenic Effects of Loganic Acid in 3T3-L1 Preadipocytes and Ovariectomized Mice. Molecules 2018, 23, 1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesavan, R.; Chandel, S.; Upadhyay, S.; Bendre, R.; Ganugula, R.; Potunuru, U.R.; Giri, H.; Sahu, G.; Kumar, P.U.; Reddy, G.B.; et al. Gentiana lutea exerts anti-atherosclerotic effects by preventing endothelial inflammation and smooth muscle cell migration. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Kucharska, A.Z.; Szumny, A.; Sokół-Łetowska, A.; Piórecki, N.; Klymenko, S.V. Iridoids and anthocyanins in cornelian cherry (Cornus mas L.) cultivars. J. Food Compos. Anal. 2015, 40, 95–102. [Google Scholar] [CrossRef]
- Yeh, S.T. Using trapezoidal rule for the area under a curve calculation. In Proceedings of the Twenty-Seventh Annual SAS. User Group International (SUGI) Conference, Orlando, FL, USA, 14–17 April 2002; Volume 27, pp. 229–237. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Souza, F.; Duncan, W.; Carvalho, R. Hematology and plasma biochemistry in rats fed with diets enriched with fatty fishes from Amazon region. Rev. Nutr. 2014, 27, 547–555. [Google Scholar] [CrossRef] [Green Version]
- Vitak, Y.; Wasser, S.P.; Nevo, E.; Sybirna, N.O. The effect of the medicinal mushrooms Agaricus brasiliensis and Ganodermalucidum (higher basidiomycetes) on the erythron system in normal and streptozotocin-induced diabetic rats. Int. J. Med. Mushrooms 2015, 17, 277–286. [Google Scholar] [CrossRef]
- Brodyak, I.V.; Sybirna, N.O. Methods of Research of Blood System; Ivan Franko National University of Lviv: Lviv, Ukraine, 2018; pp. 69–73. [Google Scholar]
- Rizvi, S.I.; Zaid, M. A intracellular reduced glutathione content in normal and type 2 diabetic erythrocytes: Effect of insulin and (-)epicatechin. J. Physiol. Pharmacol. 2001, 52, 483–488. [Google Scholar]
- Strugała, P.; Dzydzan, O.; Brodyak, I.; Kucharska, A.Z.; Kuropka, P.; Liuta, M.; Kaleta-Kuratewicz, K.; Przewodowska, A.; Michałowska, D.; Gabrielska, J.; et al. Antidiabetic and Antioxidative Potential of the Blue Congo Variety of Purple Potato Extract in Streptozotocin-Induced Diabetic Rats. Molecules 2019, 24, 3126. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Demkovych, A.; Bondarenko, Y.; Hasiuk, P.A. Oxidative modification of proteins in the process of experimental periodontitis development. Interv. Med. Appl. Sci. 2017, 9, 218–221. [Google Scholar] [CrossRef]
- Endo, Y.; Li, C.M.; Tagiri-Endo, M.; Fujimoto, K. A Modified Method for the Estimation of Total Carbonyl Compounds in Heated and Frying Oils Using 2-Propanol as a Solvent. J. Am. Oil Chem. Soc. 2001, 78, 1021–1025. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Friedlander, M.; Capeillère-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A.T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49, 1304–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalousova, M.; Skrha, J.; Zima, T. Advanced glycation end-products and advanced oxidation protein products in patients with diabetes mellitus. Physiol. Res. 2002, 51, 597–604. [Google Scholar] [PubMed]
- Putta, S.; Kilari, E.K. A review on methods of estimation of advanced glycation end products. World J. Pharm. Res. 2015, 4, 689–699. [Google Scholar]
- Kakkar, P.; Das, B.; Viswanathan, P.N. A modified spectrophotometric assay of superoxide dismutase. Ind. J. Biochem. Biophys. 1984, 21, 130–132. [Google Scholar]
- Góth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Melekh, B.; Ilkiv, I.; Lozynskyi, A.; Sklyarov, A. Antioxidant enzyme activity and lipid peroxidation in rat liver exposed to celecoxib and lansoprazole under epinephrine-induced stress. J. Appl. Pharm. Sci. 2017, 7, 94–99. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, D.M.; Spooner, R.J. Glutathione Reductase. In Methods of Enzymatic Analysis, 3rd ed.; Bergmeyer, H.U., Bergmeyer, J., GraBI, M., Eds.; Verlag Chemie: Weinheim, Germany, 1983; pp. 281–304. [Google Scholar]
- Suh, H.W.; Lee, K.B.; Kim, K.S.; Yang, H.J.; Choi, E.K.; Shin, M.H.; Park, Y.S.; Na, Y.C.; Ahn, K.S.; Jang, Y.P.; et al. A bitter herbal medicine Gentiana scabra root extract stimulates glucagon-like peptide-1 secretion and regulates blood glucose in db/db mouse. J. Ethnopharmacol. 2015, 172, 219–226. [Google Scholar] [CrossRef]
- Xie, C.; Wang, X.; Young, R.L.; Horowitz, M.; Rayner, C.K.; Wu, T. Role of Intestinal Bitter Sensingmin Enteroendocrine Hormone Secretion and Metabolic Control. Front. Endocrinol. (Laussane) 2018, 9, 576. [Google Scholar] [CrossRef]
- Huang, T.N.; Lu, K.N.; Pai, Y.P.; Chin, H.; Huang, C.J. Role of GLP-1 in the hypoglycemic effects of wild bitter gourd. Evid. Based Complement. Altern. Med. 2013, 2013, 625892. [Google Scholar] [CrossRef] [Green Version]
- Bouderbala, S.; Prost, J.; Lacaille-Dubois, M.A.; Bouchenak, M. Iridoid extracts from Ajuga iva increase the antioxidant enzyme activities in red blood cells of rats fed a cholesterol-rich diet. Nutr. Res. 2010, 30, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Lehrke, M.; Lazar, M.A. The many faces of PPARgamma. Cell 2005, 123, 993–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.H.; Yamabe, N.; Noh, J.S.; Kang, K.S.; Tanaka, T.; Yokozawa, T. The beneficial effects of morroniside on the inflammatory response and lipid metabolism in the liver of db/db mice. Biol. Pharm. Bull. 2009, 32, 1734–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.H.; Noh, J.S.; Tanaka, T.; Yokozawa, T. Effects of morroniside isolated from Corni Fructus on renal lipids and inflammation in type 2 diabetic mice. J. Pharm. Pharmacol. 2010, 62, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Yamabe, N.; Noh, J.S.; Park, C.H.; Kang, K.S.; Shibahara, N.; Tanaka, T.; Yokozawa, T. Evaluation of loganin, iridoid glycoside from Corni Fructus, on hepatic and renal glucolipotoxicity and inflammation in type 2 diabetic db/db mice. Eur. J. Pharmacol. 2010, 648, 179–187. [Google Scholar] [CrossRef]
- Gryszczyńska, B.; Budzyń, M.; Formanowicz, D.; Formanowicz, P.; Krasiński, Z.; Majewska, N.; Iskra, M.; Kasprzak, M.P. Advanced Oxidation Protein Products and Carbonylated Proteins Levels in Endovascular and Open Repair of an Abdominal Aortic Aneurysm: The Effect of Pre-, Intra-, and Postoperative Treatment. BioMed Res. Int. 2019, 2019, 7976043. [Google Scholar] [CrossRef] [Green Version]
- Capeillere-Blandin, C.; Gausson, V.; Descamps-Latscha, B.; Witko-Sarsat, V. Biochemical and spectrophotometric significance of advanced oxidized protein products. Biochim. Biophys. Acta 2004, 1689, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Xiao, Y.; Xu, A.; Zhou, Z. Neutrophils in type 1 diabetes. J. Diabetes Investig. 2016, 7, 652–663. [Google Scholar] [CrossRef]
- Heilman, K.; Zilmer, M.; Zilmer, K.; Lintrop, M.; Kampus, P.; Kals, J.; Tillmann, V. Arterial stiffness, carotid artery intima-media thickness and plasma myeloperoxidase level in children with type 1 diabetes. Diabetes Res. Clin. Pract. 2009, 84, 168–173. [Google Scholar] [CrossRef]
- Bianchi, C.; Del Prato, S. Metabolic memory and individual treatment aims in type 2 diabetes—outcome-lessons learned from large clinical trials. Rev. Diabet. Stud. 2011, 8, 432–440. [Google Scholar] [CrossRef] [Green Version]
- Maratou, E.; Dimitriadis, G.; Kollias, A.; Boutati, E.; Lambadiari, V.; Mitrou, P.; Raptis, S.A. Glucose transporter expression on the plasma membrane of resting and activated white blood cells. Eur. J. Clin. Investig. 2007, 37, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Vrhovac, I.; Breljak, D.; Sabolić, I. Glucose transporters in the mammalian blood cells. Periodicum Biologorum 2014, 116, 131–138. [Google Scholar]
- Martin, S.P.; McKinney, G.R.; Green, R.; Becker, C. The influence of glucose, fructose, and insulin on the metabolism of leukocytes of healthy and diabetic subjects. J. Clin. Investig. 1953, 32, 1171–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmoch, T.; Uhle, F.; Siegler, B.H.; Fleming, T.; Morgenstern, J.; Nawroth, P.P.; Weigand, M.A.; Brenner, T. The Glyoxalase System and Methylglyoxal-Derived Carbonyl Stress in Sepsis: Glycotoxic Aspects of Sepsis Pathophysiology. Int. J. Mol. Sci. 2017, 18, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stjernholm, R.L. Carbohydrate Metabolism in Leukocytes. J. Bact. 1967, 93, 1657–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abirami, A.; Sinsinwar, S.; Rajalakshmi, P.; Brindha, P.; Rajesh, Y.B.R.D.; Vadivel, V. Antioxidant and cytoprotective properties of loganic acid isolated from seeds of Strychnos potatorum L. against heavy metal induced toxicity in PBMC model. Drug Chem. Toxicol. 2019, 1–11. [Google Scholar] [CrossRef]
- Gomes, E.C.; Silva, A.N.; de Oliveira, M.R. Oxidants, antioxidants, and the beneficial roles of exercise-induced production of reactive species. Oxid. Med. Cell. Longev. 2012, 2012, 756132. [Google Scholar] [CrossRef]
- Bila, I.; Dzydzan, O.; Brodyak, I.; Sybirna, N. Agmatine prevents oxidative-nitrative stress in blood leukocytes under streptozotocin-induced diabetes. Open Life Sci. 2019, 14, 299–310. [Google Scholar] [CrossRef]
- Daiber, A.; Mader, M.; Stamm, P.; Zinßius, E.; Kröller-Schön, S.; Oelze, M.; Münzel, T. Oxidative Stress and Vascular Function. Cell Memb. Free Radic. Res. 2013, 5, 221–231. [Google Scholar]
- Olivares-Vicente, M.; Sanchez-Marzo, N.; Encinar, J.A.; de la Luz Cadiz-Gurrea, M.; Lozano-Sanchez, J.; Segura-Carretero, A.; Arraez-Roman, D.; Riva, C.; Barrajón-Catalán, E.; Herranz-López, M.; et al. The Potential Synergistic Modulation of AMPK by Lippia citriodora Compounds as a Target in Metabolic Disorders. Nutrients 2019, 11, 2961. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.P.; Brown, J.R.; Sag, D.; Zhang, L.; Suttles, J. Adenosine 59-Monophosphate–Activated Protein Kinase Regulates IL-10–Mediated Anti-Inflammatory Signaling Pathways in Macrophages. J. Immunol. 2015, 194, 584–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawczyk, C.M.; Holowka, T.; Sun, J.; Blagih, J.; Amiel, E.; DeBerardinis, R.J.; Cross, J.R.; Jung, E.; Thompson, C.B.; Jones, R.G.; et al. Toll-like receptor-induced changes in glycolytic metabolism regulate dendritic cell activation. Blood 2010, 115, 4742–4749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, E.; Nishikawa, T. Oxidative stress: A cause and therapeutic target of diabetic complications. J. Diabetes Investig. 2010, 1, 90–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | UV λmax (nm) | [M−H]− (m/z) | Other Ions (m/z) | Content (mg/100 g dw) |
|---|---|---|---|---|
| Iridoids | ||||
| Loganic acid 1 | 246 | 375 | 213 | 183.1 ± 27.4 |
| Loganic acid 2 a | 246 | 375 | 213 | 70,228.5 ± 548.6 |
| Loganic acid 3 | 246 | 375 | 213 | 3581.7 ± 73.1 |
| Total | 73,993.3 | |||
| Phenolic acids | ||||
| Caftaric acid 1 | 327 | 179/311 | 149/135 | 721.7 ± 10.1 |
| Caftaric acid 2 a | 328 | 179/311 | 149/135 | 1110.2 ± 20.0 |
| Total | 1831.9 | |||
| Parameters | Groups | ||
|---|---|---|---|
| Control | DM | DM + LA | |
| Leukocytes, 103/µL | 10.1 ± 0.8 | 11.2 ± 0.7 | 12.0 ± 0.8 |
| Banded neutrophilis, % | 1.4 ± 0.2 | 1.3 ± 0.4 | 1.5 ± 0.3 |
| Segmented neutrophilis, % | 21.7 ± 1.3 | 16.3 ± 1.6 a | 19.2 ± 1.8 |
| Eosinophils, % | 1.2 ± 0.4 | 1.0 ± 0.3 | 1.3 ± 0.3 |
| Lymphocytes, % | 74.4 ± 1.5 | 80.4 ± 1.3 a | 77.5 ± 2.6 |
| Monocytes, % | 0.5 ± 0.2 | 0.4 ± 0.3 | 0.5 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dzydzan, O.; Brodyak, I.; Sokół-Łętowska, A.; Kucharska, A.Z.; Sybirna, N. Loganic Acid, an Iridoid Glycoside Extracted from Cornus mas L. Fruits, Reduces of Carbonyl/Oxidative Stress Biomarkers in Plasma and Restores Antioxidant Balance in Leukocytes of Rats with Streptozotocin-Induced Diabetes Mellitus. Life 2020, 10, 349. https://doi.org/10.3390/life10120349
Dzydzan O, Brodyak I, Sokół-Łętowska A, Kucharska AZ, Sybirna N. Loganic Acid, an Iridoid Glycoside Extracted from Cornus mas L. Fruits, Reduces of Carbonyl/Oxidative Stress Biomarkers in Plasma and Restores Antioxidant Balance in Leukocytes of Rats with Streptozotocin-Induced Diabetes Mellitus. Life. 2020; 10(12):349. https://doi.org/10.3390/life10120349
Chicago/Turabian StyleDzydzan, Olha, Iryna Brodyak, Anna Sokół-Łętowska, Alicja Z. Kucharska, and Natalia Sybirna. 2020. "Loganic Acid, an Iridoid Glycoside Extracted from Cornus mas L. Fruits, Reduces of Carbonyl/Oxidative Stress Biomarkers in Plasma and Restores Antioxidant Balance in Leukocytes of Rats with Streptozotocin-Induced Diabetes Mellitus" Life 10, no. 12: 349. https://doi.org/10.3390/life10120349