
Parameter Estimation Analysis in a Model of Honey Production


Abstract
:1. Introduction
2. Mathematical Model
3. Parameter Identification
4. Numerical Experiments
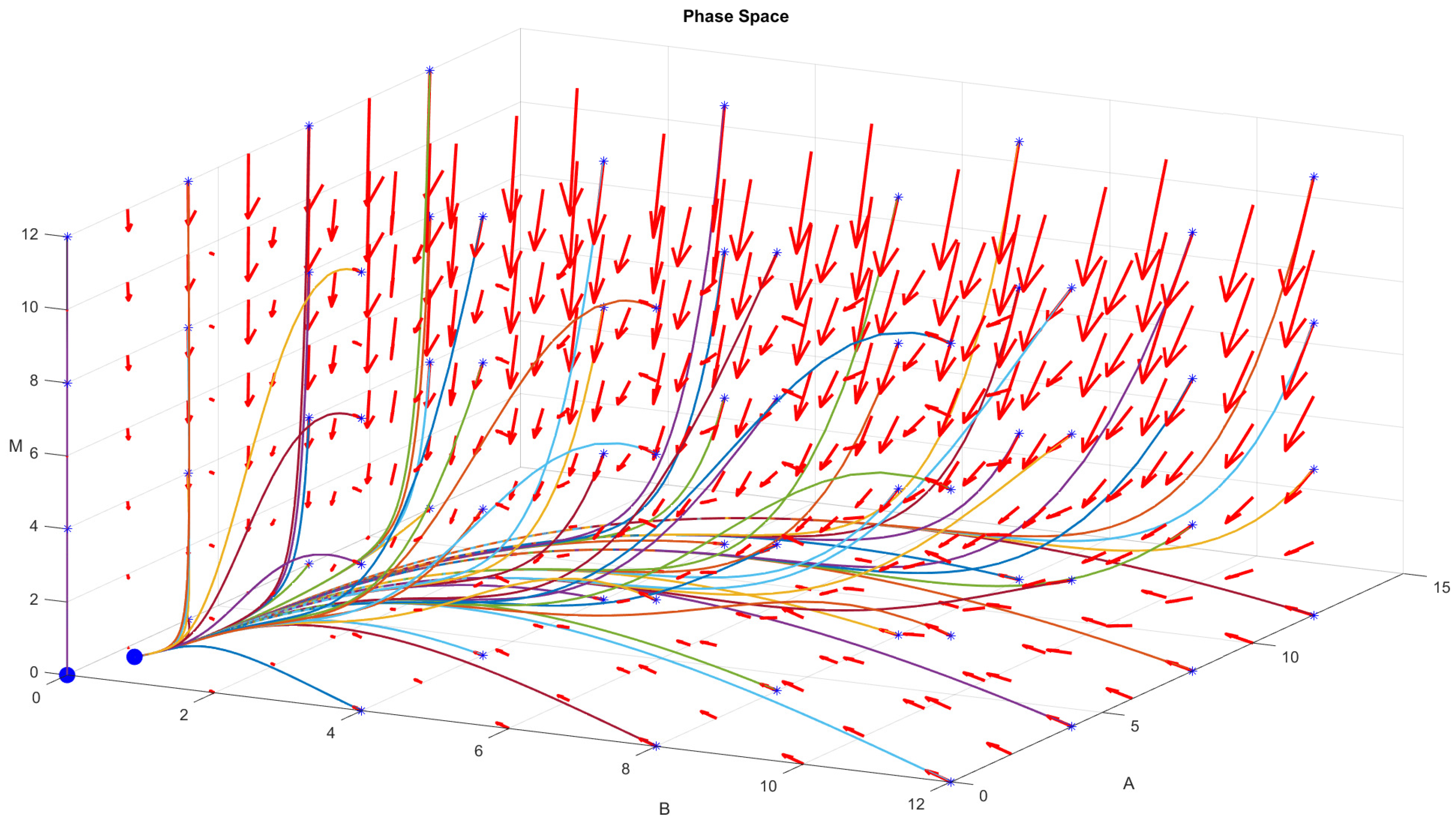
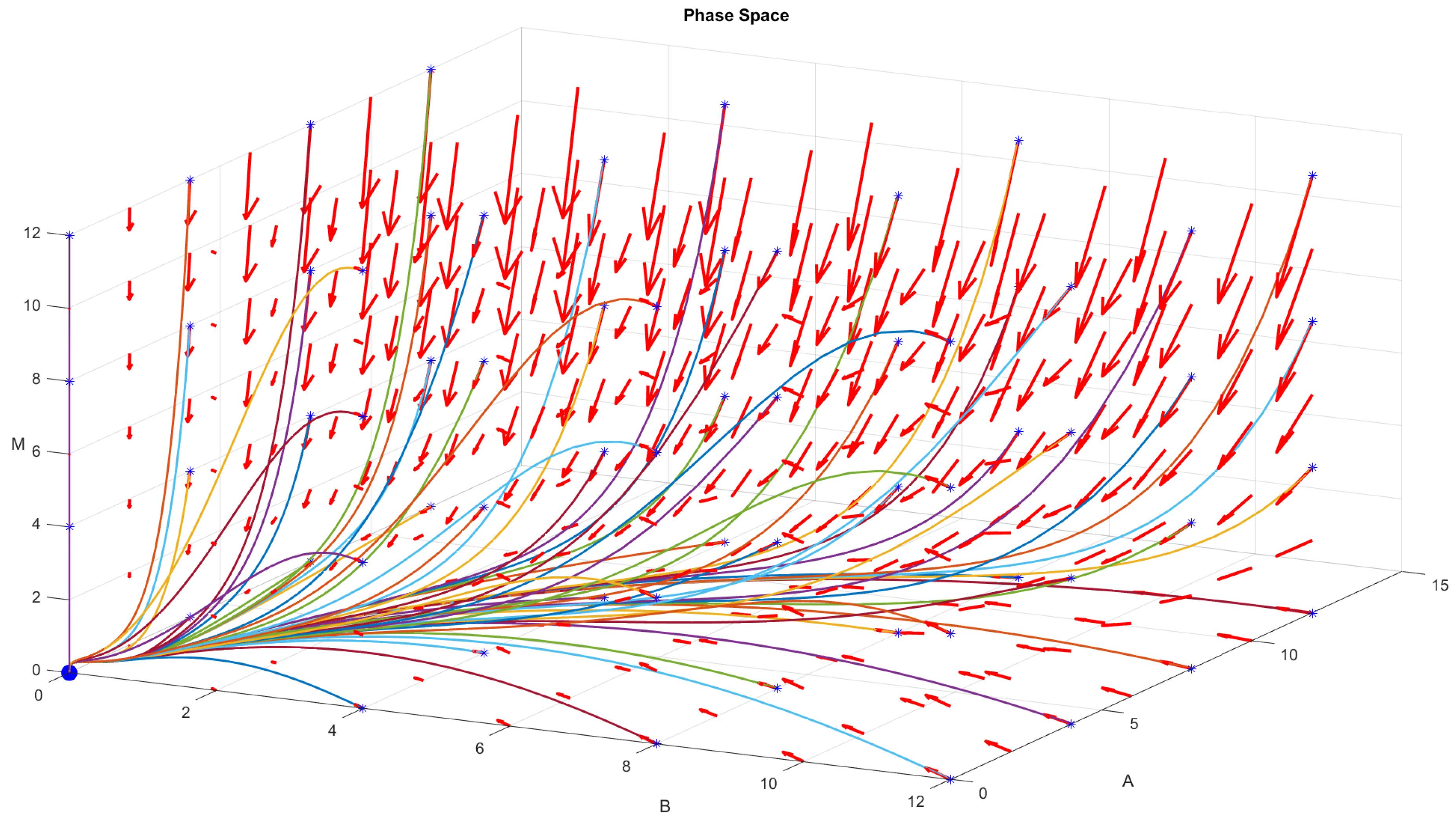
4.1. Numerical Procedure
4.2. Direct Problem
4.3. Inverse Problem
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Woodard, S.; Fischman, B.; Venkat, A.; Hudson, M.; Varala, K.; Cameron, S.; Clark, A.; Robinson, G. Genes involved in convergent evolution of eusociality in bees. Proc. Natl. Acad. Sci. USA 2011, 108, 7472–7477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matilla, H.; Seeley, T. Genetic diversity in honey bee colonies enhances productivity and fitness. Science 2007, 317, 362–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farouk, K.; Palmera, K.; Sepúlveda, P. Abejas. In InfoZoa Boletín de Zoología; Universidad del Magdalena: Santa Marta, Colombia, 2014; Volume 6. [Google Scholar]
- Bulgarian Honey. 2023. Available online: https://www.bulgarianhoney.com/quality.htm (accessed on 7 February 2023).
- Khoury, D.S.; Myerscough, M.R.; Barron, A.B. A quantitative model of honey bee colony population dynamics. PLoS ONE 2011, 6, e18491. [Google Scholar] [CrossRef] [Green Version]
- Khoury, D.S.; Barron, A.B.; Meyerscough, M.R. Modelling food and population dynamics honey bee colonies. PLoS ONE 2013, 8, e0059084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyerscough, M.R.; Khoury, D.S.; Ronzani, S.; Barron, A.B. Why do hives die? Using mathematics to solve the problem of honey bee colony collapse. In The Role and Importance of Mathematics in Innovation: Proceedings of the Forum “Math-for-Industry”; Anderssen, B., Ed.; Springer: Singapore, 2017; Volume 25, pp. 35–50. [Google Scholar]
- Russel, S.; Barron, A.B.; Harris, D. Dynamics modelling of honeybee (Apis mellifera) colony growth and failure. Ecol. Model. 2013, 265, 138–169. [Google Scholar]
- Booton, R.D.; Iwasa, Y.; Marshall, J.A.R.; Childs, D.Z. Stress-mediated Allee effects can cause the sudden collapse of honey bee colonies. J. Theor. Biol. 2017, 420, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Finley, J.; Camazine, S.; Frazier, M. The epidemic of honey bee colony losses during the 1995–1996 season. Am. Bee J. 1996, 136, 805–808. [Google Scholar]
- Amdam, G.V.; Omholt, S.W. The hive bee to forager transition in honeybee colonies: The double repressor hypothesis. J. Theor. Biol. 2003, 223, 451–464. [Google Scholar] [CrossRef]
- van der Zee, R.; Pisa, L.; Andronov, S.; Brodschneider, R.; Charriere, J.D.; Chlebo, R.; Coffey, M.F.; Cralisheim, K.; Dahle, B.; Gajda, A.; et al. Managed honey bee colony losses in Canada, China, Europe, Israel and Turkey for the winters of 2008–2009 and 2009–2010. J. Apic. Res. 2012, 51, 100–114. [Google Scholar] [CrossRef] [Green Version]
- Bailey, L. The ‘Isle of Wight Disease’: The Origin and Significance of the Myth. Bee World 1964, 45, 32–37. [Google Scholar] [CrossRef]
- Kulincevic, J.M.; Rothenbuhler, W.C.; Rinderer, T.E. Disappearing disease. Part 1—Effects of certain protein sources given to honey bee colonies in Florida. Am. Bee J. 1982, 122, 189–191. [Google Scholar]
- Dornberger, L.; Mitchell, C.; Hull, B.; Ventura, W.; Shopp, H.; Kribs-Zaleta, C.; Kojouharov, H.; Grover, J. Death of the Bees: A Mathematical Model of Colony Collapse Disorder; Technical Report 2012-12, Mathematics Preprint Series; University of Texas at Arlington Mathematics Department: Arlington, TX, USA, 2012. [Google Scholar]
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors associated with honey bee colony losses: A mini-review. Vet. Sci. 2020, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, S.; Mirzaie, M. A mathematical model of honey bee colony dynamics to predict the effect of pollen on colony failure. PLoS ONE 2019, 14, e0225632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betti, M.I.; Wahl, L.M.; Zamir, M. Reproduction number and asymptotic stability for the dynamics of a honey bee colony with continuous age structure. Bull. Math. Biol. 2017, 79, 1586–1611. [Google Scholar] [CrossRef] [Green Version]
- Switanek, M.; Crailsheim, K.; Truhetz, H.; Brodschneider, R. Modelling seasonal effects of temperature and precipitation on honey bee winter mortality in a temperate climate. Sci. Total Environ. 2017, 579, 1581–1587. [Google Scholar] [CrossRef]
- Ratti, V.; Kevan, P.G.; Eberl, H.J. A mathematical model of forager loss in honeybee colonies infested with Varroa destructor and the acute bee paralysis virus. Bull. Math. Biol. 2017, 79, 1218–1253. [Google Scholar] [CrossRef]
- Becher, M.A.; Osborne, J.L.; Thorbek, P.; Kennedy, P.J.; Grimm, V. Review: Towards a systems approach for understanding honeybee decline: A stocktaking and synthesis of existing models. J. Appl. Ecol. 2013, 50, 868–880. [Google Scholar] [CrossRef] [Green Version]
- Torres, D.J.; Ricoy, V.M.; Roybal, S. Modelling honey bee populations. PLoS ONE 2015, 10, e0130966. [Google Scholar] [CrossRef]
- Yıldız, T.A. A fractional dynamical model for honeybee colony population. Int. J. Biomath. 2018, 11, 1850063. [Google Scholar] [CrossRef]
- Romero-Leiton, J.P.; Gutierrez, A.; Benavides, I.F.; Molina, O.E.; Pulgarín, A. An approach to the modeling of honey bee colonies. Web Ecol. 2013, 22, 7–19. [Google Scholar] [CrossRef]
- Atanasov, A.Z.; Georgiev, S.G.; Vulkov, L.G. Reconstruction analysis of honeybee colony collapse disorder modeling. Optim. Eng. 2021, 22, 2481–2503. [Google Scholar] [CrossRef]
- Atanasov, A.Z.; Georgiev, S.G.; Vulkov, L.G. Parameter identification of Colony Collapse Disorder in honeybees as a contagion. In Modelling and Development of Intelligent Systems; Simian, D., Stoica, L.F., Eds.; Springer: Dordrecht, The Netherlands, 2021; Volume 1341, pp. 363–377. [Google Scholar]
- Hong, W.; Chen, B.; Lu, Y.; Lu, C.; Liu, S. Using system equalization principle to study the effects of multiple factors to the development of bee colony. Ecol. Model. 2022, 470, 110002. [Google Scholar] [CrossRef]
- Hundsdorfer, W.; Vermer, J. Numerical Solution of Time-Dependent Advection-Diffusion-Reaction Equations; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Marchuk, G.I. Adjoint Equations and Analysis of Complex Systems; Kluwer: Dordrecht, The Netherlands, 1995. [Google Scholar]
- Marchuk, G.I.; Agoshkov, V.I.; Shutyaev, V.P. Adjoint Equations and Perturbation Algorithms in Nonlinear Problems; CRC Press: Boca Raton, FL, USA, 1996. [Google Scholar]
- Winston, W.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, MA, USA, 1991. [Google Scholar]
- Ma, C.; Jiang, L. Some research on Levenberg–Marquardt method for the nonlinear equations. Appl. Math. Comp. 2007, 184, 1032–1040. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | ||||||
|---|---|---|---|---|---|---|
| 0.018 | 0.02 | 0.0274 | 0.0094 | 0.5234 | ||
| 0.12 | 0.10 | 0.0991 | 0.0209 | 0.1746 | ||
| 0.571 | 0.50 | 0.3633 | 0.2077 | 0.3638 | ||
| 0.1 | 0.20 | 0.1886 | 0.0886 | 0.8862 | ||
| 0.95 | 1.00 | 0.8992 | 0.0508 | 0.0535 |
| Parameter | ||||||
|---|---|---|---|---|---|---|
| 0.018 | 0.02 | 0.0269 | 0.0089 | 0.4942 | ||
| 0.12 | 0.10 | 0.0989 | 0.0211 | 0.1758 | ||
| 0.571 | 0.50 | 0.3731 | 0.1979 | 0.3465 | ||
| 0.1 | 0.20 | 0.1869 | 0.0869 | 0.8689 | ||
| 0.95 | 1.00 | 0.8927 | 0.0573 | 0.0603 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atanasov, A.Z.; Georgiev, S.G.; Vulkov, L.G. Parameter Estimation Analysis in a Model of Honey Production. Axioms 2023, 12, 214. https://doi.org/10.3390/axioms12020214
Atanasov AZ, Georgiev SG, Vulkov LG. Parameter Estimation Analysis in a Model of Honey Production. Axioms. 2023; 12(2):214. https://doi.org/10.3390/axioms12020214
Chicago/Turabian StyleAtanasov, Atanas Z., Slavi G. Georgiev, and Lubin G. Vulkov. 2023. "Parameter Estimation Analysis in a Model of Honey Production" Axioms 12, no. 2: 214. https://doi.org/10.3390/axioms12020214