
Late Cretaceous (Santonian to Campanian) Palynological Records and Paleoclimatic Significance from Borehole ZKY2-1, Songliao Basin

(This article belongs to the Section Biomineralization and Biominerals)
Abstract
:1. Introduction
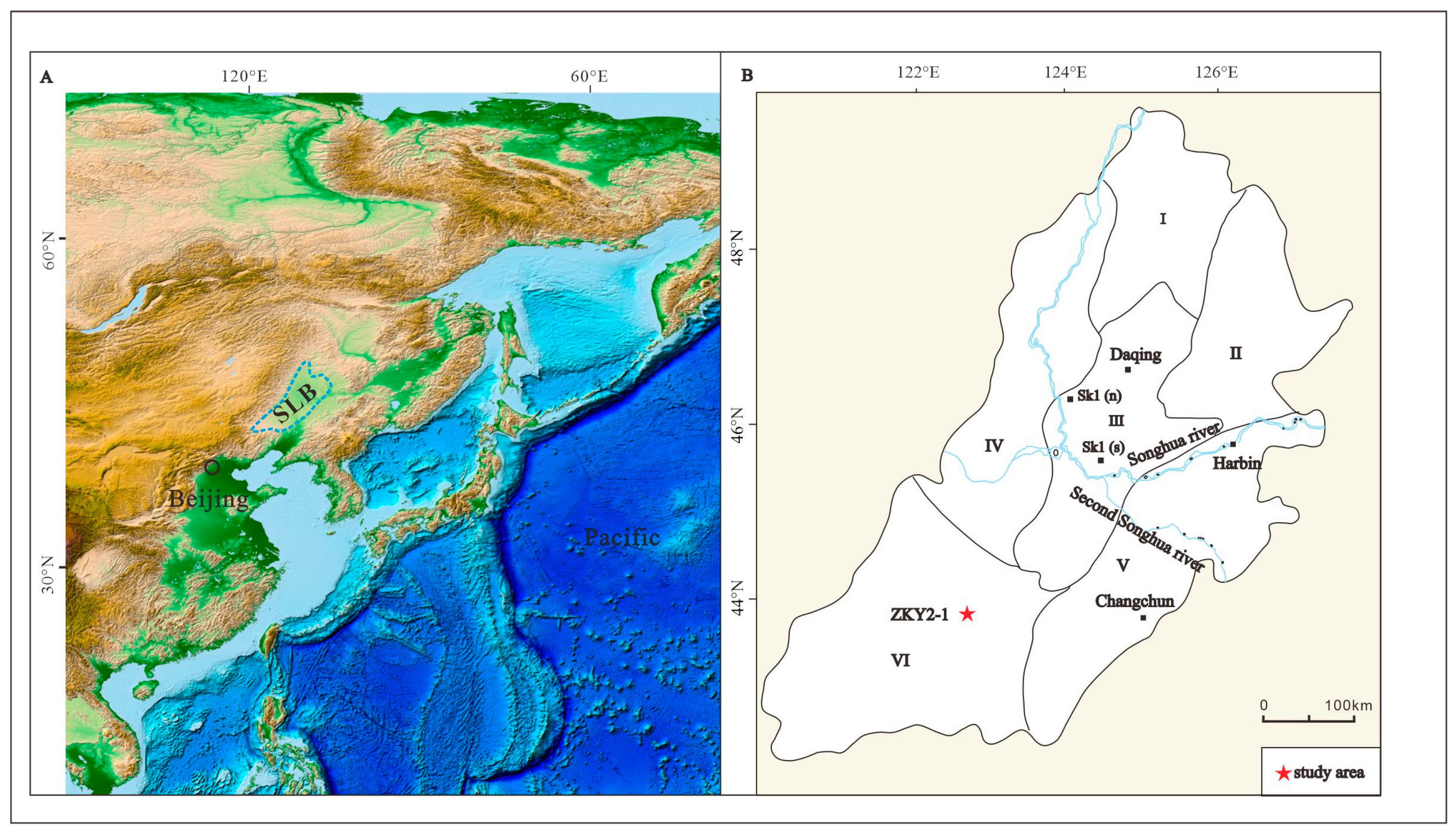
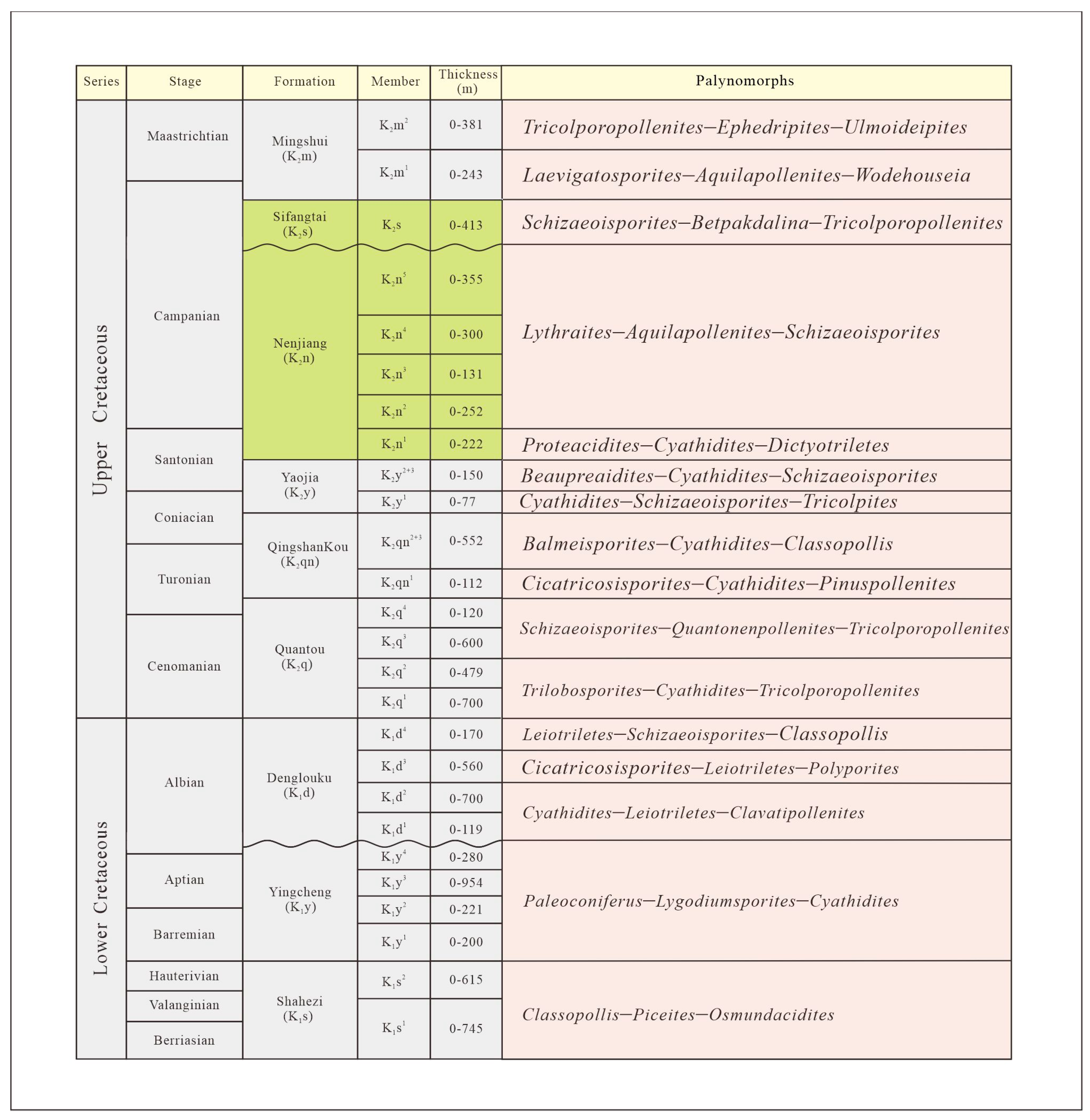
2. Geological Setting
3. Materials and Methods
4. Results
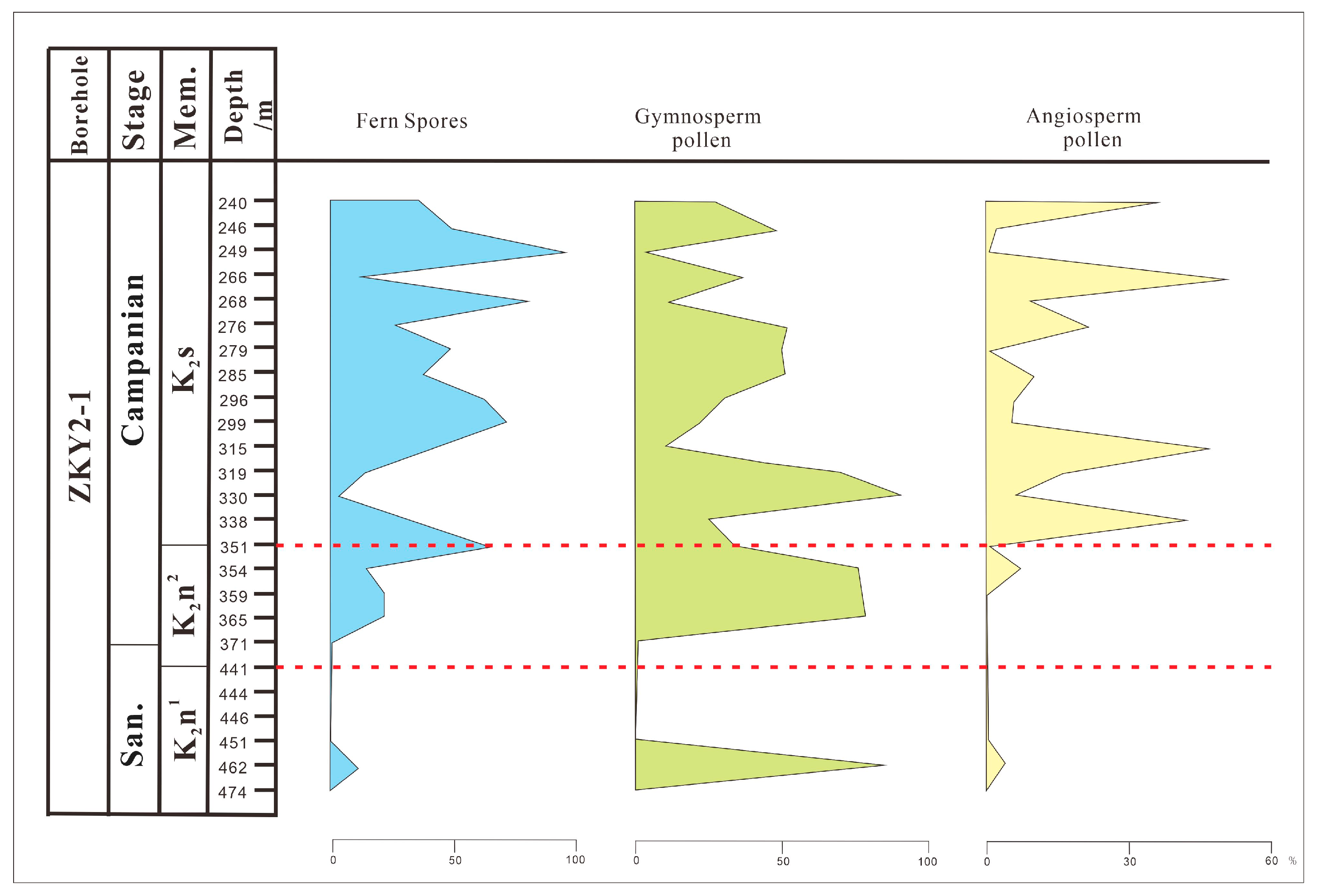
Palynomorphs from Borehole ZKY2-1
5. Discussion
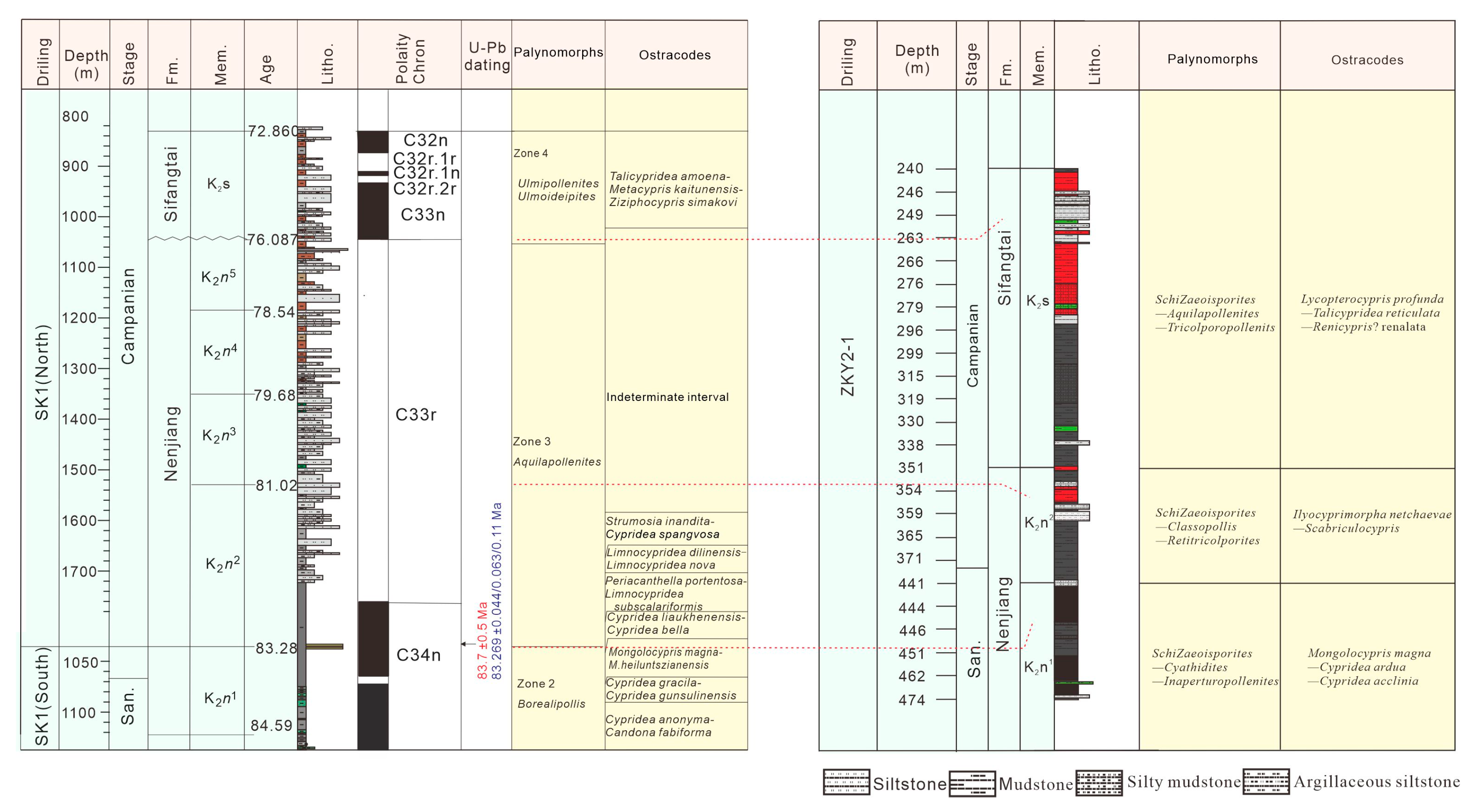
5.1. Stratigraphic Correlation between Boreholes ZKY2-1 and SK1
5.2. Vegetation Change during the Santonian–Campanian Period
5.3. Climate Change during the Late Santonian–Campanian Period
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Brien, C.L.; Robinson, S.A.; Pancost, R.D.; Sinninghe Damsté, J.S.; Schouten, S.; Lunt, D.J.; Alsenz, H.; Bornemann, A.; Bottini, C.; Brassell, S.C.; et al. Cretaceous sea-surface temperature evolution: Constraints from TEX86 and planktonic foraminiferal oxygen isotopes. Earth Sci. Rev. 2017, 172, 224–247. [Google Scholar] [CrossRef]
- Haq, B.U. Cretaceous eustasy revisited. Glob. Planet Chang. 2014, 113, 44–58. [Google Scholar] [CrossRef]
- Hay, W.W. Toward understanding Cretaceous climate–An updated review. Sci. China Earth Sci. 2017, 60, 5–19. [Google Scholar] [CrossRef]
- Jenkyns, H.C. Geochemistry of oceanic anoxic events. Geochem. Geophys. Geosyst. 2010, 11, 3. [Google Scholar] [CrossRef]
- Sames, B.; Wagreich, M.; Wendler, J.E.; Haq, B.U.; Conrad, C.P.; Melinte-Dobrinescu, M.C.; Hu, X.; Wendler, I.; Wolfgring, E.; Yilmaz, I.Ö.; et al. Review: Short-term sea–level changes in a greenhouse world–A view from the Cretaceous. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 441, 393–411. [Google Scholar] [CrossRef] [Green Version]
- Luft-Souza, F.; Fauth, G.; Bruno, M.D.R.; De Lira Mota, M.A.; Vázquez-García, B.; Santos Filho, M.A.B.; Terra, G.J.S. Sergipe–Alagoas Basin, Northeast Brazil: A reference basin for studies on the early history of the South Atlantic Ocean. Earth Sci. Rev. 2022, 229, 104034. [Google Scholar] [CrossRef]
- Huber, B.T.; MacLeod, K.G.; Watkins, D.K.; Coffin, M.F. The rise and fall of the Cretaceous Hot Greenhouse climate. Glob. Planet. Chang. 2018, 167, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Petrizzo, M.R.; MacLeod, K.G.; Watkins, D.K.; Wolfgring, E.; Huber, B.T. Late Cretaceous paleoceanographic evolution and the onset of cooling in the Santonian at southern high latitudes (IODP site U1513, SE Indian Ocean). Paleoceanogr. Paleoclimatol. 2022, 37, e2021PA004353. [Google Scholar] [CrossRef]
- Forster, A.; Schouten, S.; Moriya, K.; Wilson, P.A.; Sinninghe Damsté, J.S. Tropical warming and intermittent cooling during the Cenomanian/Turonian Oceanic Anoxic Event 2: Sea surface temperature records from the equatorial Atlantic. Paleoceanography 2007, 22. [Google Scholar] [CrossRef]
- Van Helmond, N.A.; Sluijs, A.; Reichart, G.J.; Sinninghe Damsté, J.S.; Slomp, C.P.; Brinkhuis, H. A perturbed hydrological cycle during Oceanic Anoxic Event. Geology 2014, 42, 123–126. [Google Scholar] [CrossRef]
- Zhang, L.M.; Wang, C.S.; Wignall, P.B.; Kluge, T.; Wan, X.; Wang, Q.; Gao, Y. Deccan volcanism caused coupled pCO2 and terrestrial temperature rises, and pre-impact extinctions in northern China. Geology 2018, 46, 271–274. [Google Scholar] [CrossRef]
- Perez Loinaze, V.S.; Giordano, S.R.; Limarino, C.O. Late Cretaceous palynomorphs from the Golfo San Jorge Basin, Argentina. J. S. Am. Earth Sci. 2021, 107, 103151. [Google Scholar] [CrossRef]
- Song, Z.C.; Shang, Y.K.; Liu, Z.S.; Huang, P.; Wang, X.F.; Qian, L.J.; Du, B.A.; Zhang, D.H. Palynology from China Volume II. In Mesozoic Palynology; Science Press: Beijing, China, 2000. [Google Scholar]
- Heimhofer, U.; Hochuli, P.A.; Burla, S.; Oberli, F.; Adatte, T.; Dinis, J.L.; Weissert, H. Climate and vegetation history of western Portugal inferred from Albian near-shore deposits (Gale Formation, Lusitanian Basin). Geol. Mag. 2012, 149, 1046–1064. [Google Scholar] [CrossRef] [Green Version]
- Li, J.G.; Batten, D.J.; Zhang, Y. Palynological record from a composite core through Late Cretaceous-Early Paleocene deposits in the Songliao Basin, Northeast China and its biostratigraphic implications. Cretac. Res. 2011, 32, 1–12. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Dai, S.; Heimhofer, U.; Wu, M.; Wang, Z.; Pan, B. Palynological records from two cores in the Gongpoquan Basin, central East Asia: Evidence for floristic and climatic change during the Late Jurassic to Early Cretaceous. Rev. Palaeobot. Palynol. 2014, 204, 1–17. [Google Scholar] [CrossRef]
- Gao, R.Q.; Zhao, C.B.; Qiao, X.Y. Palynology of Cretaceous Petroleum Strata in Songliao Basin; The Geological Publishing House: Beijing, China, 1999. [Google Scholar]
- Song, Z.C.; Zheng, Y.H.; Li, M.Y.; Zhang, Y.Y.; Wang, W.M.; Wang, D.N.; Zhao, C.B.; Zhou, S.F.; Zhu, Z.H.; Zhao, Y.N. Palynology from China Vol. 1 Late Cretaceous and Tertiary Palynology; Science Press: Beijing, China, 1999. [Google Scholar]
- Zhang, M.Z.; Dai, S.; Ji, L.M.; Du, B.X.; Hu, S.S. Potential links between typical greenhouse climate of the Mid-Cretaceous and radiation evolution of early angiosperms. In Proceedings of the Abstracts of the 12th National Congress and 29th Annual Conference of the Paleontological Society of China, Zhengzhou, China, 17 September 2018; pp. 140–141. [Google Scholar]
- Feng, Z.H.; Fang, W.; Wang, X.; Huang, C.Y.; Huo, Q.L.; Zhang, J.H.; Huang, Q.H.; Zhang, L. Micropaleontological and molecular fossil evidence for controlling oil shale formation by transgression in Songliao Basin. Sci. China (Ser. D) Geosci. 2009, 39, 1375–1386. [Google Scholar]
- Wang, C.S.; Feng, Z.Q.; Zhang, L.M.; Huang, Y.; Cao, K.; Wang, P.; Zhao, B. Cretaceous paleogeography and paleoclimate and the setting of SKI borehole sites in Songliao Basin, northeast China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 385, 17–30. [Google Scholar] [CrossRef]
- Wan, X.Q.; Zhao, J.; Scott, R.W.; Wang, P.; Feng, Z.; Huang, Q.; Xi, D. Late Cretaceous stratigraphy, Songliao Basin, NE China: SK1 cores. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 385, 31–43. [Google Scholar] [CrossRef]
- Xi, D.P.; Wan, X.Q.; Li, G.B.; Li, G. Integrated stratigraphy and time frame of the Cretaceous in China. Chin. Sci. (Earth Sci.) 2019, 49, 257–288. [Google Scholar]
- Xi, D.P.; Li, S.; Jing, X.; Huang, Q.H.; Wang, C.; Si, W.M.; Wan, X.Q. Late Cretaceous paleoenvironment and paleoclimate change during large lake transgression in the Songliao Basin, NE China. Earth Sci. Front. 2009, 16, 123. [Google Scholar]
- Jing, X. Late Cretaceous Palynological Fossil Assemblages and Their Paleoclimatic Records from Eastern Songliao Basin. Master’s Thesis, China University of Geosciences, Beijing, China, 2011. [Google Scholar]
- Zhao, J. Pollen, Algae and Paleoclimate and Paleoclimate Conditions of Early to Middle Cretaceous in Songliao Basin. Master’s Thesis, China University of Geosciences, Beijing, China, 2013. [Google Scholar]
- Ye, Y.Q. Classification and Biostratigraphic Correlation of Nonmarine Ostracods from the Late Cretaceous Nenjiang Formation to the Sifangtai Formation in Southwestern Songliao Basin. Master’s Thesis, China University of Geosciences, Beijing, China, 2020. [Google Scholar]
- Xi, D.P.; Wan, X.Q.; Feng, Z.Q.; Li, S.; Feng, Z.H.; Jian, J.Z.; Jing, X.; Si, W.M. The discovery of Late Cretaceous foraminifera from Songliao Basin: Evidence from the lake–sea communication of the SK–1 well. Chin. Sci. Bull. 2010, 55, 3433–3436. [Google Scholar]
- Scott, R.W.; Wan, X.; Wang, C.; Huang, Q. Late Cretaceous chronostratigraphy (Turonian-Maastrichtian): SK1 core Songliao Basin, China. Geosci. Front. 2012, 3, 357–367. [Google Scholar] [CrossRef]
- Xi, D.P.; Li, S.; Wan, X.; Jing, X.; Huang, Q.; Colin, J.; Wang, Z.; Si, W. Late Cretaceous biostratigraphy and paleoenvironmental reconstruction based on non-marine ostracodes nonmarine ostracodes from well SK1 (south), Songliao Basin, northeast China. Hydrobiologia 2012, 688, 113–123. [Google Scholar] [CrossRef]
- Qu, H.; Xi, D.; Li, S.; Colin, J.P.; Huang, Q.; Wan, X. Late Cretaceous-Early Paleocene ostracod biostratigraphy of scientific drilling Sk1(N) in the Songliao Basin, northeast China. J. Paleontol. 2014, 88, 786–799. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wan, X.Q.; Xi, D.P.; Jing, X.; Li, W.; Huang, Q.H.; Zhang, J.Y. Late Cretaceouspalynology and paleoclimate change: Evidence from the SK1 (south) core, Songliao Basin, NE China. Sci. China (Earth Sci.) 2013, 57, 2985–2997. [Google Scholar] [CrossRef]
- Ye, D.Q.; Huang, Q.H.; Zhang, Y. Cretaceous Ostracod Biostratigraphy in Songliao Basin; Petroleum Industry Press: Beijing, China, 2002. [Google Scholar]
- Cheng, J.H.; He, C.C. Non-marine Cretaceous dinoflagellate biostratigraphy. Journal of stratigraphy. J. Stratigr. 2012, 36, 229–240. [Google Scholar]
- Deng, C.L.; He, H.Y.; Pan, Y.X.; Zhu, R.X. Chronology of the terrestrial Upper Cretaceous in the Songliao Basin, northeast Asia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 385, 44–54. [Google Scholar] [CrossRef]
- Wu, H.C.; Zhang, S.H.; Jiang, G.Q.; Hinnov, L.; Yang, T.; Li, H.; Wan, X.; Wang, C. Astrochronology of the Early Turonian-Early Campanian terrestrial succession in the Songliao Basin, northeastern China and its implication for long-period behavior of the Solar System. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 385, 55–70. [Google Scholar] [CrossRef]
- Wu, H.C.; Zhang, S.; Hinnov, L.A.; Jiang, G.; Yang, T.; Li, H.; Wan, X.; Wang, C. Cyclostratigraphy and orbital tuning of the terrestrial Upper Santonian-Lower Danian in Songliao Basin, northeastern China. Earth Planet. Sci. Lett. 2014, 407, 82–95. [Google Scholar] [CrossRef]
- Ma, X.J.; Wu, H.C.; Fang, Q.; Shi, M.; Zhang, S.; Yang, T.; Li, H.; Wang, C. A floating astronomical time scale for the early Late Cretaceous continental strata in the Songliao Basin, Northeastern China. Acta Geol. Sin. Engl. Ed. 2020, 94, 27–37. [Google Scholar] [CrossRef]
- Wang, T.T.; Wang, C.S.; Ramezani, J.; Wan, X.; Yu, Z.; Gao, Y.; He, H.; Wu, H. High-precision geochronology of the Early Cretaceous Yingcheng Formation and its stratigraphic implications for Songliao Basin, China. Geosci. Front. 2022, 13, 101386. [Google Scholar] [CrossRef]
- Wang, P.J.; Xie, X.A.; Mattern, F.; Ren, Y.G.; Zhu, D.F.; Sun, X.M. The Cretaceous Songliao Basin: Volcanogenic succession, sedimentary sequence and tectonic evolution, NE China. Acta Geol. Sin. (Engl.) 2011, 81, 1002–1011. [Google Scholar]
- Wang, Y.Q.; Sames, B.; Liao, H.Y.; Xi, D.; Pan, Y. Late Cretaceous ostracod fauna from the Shenjiatun section (Songliao Basin, Northeast China): Biostratigraphic and palaeoecological implications. Cret. Res. 2017, 78, 174–190. [Google Scholar] [CrossRef]
- Yan, J.J.; Xi, D.P.; Yu, T.; Wan, X.Q. Biostratigraphy and environmental changes of the lower Nenjiang Formation in Qingshankou area, Songliao Basin. J. Stratigr. 2007, 03, 296–302. [Google Scholar]
- Kohei, Y. Changes of Palynophytes During the Campanian to Maastricht (Late Cretaceous) in East Asia. Ph.D. Thesis, China University of Geosciences, Beijing, China, 2016. [Google Scholar]
- Yu, Z.Q.; He, H.Y.; Deng, C.L.; Xi, D.; Qin, Z.; Wan, X.; Wang, C.; Zhu, R. New geochronological constraints for the Upper Cretaceous Nenjiang Formation in the Songliao Basin, NE China. Cret. Res. 2019, 102, 160–169. [Google Scholar] [CrossRef]
- Cheng, Y.; Li, Y.; Wang, S.; Li, Y.; Ao, C.; Li, J.; Sun, L.; Li, H.; Zhang, T. Late Cretaceoust tectono–magmatic event in Songliao Basin, geochronology and geochemistry analysis, NE China: New Insights from mafic dyke geochronology and geochemistry analysis. Geol. J. 2018, 53, 2991–3008. [Google Scholar] [CrossRef]
- Xie, X.; Yun, X.L. Three Cretaceous biota in Songliao Basin. In Proceedings of the China Conference, Guilin, China, 1 January 2013. [Google Scholar]
- Kong, H.; Chen, C.R.; Dang, Y.M.; Yang, J.G.; Huang, Q.H.; Zhao, C.B. Review of three Cretaceous biota in Songliao Basin. J. Paleontol. 2006, 45, 416–424. [Google Scholar]
- Huang, Q.H.; Chen, C.R.; Wang, P.Z.; Han, M.X.; Li, X.J.; Wu, D.Q. Late Cretaceous biological evolution and paleolake anoxic event in Songliao Basin. Acta Micropalaeontol. Sin. 1998, 15, 417–425. [Google Scholar]
- Xi, D.P.; Wan, X.Q.; Feng, Z.Q.; Li, S.; Feng, Z.; Jia, J.; Jing, X.; Si, W. Discovery of Late Cretaceous foraminifera in the Songliao Basin: Evidence from SK-1 and implications for identifying seawater incursions. Chin. Sci. Bull. 2011, 56, 253–256. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.F.; Wang, P.J.; Wang, C.S.; Ren, Y.G.; Wang, G.D.; Liu, W.Z.; Cheng, R.H. South Hole location, core profile characteristics and distribution of special lithologic layers of Songke 1 well. Acta Geol. Sin. 2008, 5, 669–675. [Google Scholar]
- Feng, Z.Q.; Zhang, S.; Timothy, A.; Feng, Z.H.; Xie, X.N.; Zhao, B.; Fu, X.L.; Wang, C.S. Lacustrine turbidite channels and fans in the Mesozoic Songliao Basin, China. Basin Res. 2010, 22, 96–107. [Google Scholar] [CrossRef]
- Cheng, R.H.; Wang, G.D.; Wang, P.J.; Gao, Y. Sedimentary microfacies and evolution of sedimentary environment of Sifangtai Formation and Mingshui Formation in the north hole of Songke 1 Well. Earth Sci. Front. 2009, 16, 85–95. [Google Scholar]
- Wang, G.D.; Cheng, R.H.; Wang, P.J.; Gao, Y.F.; Wang, G.S.; Ren, Y.G.; Huang, Q.H. Fine description of the sedimentary sequence of the Upper Cretaceous Sifangtai Formation in the Songke 1 well in the Songliao Basin at centimeter scale: Lithology, lithofacies and cycle. Earth Sci. Front. 2011, 18, 263–284. [Google Scholar]
- Nichols, D.J.; Sweet, A.R. Biostratigraphy of Upper Cretaceous Nonmarine Palynofloras in a north-south Transect of the Western Interior Basin; Geological Association of Canada–Special: Waterloo, ON, Canada, 1993; Volume 39, pp. 539–584. [Google Scholar]
- Srivastava, S.K. Ephedralean pollen from the Upper Cretaceous Edmonton Formation of Alberta (Canada) and their paleoecological significance. Can. J. Earth Sci. 1968, 5, 211–221. [Google Scholar] [CrossRef]
- Barrón, E.; Peyrot, D.; Rodríguez-Lopez, J.P.; Meléndez, N.; López del Valle, R.L.; Najarro, M.; Rosales, I.; Comas-Rengifo, M.J. Palynology of Aptian and Upper Albian (Lower Cretaceous) amber-bearing outcrops of the southern margin of the Basque-Cantabrian basin (northern Spain). Cret. Res. 2015, 52, 292–312. [Google Scholar] [CrossRef]
- Nichols, D.J.; Jacobson, S.R. Cretaceous biostratigraphy in the Wyoming thrust belt. Mt Geol. 1982, 19, 73–78. [Google Scholar] [CrossRef]
- Crane, P.R.; Friis, E.M.; Pedersen, K.R. The origin and early diversification of angiosperms. Nature 1995, 374, 27–33. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Ji, L.M.; Du, B.X.; Dai, S.; Hou, X. Palynology of the Early Cretaceous Hanxia Section in the Jiuquan Basin, Northwest China: The discovery of diverse early angiosperm pollen and paleoclimatic significance. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 440, 297–306. [Google Scholar] [CrossRef]
- Wang, X. The dawn angiosperms: Uncovering the origin of flowering plants. Lect. Notes Earth Sci. 2010, 121, 1–4. [Google Scholar] [CrossRef]
- Sauquet, H.; von Balthazar, M.; Magallón, S.; Doyle, J.A.; Endress, P.K.; Bailes, E.J.; Barroso de Morais, E.; Bull-Hereñu, K.; Carrive, L.; Chartier, M.; et al. The ancestral flower of angiosperms and its early diversification. Nat. Commun. 2017, 8, 16047. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.; Liang, F.; Yang, T.; Zhang, S.Q. Late Cretaceous Flora from Jiayin of Heilongjiang; Shanghai Science and Technology Press Education Press: Shanghai, China, 2020. [Google Scholar]
- Sun, G.; Dong, Z.M.; Akhmetiev, M.; Markevich, V.; Godefroit, P.; Dlicher, D.L.; Sun, C.L.; Sun, Y.M.; Golovneva, L. Late Cretaceous-Paleocene Biota and the K–Pg Boundary from Jiayin of Heilongjiang, China; Shanghai Science, Technology and Education Publishing House: Shanghai, China, 2014; pp. 1–194. [Google Scholar]
- Duan, Z.H. Classopollis and its paleoclimatic significance. Coal Geol. Explor. 1991, 6, 14–21. [Google Scholar]
- Vakhrameev, V.A. Pollen Classopollis: Indicator of Jurassic and Cretaceous climate. Palaeobotanists 1981, 28, 301–307. [Google Scholar] [CrossRef]
- Abbink, O.; Targarona, J.; Brinkhuis, H.; Visscher, H. Late Jurassic to earliest Cretaceous palaeoclimatic evolution of the southern North Sea. Glob. Planet Chang. 2001, 30, 231–256. [Google Scholar] [CrossRef]
- Abbink, O.A.; van Konijnenburg-Van Cittert, J.H.A.; Visscher, H. A sporomorph ecogroup model for the northwest European Jurassic-Lower Cretaceous: Concepts and framework. Neth. J. Geosci. 2004, 83, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.H. Paleoclimatic indicative significance of Mesozoic main plant fossils. J. Palaeogeogr. 2007, 9, 559–574. [Google Scholar]
- Lin, M.Q. Late Jurassic to Early Cretaceous Palynological Flora in Northern China and Tibet and Its Paleoenvironmental Significance. Ph.D. Thesis, University of Science and Technology of China, Hefei, China, 2020. [Google Scholar]
- Stukins, S.; Jolley, D.W.; McIlroy, D.; Hartley, A.J. Middle Jurassic vegetation dynamics from allochthonous palynological assemblages: An example from a marginal marine depositional setting; Lajas Formation, Neuquen Basin, Argentina. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 392, 117–127. [Google Scholar] [CrossRef]
- Li, J.G.; Wu, Y.X.; Batten, D.J.; Lin, M. Vegetation and climate of the central and northern Qinghai-Xizang plateau from the Middle Jurassic to the end of the Paleogene inferred from palynology. J. Asian Earth Sci. 2019, 175, 35–48. [Google Scholar] [CrossRef]
- Li, J.G.; Peng, J.G.; Zhang, Q.Q. Cretaceous sporopollen assemblages in the Gangbaqaqila section, Tibet. J. Paleontol. 2016, 5, 346–366. [Google Scholar]
- Slater, S.M.; Twitchett, R.J.; Danise, S.; Vajda, V. Substantial vegetation response to Early Jurassic global warming with impacts on oceanic anoxia. Nat. Geosci. 2019, 12, 462–467. [Google Scholar] [CrossRef]
- Gao, R.Q.; Zhang, Y.; Cui, T.C. Cretaceous Petroleum Strata in Songliao Basin; Petroleum Industry Press: Beijing, China, 1994. [Google Scholar]
- Markevich, V.S.; Ashraf, A.R.; Bugdaeva, E.V. The Maastrichtian-Danian palynological assemblages from Wuyun of Jiayin nearby the Heilongjiang (Amur) River. In Ancient Life and Modern Approaches; The 2nd International Congress of Paleontology; IPC: Beijing, China, 2006; pp. 526–527. [Google Scholar]
- Steinig, S.; Dummann, W.; Park, W.; Latif, M.; Kusch, S.; Hofmann, P.; Flögel, S. Evidence for a regional warm bias in the Early Cretaceous TEX86 record. Earth Planet. Sci. Lett. 2020, 539, 116184. [Google Scholar] [CrossRef]
- Ji, L.M.; Zhang, M.Z.; Song, Z.G. The palynological record from Coniacian to Lower Campanian continental sequences in the Songliao Basin, northeastern China and its implications for palaeoclimate. Cret. Res. 2015, 56, 226–236. [Google Scholar] [CrossRef]
- Wan, X.Q.; Wu, H.C.; Xi, D.P.; Liu, M.Y.; Qin, Z.H. Terrestrial biota and climate environment evolution during the Cretaceous Greenhouse Period in Northeast China. Earth Sci. Front. 2017, 24, 18–31. [Google Scholar] [CrossRef]
- Keller, G. Cretaceous climate, volcanism, impacts, and biotic effects. Cret. Res. 2008, 29, 754–771. [Google Scholar] [CrossRef]
- Coiffard, C.; Gomez, B. The rise to dominance of the angiosperm kingdom: Dispersal, habitat widening and evolution during the Late Cretaceous of Europe. Lethaia 2010, 43, 164–169. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Osmundacidites | O. wellmanii | Cibotiumspora | Todisporites | Pterisisporites | Hymenophyllumsporites | Salviniaspora | Schizaeoisporites | S. digitatoides | S. laevigataeformis | S. cretaceous | S. praeclarus | S. evidens | S. kulandyensis | S. sunjiawanensis | S. certus | S. applanatus | S. regularis | S. palaeocenicus | S. minor | Trilobosporites | Concavissmisporites | Lygodiumsporites | Lygodioisporites | Cicatricosisporites | C. minor | Densoisporites | Pteridium | Cyathidites | Cyathidites minor | Deltoidospora | Klukisporites | Toroisporis | Triporoletes | T. laevigatus | Foraminisporis | Biretisporites | Polycingulatisporites | Gabonisporites | Punctatisporites | Laevigatosporites | Leiotriletes | Fern spores | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 240 | K2s | 1 | 1 | 2 | 10 | 8 | 10 | 1 | 2 | 4 | 1 | 4 | 2 | 8 | 2 | 10 | 4 | 2 | 3 | 1 | 1 | 3 | 80 | |||||||||||||||||||||
| 246 | K2s | 4 | 7 | 9 | 1 | 2 | 10 | 6 | 39 | |||||||||||||||||||||||||||||||||||
| 249 | K2s | 8 | 153 | 30 | 191 | |||||||||||||||||||||||||||||||||||||||
| 266 | K2s | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||
| 268 | K2s | 46 | 24 | 12 | 2 | 84 | ||||||||||||||||||||||||||||||||||||||
| 276 | K2s | 1 | 2 | 3 | 6 | |||||||||||||||||||||||||||||||||||||||
| 279 | K2s | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||
| 285 | K2s | 6 | 1 | 3 | 16 | 5 | 1 | 32 | ||||||||||||||||||||||||||||||||||||
| 296 | K2s | 6 | 18 | 5 | 7 | 1 | 2 | 39 | ||||||||||||||||||||||||||||||||||||
| 299 | K2s | 1 | 1 | 1 | 7 | 3 | 1 | 1 | 1 | 2 | 1 | 30 | 10 | 4 | 1 | 16 | 2 | 4 | 86 | |||||||||||||||||||||||||
| 315 | K2s | 12 | 3 | 2 | 2 | 2 | 1 | 7 | 2 | 2 | 4 | 37 | ||||||||||||||||||||||||||||||||
| 319 | K2s | 2 | 6 | 1 | 1 | 1 | 3 | 1 | 1 | 1 | 2 | 3 | 4 | 26 | ||||||||||||||||||||||||||||||
| 330 | K2s | 2 | 1 | 3 | ||||||||||||||||||||||||||||||||||||||||
| 338 | K2s | 1 | 2 | 29 | 5 | 1 | 1 | 5 | 1 | 6 | 1 | 1 | 1 | 5 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 67 | ||||||||||||||||||||||
| 351 | K2n2 | 1 | 1 | 2 | ||||||||||||||||||||||||||||||||||||||||
| 354 | K2n2 | 5 | 2 | 1 | 1 | 1 | 1 | 11 | ||||||||||||||||||||||||||||||||||||
| 359 | K2n2 | 8 | 1 | 2 | 1 | 12 | ||||||||||||||||||||||||||||||||||||||
| 365 | K2n2 | 19 | 19 | |||||||||||||||||||||||||||||||||||||||||
| 371 | K2n2 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 441 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 444 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 446 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 451 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 462 | K2n1 | 6 | 10 | 10 | 3 | 1 | 1 | 2 | 1 | 34 | ||||||||||||||||||||||||||||||||||
| 474 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| Podocarpidites | P. minisculus | Pinaceae | Pinuspollenites | Abietineaepollenites | Piceaepollenites | P. tobolicus | P. sp. | Cedripites | Laricoidites | Rugubivesiculites | Araucariacites | Cycadopites | Ephedripites | E. fusiformis | Regalipollenites | Jugella | Inaperturopollenites | Chasmatosporites | Exesipollenites | E. tumulus | Rotundipollis | Psophosphaera | Concentrisporites | Callialasporites | Jiaohepollis | Parvisaccites | Parcisporites | Cerebropollenites | Classopollis | C. annulatus | C. classoides | Pseudopicea | Piceites | Paleoconiferus | Gymnosperm pollen | |||||||||
| 240 | K2s | 4 | 8 | 2 | 4 | 5 | 2 | 2 | 1 | 4 | 9 | 19 | 1 | 61 | ||||||||||||||||||||||||||||||
| 246 | K2s | 2 | 2 | 1 | 11 | 1 | 4 | 2 | 12 | 1 | 2 | 38 | ||||||||||||||||||||||||||||||||
| 249 | K2s | 5 | 1 | 6 | ||||||||||||||||||||||||||||||||||||||||
| 266 | K2s | 2 | 1 | 3 | ||||||||||||||||||||||||||||||||||||||||
| 268 | K2s | 2 | 4 | 1 | 2 | 1 | 1 | 11 | ||||||||||||||||||||||||||||||||||||
| 276 | K2s | 1 | 9 | 2 | 12 | |||||||||||||||||||||||||||||||||||||||
| 279 | K2s | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||
| 285 | K2s | 11 | 4 | 11 | 1 | 2 | 6 | 4 | 4 | 43 | ||||||||||||||||||||||||||||||||||
| 296 | K2s | 2 | 12 | 2 | 2 | 18 | ||||||||||||||||||||||||||||||||||||||
| 299 | K2s | 1 | 2 | 2 | 1 | 3 | 1 | 2 | 7 | 1 | 1 | 5 | 26 | |||||||||||||||||||||||||||||||
| 315 | K2s | 1 | 3 | 2 | 8 | |||||||||||||||||||||||||||||||||||||||
| 319 | K2s | 1 | 2 | 9 | 29 | 31 | 3 | 8 | 1 | 2 | 8 | 2 | 1 | 1 | 1 | 4 | 1 | 1 | 25 | 1 | 1 | 1 | 2 | 135 | ||||||||||||||||||||
| 330 | K2s | 4 | 8 | 2 | 3 | 2 | 2 | 1 | 2 | 2 | 39 | 1 | 2 | 1 | 69 | |||||||||||||||||||||||||||||
| 338 | K2s | 2 | 6 | 2 | 3 | 1 | 1 | 2 | 1 | 2 | 1 | 14 | 1 | 1 | 1 | 6 | 1 | 1 | 46 | |||||||||||||||||||||||||
| 351 | K2n2 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||
| 354 | K2n2 | 1 | 3 | 6 | 3 | 2 | 1 | 1 | 2 | 3 | 7 | 57 | ||||||||||||||||||||||||||||||||
| 359 | K2n2 | 2 | 4 | 1 | 1 | 5 | 20 | 6 | 1 | 1 | 41 | |||||||||||||||||||||||||||||||||
| 365 | K2n2 | 21 | 33 | 2 | 12 | 68 | ||||||||||||||||||||||||||||||||||||||
| 371 | K2n2 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 441 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 444 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 446 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 451 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 462 | K2n1 | 2 | 28 | 48 | 20 | 20 | 2 | 4 | 11 | 1 | 40 | 8 | 8 | 2 | 14 | 2 | 6 | 18 | 3 | 4 | 271 | |||||||||||||||||||||||
| 474 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| Cupuliferoipollenites | Cupuliferoidaepollenites | Ulmoideipites | Ulmipollenites | Liquidambarpollenites | Celtispollenites | Magnolipollis | Liliacidites | Margocolporites | Potamogetonacidites | Granwellia | Callistopollenites | Asteropollis | Clavatipollenites | Aquilapollenites | A. minor | A. spinulosus | Lythraites | Integricorpus | Xuippllis | Brenneripollis | Polyporites | Sabalpollenites | Triphyllopollis trigonos | Retitrescolpites | Retitricolpites | Tricolpopollenites | T. mollis | Retitricolporites | Angiosperm pollen | |||||||||||||||
| 240 | K2s | 1 | 3 | 4 | 2 | 4 | 4 | 4 | 4 | 6 | 6 | 2 | 3 | 3 | 4 | 12 | 6 | 5 | 77 | |||||||||||||||||||||||||
| 246 | K2s | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||
| 249 | K2s | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||
| 266 | K2s | 4 | ||||||||||||||||||||||||||||||||||||||||||
| 268 | K2s | 1 | 1 | 7 | 9 | |||||||||||||||||||||||||||||||||||||||
| 276 | K2s | 5 | 5 | |||||||||||||||||||||||||||||||||||||||||
| 279 | K2s | |||||||||||||||||||||||||||||||||||||||||||
| 285 | K2s | 1 | 1 | 1 | 3 | 2 | 8 | |||||||||||||||||||||||||||||||||||||
| 296 | K2s | 2 | 1 | 3 | ||||||||||||||||||||||||||||||||||||||||
| 299 | K2s | 2 | 2 | 2 | 6 | |||||||||||||||||||||||||||||||||||||||
| 315 | K2s | 3 | 5 | 2 | 1 | 6 | 2 | 4 | 10 | 6 | 39 | |||||||||||||||||||||||||||||||||
| 319 | K2s | 2 | 11 | 1 | 1 | 3 | 2 | 2 | 2 | 1 | 5 | 30 | ||||||||||||||||||||||||||||||||
| 330 | K2s | 1 | 1 | 2 | 4 | |||||||||||||||||||||||||||||||||||||||
| 338 | K2s | 1 | 1 | 7 | 1 | 4 | 1 | 1 | 23 | 1 | 2 | 1 | 1 | 2 | 2 | 3 | 2 | 4 | 5 | 4 | 15 | 83 | ||||||||||||||||||||||
| 351 | K2n2 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 354 | K2n2 | 1 | 1 | 2 | 2 | 6 | ||||||||||||||||||||||||||||||||||||||
| 359 | K2n2 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 365 | K2n2 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 371 | K2n2 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 441 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 444 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 446 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 451 | K2n1 | 0 | ||||||||||||||||||||||||||||||||||||||||||
| 462 | K2n1 | 2 | 2 | 6 | 11 | |||||||||||||||||||||||||||||||||||||||
| 474 | K2n1 | 0 |
| Vegetation Types | ||||
|---|---|---|---|---|
| Coniferous Forest | Evergreen Broadfeaf Forest | Deciduous Broadleaf Forest | Shrub | Herb |
| Classopollis | Cyathidites | Ulmoideipites | Concavissimisporites | Laevigatosporites |
| Podocarpidites | Cibotiumspora | Liquidambarpollenites | Klukisporites | Schizaeoisporites |
| Cedripites | Cycadopites | Ulmipollenites | Lygodiumsporites | Deltoidospora |
| Piceaepollenites | Magnolipollis | Lygodioisporites | Osmundacidites | |
| Abietspollenites | Aquilapollenites | Todisporites | ||
| Pinuspollenites | Integricorpus | Densoisporites | ||
| Araucariacites | Polyporites | |||
| Parvisaccites | Lythraites | |||
| Palaeoconiferus | Cicatricosisporites | |||
| Piceites | Ephedripites | |||
| Inaperturopollenites | ||||
| Psophosphaera | ||||
| Palynomorphs | Climate type | References |
|---|---|---|
| Classopollis | Hot Type | [60,64,65,66,67,68,69,70,71,72,73,74] |
| Foraminisporis | Warm Type | |
| Osmundacidites | Warm Type | |
| Callialasporites | Warm Type | |
| Inaperturopollenites | Warm Type | |
| Concentrisporites | Cool Type | |
| Exesipollenites | Cool Type | |
| Pinaceae | Cool Type | |
| Abietineaepollenites | Cool Type | |
| Pinuspollenites | Cool Type | |
| Cedripites | Cool Type | |
| Parvisaccites | Cool Type | |
| Piceaepollenites | Cool Type | |
| Piceites | Cool Type |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Z.; Xi, D.; Sun, L.; Zhao, J.; Yang, W.; Ye, Y.; Meng, X.; Wan, X. Late Cretaceous (Santonian to Campanian) Palynological Records and Paleoclimatic Significance from Borehole ZKY2-1, Songliao Basin. Minerals 2023, 13, 338. https://doi.org/10.3390/min13030338
Zhou Z, Xi D, Sun L, Zhao J, Yang W, Ye Y, Meng X, Wan X. Late Cretaceous (Santonian to Campanian) Palynological Records and Paleoclimatic Significance from Borehole ZKY2-1, Songliao Basin. Minerals. 2023; 13(3):338. https://doi.org/10.3390/min13030338
Chicago/Turabian StyleZhou, Zihan, Dangpeng Xi, Lixin Sun, Jing Zhao, Wanshu Yang, Yunqi Ye, Xinyu Meng, and Xiaoqiao Wan. 2023. "Late Cretaceous (Santonian to Campanian) Palynological Records and Paleoclimatic Significance from Borehole ZKY2-1, Songliao Basin" Minerals 13, no. 3: 338. https://doi.org/10.3390/min13030338