
Phosphate Record in Pleistocene-Holocene Sediments from Denisova Cave: Formation Mechanisms and Archaeological Implications

 ,
,
Abstract
:1. Introduction
- i.
- Reconstructing the paleochemistry of archaeological sediments in order to estimate the preservation of remains in a given environment after deposition and during diagenesis;
- ii.
- Detecting secondary indicators that appeared and/or survived during diagenesis being stable in the altered environment and justifying their suitability for interpretation of past conditions and events.
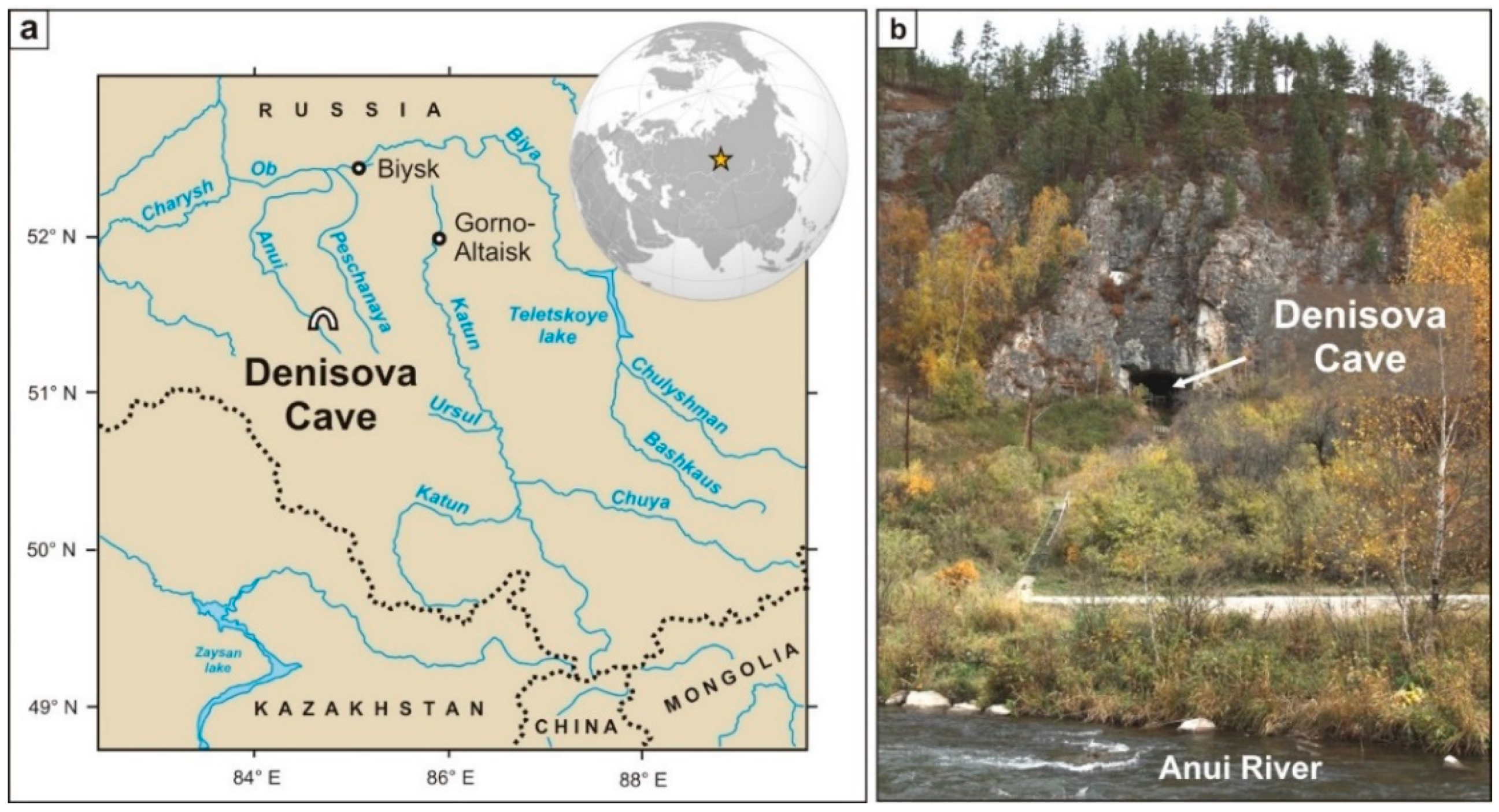
2. Denisova Cave
2.1. Historic Background
2.2. Geological and Archaeological Background
3. Authigenic Phosphates in Cave Sediments
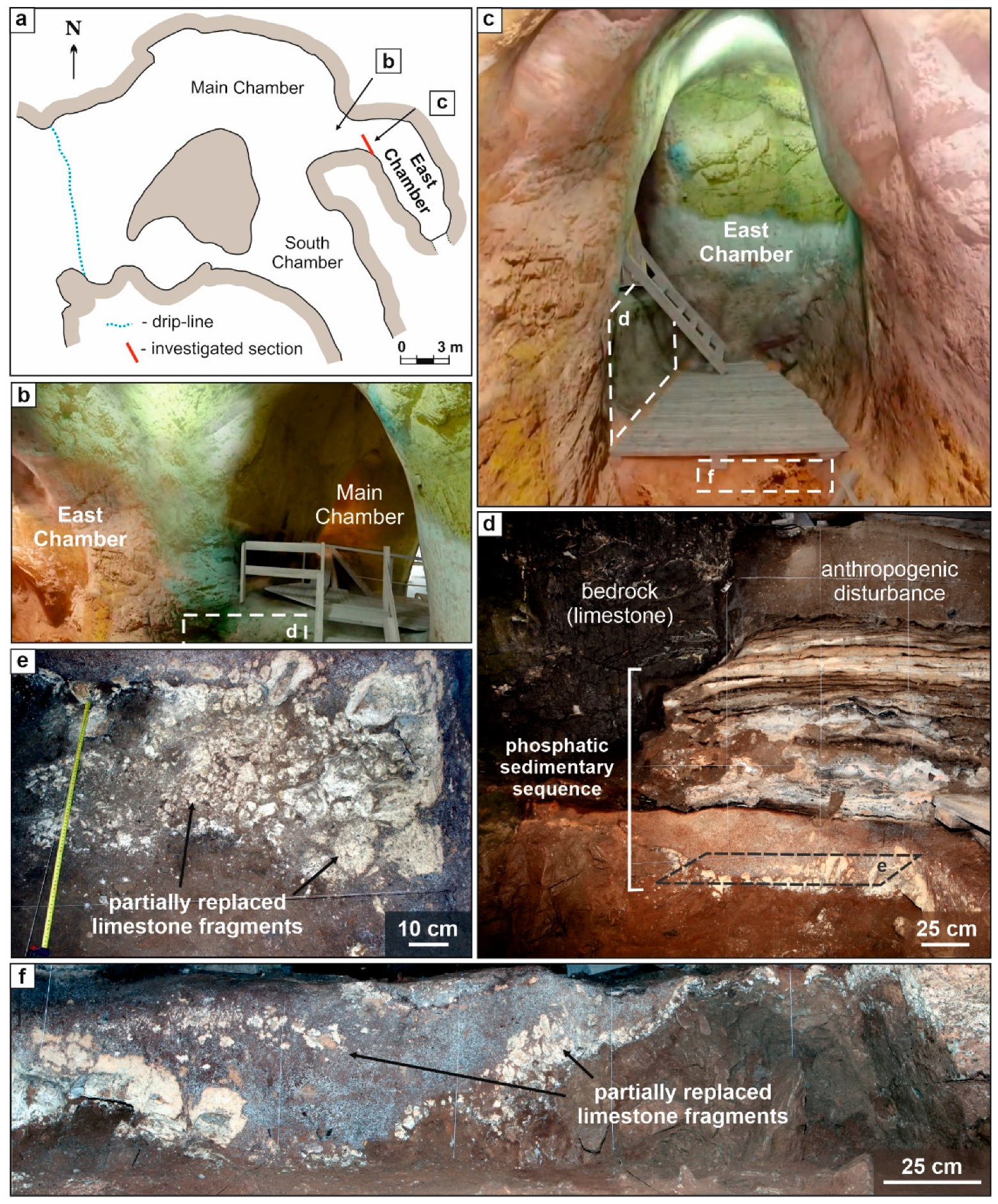
4. Site Description
Sedimentary Sequence of East Chamber
5. Materials and Methods
5.1. Sampling
5.2. Analytical Procedures
6. Results
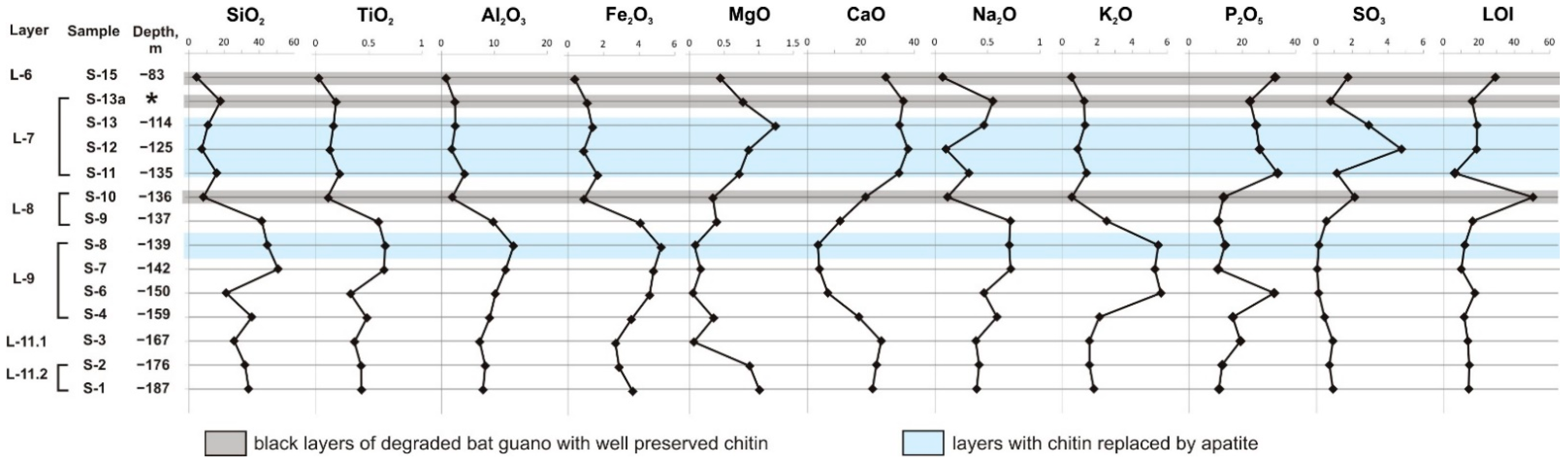
6.1. Bulk Rock Chemistry
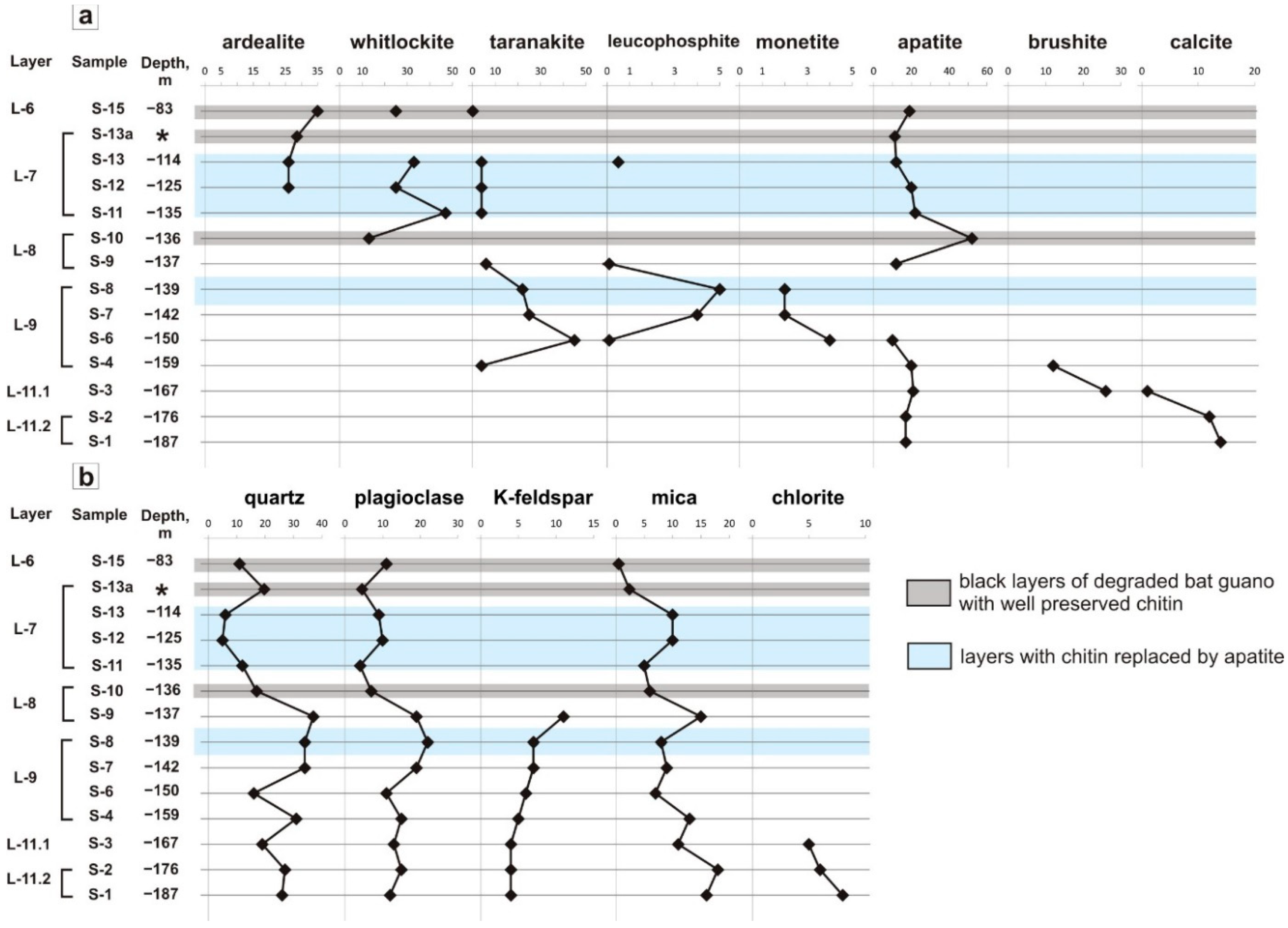
6.2. Depth Profiles of Phosphate Minerals in the East Chamber
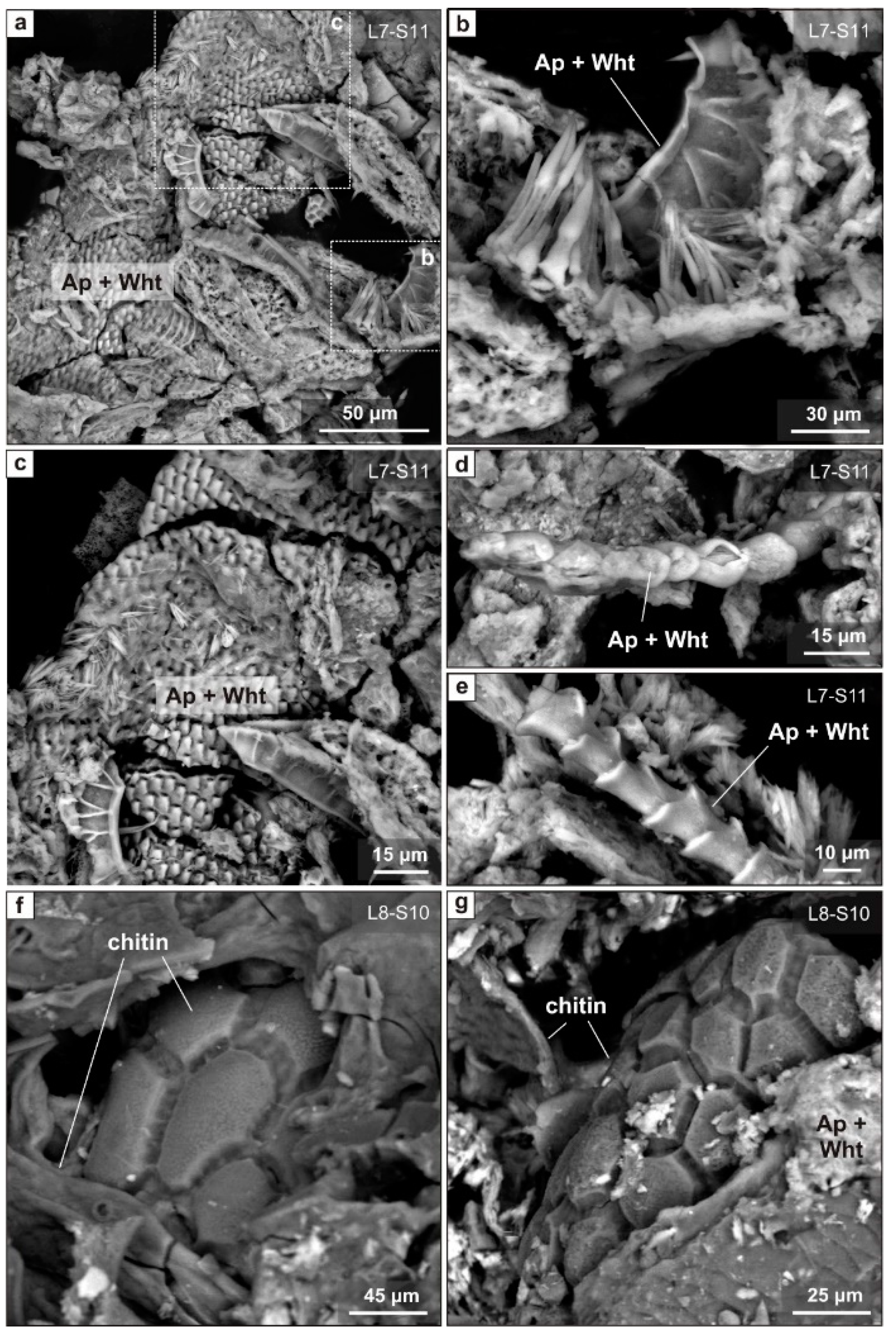
6.3. Distribution of Phosphate Assemblages by Layer
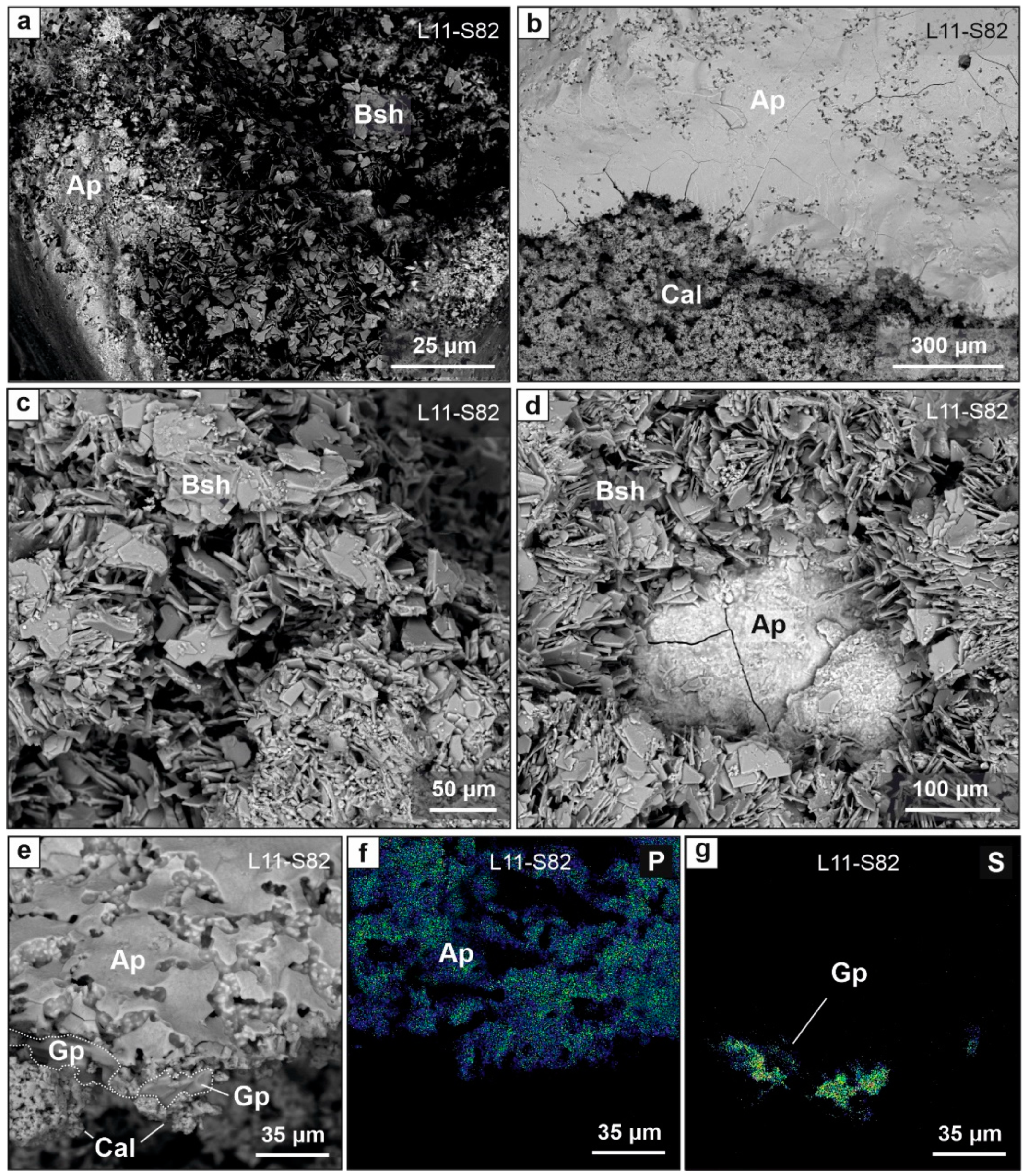
6.4. Effect of Phosphate Leaching on Stone Tools
7. Discussion
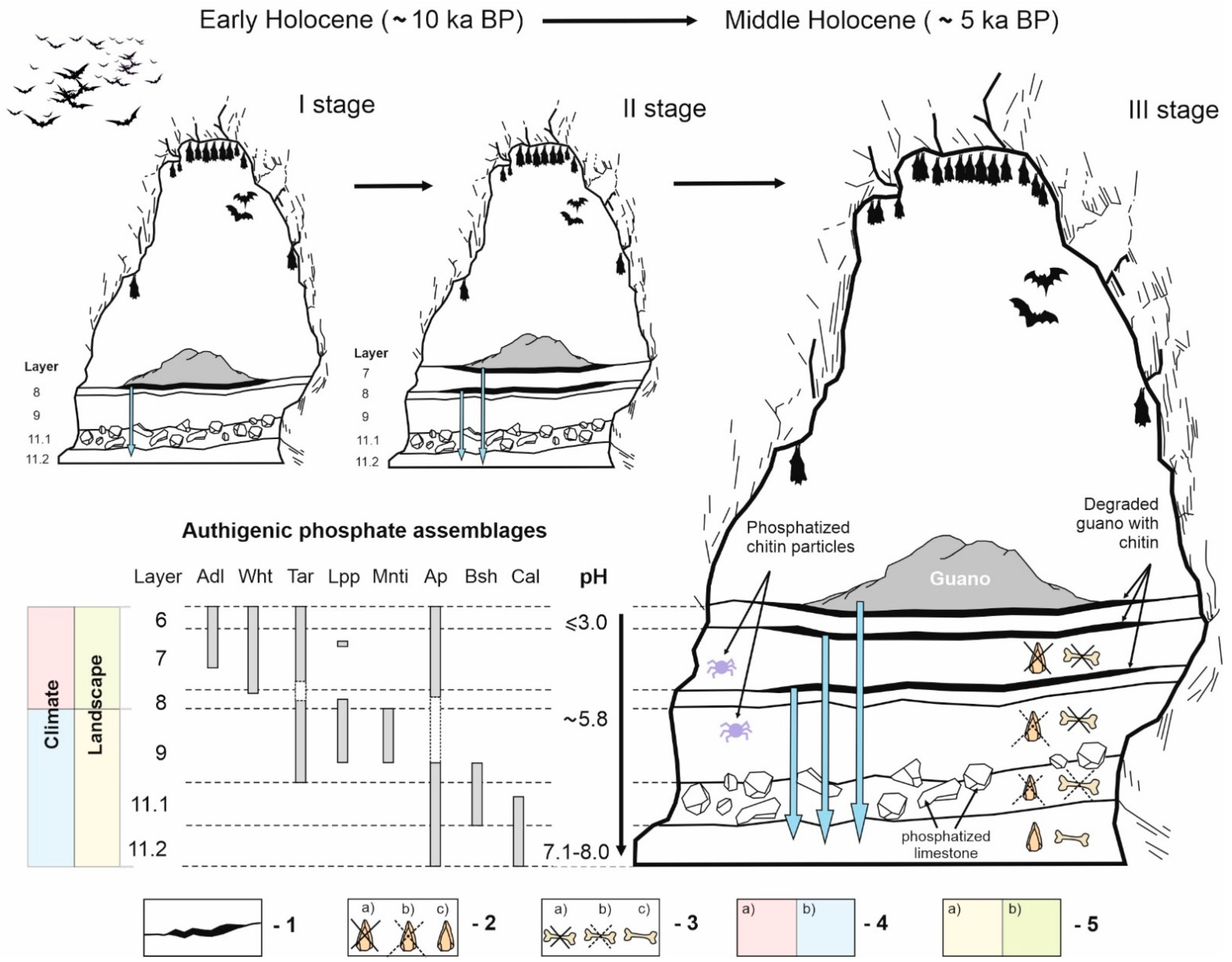
7.1. Authigenic Phosphates in Sedimentary Sequence of East Chamber: Diversity and Formation Conditions
7.2. Preservation of Archaelogical and Paleontological Findings in the East Chamber: Biogenic Processes and Acidic Leaching
7.3. Dissolved Bones: Source of Phosphorus in Cave Sediments
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shahack-Gross, R.; Berna, F.; Karkanas, P.; Weiner, S. Bat guano and preservation of archaeological remains in cave sites. J. Archeol. Sci. 2004, 31, 1259–1272. [Google Scholar] [CrossRef]
- Karkanas, P. Preservation of anthropogenic materials under different geochemical processes: A mineralogical approach. Quat. Int. 2010, 214, 63–69. [Google Scholar] [CrossRef]
- Wurster, C.M.; Munksgaard, N.; Zwart, C.; Bird, M. The biogeochemistry of insectivorous cave guano: A case study from insular Southeast Asia. Biogeochemistry 2015, 124, 163–175. [Google Scholar] [CrossRef]
- Shunkov, M.V.; Kulik, N.A.; Kozlikin, M.B.; Sokol, E.V.; Miroshnichenko, L.V.; Ulianov, V.A. The phosphates of Pleistocene-Holocene sediments of the Eastern Gallery of Denisova Cave. Doklady Earth Sci. 2018, 478, 46–50. [Google Scholar] [CrossRef]
- Birkeland, P.W. Soils and Geomorphology; Oxford Univ. Press: New York, NY, USA, 1999; 430p. [Google Scholar]
- Bohn, H.L.; Myer, R.A.; O’Connor, G.A. Soil Chemistry; John Wiley & Sons: Oxford, UK, 2002. [Google Scholar]
- Retallack, G.J. Soils of The Past: An Introduction to Paleopedology; Blackwell: Oxford, UK, 2001; 600p. [Google Scholar]
- Canti, M.; Huisman, D.J. Scientific advances in geoarchaeology during the last twenty years. J. Archeol. Sci. 2015, 56, 96–108. [Google Scholar] [CrossRef] [Green Version]
- Derevianko, A.P.; Shunkov, M.V.; Agadjanian, A.K.; Baryshnikov, G.F.; Malaeva, E.M.; Ulianov, V.A.; Kulik, N.A.; Postnov, A.V.; Anoikin, A.A. Paleoenvironment and Paleolithic Human Occupation of Gorny Altai; Derevianko, A.P., Shunkov, M.V., Eds.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2003; 448p. (In Russian) [Google Scholar]
- Derevianko, A.P.; Shunkov, M.V.; Kozlikin, M.B. Who Were the Denisovans? Archeol. Ethnol. Anthropol. Eurasia 2020, 48, 3–32. [Google Scholar] [CrossRef]
- Karkanas, P.; Goldberg, P. Site formation processes at Pinnacle point Cave 13B (Mossel Bay, Western Cape Province, South Africa): Resolving stratigraphic and depositional complexities with micromorphology. J. Hum. Evol. 2010, 59, 256–273. [Google Scholar] [CrossRef]
- Henderson, P.; Marlow, C.A.; Molleson, T.; Williams, C.T. Patterns of chemical change during bone fossilization. Nature 1983, 306, 358–360. [Google Scholar] [CrossRef]
- Kohn, M.J.; Cerling, T.E. Stable isotope compositions of biological apatite. In Phosphates-Geochemical, Geobiological, and Materials Importance; Kohn, M.J., Rakovan, J., Hughes, J.M., Eds.; Mineralogical Society of America and Geochemical Society: Washington, DC, USA, 2002; pp. 455–488. [Google Scholar]
- Kohn, M.J. Models of diffusion-limited uptake of trace elements in fossils and rates of fossilization. Geochim. Cosmochim. Acta 2008, 72, 3758–3770. [Google Scholar] [CrossRef]
- Trueman, C.N.; Tuross, N. Trace elements in modern and ancient bone. In Phosphates-Geochemical, Geobiological, and Materials Importance; Kohn, M.J., Rakovan, J., Hughes, J.M., Eds.; Mineralogical Society of America and Geochemical Society: Washington, DC, USA, 2002; pp. 489–522. [Google Scholar]
- Cid, A.S.; Anjos, R.M.; Zamboni, C.B.; Cardoso, R.; Muniz, M.; Corona, A.; Valladares, D.L.; Kovacs, L.; Macario, K.; Perea, D.; et al. Na, K, Ca, Mg, and U-series in fossil bone and the proposal of a radial diffusion-adsorption model of uranium uptake. J. Env. Radioact. 2014, 136, 131–139. [Google Scholar] [CrossRef]
- Keely, H.C.M.; Hudson, G.E.; Evans, J. Trace element contents of human bones in various states of preservation. 1. The soil silhouette. J. Archeol. Sci. 1977, 4, 19–24. [Google Scholar] [CrossRef]
- Hedges, R.E.; Millard, A.R. Bones and groundwater: Towards the modelling of diagenetic processes. J. Archeol. Sci. 1995, 22, 155–164. [Google Scholar] [CrossRef]
- Berna, F.; Matthews, A.; Weiner, S. Solubilities of bone mineral from archaeological sites: The recrystallization window. J. Archeol. Sci. 2004, 31, 867–882. [Google Scholar] [CrossRef]
- Karkanas, P.; Bar-Yosef, O.; Goldberg, P.; Weiner, S. Diagenesis in prehistoric caves: The use of minerals that form in situ to assess the completeness of the archaeological record. J. Archeol. Sci. 2000, 27, 915–929. [Google Scholar] [CrossRef]
- Weiner, S.; Goldberg, P.; Bar-Yosef, O. Bone preservation in Kebara Cave, Israel using on-site Fourier transform infrared spectrometry. J. Archeol. Sci. 1993, 20, 613–627. [Google Scholar] [CrossRef]
- Onac, B.P.; Breban, R.; Kearns, J.; Tamas, T. Unusual minerals related to phosphate deposits in Cioclovina Cave, Sureanu Mts. (Romania). Theor. Appl. Karstol. 2002, 15, 27–34. [Google Scholar]
- Onac, B.P.; Forti, P. Minerogenetic mechanisms occurring in the cave environment: An overview. Int. J. Speleol. 2011, 40, 79–98. [Google Scholar] [CrossRef] [Green Version]
- Audra, P.; De Waele, J.; Bentaleb, I.; Chroňáková, A.; Krištůfek, V.; D’Angeli, I.M.; Carbone, C.; Madonia, G.; Vattano, M.; Scopelliti, G.; et al. Guano-related phosphaterich minerals in European caves. Int. J. Speleol. 2019, 48, 75–105. [Google Scholar] [CrossRef] [Green Version]
- Krause, J.; Fu, Q.; Good, J.M.; Viola, B.; Shunkov, M.V.; Derevianko, A.P.; Pääbo, S. The Complete Mitochondrial DNA Genome of an Unknown Hominin from Southern Siberia. Nature 2010, 464, 894–897. [Google Scholar] [CrossRef]
- Reich, D.; Green, R.E.; Kircher, M.; Krause, J.; Patterson, N.; Durand, E.Y.; Viola, B.; Briggs, A.W.; Stenzel, U.; Johnson, P.L.F.; et al. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 2010, 468, 1053–1060. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.; Kircher, M.; Gansauge, M.-T.; Li, H.; Racimo, F.; Mallick, S.; Schraiber, J.G.; Jay, F.; Prüfer, K.; Filippo, C.d.; et al. A High-Coverage Genome Sequence from an Archaic Denisovan Individual. Science 2012, 338, 222–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slon, V.; Mafessoni, F.; Vernot, B.; de Filippo, C.; Grote, S.; Viola, B.; Hajdinjak, M.; Peyrégne, S.; Nagel, S.; Brown, S.; et al. The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature 2018, 561, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Okladnikov, A.P.; Ovodov, N.D. Denisova Cave paleolithic site in Altai Mountains. In Archaeological Discoveries 1977; Rybakov, B.A., Ed.; Nauka: Moscow, Russia, 1978; pp. 266–268. (In Russian) [Google Scholar]
- Derevianko, A.P.; Molodin, V.I. Denisova Cave; Nauka: Novosibirsk, Russia, 1994; 262p. (In Russian) [Google Scholar]
- Derevianko, A.P.; Shunkov, M.V.; Anoikin, A.A.; Ulyanov, A.V. The upper paleolithic of Denisova Cave: New results. In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Molodin, V.I., Eds.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2000; Volume 6, pp. 93–98. (In Russian) [Google Scholar]
- Jacobs, Z.; Li, B.; Shunkov, M.V.; Kozlikin, M.B.; Bolikhovskaya, N.S.; Agadjanian, A.K.; Uliyanov, V.A.; Vasiliev, S.K.; O’Gorman, K.; Derevianko, A.P.; et al. Timing of archaic hominin occupation of Denisova Cave in southern Siberia. Nature 2019, 565, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Filippov, A.G. Denisova Cave. In Atlas of the Caves of Russia; Shelepin, A.L., Ed.; Russian Geographical Society, Russian Union of Speleologist: Moscow, Russia, 2019; pp. 485–487. (In Russian) [Google Scholar]
- Vistingauzen, V.K. Speleological zoning of Altai. Bull. AB RGS 2019, 1, 17–27. (In Russian) [Google Scholar] [CrossRef]
- Marinin, A.M. Karst and Caves of Altai; Novosibirsk State Pedagogical University: Novosibirsk, Russia, 1990; 148p. (In Russian) [Google Scholar]
- Bolikhovskaya, N.S.; Kozlikin, M.B.; Shunkov, M.V.; Ulianov, V.A.; Faustov, S.S. New data in palynology of unique Paleolithic site Denisova Cave in the North-Western Altai. Bull. Moscow Soc. Nat. Biol. Ser. 2017, 122, 46–60. (In Russian) [Google Scholar]
- Shunkov, M.V.; Agadzhanyan, A.K. Microclimate of Denisova Cave. In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Molodin, V.I., Eds.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2007; Volume 13, pp. 162–166. (In Russian) [Google Scholar]
- Vasenkov, D.A. Chiroptera Mammalia in highliands of southwestern Siberia. Ph.D. Thesis, Institute of Systematics and Ecology of Animals, Novosibirsk, Russia, 2009. (In Russian). [Google Scholar]
- Zhigalin, A.V. Mammalia, Chiroptera, Vespertilionidae of the Altai-Sayan Mountain Province and its Surroundings: Fauna and Ecology. Ph.D. Thesis, National Research Tomsk State University, Tomsk, Russia, 2018. (In Russian). [Google Scholar]
- Zhigalin, A.V.; Gadzhiev, A.A.; Daudova, M.G.; Salimkhanov, N.G.; Shestopalov, A.M. Ecology of chiroptera bats in Altai-Sayan region of Southern Siberia. S. Russ. Ecol. Develop. 2019, 14, 9–25. [Google Scholar] [CrossRef] [Green Version]
- Leonov, A.V.; Bobkov, A.E.; Baturin, Y.M.; Anikushkin, M.N.; Rys, I.V.; Kozlikin, M.B.; Shunkov, M.V.; Derevianko, A.P. Development of a virtual 3d model of Denisova Cave in the Altai mountains. Archeol. Ethnol. Anthropol. Eurasia 2014, 3, 14–20. [Google Scholar] [CrossRef]
- Wilson, M.J.; Bain, D.C. Occurrence of leucophosphite in a soil from Elephant Island, British Antarctic Territory. Am. Mineral. 1976, 61, 1027–1028. [Google Scholar]
- Simas, F.N.B.; Schaefer, C.E.G.R.; Melo, V.F.; Albuquerque-Filho, M.R.; Michel, R.F.M.; Pereira, V.V.; Gomes, M.R.M.; da Costa, L.M. Ornithogenic cryosols from maritime Antarctica: Phosphatization as a soil forming process. Geoderma 2007, 138, 191–203. [Google Scholar] [CrossRef]
- McFarlane, D.A.; Keeler, R.C.; Mizutani, H. Ammonia volatilization in a Mexican bat cave ecosystem. Biogeochem 1995, 30, 1–8. [Google Scholar] [CrossRef]
- Karkanas, P.; Rigaud, J.P.; Simek, J.F.; Albert, R.M.; Weiner, S. Ash bones and guano: A study of the minerals and phytoliths in the sediments of Grotte XVI, Dordogne, France. J. Archeol. Sci. 2002, 29, 721–732. [Google Scholar] [CrossRef]
- Fiore, S.; Laviano, R. Brushite, hydroxylapatite, and taranakite from Apulian caves (southern Italy): New mineralogical data. Am. Mineral. 1991, 76, 1722–1727. [Google Scholar]
- Rellini, I.; Firpo, M.; Martino, G.; Riel-Salvatore, J.; Maggi, R. Climate and environmental changes recognized by micromorphology in Paleolithic deposits at Arene Candide (Liguria, Italy). Quat. Int. 2013, 315, 42–55. [Google Scholar] [CrossRef]
- Forbes, M.S.; Bestland, E.A. Guano-derived deposits within the sandy cave fills of Naracoorte, South Australia. Alcheringa: Australas. J. Palaeontol. 2006, 30, 129–146. [Google Scholar] [CrossRef]
- Onac, B.P.; Vereş, D.S. Sequence of secondary phosphates deposition in a karst environment: Evidence from Magurici Cave (Romania). Europ. J. Mineral. 2003, 15, 741–745. [Google Scholar] [CrossRef]
- Dumitraş, D.G.; Marincea, Ş.; Bilal, E.; Hatert, F. Apatite-(CaOH) in the fossil bat guano deposit from the “dry” Cioclovina Cave, Sureanu Mountains, Romania. Can. Mineral. 2008, 46, 431–445. [Google Scholar] [CrossRef]
- Marincea, Ş.; Dumitraş, D.G.; Diaconu, G.; Bilal, E. Hydroxylapatite, brushite and ardealite in the bat guano deposit from Peştera Mare de la Mereşti, Perşani Mountains, Romania. Neues Jahrb. für Mineral. Abhandlungen 2004, 10, 464–488. [Google Scholar] [CrossRef]
- Dumitraş, D.G.; Marincea, Ş. Sequential dehydration of the phosphate-sulfate association from Gura Dobrogei Cave, Dobrogea, Romania. Europ. J. Mineral. 2021, 33, 329–340. [Google Scholar] [CrossRef]
- Tămaş, T.; Miheţ, O.; Giurgiu, A. Mineralogy of bat guano deposits from Huda lui Papară cave (Trascău Mountains, Romania). Carpath. J. Earth Env. Sci. 2014, 9, 25–32. [Google Scholar]
- Dumitraş, D.G.; Marincea, Ş.; Fransolet, A.M. Brushite in the bat guano deposit from the “dry” Cioclovina Cave (Sureanu Mountains, Romania). Neues Jahrb. für Mineral. Abhandlungen 2004, 180, 45–64. [Google Scholar] [CrossRef]
- Onac, B.P.; Sumrall, J.; Mylroie, J.E.; Kearns, J.B. Cave minerals of San Salvador Island, Bahamas; University of South Florida: Tampa Library: Tampa, FL, USA, 2009; 70p. [Google Scholar]
- Snow, M.R.; Pring, A.; Allen, N. Minerals of the Wooltana Cave, Flinders Ranges, South Australia. Trans. R. Soc. S. Aust. 2014, 138, 214–230. [Google Scholar] [CrossRef]
- Dumitraş, D.G. A re-investigation of ardealite from the type locality, the “dry” Cioclovina Cave (Şureanu Mountains, Romania). Europ. J. Mineral. 2017, 529, 1055–1066. [Google Scholar] [CrossRef]
- Goldberg, P.S.; Nathan, Y. The phosphate mineralogy of et-Tabun cave, Mount Carmel, Israel. Mineral. Mag. 1975, 40, 253–258. [Google Scholar] [CrossRef]
- Quattropani, L.; Charlet, L.; De Lumley, H.; Menu, M. Early Palaeolithic bone diagenesis in the Arago cave at Tautavel, France. Mineral. Mag. 1999, 63, 801–812. [Google Scholar] [CrossRef]
- Onac, B.P.; Kearns, J.; Breban, R.; Cĩntă Pânzaru, S. Variscite (AlPO4·2H2O) from Cioclovina Cave (Sureanu Mountains, Romania): A tale of a missing phosphate. Studia UBB Geol. 2004, 49, 3–14. [Google Scholar] [CrossRef]
- Giurgiu, A.; Tămaş, T. Mineralogical data on bat guano deposits from three Romanian caves. Studia UBB Geol. 2013, 58, 13–18. [Google Scholar] [CrossRef]
- Marincea, Ş.; Dumitraş, D.G. The occurrence of taranakite in the in the “dry” Cioclovina Cave (Sureanu Mountains, Romania). Neues Jahrb. für Mineral. Abhandlungen 2003, 3, 127–144. [Google Scholar] [CrossRef]
- Dumitraş, D.G.; Marincea, Ş.; Diaconu, G. Leucophosphite and taranakite in the bat-guano deposit from Lazului Cave (Mehedinți Mountains). Rom. J. Miner. Depos. 2002, 80, 23–25. [Google Scholar]
- Morley, M.V.; Goldberg, P.; Uliyanov, V.A.; Kozlikin, M.B.; Shunkov, M.V.; Derevianko, A.P.; Jacobs, Z.; Roberts, R.G. Hominin and animal activities in the microstratigraphic record from Denisova Cave (Altai Mountains, Russia). Sci. Rep. 2019, 9, 13785. [Google Scholar] [CrossRef]
- Ulyanov, V.A.; Shunkov, M.V. Some features of sedimentation in the East Chamber of Denisova Cave. In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Molodin, V.I., Eds.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2013; Volume 19, pp. 159–162. (In Russian) [Google Scholar]
- Agadzhanyan, A.K.; Shunkov, M.V.; Kozlikin, M.B. Small Vertebrates from the Pleistocene Sediments of the East Chamber of Denisova Cave, Northwestern Altai Mountains. Paleontol. J. 2021, 55, 331–342. [Google Scholar] [CrossRef]
- Vasiliev, S.K.; Kozlikin, M.B.; Shunkov, M.V. Megafaunal Remains from the upper portion of pleistocene deposits in South Chamber of Denisova Cave. In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Molodin, V.I., Eds.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2018; Volume 24, pp. 37–41. (In Russian) [Google Scholar]
- Derevianko, A.P.; Shunkov, M.V.; Kozlikin, M.B.; Ulyanov, V.A.; Chekha, A.M. The upper Paleolithic of the East Chamber in Denisova Cave: New data. In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Molodin, V.I., Eds.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2014; Volume 20, pp. 50–53. (In Russian) [Google Scholar]
- Sokol, E.V.; Kokh, S.N.; Kozmenko, O.A.; Lavrushin, V.Y.; Belogub, E.V.; Khvorov, P.V.; Kikvadze, O.E. Boron in an onshore mud volcanic environment: Case study from the Kerch Peninsula, the Caucasus continental collision zone. Chem. Geol. 2019, 525, 58–81. [Google Scholar] [CrossRef]
- Shatsky, V.; Sitnikova, E.; Kozmenko, O.; Palessky, S.; Nikolaeva, I.; Zayachkovsky, A. Behavior of incompatible elements during ultrahigh-pressure metamorphism (by the example of rocks of the Kokchetav massif). Russ. Geol. Geophys. 2006, 47, 482–496. [Google Scholar]
- Sakae, T.; Nagata, H.; Sudo, T. The crystal structure of synthetic calcium phosphate-sulfate hydrate, Ca2HPO4SO4·4H2O, and its relation to brushite and gypsum. Am. Mineral. 1978, 63, 520–527. [Google Scholar]
- Sakae, T.; Sudo, T. Taranakite from the Onino-Iwaya limestone cave at Hiroshima Prefecture, Japan: A new occurrence. Am. Mineral. 1975, 60, 331–334. [Google Scholar]
- Capitelli, F.; Bosi, F.; Capelli, S.C.; Radica, F.; Della Ventura, G. Neutron and XRD single-crystal diffraction study and vibrational properties of whitlockite, the natural counterpart of synthetic tricalcium phosphate. Crystals 2021, 11, 225. [Google Scholar] [CrossRef]
- International Centre for Diffraction Data. PDF-4+. Powder Diffraction File; Windows; International Centre for Diffraction Data: Newtown Square, PA, USA, 2009. [Google Scholar]
- Xu, G.; Aksay, I.A.; Groves, J.T. Continuous crystalline carbonate apatite thin films. A biomimetic approach. J. Am. Chem. Soc. 2001, 123, 2196–2203. [Google Scholar] [CrossRef]
- Goldberg, M.A.; Krohicheva, P.A.; Fomin, A.S.; Khairutdinova, D.R.; Antonova, O.S.; Baikin, A.S.; Smirnov, V.V.; Fomina, A.A.; Leonov, A.V.; Mikheev, I.V.; et al. In situ magnesium calcium phosphate cements formation: From one pot powders precursors synthesis to in vitro investigations. Bioact. Mater. 2020, 5, 644–658. [Google Scholar] [CrossRef]
- Cacciotti, I. Cationic and anionic substitutions in hydroxyapatite. In Handbook of Bioceramics and Biocomposites; Springer: Berlin/Heidelberg, Germany, 2016; pp. 145–211. [Google Scholar]
- Beaufils, S.; Rouillon, T.; Millet, P.; Le Bideau, J.; Weiss, P.; Chopart, J.P.; Daltin, A.L. Synthesis of calcium-deficient hydroxyapatite nanowires and nanotubes performed by template-assisted electrodeposition. Mater. Sci. Engineer. C 2019, 98, 333–346. [Google Scholar] [CrossRef]
- Minh, D.P.; Tran, N.D.; Nzihou, A.; Sharrock, P. Carbonate-containing apatite (CAP) synthesis under moderate conditions starting from calcium carbonate and orthophosphoric acid. Mater. Sci. Eng. C 2013, 33, 2971–2980. [Google Scholar] [CrossRef] [Green Version]
- Li, J. Structural characterisation of apatite-like materials. Ph.D. Thesis, University of Birmingham, Birmingham, UK, 2010. [Google Scholar]
- International Centre for Diffraction Data. PDF-2. Powder Diffraction File; Windows; Newtown Square: International Centre for Diffraction Data: Newtown Square, PA, USA, 2005. [Google Scholar]
- Markin, S.V. Natural scientific and archaeological evidence from the pleistocene layers of Kaminnaya cave. In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Ed.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2021; Volume 26, pp. 182–187. (In Russian) [Google Scholar] [CrossRef]
- Derevianko, A.P.; Glinskii, S.V.; Dergacheva, M.I.; Dupal, T.A.; Efremov, S.A.; Zenin, A.N.; Krivoshapkin, A.I.; Kulikov, O.I.; Malaeva, E.M.; Markin, S.V.; et al. The Problems of Paleoecology, Geology and Archaeology of the Paleolithic of the Altai; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 1998; 312p. (In Russian) [Google Scholar]
- Nriagu, J.O. Phosphate-clay mineral relations in soils and sediments. Can. J. Earth Sci. 1976, 13, 717–736. [Google Scholar] [CrossRef]
- Savenko, A.V. The possibility of phosphatization of silicates in the supergene zone. Geochem. Int. 2015, 53, 87–94. [Google Scholar] [CrossRef]
- Roncal-Herrero, T.; Oelkers, E.H. Does variscite control phosphate availability in acidic natural waters? An experimental study of variscite dissolution rates. Geochim. Cosmochim. Acta 2011, 75, 416–426. [Google Scholar] [CrossRef]
- Liu, P.L.; Sherman, G.D.; Swindale, L.D. Laboratory formation and characterization of taranakite in a hydrol humic Latosol soil from Hawaii. Pacific Sci. 1966, 20, 496–506. [Google Scholar]
- Karkanas, P.; Kyparissi-Apostolika, N.; Bar-Yosef, O.; Weiner, S. Mineral assemblages in Theopetra, Greece: A framework for understanding diagenesis in a prehistoric cave. J. Archeol. Sci. 1999, 26, 1171–1180. [Google Scholar] [CrossRef] [Green Version]
- Krauskopf, K.B.; Bird, D.K. Introduction to Geochemistry; McGraw-Hill Book Company: New York, NY, USA, 1979; 617p. [Google Scholar]
- Eltantawy, M.M.; Belokon, M.A.; Belogub, E.V.; Ledovich, O.I.; Skorb, E.V.; Ulasevich, S.A. Self-Assembled Liesegang Rings of Hydroxyapatite for Cell Culturing. Adv. NanoBiomed Res. 2021, 1, 2000048. [Google Scholar] [CrossRef]
- Kizalaite, A.; Grigoraviciute-Puroniene, I.; Asuigui, D.R.C.; Stoll, S.L.; Cho, S.H.; Sekino, T.; Kareiva, A.; Zarkov, A. Dissolution-Precipitation Synthesis and Characterization of Zinc Whitlockite with Variable Metal Content. ACS Biomater. Sci. Eng. 2021, 7, 3586–3593. [Google Scholar] [CrossRef]
- Zhou, J.M.; Liu, C.; Huang, P.M. Perturbation of taranakite formation by ferrous and ferric iron under acidic conditions. Soil Sci. Soc. Am. J. 2000, 64, 885–892. [Google Scholar] [CrossRef]
- Toshima, T.; Hamai, R.; Tafu, M.; Takemura, Y.; Fujita, S.; Chohji, T.; Tanda, S.; Li, S.; Qin, G.W. Morphology control of brushite prepared by aqueous solution synthesis. J. Asian. Ceram. Soc. 2014, 2, 52–56. [Google Scholar] [CrossRef]
- Veresov, A.G.; Putlyaev, V.I.; Tret’yakov, Y.D. Chemistry of calcium phosphate-based inorganic materials. Ross. Khim. Zh. 2004, 48, 52–63. (In Russian) [Google Scholar]
- Boanini, E.; Silingardi, F.; Gazzano, M.; Bigi, A. Synthesis and hydrolysis of brushite (DCPD): The role of ionic substitution. Crystal Growth Design 2021, 21, 1689–1697. [Google Scholar] [CrossRef]
- Vasiliev, S.K.; Shunkov, M.V.; Kozlikin, M.B. Pleistocene climate in the Northwestern Altai reconstructed from megafauna fossils in the East Chamber of Denisova Cave. In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Molodin, V.I., Eds.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2017; Volume 23, pp. 60–64. (In Russian) [Google Scholar]
- Brown, S.; Kozlikin, M.; Shunkov, M.; Derevianko, A.; Higham, T.; Douka, K.; Korzow Richter, K. Examining collagen preservation through glutamine deamination at Denisova Cave. J. Archeol. Sci. 2021, 133, 105454. [Google Scholar] [CrossRef]
- Brown, S.; Wang, N.; Oertle, A.; Kozlikin, M.B.; Shunkov, M.V.; Derevianko, A.P.; Comeskey, D.; Jope-Street, B.; Harvey, V.L.; Chowdhury, M.P.; et al. Zooarchaeology through the lens of collagen fingerprinting at Denisova Cave. Sci. Rep. 2021, 11, 15457. [Google Scholar] [CrossRef] [PubMed]
- Zavala, E.I.; Jacobs, Z.; Vernot, B.; Shunkov, M.V.; Kozlikin, M.B.; Derevianko, A.P.; Essel, E.; de Fillipo, C.; Nagel, S.; Richter, J.; et al. Pleistocene sediment DNA reveals hominin and faunal turnovers at Denisova Cave. Nature 2021, 595, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Massilani, D.; Morley, M.W.; Mentzer, S.M.; Aldeias, V.; Vernot, B.; Miller, C.; Stahlschmidt, M.; Kozlikin, M.B.; Shunkov, M.V.; Derevianko, A.P.; et al. Microstratigraphic preservation of ancient faunal and hominin DNA in Pleistocene cave sediments. PNAS 2022, 119, e2113666118. [Google Scholar] [CrossRef]
- Kulik, N.A.; Kozlikin, M.B.; Miroshnichenko, L.V.; Shunkov, M.V. Impact of phosphate mineralization of sediments on the preservation of stone artifacts (based on evidence from Denisova Cave). In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Ed.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2020; Volume 24, pp. 142–147. (In Russian) [Google Scholar] [CrossRef]
- Agadzhanyan, A.K.; Rosina, V.V. Bats from the Pleistocene deposits of Denisova Cave. In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Molodin, V.I., Eds.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2001; Volume 8, pp. 33–36. (In Russian) [Google Scholar]
- Agadzhanyan, A.K.; Rosina, V.V.; Shunkov, M.V. The role of bats in the Pleistocene biogeny of the Denisova Cave area. In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Molodin, V.I., Eds.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2004; Volume 10, pp. 6–11. (In Russian) [Google Scholar]
- Rosina, V.V. The Pleistocene and Holocene History of Chiroptera Mammalia fauna in the Northwestern Altai. Ph.D. Thesis, Paleontological Institute of Russian Academy of Sciences, Moscow, Russia, 2004. (In Russian). [Google Scholar]
- Derevianko, A.P.; Shunkov, M.V.; Fedorchenko, A.Y.; Kozlikin, M.B. A Palaeolithic diadem from Denisova Cave. In Problems of Archaeology, Ethnography, Anthropology of Siberia and Neighboring Territories; Derevianko, A.P., Molodin, V.I., Eds.; Institute of Archeology and Ethnography of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2018; Volume 24, pp. 87–90. (In Russian) [Google Scholar]
- Bannikov, A.G.; Vtorov, P.P.; Gladkova, T.D.; Kuzyakin, A.P.; Naumov, S.P.; Novikov, G.A.; Rogacheva, E.V.; Tomilin, A.G.; Flint, V.E. Mammals, Eutheria. In The Life of Animals, 6 Volumes; Naumov, S.P., Kuzyakin, A.P., Eds.; Prosveshchenie: Moscow, Russia, 1971; 627p. (In Russian) [Google Scholar]
- Constantz, B.R.; Ison, I.C.; Fulmer, M.T.; Poser, R.D.; Smith, S.T.; VanWagoner, M.; Rosssteven, J.; Goldsteinjesse, A.; Jupiter, B.; Rosenthal, D.I. Skeletal repair by in situ formation of the mineral phase of bone. Science 1995, 267, 1796–1799. [Google Scholar] [CrossRef] [PubMed]























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Association | Aggregate Type | Features | Genesis | Cave Sites |
|---|---|---|---|---|
| Hydroxylapatite (carbonate-rich hydroxylapatite or dahllite) Ca5(PO4,CO3)3(OH) hexagonal | ||||
| Cal Gp Bsh Adl | (i) Massive aggregates and crusts covering fragments of carbonate host rock (ii) Nodules and veins in the guano layer (iii) Pseudomorph after bones | (i) Carbonate-, sulfate- and Zn-bearing (ii) Low crystallinity | Interaction between near neutral phosphatic solution and CaCO3 | IL: Kebara *, Etzba * and Ornit * [1] IT: Arene Candide *, Cocci, Salnitro, Eremita and Apulian Caves [24,46,47] AU: Robertson * [48] RO: Cioclovina *, Măgurici, Mare de la Mereşti, Gura Dobrogei and Huda lui Papară [49,50,51,52,53] |
| Brushite CaHPO4·2H2O monoclinic | ||||
| Ap Cal Adl Gp | (i) Crusts, nodules, veins in the guano layer (ii) Crusts over fragments of carbonate host rock or aggregates of hydroxylapatite | Sulfate-bearing | Interaction between slightly acidic phosphatic solution and CaCO3 | RO: Cioclovina *, Măgurici, Mare de la Mereşti, Gura Dobrogei and Huda lui Papară [49,50,51,52,53,54] ES: Serres Centrals [24] FR: Raganeous [24] IT: Salnitro and Apulian Caves [24,46] SV: Lighthouse, Reckley Hill, Midget Horror and Crescent Top [55] |
| Monetite CaHPO4 triclinic | ||||
| Bsh Bss Fan | (i) Dispersed grains in guano-limestone sequence (ii) Pseudomorph after brushite | No data | (i) Dehydration of brushite in dry condition (ii) Dehydration of brushite due to microbial exothermic reactions | RO: Măgurici and Gura Dobrogei [49,52] SV: Lighthouse and Dance Hall [55] |
| Whitlockite Ca9(Mg,Fe2+)(PO4)6(HPO4) trigonal | ||||
| Ap | (i) Powdered aggregates on carbonate rock surface (ii) Pseudomorph after bones | Sulfate-bearing | (i) Interaction between acidic phosphatic solution and Mg-calcite or dolomite (ii) Dissolution of apatite and subsequent precipitation from low-Ca solutions | MY: Niah [3] FR: Guano [24] SV: Lighthouse, Reckley Hill and Altar [55] AU: Wooltana [56] |
| Ardealite Ca2(SO4)(HPO4)·4H2O monoclinic | ||||
| Gp Bsh Ap | (i) Crusts and intergranular pore infillings in sulfate-phosphate mineral aggregates (ii) Powdery material near the guano-limestone contact | No data | Interaction between acidic phosphatic solution and CaCO3 | IL: Ornit * [1] SV: Lighthouse [55] RO: Cioclovina *, Măgurici, Mare de la Mereşti, Gura Dobrogei and Huda lui Papară [49,50,51,52,53,54,55,56,57] |
| Crandallite CaAl3(PO4)2(OH)5·H2O trigonal | ||||
| Gp Ap Cal | (i) Nodules (ii) Pseudomorph after bones | No data | Interaction between slightly acidic phosphatic solution and Ca-Al sediments | IL: Et-Tabun * [58] FR: Arago * [59] IT: Cocci and Carburangeli [24] |
| Variscite Al(PO4)·2H2O orthorhombic | ||||
| Tar Lpp | Crusts, nodules and veinlets within the argillaceous material | No data | Interaction between acidic phosphatic solution and argillaceous matter | IL: Ornit * [1] RO: Cioclovina *, Gaura cu Muscă and Gaura Haiducească [60,61] MY: Batu, Gangub and Makangit [3] |
| Taranakite (K,Na,NH4)3(Al,Fe3+)5(HPO4)6(PO4)2·18H2O) trigonal | ||||
| Lpp Var | Nodules in clay matrix | NH4+- and Fe3+-bearing | Interaction between acidic phosphatic solution and argillaceous matter | IL: Ornit * [1] RO: Cioclovina *, Măgurici, Gura Dobrogei, Huda lui Papară, Gaura Haiducească, Gaura Muscă and Lazului [49,52,53,61,62,63] MY: Gangub and Makangit [3] IT: Eremita, Palombara, Corona’e sa Craba and Apulian Caves [24,46] |
| Leucophosphite KFe3+2(PO4)2(OH)·2H2O monoclinic | ||||
| Tar Var | Fine-grained crusts located at the center of guano layer enriched in clay material | Al3+-bearing | Interaction between acidic phosphatic solution and Fe-rich siliciclastic matter | MY: Batu, Gangub and Makangit [3] IT: Corona ’e sa Craba [24] RO: Gaura cu Muscă and Lazului [61,63] |
| Layer | Sample | SiO2 | TiO2 | Al2O3 | Fe2O3 | MnO | MgO | CaO | Na2O | K2O | P2O5 | SO3 | ZnO | CuO | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L-6 | S-15 | 4.34 | 0.03 | 0.87 | 0.39 | 0.09 | 0.44 | 29.22 | 0.07 | 0.51 | 32.31 | 1.77 | 0.57 | 0.05 | 99.97 |
| L-7 | S-13a | 17.91 | 0.19 | 2.57 | 1.08 | 0.08 | 0.77 | 35.84 | 0.55 | 1.23 | 22.91 | 0.78 | 0.10 | 0.05 | 100.23 |
| L-7 | S-13 | 10.81 | 0.16 | 2.62 | 1.38 | 0.09 | 1.24 | 34.37 | 0.47 | 1.28 | 25.15 | 2.93 | 0.36 | 0.06 | 99.92 |
| L-7 | S-12 | 7.28 | 0.13 | 1.91 | 0.88 | 0.05 | 0.85 | 37.55 | 0.10 | 0.87 | 26.53 | 4.76 | 0.27 | 0.07 | 99.96 |
| L-7 | S-11 | 15.83 | 0.22 | 4.38 | 1.67 | 0.05 | 0.72 | 34.23 | 0.32 | 1.37 | 33.27 | 1.14 | 0.33 | 0.04 | 99.96 |
| L-8 | S-10 | 8.17 | 0.11 | 1.93 | 0.89 | 0.02 | 0.34 | 21.66 | 0.12 | 0.53 | 12.94 | 2.14 | 0.36 | 0.18 | 99.89 |
| L-8 | S-9 | 41.45 | 0.59 | 9.82 | 4.07 | 0.15 | 0.39 | 12.12 | 0.72 | 2.53 | 10.92 | 0.55 | 0.12 | <0.02 | 99.91 |
| L-9 | S-8 | 44.64 | 0.65 | 13.63 | 5.23 | 0.01 | 0.07 | 3.85 | 0.70 | 5.47 | 13.49 | 0.14 | 0.09 | <0.02 | 99.97 |
| L-9 | S-7 | 50.74 | 0.64 | 12.12 | 4.81 | 0.03 | 0.16 | 4.38 | 0.72 | 5.28 | 10.89 | 0.05 | 0.08 | 0.02 | 100.00 |
| L-9 | S-6 | 21.20 | 0.33 | 10.21 | 4.59 | 0.03 | 0.05 | 7.57 | 0.47 | 5.62 | 31.99 | 0.13 | 0.09 | <0.02 | 99.98 |
| L-9 | S-4 | 35.75 | 0.48 | 9.13 | 3.56 | 0.18 | 0.35 | 19.13 | 0.59 | 2.10 | 16.41 | 0.45 | 0.08 | <0.02 | 99.96 |
| L-11.1 | S-3 | 25.71 | 0.36 | 7.26 | 2.67 | 0.14 | 0.06 | 27.55 | 0.39 | 1.54 | 19.32 | 0.92 | 0.03 | <0.02 | 99.74 |
| L-11.2 | S-2 | 31.87 | 0.43 | 8.30 | 2.88 | 0.08 | 0.87 | 25.77 | 0.42 | 1.54 | 12.49 | 0.73 | 0.04 | <0.02 | 99.96 |
| L-11.2 | S-1 | 33.96 | 0.43 | 7.88 | 3.64 | 0.06 | 1.01 | 24.29 | 0.40 | 1.79 | 11.29 | 0.92 | 0.05 | <0.02 | 99.97 |
| Layer | Sample | Corg | H | N | S |
|---|---|---|---|---|---|
| L-6 | S-15 | 7.68 | 1.34 | 2.17 | 2.86 |
| L-7 | S-13a | 3.39 | 0.97 | 0.49 | <0.3 |
| L-7 | S-13 | <0.3 | 1.20 | <0.3 | 3.44 |
| L-7 | S-12 | 0.69 | 1.17 | <0.3 | 3.48 |
| L-7 | S-11 | 0.81 | 0.45 | 0.33 | 0.44 |
| L-8 | S-10 | 27.84 | 1.90 | 7.88 | 0.39 |
| L-8 | S-9 | 4.77 | 0.97 | 0.95 | <0.3 |
| L-9 | S-8 | 1.07 | 1.07 | <0.3 | <0.3 |
| L-9 | S-7 | 1.09 | 0.85 | <0.3 | <0.3 |
| L-9 | S-6 | 1.74 | 1.51 | 0.37 | <0.3 |
| L-9 | S-4 | 1.83 | 0.88 | 0.42 | <0.3 |
| Phase | Taranakite (K,Na,NH4)3(Al,Fe3+)5(HPO4)6(PO4)2·18H2O) | Brushite Ca(HPO4)·2H2O | Ardealite Ca2(SO4)(HPO4)·4H2O | Whitlockite Ca9(Mg,Fe2+)(PO4)6(HPO4) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Layer 9 | Layer 11.1 | Layer 7 | Layer 7 | |||||||||||||
| Sample | Theor. | 1 | S-8 | S-8 | Theor. | 2 | S-82 | S-82 | Theor. | 3 | S-12 | S-12 | Theor. | 4 | S-12 * | S-11 ** |
| K2O | 10.53 | 7.09 | 7.6 | 8.9 | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | n.a. | 0.3 | 2.1 | |||
| Al2O3 | 18.99 | 19.03 | 20.8 | 21.6 | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | 0.22 | <0.3 | <0.3 | |||
| Fe2O3 | 0.42 | 1.5 | 1.6 | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | ||||
| CaO | n.a. | 0.7 | 0.7 | 41.24 | 40.53 | 42.7 | 41.8 | 32.58 | 31.14 | 31.4 | 31.3 | 47.05 | 46.16 | 45.3 | 46.8 | |
| SrO | n.a. | 1.4 | 1.6 | n.a. | 0.9 | 0.8 | n.a. | <0.3 | <0.3 | 0.44 | 1.4 | 0.6 | ||||
| MgO | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | 2.52 | 2.03 | 2.3 | 1.2 | |||
| FeO | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | 3.23 | 2.32 | 0.9 | 0.4 | |||
| Na2O | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | 0.80 | 0.5 | <0.3 | ||||
| CuO | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | n.a. | 0.4 | <0.3 | ||||
| ZnO | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | n.a. | <0.3 | <0.3 | n.a. | 0.5 | <0.3 | ||||
| P2O5 | 42.30 | 42.32 | 41.0 | 40.7 | 32.59 | 32.12 | 32.7 | 31.4 | 20.62 | 21.31 | 20.7 | 21.5 | 46.35 | 47.17 | 44.5 | 47.9 |
| SO3 | n.a. | <0.3 | <0.3 | n.a. | 1.2 | 0.9 | 23.26 | 22.75 | 23.2 | 22.8 | n.a. | 2.5 | <0.3 | |||
| H2O | 28.18 | 31.10 | n.a. | n.a. | 26.17 | 26.39 | n.a. | n.a. | 23.55 | 24.86 | n.a. | n.a. | 0.84 | 0.82 | n.a. | n.a. |
| Total | 100.00 | 99.96 | 73.0 | 75.1 | 100.00 | 99.04 | 77.5 | 74.9 | 100.00 | 100.06 | 75.3 | 75.6 | 100.00 | 99.96 | 98.3 | 99.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokol, E.V.; Kozlikin, M.B.; Kokh, S.N.; Nekipelova, A.V.; Kulik, N.A.; Danilovsky, V.A.; Khvorov, P.V.; Shunkov, M.V. Phosphate Record in Pleistocene-Holocene Sediments from Denisova Cave: Formation Mechanisms and Archaeological Implications. Minerals 2022, 12, 553. https://doi.org/10.3390/min12050553
Sokol EV, Kozlikin MB, Kokh SN, Nekipelova AV, Kulik NA, Danilovsky VA, Khvorov PV, Shunkov MV. Phosphate Record in Pleistocene-Holocene Sediments from Denisova Cave: Formation Mechanisms and Archaeological Implications. Minerals. 2022; 12(5):553. https://doi.org/10.3390/min12050553
Chicago/Turabian StyleSokol, Ella V., Maxim B. Kozlikin, Svetlana N. Kokh, Anna V. Nekipelova, Natalya A. Kulik, Viktoria A. Danilovsky, Pavel V. Khvorov, and Michael V. Shunkov. 2022. "Phosphate Record in Pleistocene-Holocene Sediments from Denisova Cave: Formation Mechanisms and Archaeological Implications" Minerals 12, no. 5: 553. https://doi.org/10.3390/min12050553