
Mud Crab’s Mottled, Deep-Blue Exoskeleton: Surface Morphology and Internal Microstructure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Preparation
2.2. Laser Scanning Microscope Observation
2.3. SEM Observation and EDS Analysis
2.4. Fracture-Surface Observation by SEM
2.5. Nanoindentation Tests
3. Results and Discussion
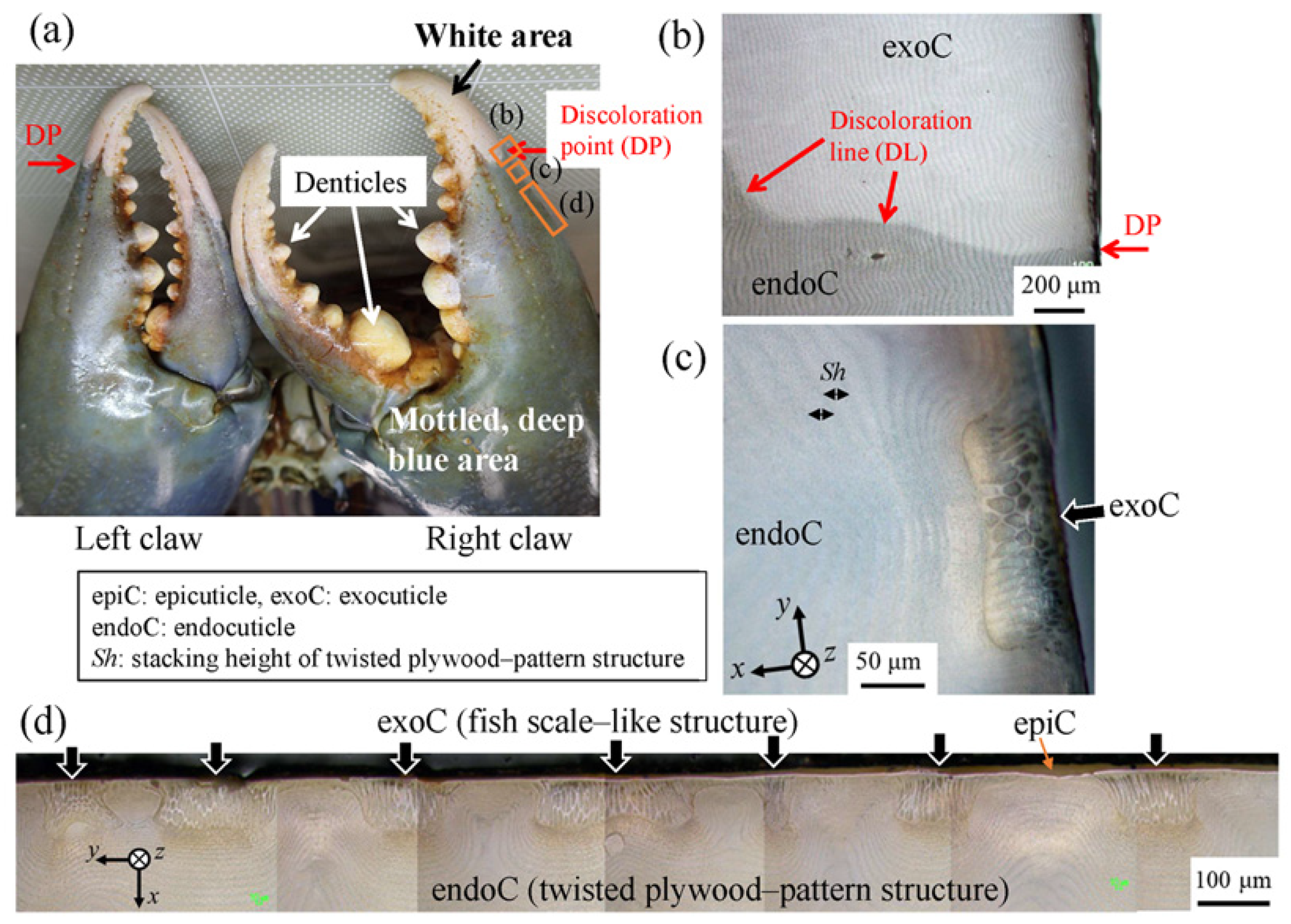
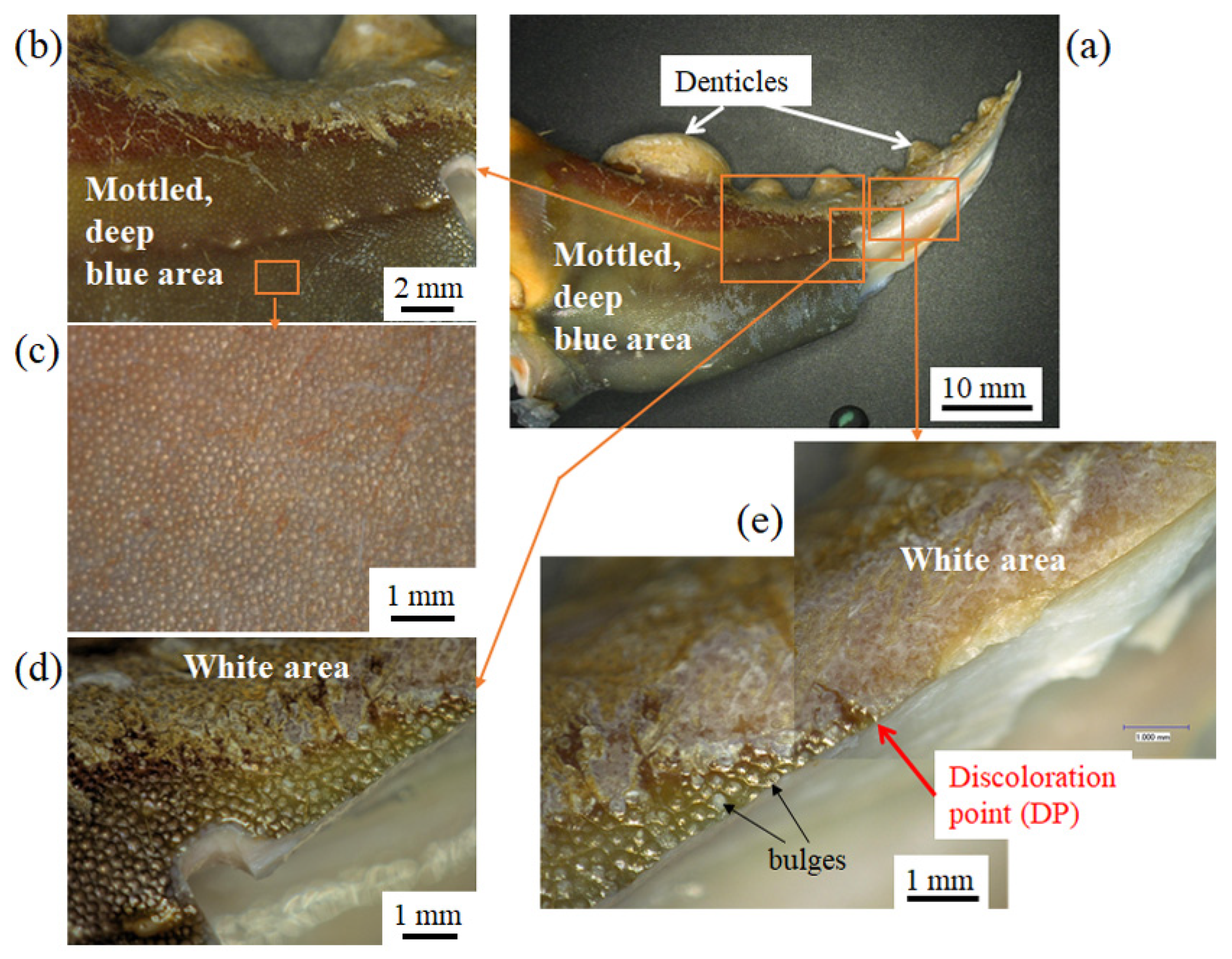
3.1. Surface Morphology
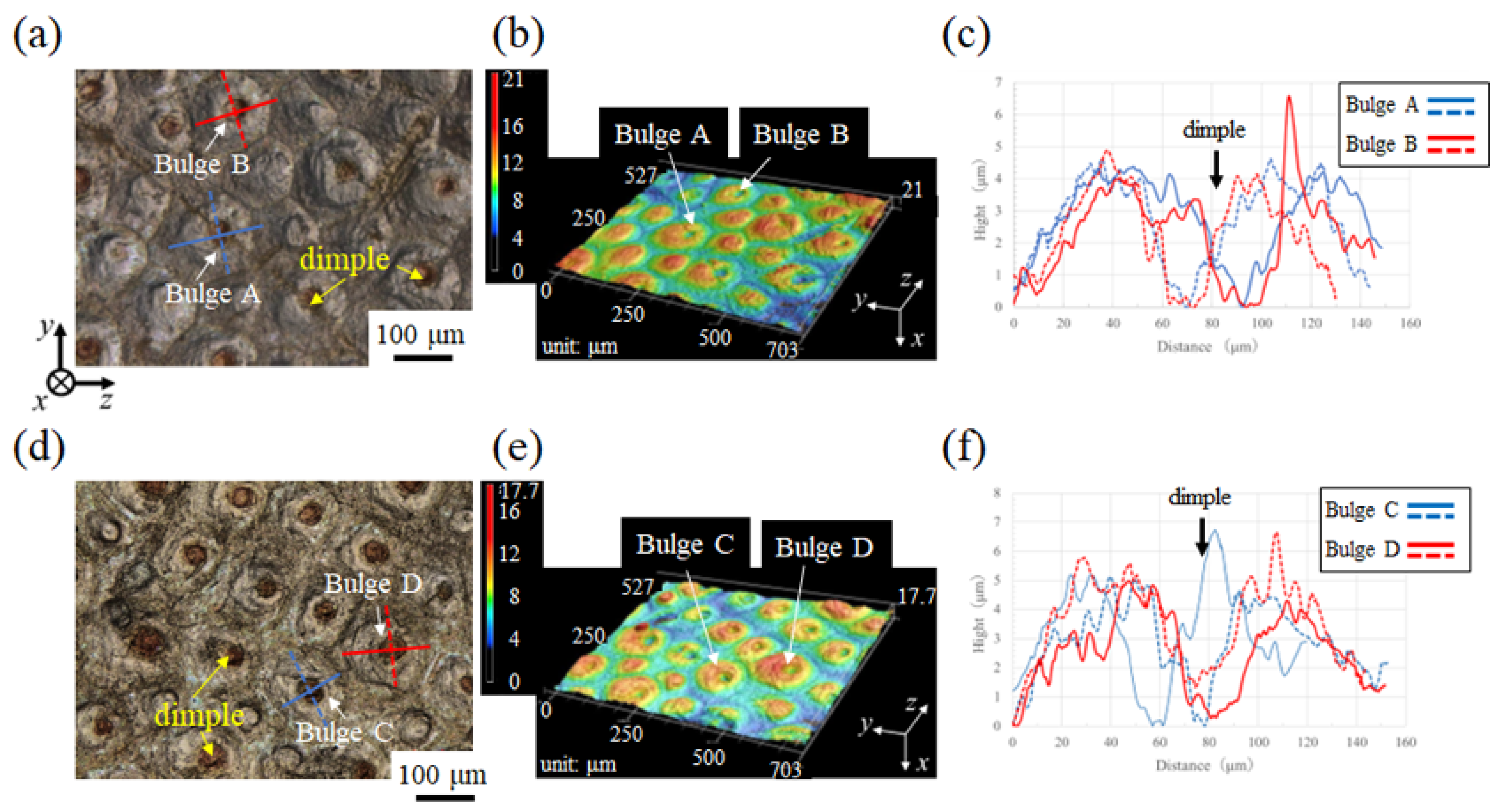
3.1.1. Fine Bulges on the Exoskeleton Surface
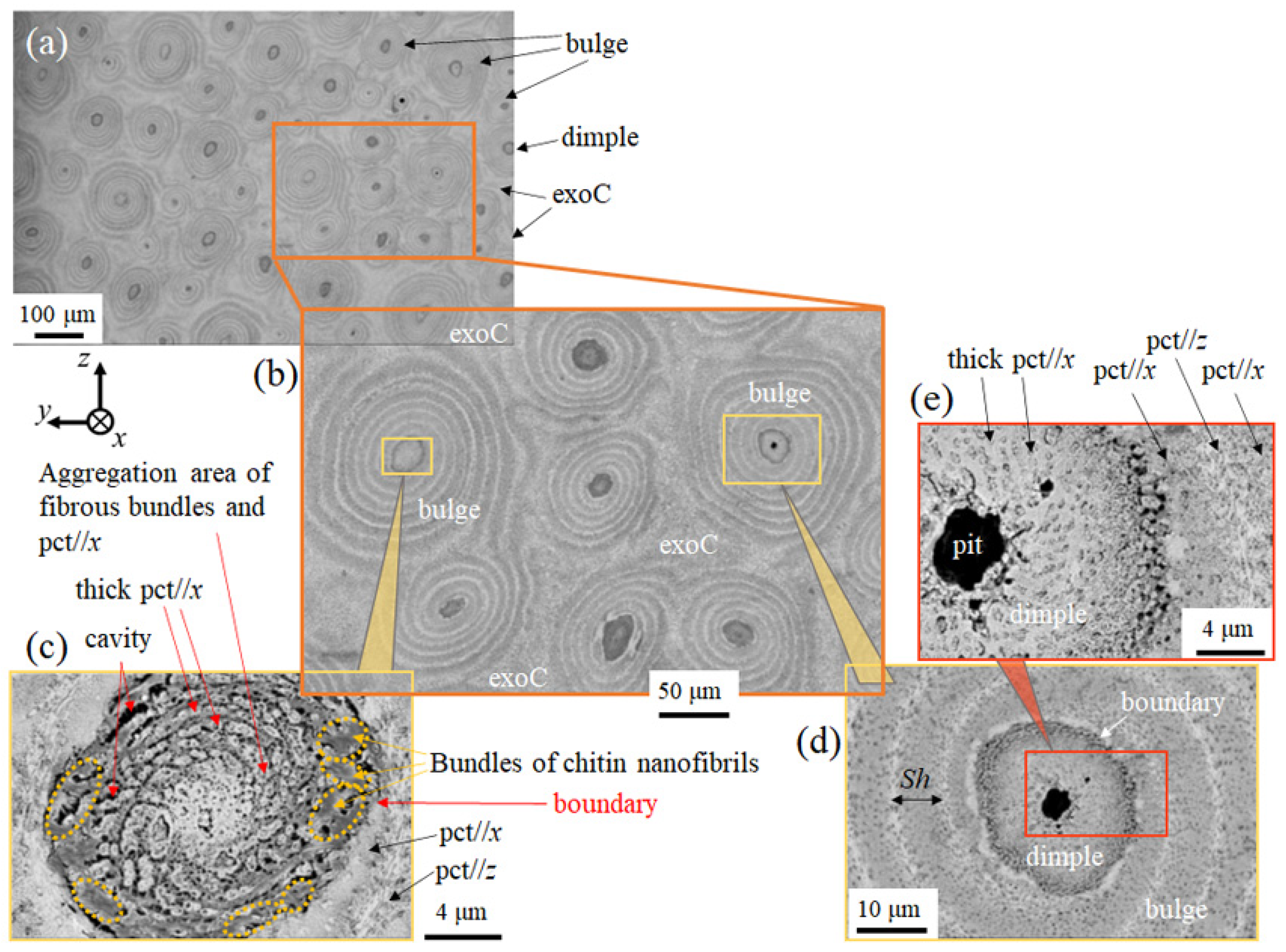
3.1.2. Surface Analysis of the Exoskeleton and Pore-Canal Tube
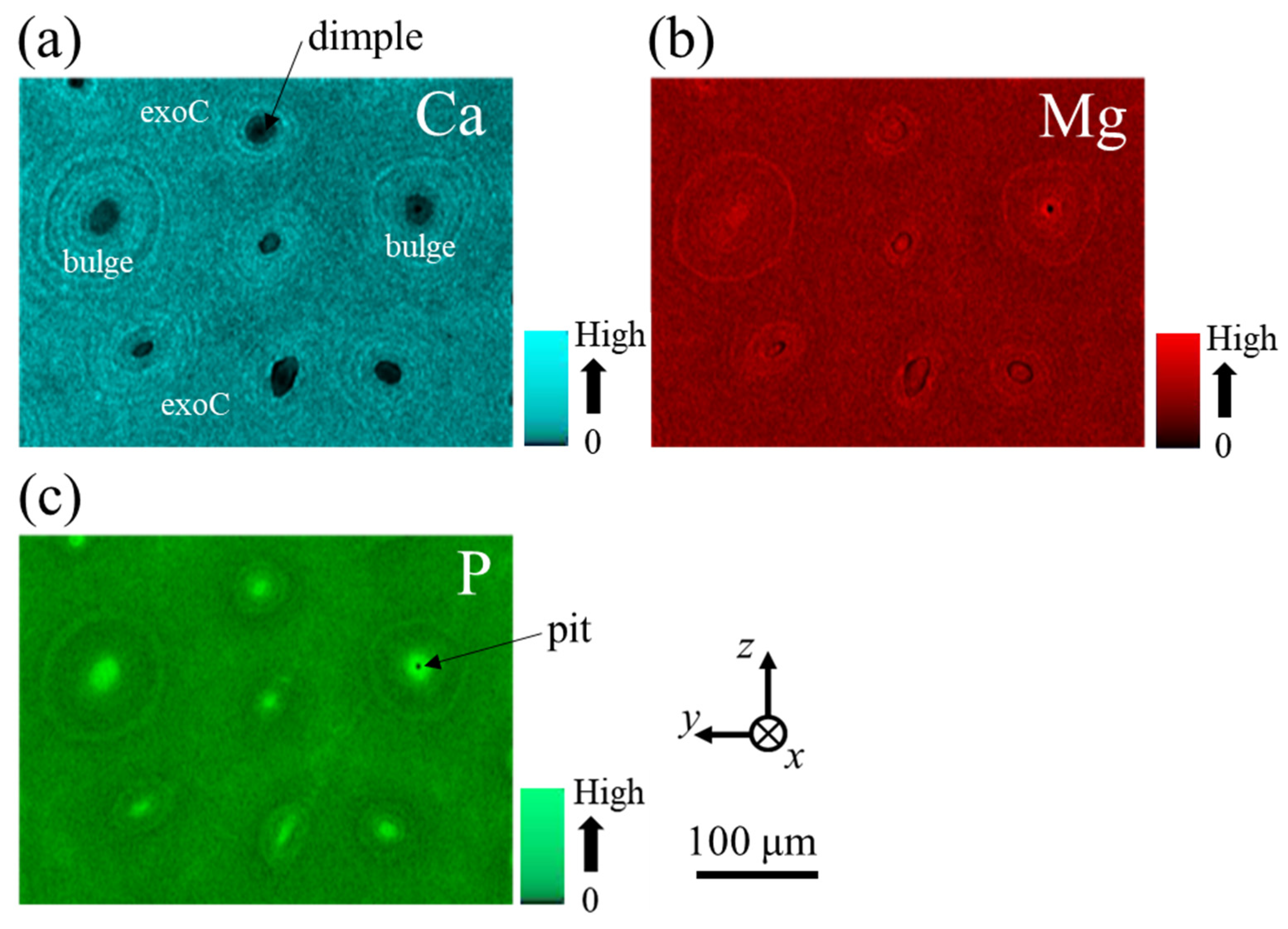
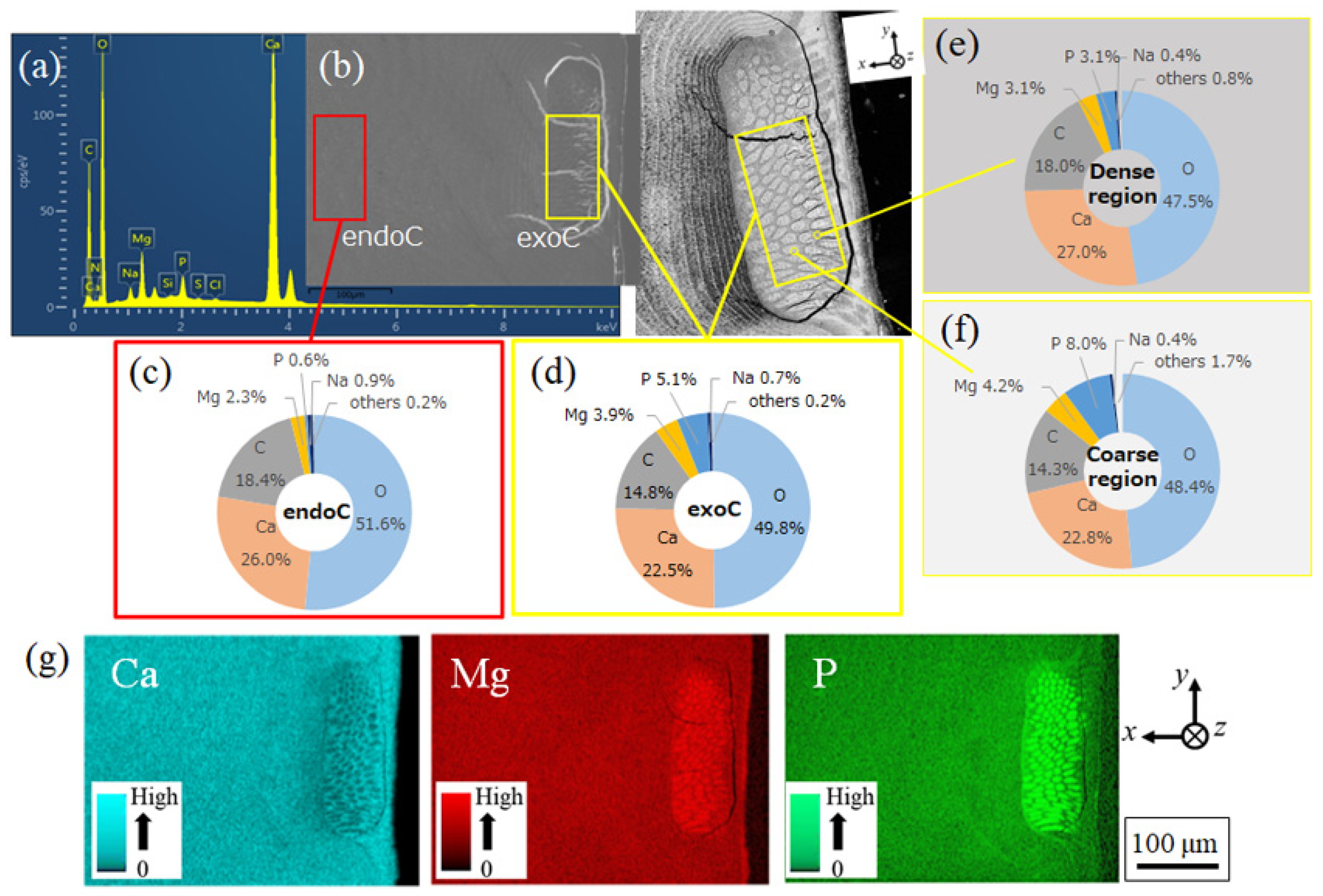
3.1.3. Microstructures of Fine Bulges and Chemical Composition
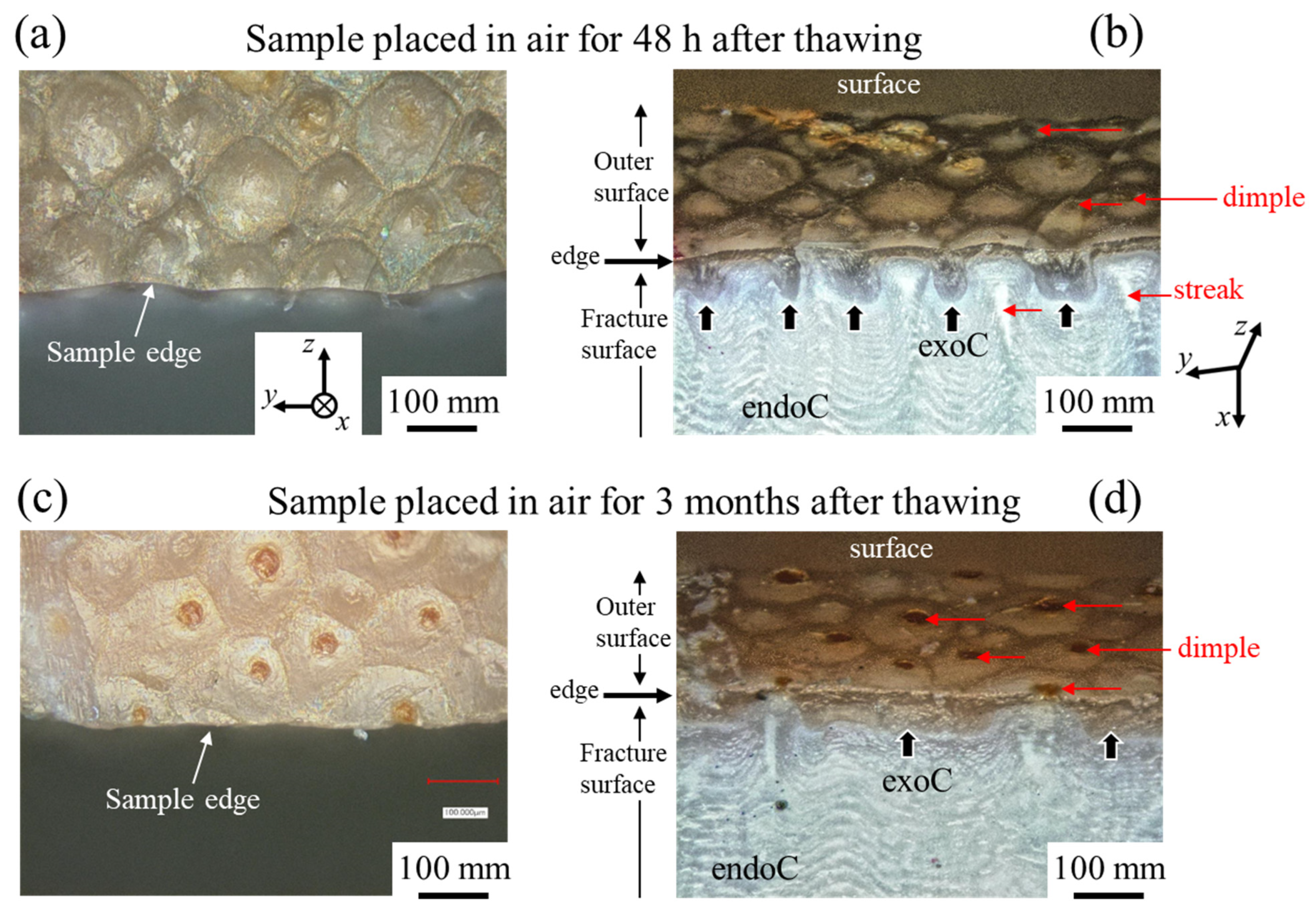
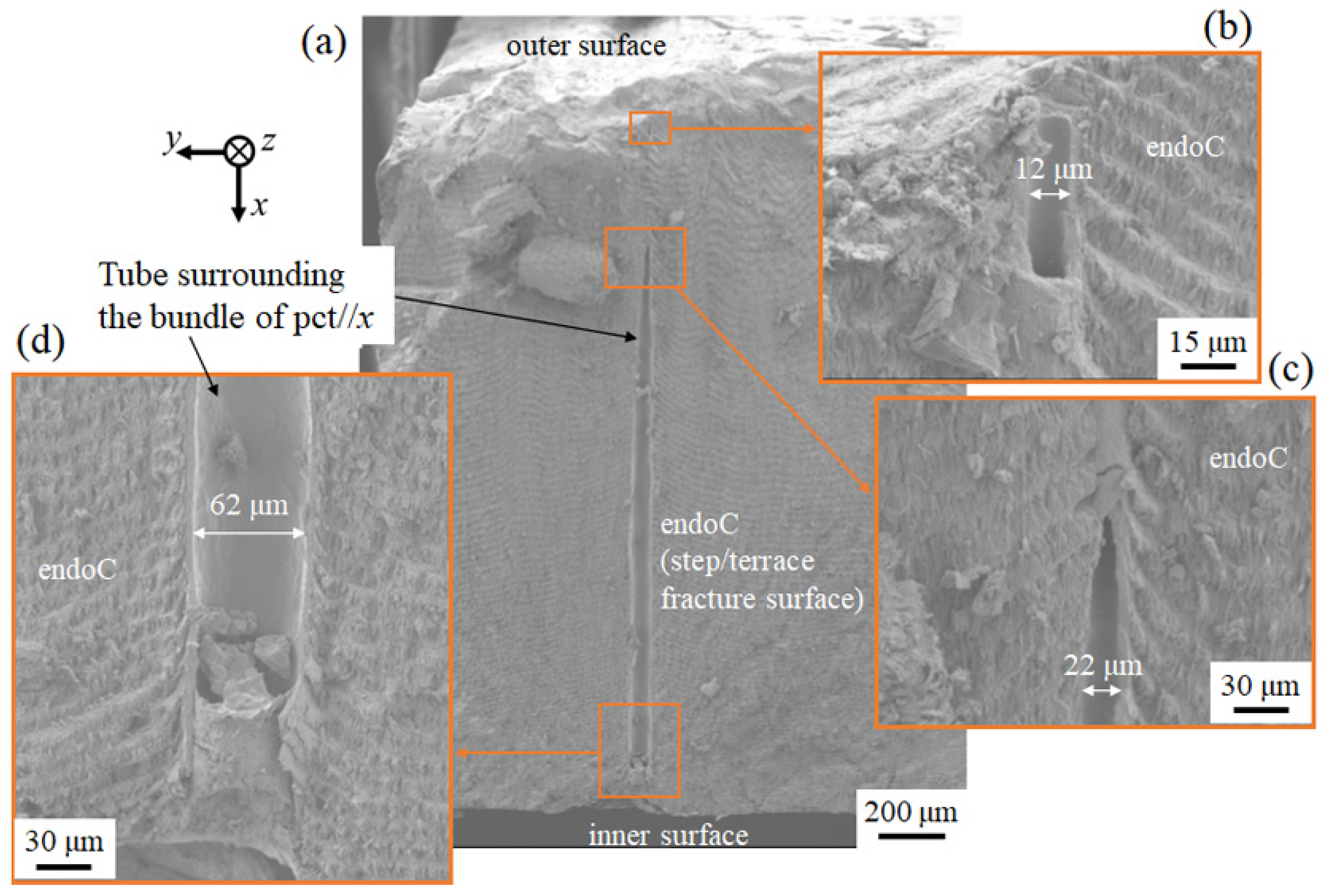
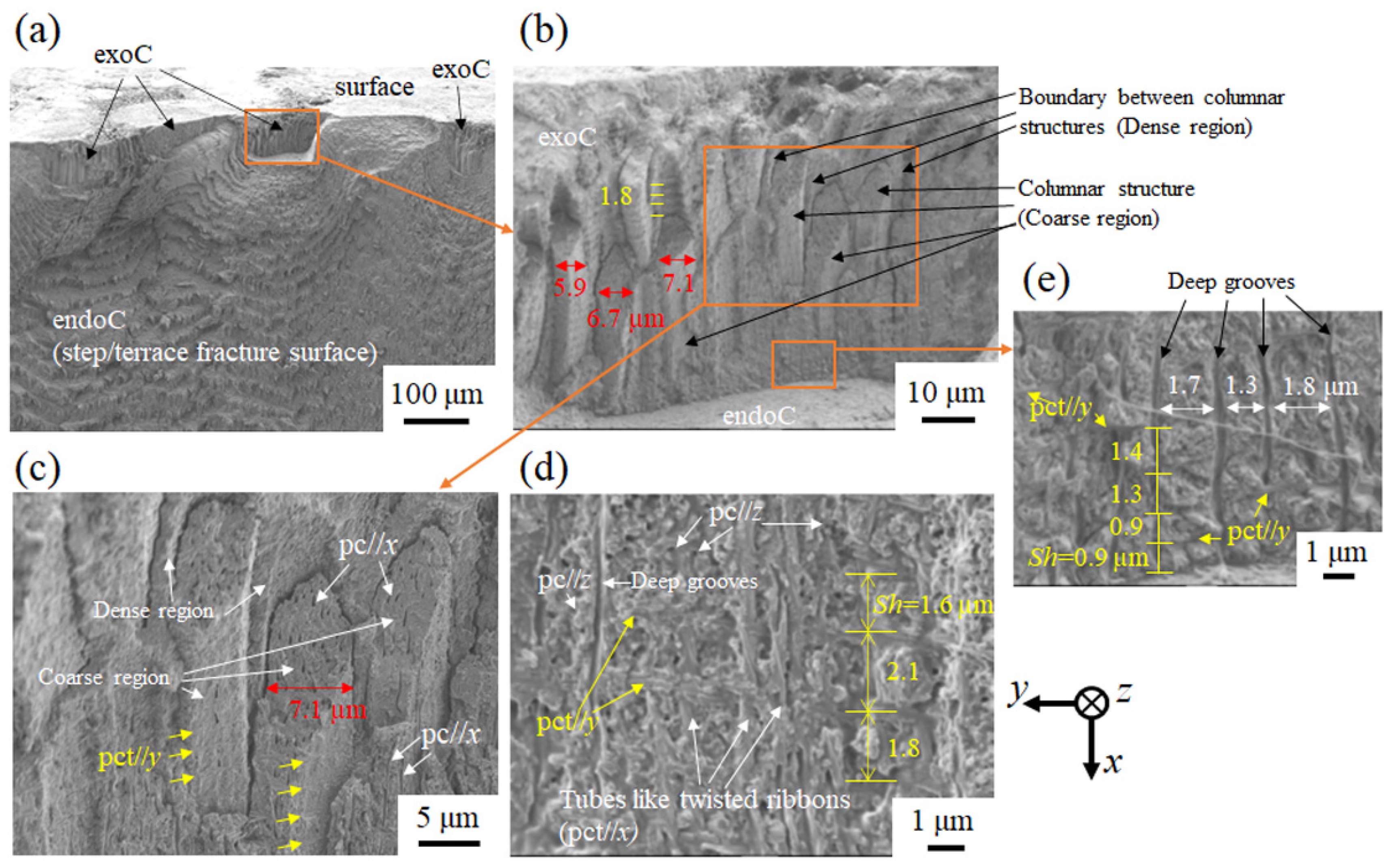
3.1.4. Fracture Surface
3.2. Fish-Scale-Like Tissue of the Exocuticle
3.2.1. Microstructures and Chemical Composition
3.2.2. Fracture Surface
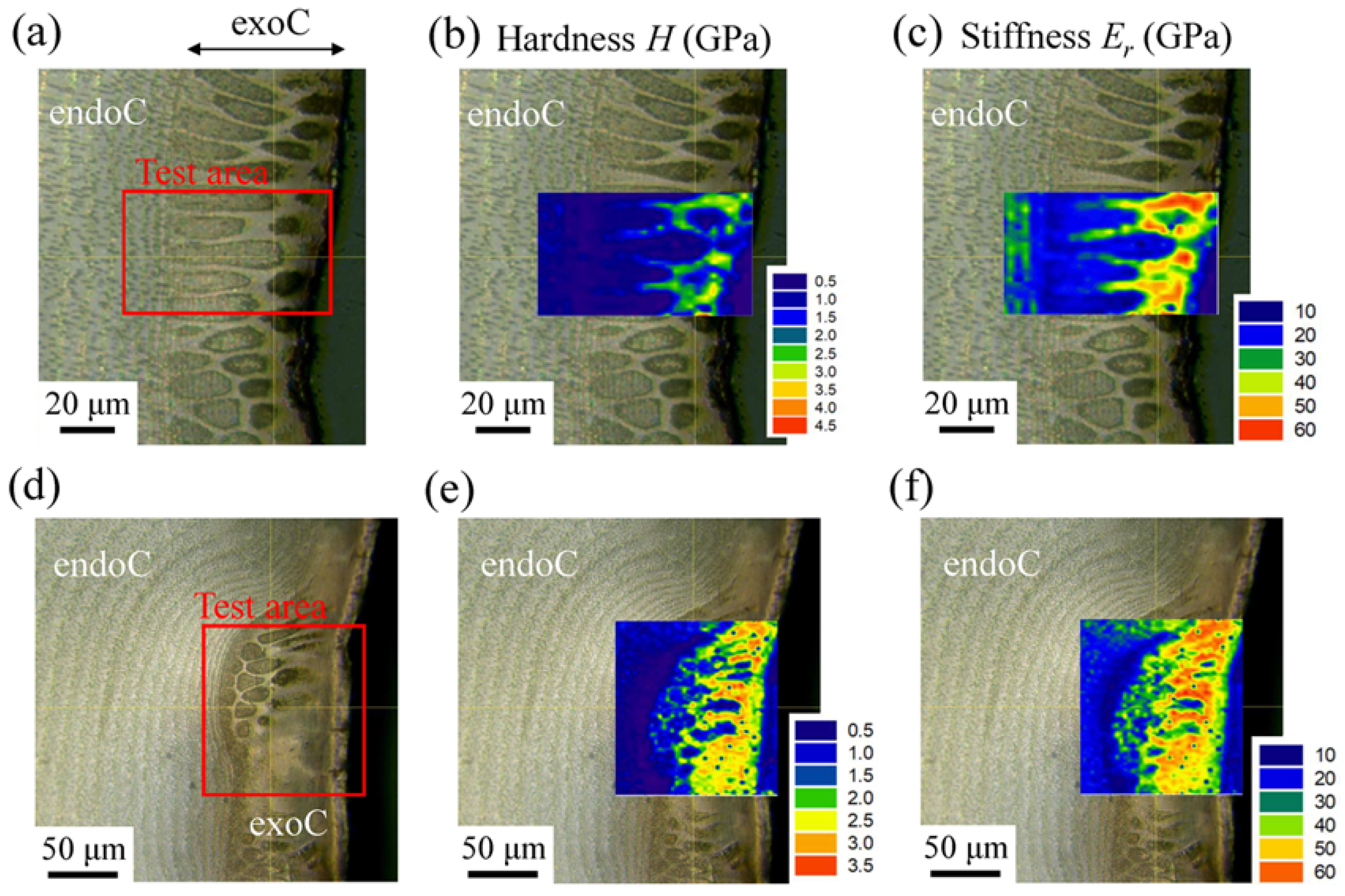
3.2.3. Mechanical Properties
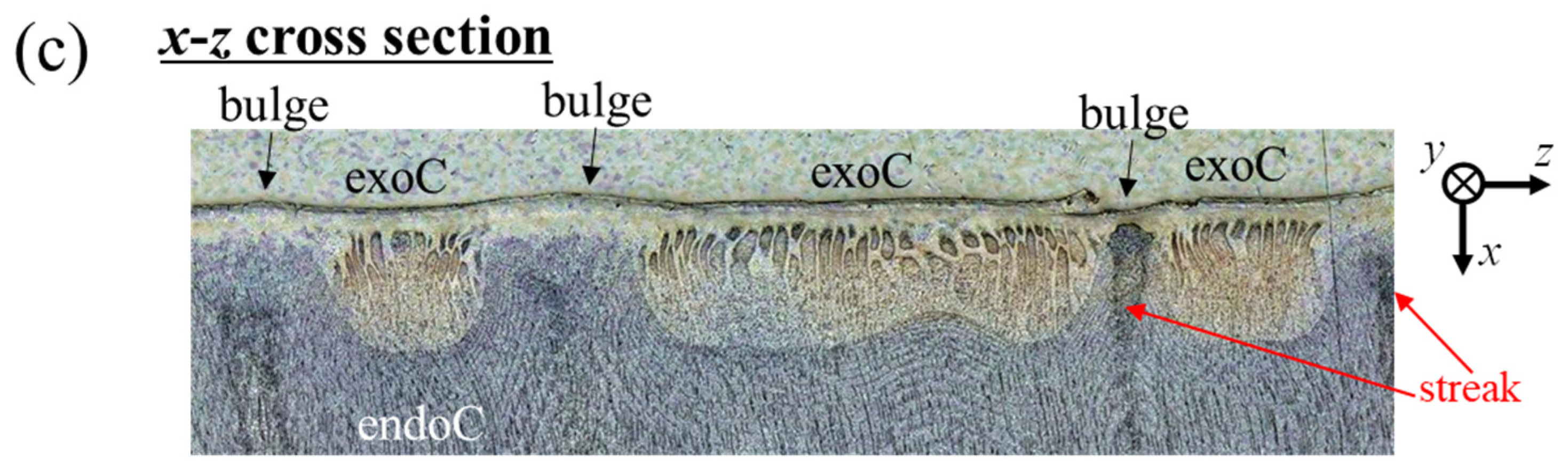
3.3. Bulges on the Mottled, Deep-Blue Surface of the Exoskeleton
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collado, A.G.; Blanco, J.M.; Gupta, M.K.; Vicentea, R.D. Advances in polymers based Multi-Material Additive-Manufacturing Techniques: State-of-art review on properties and applications. Addit. Manuf. 2022, 50, 102577. [Google Scholar] [CrossRef]
- Tiismus, H.; Kallaste, A.; Vaimann, T.; Rassõlkin, A. State of the art of additively manufactured electromagnetic materials for topology optimized electrical machines. Addit. Manuf. 2022, 55, 102778. [Google Scholar] [CrossRef]
- Zhou, Y. The Application of Ultrasound in 3D Bio-Printing. Molecules 2016, 21, 590. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.; Brooks, A.K.; Yadavalli, V.K. Nature-derived materials for the fabrication of functional biodevices. Mater. Today Bio 2020, 7, 100065. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.K.; Hazell, P.J.; Escobedo, J.P.; Wang, H. Biomimetic armour design strategies for additive manufacturing: A review. Mater. Des. 2021, 205, 109730. [Google Scholar] [CrossRef]
- Huang, W.; Restrepo, D.; Jung, J.-Y.; Su, F.Y.; Liu, Z.; Ritchie, R.O.; McKittrick, J.; Zavattieri, P.; Kisailus, D. Multiscale Toughening Mechanisms in Biological Materials and Bioinspired Designs. Adv. Mater. 2019, 31, 1901561. [Google Scholar] [CrossRef]
- Zhang, B.; Han, Q.; Zhang, J.; Han, Z.; Niu, S.; Ren, L. Advanced bio-inspired structural materials: Local properties determine overall performance. Mater. Today. 2020, 41, 177–199. [Google Scholar] [CrossRef]
- Sun, Y.; Tian, W.; Zhang, T.; Chen, P.; Li, M. Strength and toughness enhancement in 3d printing via bioinspired tool path. Mater. Des. 2020, 185, 108239. [Google Scholar] [CrossRef]
- Huber, D.R.; Eason, T.G.; Hueter, R.E.; Motta, P.J. Analysis of the bite force and mechanical design of the feeding mechanism of the durophagous horn shark Heterodontus francisci. J. Exp. Biol. 2005, 208, 3553–3571. [Google Scholar] [CrossRef] [Green Version]
- Inoue, T.; Oka, S.; Hara, T. Three-dimensional microstructure of robust claw of coconut crab, one of the largest terrestrial crustaceans. Mater. Des. 2021, 206, 109765. [Google Scholar] [CrossRef]
- Taylor, G.M. Maximum force production: Why are crabs so strong? Proc. Biol. Sci. 2000, 267, 1475–1480. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Song, Z.; Zhang, S.; Ni, Y.; Cai, S.; Gong, X.; He, L.; Yu, S. Discontinuous fibrous Bouligand architecture enabling formidable fracture resistance with crack orientation insensitivity. Proc. Natl. Acad. Sci. USA 2020, 117, 15465–15472. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, B.; Gilman, J.W. Bioinspired Bouligand cellulose nanocrystal composites: A review of mechanical properties. Phil. Trans. R. Soc. A 2017, 376, 20170050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raabe, D.; Sachs, C.; Romano, P. The crustacean exoskeleton as an example of a structurally and mechanically graded biological nanocomposite material. Acta Mater. 2005, 53, 4281–4292. [Google Scholar] [CrossRef]
- Inoue, T.; Hara, T.; Nakazato, K.; Oka, S. Superior mechanical resistance in the exoskeleton of the coconut crab, Birgus latro. Mater. Today Bio 2021, 12, 100132. [Google Scholar] [CrossRef]
- Ma, Y.; Guo, C.; Dai, N.; Shen, J.; Guan, J. Structural characterization and regulation of the mechanical properties of the carapace cuticle in tri-spine horseshoe crab (Tachypleus tridentatus). J. Mech. Behav. Biomed. Mater. 2022, 125, 104954. [Google Scholar] [CrossRef]
- Cheng, L.; Thomas, A.; Glancey, J.L.; Karlsson, A.M. Mechanical behavior of bio-inspired laminated composites. Compos. Part A Appl. Sci. Manuf. 2011, 42, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Yano, I. A histochemical study on the exocuticle with respect to its calcification and associated epidermal cells in a shore crab. Bull. Jpn. Soc. Sci. Fish. 1972, 38, 733–739. [Google Scholar] [CrossRef]
- Rose, R.; Dillaman, R. The Structure and Calcification of the Crustacean Cuticle. Am. Zool. 1984, 24, 893–909. [Google Scholar] [CrossRef]
- Fabritius, H.-O.; Karsten, E.S.; Balasundaram, K.; Hild, S.; Huemer, K.; Raabe, D. Correlation of structure, composition and local mechanical properties in the dorsal carapace of the edible crab Cancer pagurus. Z. Kristallogr. 2012, 227, 766–776. [Google Scholar] [CrossRef]
- Fabritius, H.-O.; Sachs, C.; Triguero, P.R.; Raabe, D. Influence of structural principles on the mechanics of a biological fiber-based composite material with hierarchical organization: The exoskeleton of the lobster homarus americanus. Adv. Mater. 2009, 21, 391–400. [Google Scholar] [CrossRef]
- Inoue, T.; Oka, S.; Nakazato, K.; Hara, T. Structural changes and mechanical resistance of claws and denticles in coconut crabs of different sizes. Biology 2021, 10, 1304. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Oka, S.; Nakazato, K.; Hara, T. Columnar Structure of Claw Denticles in the Coconut Crab, Birgus latro. Minerals 2022, 12, 274. [Google Scholar] [CrossRef]
- Nekvapil, F.; Pinzaru, S.C.; Barbu-Tudoran, L.; Suciu, M.; Glamuzina, B.; Tamaș, T.; Chiș, V. Color-specific porosity in double pigmented natural 3d-nanoarchitectures of blue crab shell. Sci. Rep. 2020, 10, 3019. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.A.-H.S.Y.; Meynecke, J.-Q.; Diele, K.; Nordhaus, I.; Wolff, M. Life-history, movement, and habitat use of Scylla serrata (Decapoda, Portunidae): Current knowledge and future challenges. Hydrobiologia 2016, 763, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Waugh, D.A.; Feldmann, R.M.; Schroeder, A.M.; Mutel, M.H. Differential cuticle architecture and its preservation in fossil and extant Callinectes and Scylla claws. J. Crustacean Biol. 2006, 26, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Inoue, T.; Hiroto, T.; Hara, Y.; Nakazato, K.; Oka, S. Exoskeleton of huge claws of mud crab, Scylla serrata, Tissue structure and mechanical properties. J. Mater. Sci. 2022. [Google Scholar]
- Amini, S.; Miserez, A. Wear and abrasion resistance selection maps of biological materials. Acta Biomater. 2013, 9, 7895–7907. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Lin, A.Y.; McKittrick, J.; Meyers, M.A. Structure and mechanical properties of crab exoskeletons. Acta Biomater. 2008, 4, 587–596. [Google Scholar] [CrossRef]
- Sachs, C.; Fabritius, H.; Raabe, D. Experimental investigation of the elastic–plastic deformation of mineralized lobster cuticle by digital image correlation. J. Struct. Biol. 2006, 155, 409–425. [Google Scholar] [CrossRef]
- Barthlott, W.; Mail, M.; Bhushan, B.; Koch, K. Plant Surfaces: Structures and Functions for Biomimetic Innovations. Nano-Micro Lett. 2017, 9, 1265–1305. [Google Scholar] [CrossRef] [PubMed]
- Darmanin, T.; Guittard, F. Superhydrophobic and superoleophobic properties in nature. Mater. Today 2015, 18, 273–285. [Google Scholar] [CrossRef]
- Inoue, T.; Yin, F.; Kimura, Y.; Tsuzaki, K.; Ochiai, S. Delamination Effect on Impact Properties of Ultrafine-Grained Low-Carbon Steel Processed by Warm Caliber Rolling. Met. Mater. Trans. A 2010, 41, 341–355. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, T.; Kitahara, E.; Hara, Y.; Nakazato, K. Mud Crab’s Mottled, Deep-Blue Exoskeleton: Surface Morphology and Internal Microstructure. Minerals 2022, 12, 1607. https://doi.org/10.3390/min12121607
Inoue T, Kitahara E, Hara Y, Nakazato K. Mud Crab’s Mottled, Deep-Blue Exoskeleton: Surface Morphology and Internal Microstructure. Minerals. 2022; 12(12):1607. https://doi.org/10.3390/min12121607
Chicago/Turabian StyleInoue, Tadanobu, Erina Kitahara, Yuka Hara, and Koji Nakazato. 2022. "Mud Crab’s Mottled, Deep-Blue Exoskeleton: Surface Morphology and Internal Microstructure" Minerals 12, no. 12: 1607. https://doi.org/10.3390/min12121607