
Testing the Capacity of Staphylococcus equorum for Calcium and Copper Removal through MICP Process

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Selection
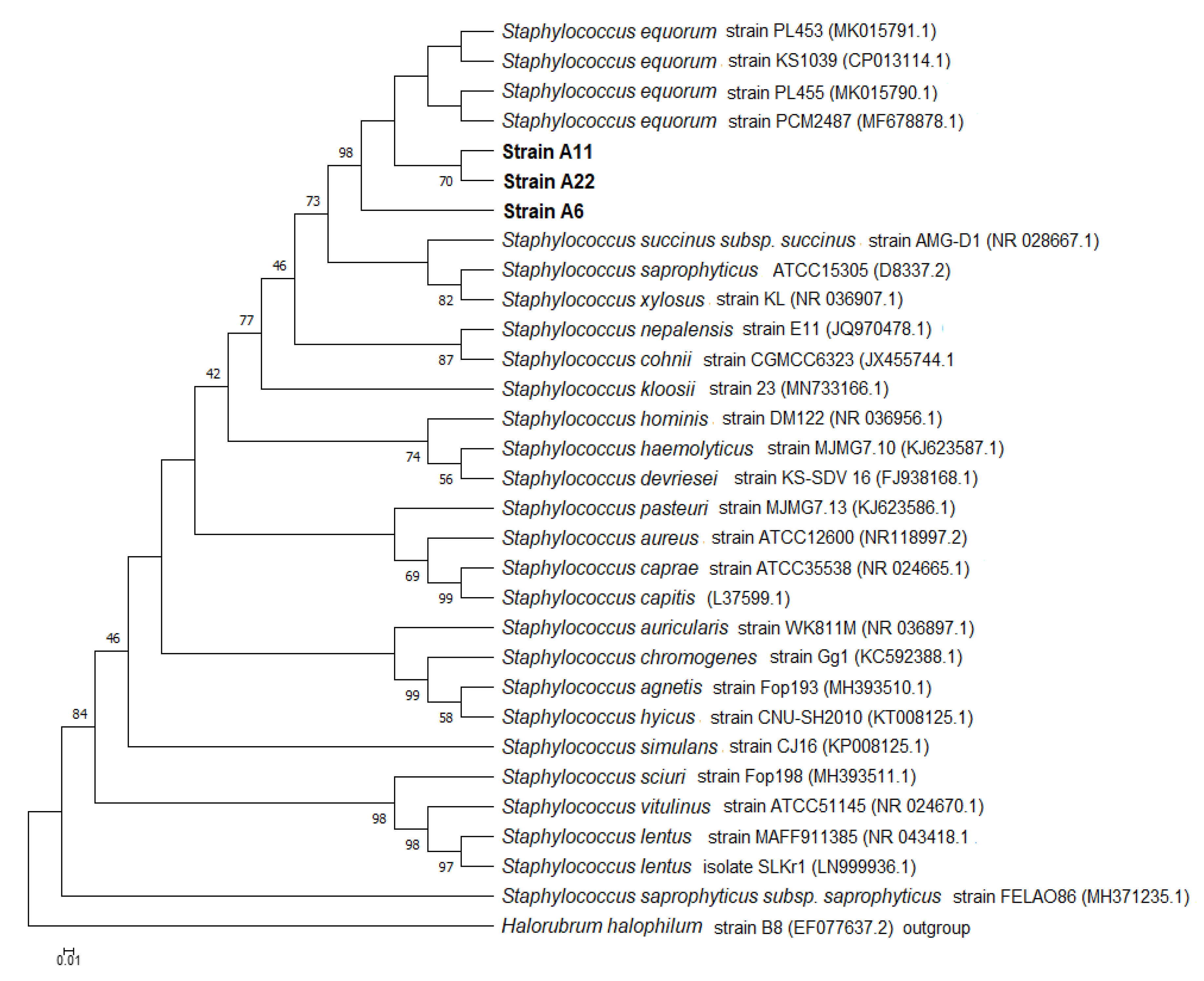
2.2. Bacterial Identification by 16S rRNA Gene, Phylogenetic Analysis and Nucleotide Sequence Accesion Numbers
2.3. Bacterial and Mineral Characterization
2.4. Copper Tolerance and Copper Removal Assay
2.5. SEM-EDX
3. Results
3.1. Isolation and Identification of Bacterial Culture with Ureolytic Activity
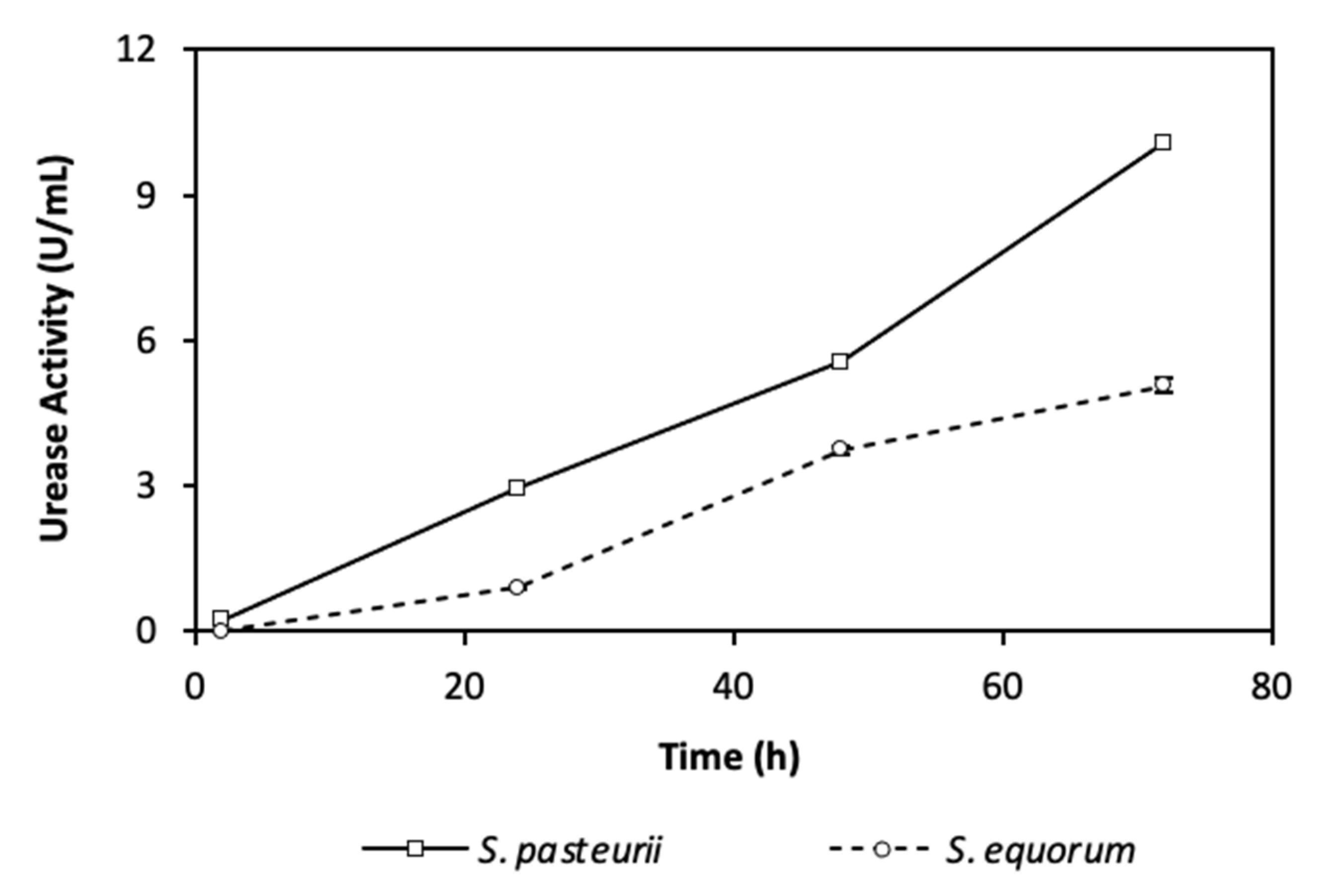
3.2. Ureolytic Activity of S. equorum
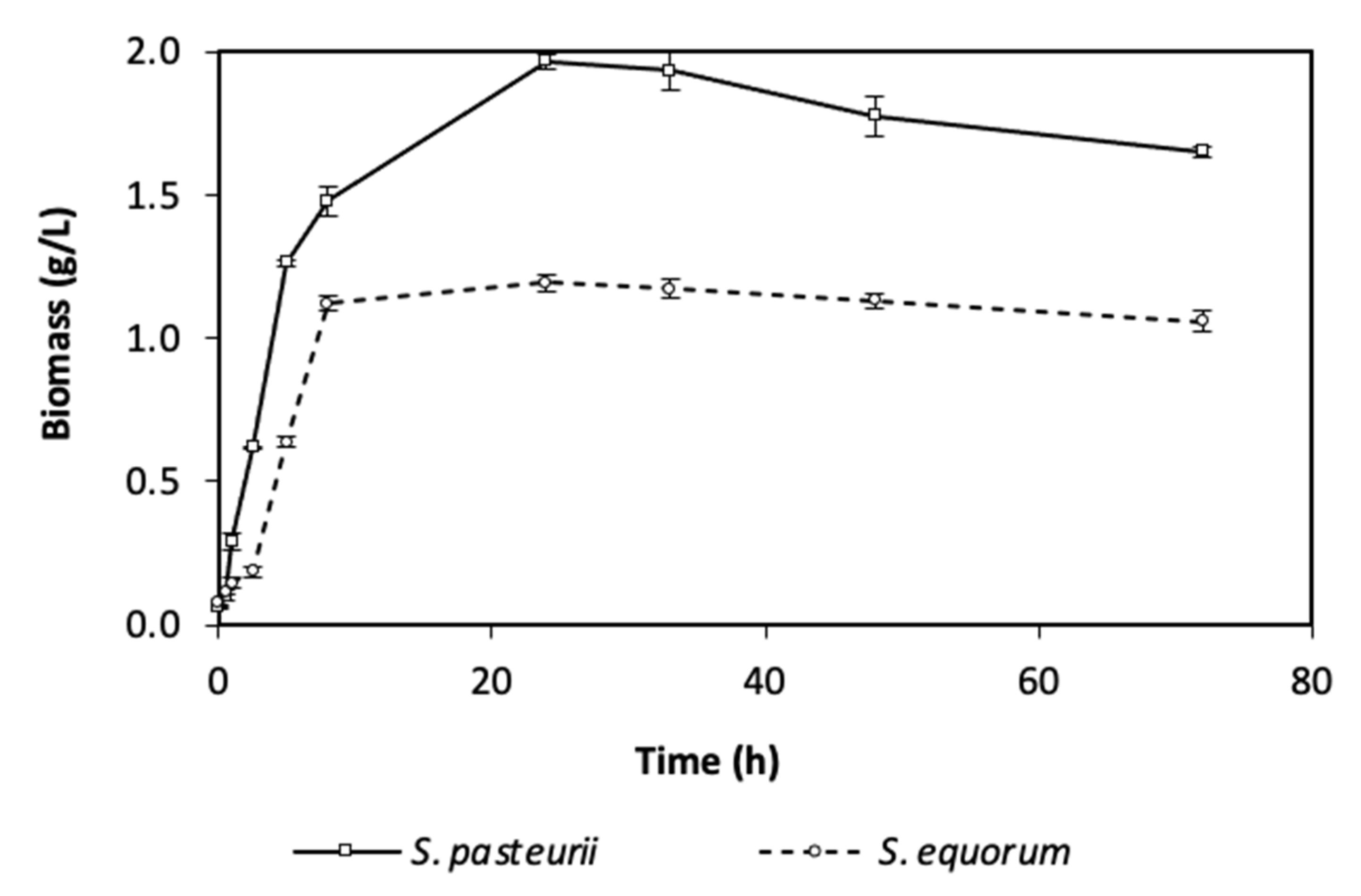
3.2.1. Bacteria Growth
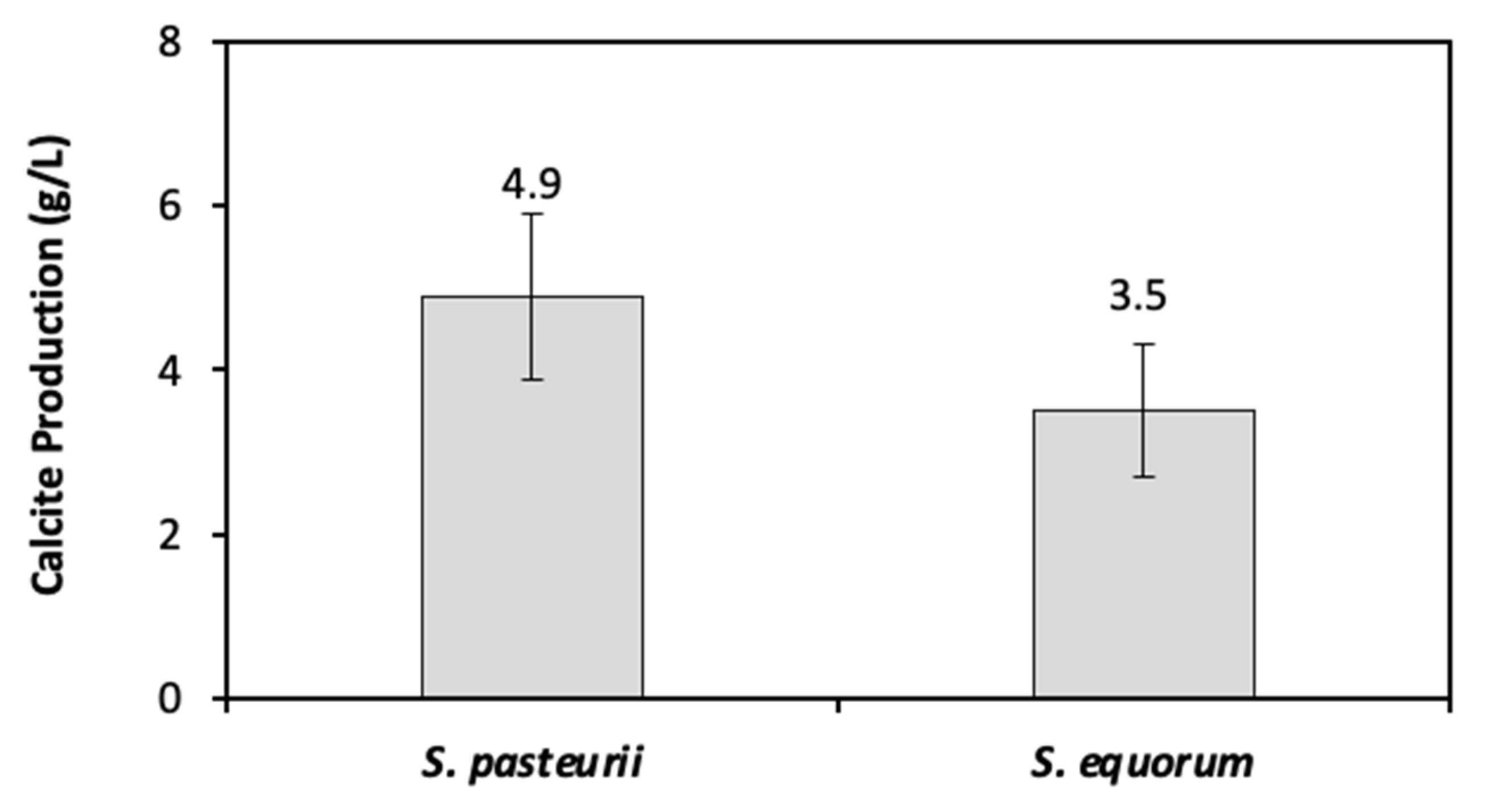
3.2.2. CaCO3 Precipitation
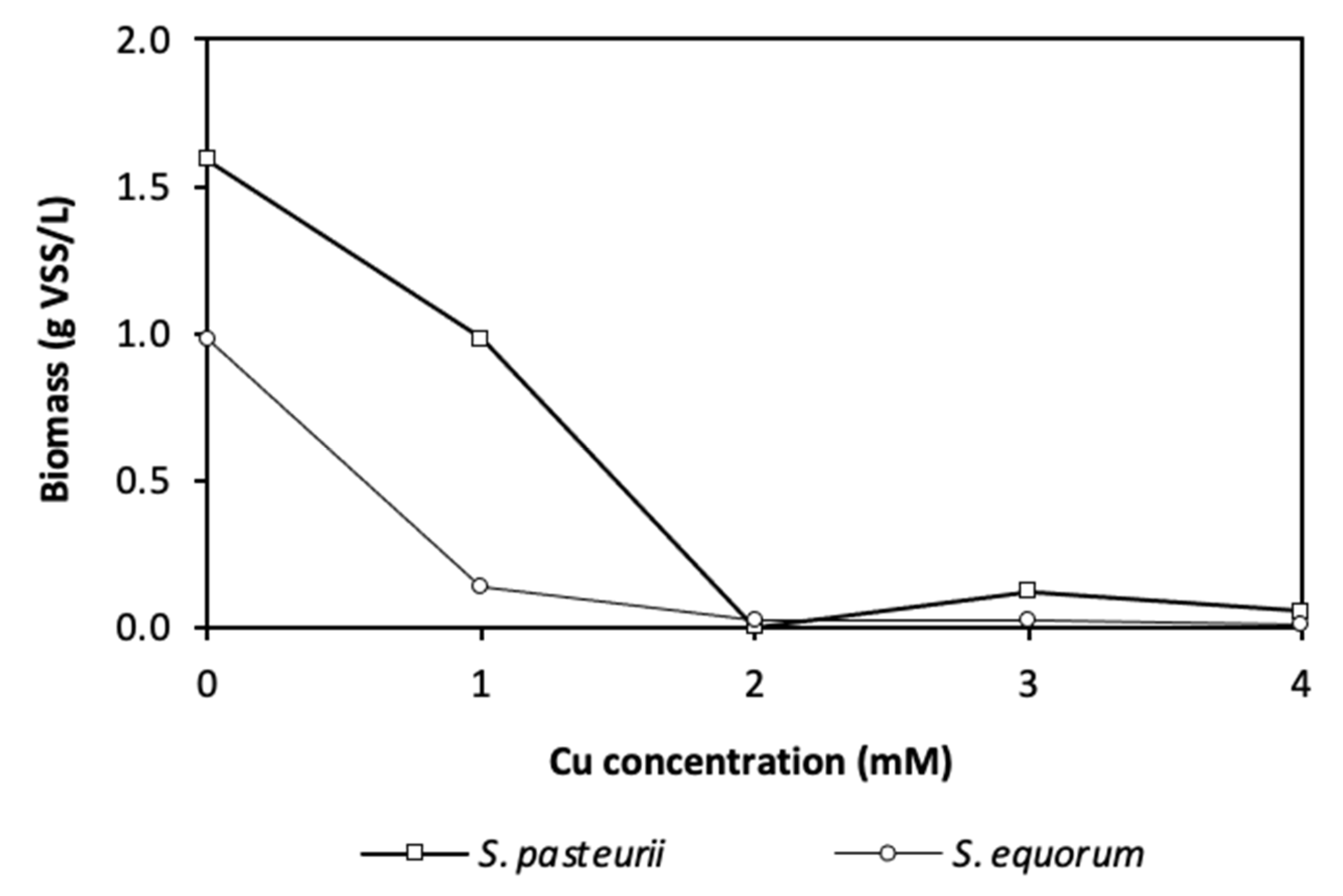
3.2.3. Copper Tolerance
3.2.4. Copper Removal
4. Discussion: Potential Application in Mining Operations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herrera-León, S.; Lucay, F.A.; Cisternas, L.A.; Kraslawski, A. Applying a multi-objective optimization approach in designing water supply systems for mining industries. The case of Chile. J. Clean. Prod. 2019, 210, 994–1004. [Google Scholar] [CrossRef]
- Le, T.M.K.; Mäkelä, M.; Schreithofer, N.; Dahl, O. A multivariate approach for evaluation and monitoring of water quality in mining and minerals processing industry. Miner. Eng. 2020, 157, 106582. [Google Scholar] [CrossRef]
- Jeldres, R.I.; Uribe, L.; Cisternas, L.A.; Gutierrez, L.; Leiva, W.H.; Valenzuela, J. The effect of clay minerals on the process of flotation of copper ores—A critical review. Appl. Clay Sci. 2019, 170, 57–69. [Google Scholar] [CrossRef]
- Zhu, G.; Wu, X.; Ge, J.; Liu, F.; Zhao, W.; Wu, C. Influence of mining activities on groundwater hydrochemistry and heavy metal migration using a self-organizing map (SOM). J. Clean. Prod. 2020, 257, 120664. [Google Scholar] [CrossRef]
- Castro, S.; Uribe, L.; Laskowski, J.S. Depression of inherently hydrophobic minerals by hydrolyzable metal cations: Molybdenite depression in seawater. In Proceedings of the IMPC 2014—27th International Mineral Processing Congress, Santiago, Chile, 20–24 October 2014; pp. 1–11. [Google Scholar]
- Kinnunen, P.; Obenaus-Emler, R.; Raatikainen, J.; Guignot, S.; Guimerà, J.; Ciroth, A.; Heiskanen, K. Review of closed water loops with ore sorting and tailings valorization for a more sustainable mining industry. J. Clean. Prod. 2020, 278, 123237. [Google Scholar] [CrossRef]
- Ramos, J.J.; Leiva, W.H.; Castillo, C.N.; Ihle, C.F.; Fawell, P.D.; Jeldres, R.I. Seawater flocculation of clay-based mining tailings: Impact of calcium and magnesium precipitation. Miner. Eng. 2020, 154, 106417. [Google Scholar] [CrossRef]
- Jeldres, R.I.; Arancibia-Bravo, M.P.; Reyes, A.; Aguirre, C.E.; Cortes, L.; Cisternas, L.A. The impact of seawater with calcium and magnesium removal for the flotation of copper-molybdenum sulphide ores. Miner. Eng. 2017, 109, 10–13. [Google Scholar] [CrossRef]
- Zanin, M.; Lambert, H.; du Plessis, C.A. Lime use and functionality in sulphide mineral flotation: A review. Miner. Eng. 2019, 143, 105922. [Google Scholar] [CrossRef]
- Torres-Aravena, Á.; Duarte-Nass, C.; Azócar, L.; Mella-Herrera, R.; Rivas, M.; Jeison, D. Can microbially induced calcite precipitation (MICP) through a ureolytic pathway be successfully applied for removing heavy metals from wastewaters? Crystals 2018, 8, 438. [Google Scholar] [CrossRef] [Green Version]
- Arias, D.; Cisternas, L.; Rivas, M. Biomineralization mediated by ureolytic bacteria applied to water treatment: A review. Crystals 2017, 7, 345. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.J.; Gerlach, R.; Lauchnor, E.; Mitchell, A.C.; Cunningham, A.B.; Spangler, L. Engineered applications of ureolytic biomineralization: A review. Biofouling 2013, 29, 715–733. [Google Scholar] [CrossRef] [Green Version]
- Krajewska, B. Urease-aided calcium carbonate mineralization for engineering applications: A review. J. Adv. Res. 2018, 13, 59–67. [Google Scholar] [CrossRef]
- Dhami, N.K.; Reddy, M.S.; Mukherjee, M.S. Biomineralization of calcium carbonates and their engineered applications: A review. Front. Microbiol. 2013, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kumari, D.; Qian, X.-Y.; Pan, X.; Achal, V.; Li, Q.; Gadd, G.M. Microbially-induced carbonate precipitation for immobilization of toxic metals. Adv. Appl. Microbiol. 2016, 94, 79–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Liu, Q.; Yu, L.; Subhonqulov, S.H. The interaction mechanism of Fe3+ and NH4+ on chalcopyrite surface and its response to flotation separation of chalcopyrite from arsenopyrite. Sep. Purif. Technol. 2021, 256, 117778. [Google Scholar] [CrossRef]
- Anbu, P.; Kang, C.H.; Shin, Y.J.; So, J.S. Formations of calcium carbonate minerals by bacteria and its multiple applications. Springerplus 2016, 5, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, C.H.; Oh, S.J.; Shin, Y.; Han, S.H.; Nam, I.H.; So, J.S. Bioremediation of lead by ureolytic bacteria isolated from soil at abandoned metal mines in South Korea. Ecol. Eng. 2015, 74, 402–407. [Google Scholar] [CrossRef]
- Kang, C.H.; Han, S.H.; Shin, Y.; Oh, S.J.; So, J.S. Bioremediation of Cd by microbially induced calcite precipitation. Appl. Biochem. Biotechnol. 2014, 172, 2907–2915. [Google Scholar] [CrossRef] [PubMed]
- Achal, V.; Pan, X.; Fu, Q.; Zhang, D. Biomineralization based remediation of As(III) contaminated soil by Sporosarcina ginsengisoli. J. Hazard. Mater. 2012, 201–202, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E. Biosequestration of heavy metals by microbially induced calcite precipitation of ureolytic bacteria. Rom. Biotechnol. Lett. 2019, 24, 147–153. [Google Scholar] [CrossRef]
- Achal, V.; Pan, X.; Zhang, D. Remediation of copper-contaminated soil by Kocuria flava CR1, based on microbially induced calcite precipitation. Ecol. Eng. 2011, 37, 1601–1605. [Google Scholar] [CrossRef]
- Yang, J.; Pan, X.; Zhao, C.; Mou, S.; Achal, V.; Al-Misned, F.A.; Mortuza, M.G.; Gadd, G.M. Bioimmobilization of heavy metals in acidic copper mine tailings soil. Geomicrobiol. J. 2016, 33, 261–266. [Google Scholar] [CrossRef]
- Li, M.; Cheng, X.; Guo, H. Heavy metal removal by biomineralization of urease producing bacteria isolated from soil. Int. Biodeterior. Biodegrad. 2013, 76, 81–85. [Google Scholar] [CrossRef]
- Dharmakeerthi, R.S.; Thenabadu, M.W. Urease activity in soils: A review. J. Natl. Sci. Counc. Sri Lanka 1996, 24, 159–195. [Google Scholar] [CrossRef] [Green Version]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [Green Version]
- Mugwar, A.J.; Harbottle, M.J. Toxicity effects on metal sequestration by microbially-induced carbonate precipitation. J. Hazard. Mater. 2016, 314, 237–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloos, W.E.; George, C.G. Identification of Staphylococcus species and subspecies with the MicroScan Pos ID and Rapid Pos ID panel systems. J. Clin. Microbiol. 1991, 29, 738–744. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Chen, X.; Zhang, Q.; Achal, V. More effective immobilization of divalent lead than hexavalent chromium through carbonate mineralization by Staphylococcus epidermidis HJ2. Int. Biodeterior. Biodegrad. 2019, 140, 67–71. [Google Scholar] [CrossRef]
- Kim, G.; Kim, J.; Youn, H. Effect of temperature, pH, and reaction duration on microbially induced calcite precipitation. Appl. Sci. 2018, 8, 1277. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000; ISBN 0-19-513585-7. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Achal, V.; Mukherjee, A.; Basu, P.C.; Reddy, M.S. Strain improvement of Sporosarcina pasteurii for enhanced urease and calcite production. J. Ind. Microbiol. Biotechnol. 2009, 36, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Achal, V.; Mukherjee, A. A review of microbial precipitation for sustainable construction. Constr. Build. Mater. 2015. [Google Scholar] [CrossRef]
- Kang, C.H.; So, J.S. Heavy metal and antibiotic resistance of ureolytic bacteria and their immobilization of heavy metals. Ecol. Eng. 2016, 97, 304–312. [Google Scholar] [CrossRef]
- Gutiérrez-Arnillas, E.; Arellano, M.; Deive, F.J.; Rodríguez, A.; Sanromán, M.Á. Unravelling the suitability of biological induction for halophilic lipase production by Halomonas sp. LM1C cultures. Bioresour. Technol. 2017, 239, 368–377. [Google Scholar] [CrossRef]
- Stocks-Fischer, S.; Galinat, J.K.; Bang, S.S. Microbiological precipitation of CaCO3. Soil Biol. Biochem. 1999, 31, 1563–1571. [Google Scholar] [CrossRef]
- Prabhakaran, P.; Ashraf, M.A.; Aqma, W.S. Microbial stress response to heavy metals in the environment. RSC Adv. 2016, 6, 109862–109877. [Google Scholar] [CrossRef]
- Jiang, N.J.; Liu, R.; Du, Y.J.; Bi, Y.Z. Microbial induced carbonate precipitation for immobilizing Pb contaminants: Toxic effects on bacterial activity and immobilization efficiency. Sci. Total Environ. 2019, 672, 722–731. [Google Scholar] [CrossRef]
- Pasquale, V.; Fiore, S.; Hlayem, D.; Lettino, A.; Huertas, F.J.; Chianese, E.; Dumontet, S. Biomineralization of carbonates induced by the fungi Paecilomyces inflatus and Plectosphaerella cucumerina. Int. Biodeterior. Biodegrad. 2019, 140, 57–66. [Google Scholar] [CrossRef]
- Radford, D.S.; Kihlken, M.A.; Borrelly, G.P.M.; Harwood, C.R.; Le Brun, N.E.; Cavet, J.S. CopZ from Bacillus subtilis interacts in vivo with a copper exporting CPx-type ATPase CopA. FEMS Microbiol. Lett. 2003, 220, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Majzlik, P.; Strasky, A.; Adam, V.; Nemec, M.; Trnkova, L.; Zehnalek, J.; Hubalek, J.; Provaznik, I.; Kizek, R. Influence of zinc(II) and copper(II) ions on Streptomyces bacteria revealed by electrochemistry. Int. J. Electrochem. Sci. 2011, 6, 2171–2191. [Google Scholar] [CrossRef] [Green Version]
- Altimira, F.; Yáñez, C.; Bravo, G.; González, M.; Rojas, L.A.; Seeger, M. Characterization of copper-resistant bacteria and bacterial communities from copper-polluted agricultural soils of central Chile. BMC Microbiol. 2012, 12, 193. [Google Scholar] [CrossRef] [Green Version]
- Kurmaç, Y. The impact of toxicity of metals on the activity of ureolytic mixed culture during the precipitation of calcium. J. Hazard. Mater. 2009, 163, 1063–1067. [Google Scholar] [CrossRef]
- Duarte-Nass, C.; Rebolledo, K.; Valenzuela, T.; Kopp, M.; Jeison, D.; Rivas, M.; Azócar, L.; Torres-Aravena, Á.; Ciudad, G. Application of microbe-induced carbonate precipitation for copper removal from copper-enriched waters: Challenges to future industrial application. J. Environ. Manag. 2020, 256, 109938. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kwon, S.; Roh, Y. Effect of divalent cations (Cu, Zn, Pb, Cd, and Sr) on microbially induced calcium carbonate precipitation and mineralogical properties. Front. Microbiol. 2021, 12, 763. [Google Scholar]
- Liu, F.; Zhou, K.; Chen, Q.; Wang, A.; Chen, W. Application of magnetic ferrite nanoparticles for removal of Cu(II) from copper-ammonia wastewater. J. Alloys Compd. 2019, 773, 140–149. [Google Scholar] [CrossRef]
- Sen, S.K.; Raut, S.; Dora, T.K.; Mohapatra, P.K. Das Contribution of hot spring bacterial consortium in cadmium and lead bioremediation through quadratic programming model. J. Hazard. Mater. 2014, 265, 47–60. [Google Scholar] [CrossRef]
- Wang, J.; Chen, C. Biosorbents for heavy metals removal and their future. Biotechnol. Adv. 2009, 27, 195–226. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Wang, Q.; Lv, M.; Chen, L. Microorganism remediation strategies towards heavy metals. Chem. Eng. J. 2019, 360, 1553–1563. [Google Scholar] [CrossRef]
- Landeta, G.; Reverón, I.; Carrascosa, A.V.; de las Rivas, B.; Muñoz, R. Use of recA gene sequence analysis for the identification of Staphylococcus equorum strains predominant on dry-cured hams. Food Microbiol. 2011, 28, 1205–1210. [Google Scholar] [CrossRef] [Green Version]
- Jeong, D.W.; Na, H.; Ryu, S.; Lee, J.H. Complete genome sequence of Staphylococcus equorum KS1039 isolated from Saeu-jeotgal, Korean high-salt-fermented seafood. J. Biotechnol. 2016, 219, 88–89. [Google Scholar] [CrossRef]
- Bockelmann, W.; Koslowsky, M.; Goerges, S.; Scherer, S.; Franz, C.M.A.P.; Heller, K.J. Growth inhibition of Listeria monocytogenes by bacteriocin-producing Staphylococcus equorum SE3 in cheese models. Food Control 2017, 71, 50–56. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Closest Species in BLASTN | E Value | Coverage (%) | Identity (%) | Homolog GenBank Accession Number |
|---|---|---|---|---|---|
| A6 | Staphylococcus equorum strain PL453 | 0.0 | 100 | 99.52 | MK015791.1 |
| A11 | Staphylococcus equorum strain PL453 | 0.0 | 99 | 99.32 | MK015791.1 |
| A22 | Staphylococcus equorum strain PL453 | 0.0 | 99 | 99.73 | MK015791.1 |
| Samples | Elemental Composition (%) | |||
|---|---|---|---|---|
| C | O | Ca | Other | |
| S. pasteurii | 7.95 | 29.72 | 51.61 | 10.72 |
| S. equorum | 9.65 | 34.94 | 43.63 | 11.78 |
| CaCO3 | 12.00 | 48.00 | 40.00 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepúlveda, S.; Duarte-Nass, C.; Rivas, M.; Azócar, L.; Ramírez, A.; Toledo-Alarcón, J.; Gutiérrez, L.; Jeison, D.; Torres-Aravena, Á. Testing the Capacity of Staphylococcus equorum for Calcium and Copper Removal through MICP Process. Minerals 2021, 11, 905. https://doi.org/10.3390/min11080905
Sepúlveda S, Duarte-Nass C, Rivas M, Azócar L, Ramírez A, Toledo-Alarcón J, Gutiérrez L, Jeison D, Torres-Aravena Á. Testing the Capacity of Staphylococcus equorum for Calcium and Copper Removal through MICP Process. Minerals. 2021; 11(8):905. https://doi.org/10.3390/min11080905
Chicago/Turabian StyleSepúlveda, Sebastián, Carla Duarte-Nass, Mariella Rivas, Laura Azócar, Andrés Ramírez, Javiera Toledo-Alarcón, Leopoldo Gutiérrez, David Jeison, and Álvaro Torres-Aravena. 2021. "Testing the Capacity of Staphylococcus equorum for Calcium and Copper Removal through MICP Process" Minerals 11, no. 8: 905. https://doi.org/10.3390/min11080905