Biominerals Fossilisation: Fish Bone Diagenesis in Plio–Pleistocene African Hominid Sites of Malawi

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
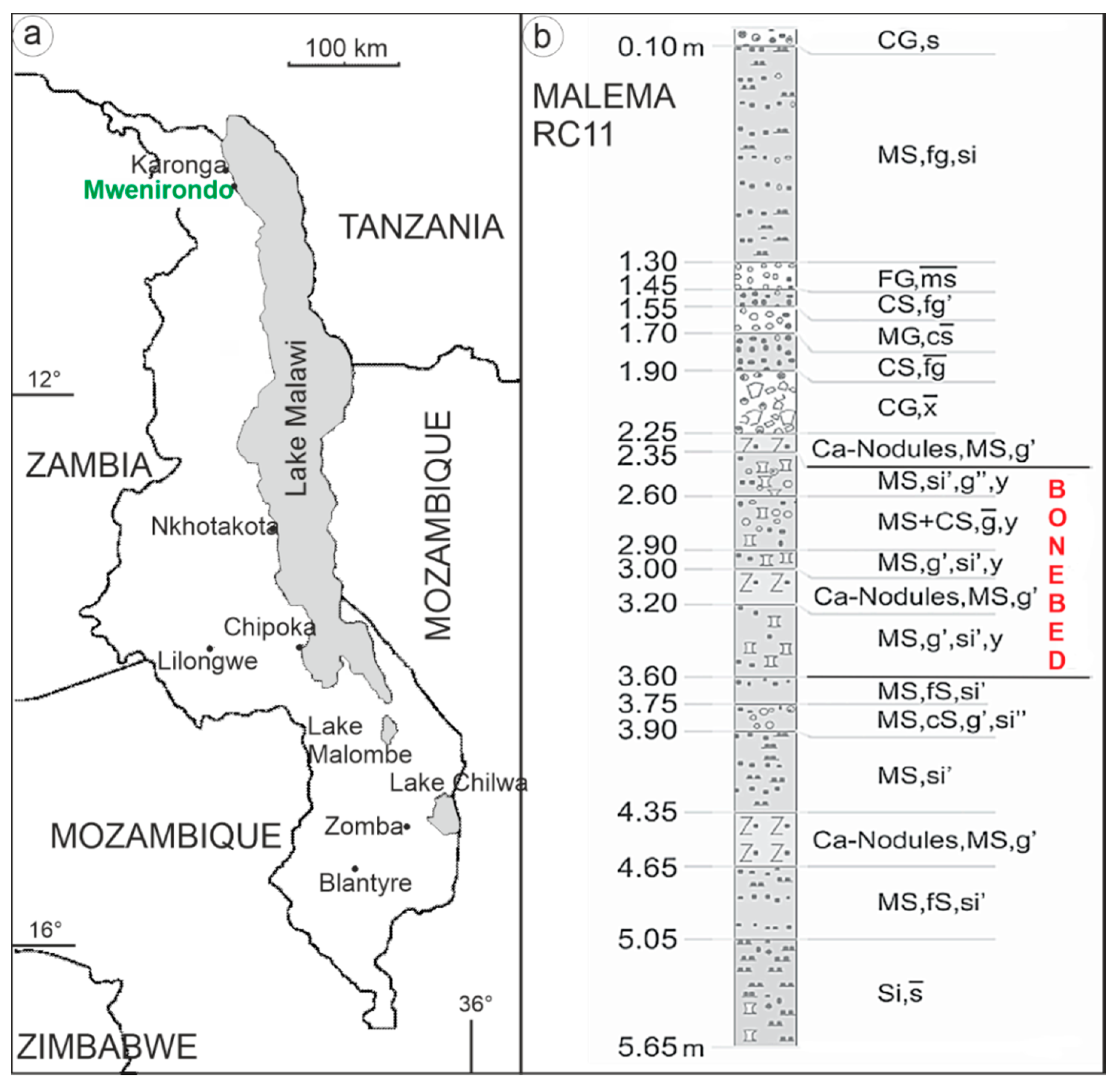
2.1. Materials
2.2. Methods
2.2.1. Sample Preparation
2.2.2. Fish Taxonomic Identifications
2.2.3. Scanning Electron Microscope (SEM)
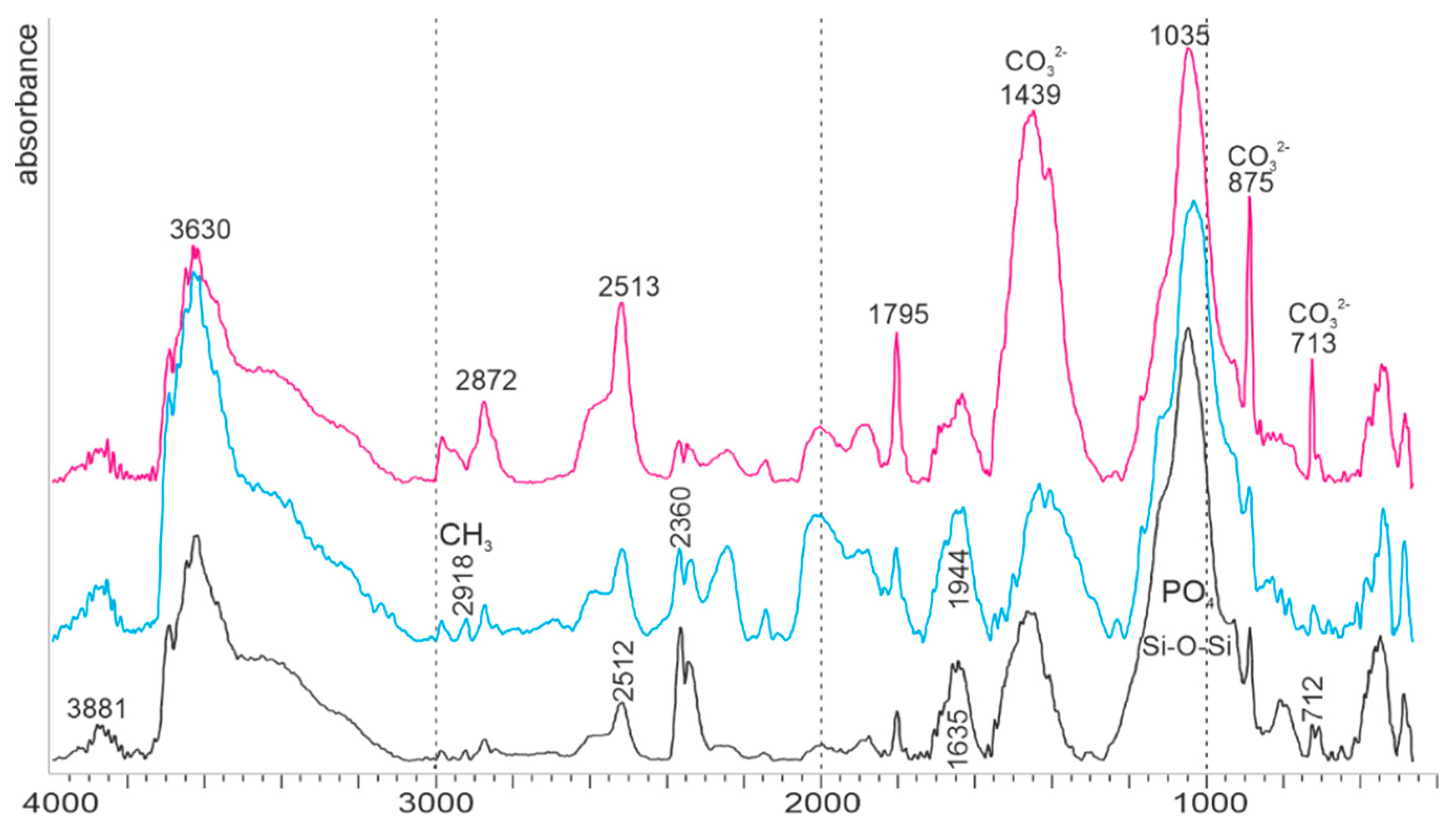
2.2.4. Fourier-Transform Infrared Spectroscopy (FTIR)
2.2.5. Chemical Analyses
2.2.6. Statistical Methods
3. Results
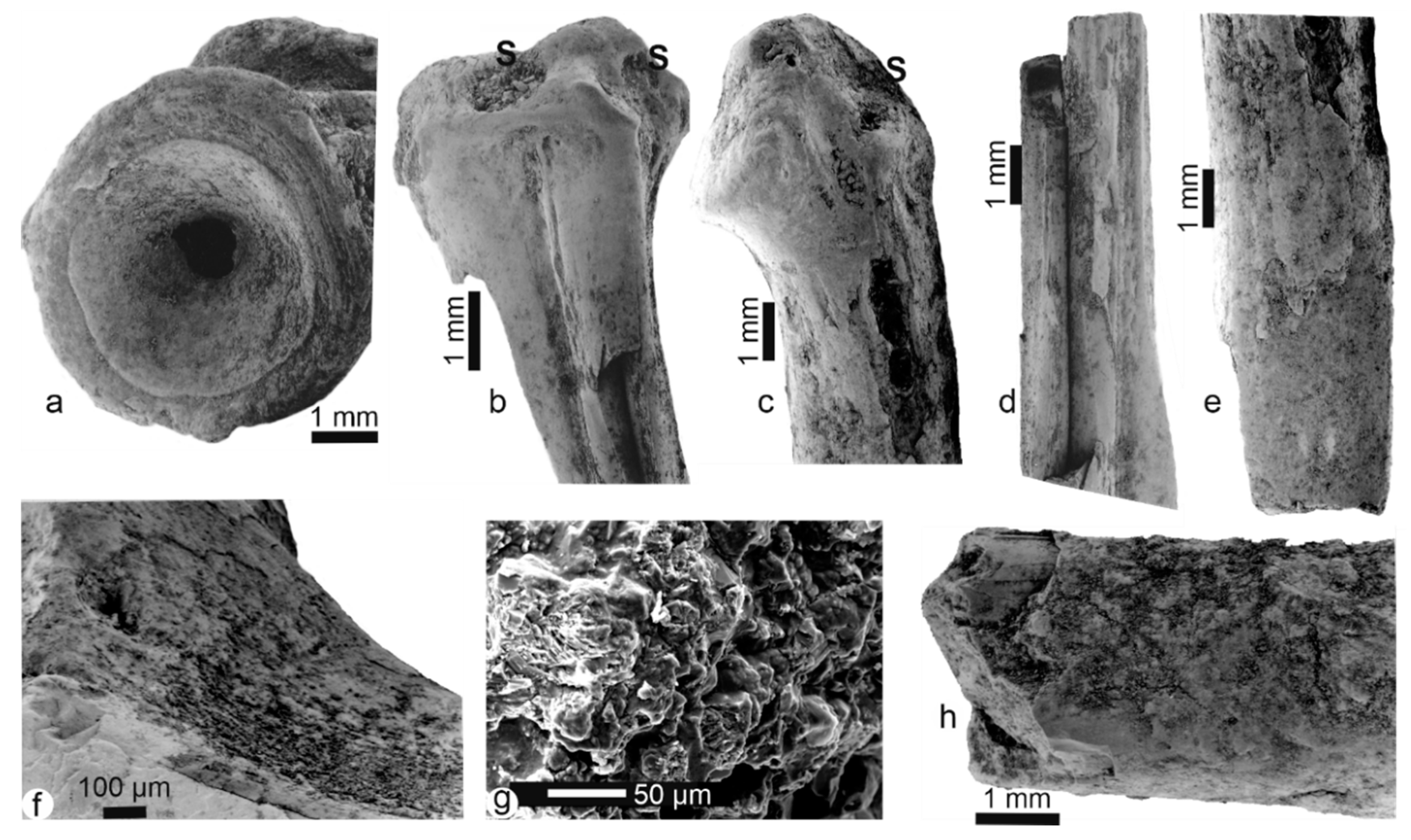
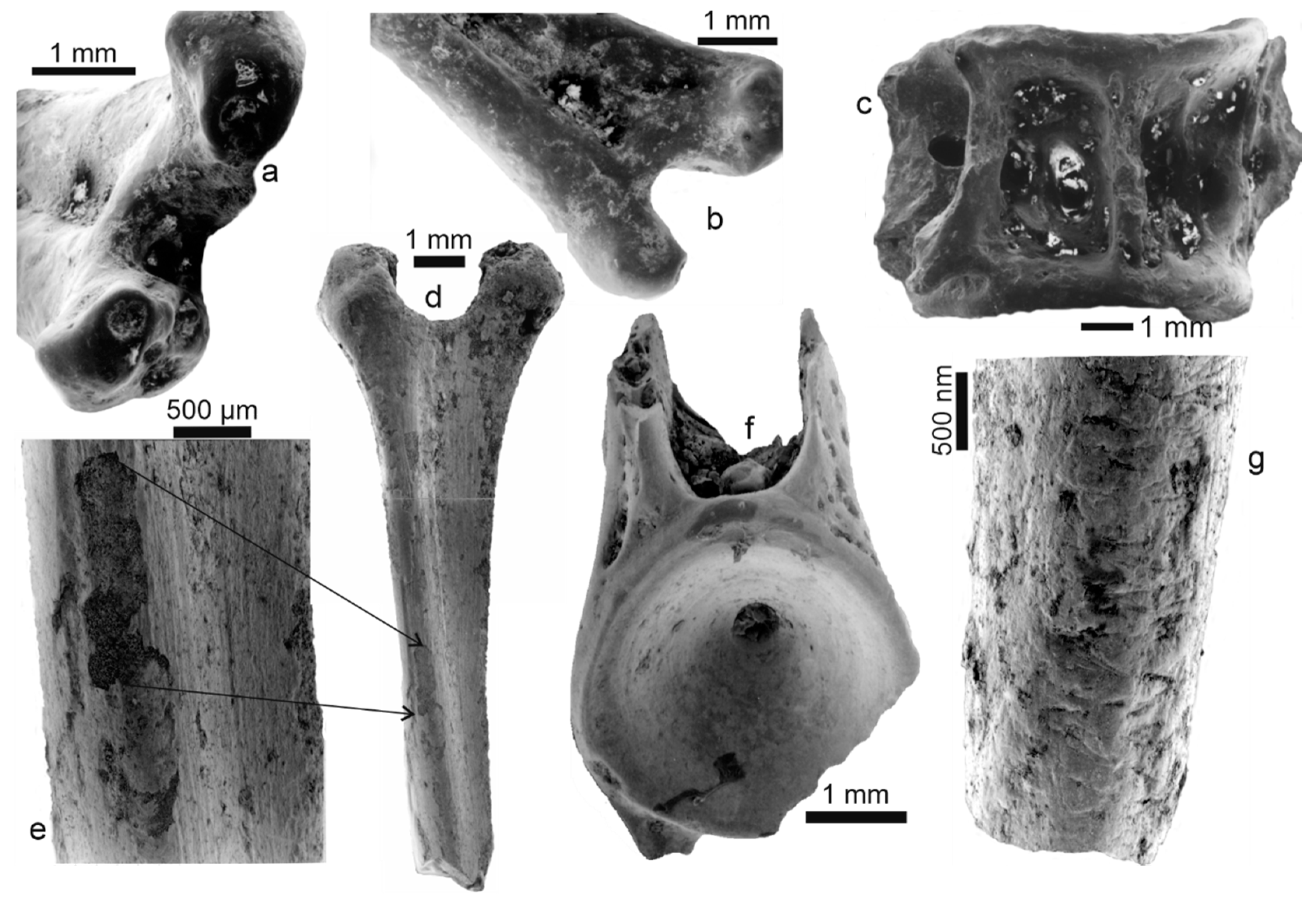
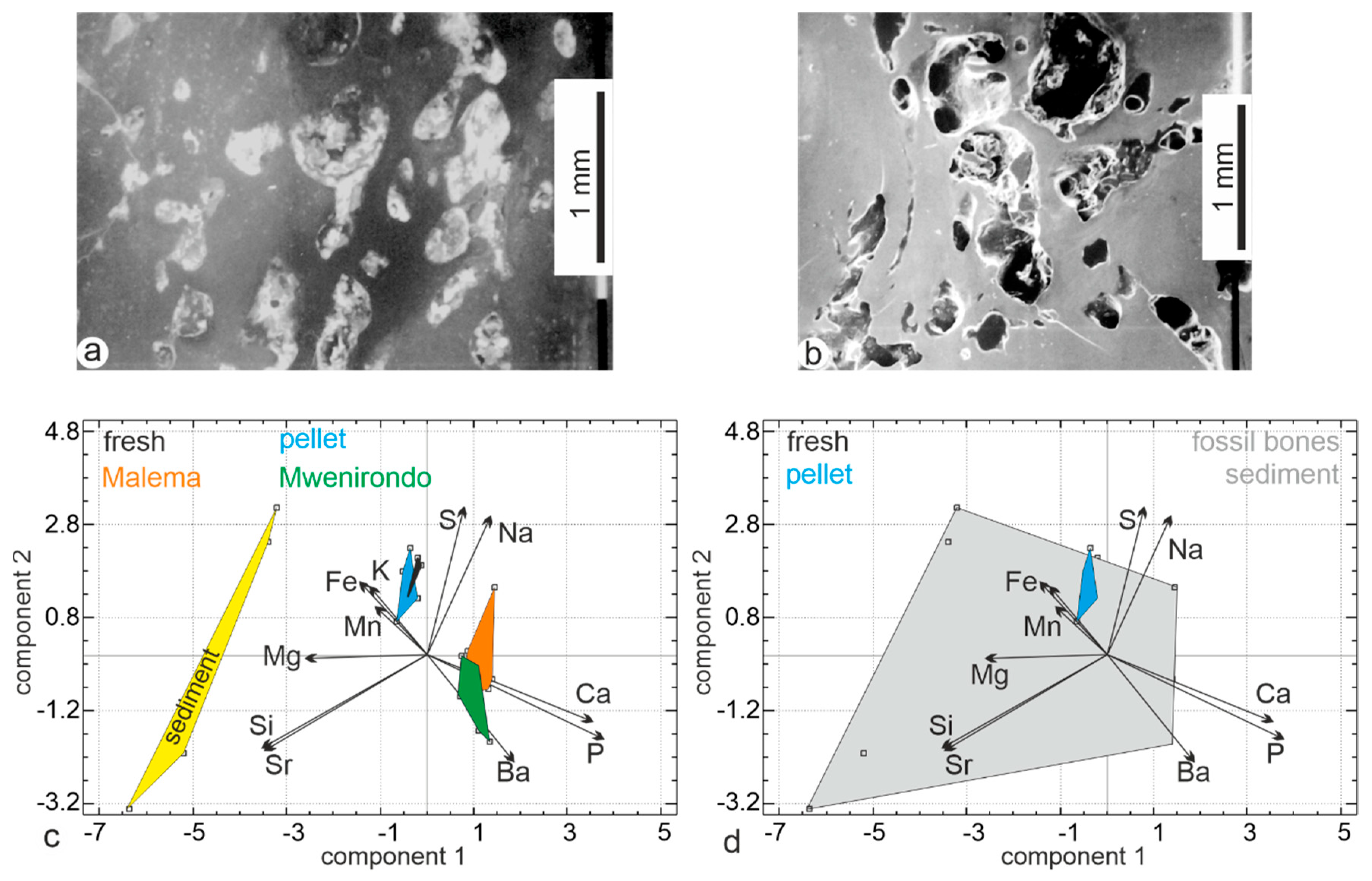
3.1. Bone Surface Preservation and Microstructures
3.1.1. Modern Fresh Bones
3.1.2. Fish-Eagle Pellets
3.1.3. Fossil Bones
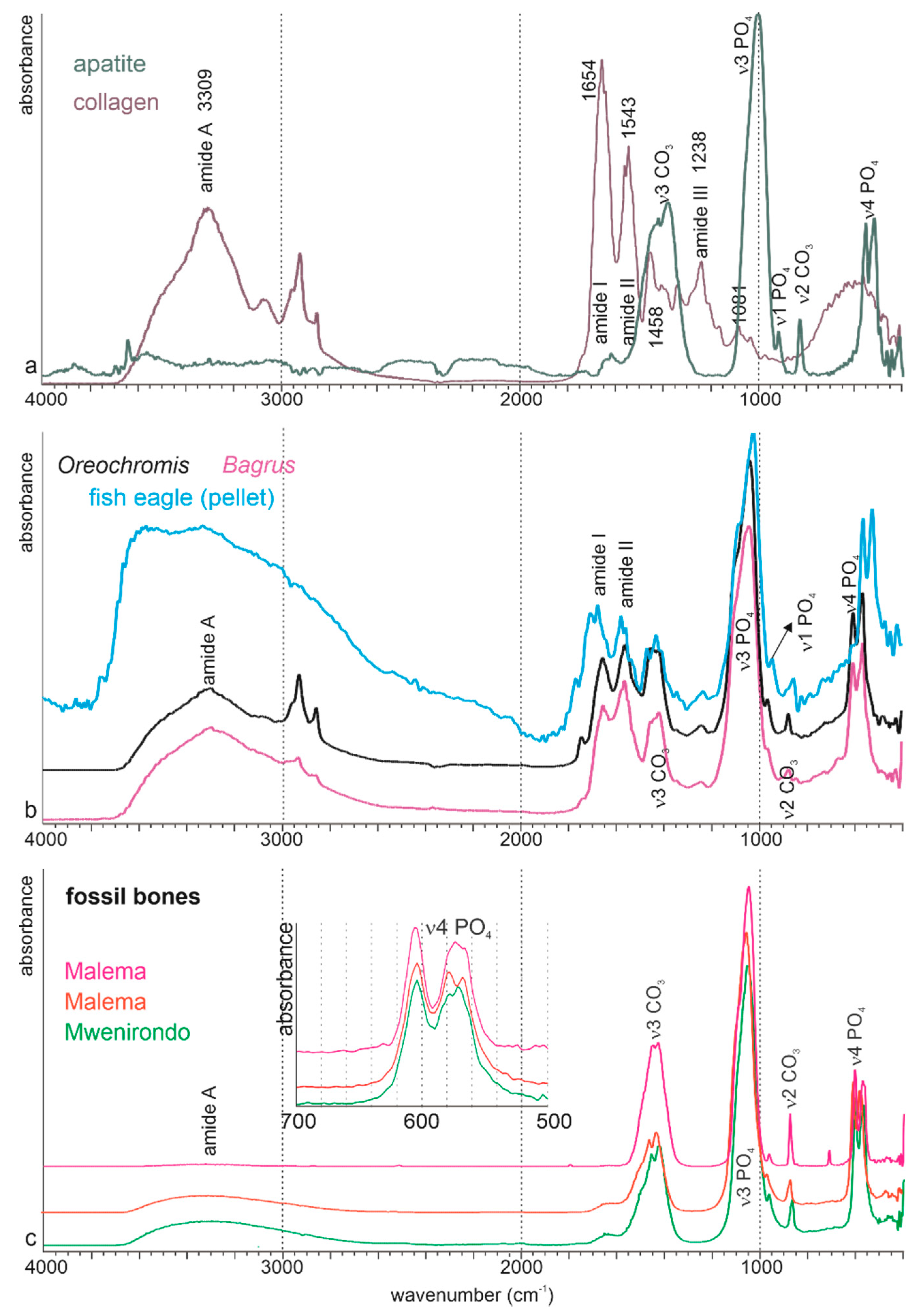
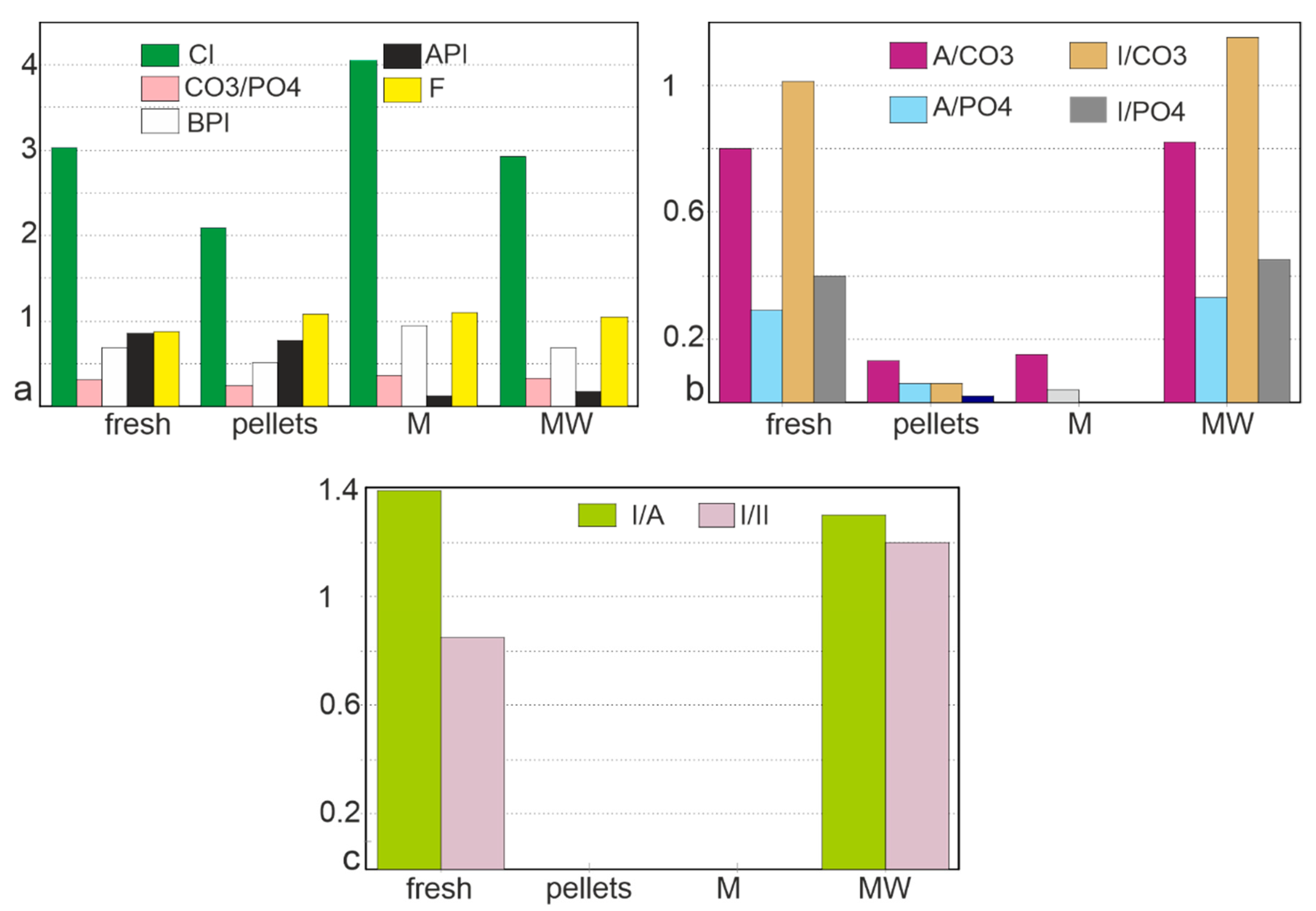
3.2. Chemical Composition
3.2.1. Bulk Composition
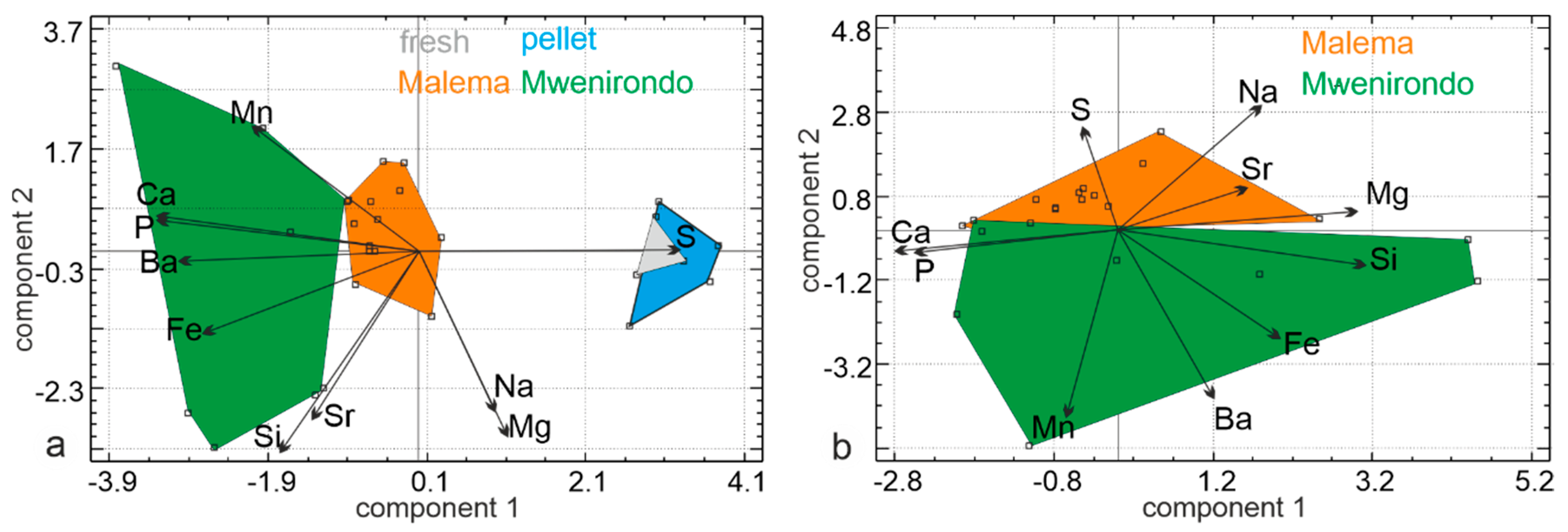
3.2.2. Elemental Composition
4. Discussion
4.1. Comparison of Fresh Bones and Bones Extracted From Regurgitation Pellets
4.1.1. Surface and Microstructure
4.1.2. Mineralogy, Composition
4.2. Comparison of Modern and Fossil Fish Bones
4.2.1. Surface and Microstructure
4.2.2. Mineralogy and Composition
4.3. Comparison of the Fossil Bones From Malema and Mwenirondo
4.4. Fossil Bones and Sediment
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dauphin, Y.; Denys, C.; Denis, A. Les mécanismes de formation des gisements de microvertébrés: Modifications de la composition chimique des os et dents de rongeurs issus de pelotes de régurgitation de rapaces. C. R. Acad. Sci. Paris 1988, 307, 603–608. [Google Scholar]
- Dauphin, Y.; Denys, C.; Denis, A. Les mécanismes de formation des gisements de microvertébrés. 2- Composition chimique élémentaire des os et dents de rongeurs provenant de pelotes de régurgitation de rapaces. Bull. Muséum Natl. Hist. Nat. 1989, 11, 211–237. [Google Scholar]
- Dauphin, Y.; Andrews, P.J.; Denys, C.; Fernandez-Jalvo, Y.; Williams, C.T. Digestion and bone modification in a modern pellet assemblage of Olduvai gorge, Tanzania. J. Taphon. 2003, 1, 209–232. [Google Scholar]
- Denys, C.; Stoetzel, E.; Andrews, P.; Bailon, S.; Rihane, A.; Huchet, J.B.; Fernandez-Jalvo, Y.; Laroulandie, V. Taphonomy of small predators multi-taxa accumulations: Palaeoecological implications. Histor. Biol. 2018, 30, 868–881. [Google Scholar] [CrossRef]
- Dauphin, Y.; Castillo-Michel, H.; Farre, B.; Mataame, A.; Rbii, K.; Rihane, A.; Stoetzel, E.; Denys, C. Identifying predation on rodent teeth through structure and composition: A case from Morocco. Micron 2015, 75, 34–44. [Google Scholar] [CrossRef]
- Denys, C.; Williams, T.; Dauphin, Y.; Andrews, P.; Fernandez-Jalvo, Y. Diagenetical changes in Pleistocene small mammal bones from Olduvai Bed I. Palaeogeogr. Palaeoclim. Palaeoecol. 1996, 126, 121–134. [Google Scholar] [CrossRef]
- Schrenk, F.; Bromage, T.G.; Gorthner, A.; Sandrock, O. Paleoecology of the Malawi Rift: Vertebrate and invertebrate faunal context of the Chiwondo Beds, northern Malawi. J. Human Evol. 1995, 29, 59–70. [Google Scholar] [CrossRef]
- Sandrock, O.; Kullmer, O.; Schrenk, F.; Juwayeyi, Y.M.; Bromage, T. Fauna, taphonomy, and ecology of the Plio-Pleistocene Chiwondo beds, Northern Malawi. In Hominin Environments in the East African Pliocene: An Assessment of the Faunal Evidence; Bobe, R., Alemseged, Z., Behrensmeyer, A.K., Eds.; Springer: Dordrecht, Germany, 2007; pp. 315–332. [Google Scholar]
- Kullmer, O.; Sandrock, O.; Kupczik, K.; Frost, S.R.; Volpato, V.; Bromage, T.G.; F Schrenk, F. Primate remains from Mwenirondo, Chiwondo Beds in northern Malawi. J. Hum. Evol. 2011, 61, 617–623. [Google Scholar] [CrossRef]
- Sandrock, O. The Taphonomy and Paleo Ecology of the Malema Hominid Site, Northern Malawi. Ph.D. Thesis, Johannes Gutenberg Universität Mainz, Mainz, Germany, 1999; p. 276. [Google Scholar]
- Sandrock, O.; Dauphin, Y.; Kullmer, O.; Abel, R.; Schrenk, F.; Denys, C. Malema: Preliminary taphonomic analysis of an African hominid locality. C. R. Acad. Sci. Paris 1999, 328, 133–139. [Google Scholar] [CrossRef]
- Lüdecke, T.; Kullmer, O.; Wacker, U.; Sandrock, O.; Fiebig, J.; Schrenk, F.; Mulch, M. Dietary versatility of Early Pleistocene hominins. Proc. Natl. Acad. Sci. USA 2018, 115, 13330–13335. [Google Scholar]
- Stewart, K.M.; Murray, A.M. Earliest fish remains from the Lake Malawi Basin, and biogeographical implications. J. Vert. Paleont. 2013, 33, 532–539. [Google Scholar] [CrossRef]
- Stewart, K.M. Fishing sites of North and East Africa in the Late Pleistocene and Holocene: Environmental change and human adaptation. BAR Int. Ser. 1989, 521, 1–292. [Google Scholar]
- Van Neer, W. New Late Tertiary fish fossils from the Sinda Region, Eastern Zaire. Afr. Study Monogr. 1992, 27–47. [Google Scholar] [CrossRef]
- Otero, O.; Pinton, A.; Mackaye, H.T.; Likius, A.; Vignaud, P.; Brunet, M. The fish assemblage associated with the Late Miocene Chadian hominid (site TM266, Toros-Menalla, Western Djurab), and its palaeoenvironmental signification. Palaeontographica 2010, A292, 21–51. [Google Scholar] [CrossRef]
- Gäb, F.; Ballhaus, C.; Stinnesback, E.; Kral, A.G.; Janssen, K.; Bierbaum, G. Experimental taphonomy of fish - rôle of elevated pressure, salinity and pH. Sci. Rep. 2020, 10, 7839. [Google Scholar] [CrossRef]
- Butler, V.L. Natural versus cultural salmonid remains: Origin of the Dalles Roadcut bones, Columbia River, Oregon, USA. J. Archaeol. Sci. 1993, 20, 1–24. [Google Scholar] [CrossRef]
- Jones, A.K.G. Fish bone survival in the digestive systems of the pig, dogs and man: Some experiments. In Fish and Archaeology: Studies in Osteometry, Taphonomy, Seasonality and Fishing Methods; BAR International series 294; Brinkhuizen, D.C., Clason, A.T., Eds.; BAR International: Oxford, UK, 1986; pp. 53–61. [Google Scholar]
- Nicholson, R.A. An investigation into the effects on fish bone passage through the human gut: Some experiments and comparisons with archaeological material. Circaea 1992, 10, 38–50. [Google Scholar]
- Van Neer, W.; Alhaique, F.; Wouters, W.; Dierickx, K.; Gala, M.; Goffette, Q.; Mariani, G.S.; Zerboni, A.; di Lernia, S. Aquatic fauna from the Takarkori rock shelter reveals the Holocene central Saharan climate and palaeohydrography. PLoS ONE 2020, 15, e0228588. [Google Scholar] [CrossRef] [Green Version]
- Stewart, K.M.; Matthiesen, D.P.; Leblanc, L.; West, J. Prey diversity and selectivity of the African fish eagle: Data from a roost in northern Kenya. Afr. J. Ecol. 1997, 35, 133–145. [Google Scholar] [CrossRef]
- Stewart, K.M.; Leblanc, L.; Matthiesen, D.P.; West, J. Microfaunal remains from a modern east African raptor roost: Patterning and implications for fossil bone scatters. Paleobiology 1999, 25, 483–503. [Google Scholar] [CrossRef]
- Guillaud, E.; Bearez, P.; Denys, C.; Raimond, S. New data on fish diet and bone digestion of the Eurasian otter (Lutra lutra) (Mammalia: Mustelidae) in central France. Europ. Zool. J. 2017, 84, 226–237. [Google Scholar] [CrossRef] [Green Version]
- Guillaud, E.; Lebreton, L.; Béarez, P. Taphonomic signature of Eurasian eagle owl (Bubo bubo) on fish remains. Folia Zool. 2018, 67, 143–153. [Google Scholar] [CrossRef]
- Betzler, C.; Ring, U. Sedimentology of the Malawi Rift: Facies and stratigraphy of the Chiwondo Beds, northern Malawi. J. Hum. Evol. 1995, 28, 23–35. [Google Scholar] [CrossRef]
- Brown, F.H.; McDougall, I.; Davies, T.; Maier, R. An integrated Plio-Pleistocene chronology for the Turkana Basin. In Ancestors: The Hard Evidence; Delson, E., Ed.; Alan R. Liss: New York, NY, USA, 1985; pp. 82–90. [Google Scholar]
- Brown, F.H.; Feibel, C.S. Revision of lithostratigraphic nomenclature in the Koobi Fora region, Kenya. J. Geol. Soc. Lond. 1986, 143, 297–310. [Google Scholar] [CrossRef]
- Schrenk, F.; Bromage, T.G.; Betzler, C.G.; Ring, U.; Juwayeyi, Y.M. Oldest Homo and Pliocene biogeography of the Malawi Rift. Nature 1993, 365, 833–836. [Google Scholar] [CrossRef]
- Bromage, T.G.; Schrenk, F.; Juwayeyi, Y.M. Palaeobiogeography of the Malawi Rift: Age and vertebrate paleontology of the Chiwondo Beds, northern Malawi. J. Hum. Evol. 1995, 28, 37–59. [Google Scholar] [CrossRef]
- Kullmer, O. The fossil suidae from the Plio-Pleistocene Chiwondo Beds of northern Malawi, Africa. J. Vert. Paleontol. 2008, 208, 208–216. [Google Scholar] [CrossRef]
- Shemesh, A. Crystallinity and diagenesis of sedimentary apatite. Geochim. Cosmochim. Acta 1990, 545, 2433–2438. [Google Scholar] [CrossRef]
- Weiner, S.; Bar-Yosef, O. States of preservation of bones from prehistoric sites in the Near East: A survey. J. Archaeol. Sci. 1990, 17, 187–196. [Google Scholar] [CrossRef]
- Sillen, A. Biogenic and diagenetic Sr/Ca in Plio-Pleistocene fossils of the Omo Shungura formation. Paleobiology 1986, 12, 311–323. [Google Scholar] [CrossRef]
- LeGeros, R.; Trautz, O.R.; Klein, E.; LeGeros, J.P. Two types of carbonate substitution in the apatite structure. Experientia 1969, 25, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Rey, C.; Renugopalakrishnan, V.; Collins, B.; Glimcher, M.J. Fourier transform infrared spectroscopic study of the carbonate ions in bone mineral during aging. Calcif. Tissue Int. 1991, 46, 251–258. [Google Scholar] [CrossRef]
- Botha, J.; Lee-Thorp, J.; Sponheimer, M. An examination of Triassic cynodont tooth enamel chemistry using Fourier Transform infrared spectroscopy. Calcif. Tissue Int. 2004, 74, 163–169. [Google Scholar] [CrossRef] [PubMed]
- El Feki, H.; Rey, C.; Vignoles, M. Carbonate ions in apatites: Infrared investigations in the v4 CO3 domain. Calcif. Tissue Int. 1991, 49, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Dauphin, Y. Potential of the Diffuse Reflectance Infrared Fourier Transform (DRIFT) method in paleontological studies of bones. Appl. Spectrosc. 1993, 47, 52–55. [Google Scholar] [CrossRef]
- Fernandez-Jalvo, Y.; Andrews, P. Atlas of Taphonomic Identifications: 1001+ Images of Fossil and Recent Mammal Bone Modification; Vertebrate Paleobiology and Paleoanthropology Series; Springer: Dordrecht, Germany, 2016; pp. 1–359. [Google Scholar]
- Fernandez-Jalvo, Y.; Andrews, P. Experimental effects of water abrasion on bone fragments. J. Taphon. 2003, 1, 147–163. [Google Scholar]
- Trueman, C.N.G.; Behrensmayer, A.K.; Tuross, N.; Weiner, S. Mineralogical and compositional changes in bones exposed on soil surfaces in Amboseli National park, Kenya: Diagenetic mechanisms and the role of sediment pore fluids. J. Archaeol. Sci. 2004, 31, 721–739. [Google Scholar] [CrossRef]
- Madejova, J.; Gates, W.P.; Petit, S. IR spectra of clay minerals. In Developments in Clay Science, 8; Gates, W., Kloprogge, J.T., Madejova, J., Bergaya, F., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; p. 620. [Google Scholar] [CrossRef]
- Vaculikova, L.; Plevova, E. Identification of clay minerals and micas in sedimentary rocks. Acta Geodyn. Geomater. 2005, 2, 165–175. [Google Scholar]
- Dauphin, Y.; Kowalski, C.; Denys, C. Assemblage data and bone and teeth modifications as an aid to paleoenvironmental interpretations of the open-air pleistocene site of Tighenif (Algeria). Quat. Res. 1994, 42, 340–342. [Google Scholar] [CrossRef]
- Denys, C.; Kowalski, K.; Dauphin, Y. Mechanical and chemical alterations of skeletal tissues in a recent Saharian accumulation of faeces from Vulpes rueppelli (Carnivora, Mammalia). Acta Zool. Cracov. 1992, 35, 265–283. [Google Scholar]
- Cohen, L.; Dean, M.; Shipov, A.; Atkins, A.; Monsonego-Ornan, E.; Shahar, R. Comparison of structural, architectural and mechanical aspects of cellular and acellular bone in two teleost fish. J. Experim. Biol. 2012, 215, 1983–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, R.A. Fish bone diagenesis in different soils. Archaeofauna 1996, 5, 79–91. [Google Scholar]
- Kölliker, A. On the different types in the microscopic structure of the skeleton of osseous fishes. Proc. R. Soc. Lond. 1857, 9, 656–668. [Google Scholar]
- Meunier, F.J.; Huysseune, A. The concept of bone tissue in Osteichthyes. Neth. J. Zool. 1992, 42, 445–458. [Google Scholar] [CrossRef]
- Currey, J.D.; Shahar, R. Cavities in the compact bone in tetrapods and fish and their effect on mechanical properties. J. Struct. Biol. 2013, 183, 107–122. [Google Scholar] [CrossRef]
- Zohar, I.; Belmaker, M.; Nadel, D.; Gafny, S.; Goren, G.; Hershkovitz, I.; Dayan, T. The living and the dead: How do taphonomic processes modify relative abundance and skeletal completeness of freshwater fish? Palaeogeogr. Palaeocl. Palaeoecol. 2008, 258, 292–316. [Google Scholar] [CrossRef]
- Maccari, F.; Ferrarini, F.; Volpi, N. Structural characterization of chondroitin sulfate from sturgeon bone. Carbohydr. Res. 2010, 145, 1575–1580. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | m/Sd | Na | Mg | Si | S | K | Mn | Fe | Sr | Ba | P | Ca | Ca/P |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fresh | m | 4440 | 1293 | 269 | 2646 | 215 | 132 | 356 | 1629 | 386 | 81,183 | 156,234 | 1.92 |
| n = 3 | sd | 366 | 142 | 148 | 912 | 140 | 98 | 122 | 299 | 51 | 3319 | 6928 | |
| Pellets | m | 3418 | 1559 | 173 | 2825 | 185 | 71 | 369 | 1578 | 200 | 88,210 | 167,708 | 1.90 |
| n = 4 | sd | 998 | 255 | 51 | 350 | 365 | 87 | 195 | 721 | 100 | 9769 | 19,498 | |
| Malema | m | 3621 | 873 | 1419 | 1283 | 117 | 860 | 13,747 | 2267 | 584 | 147,052 | 335,251 | 2.28 |
| n = 14 | sd | 905 | 186 | 2522 | 403 | 192 | 479 | 7268 | 818 | 183 | 5976 | 14,086 | |
| Mwenirondo | m | 3378 | 1001 | 3828 | 845 | 36 | 3478 | 45,552 | 2272 | 905 | 146,434 | 331,852 | 2.27 |
| n = 7 | sd | 777 | 497 | 3596 | 405 | 45 | 4181 | 18,077 | 1445 | 368 | 10626 | 26,590 | |
| Sediment | m | 2943 | 4267 | 179,947 | 565 | 1244 | 1714 | 128,620 | 15,437 | 309 | 46,768 | 88,432 | 1.89 |
| n = 4 | sd | 1681 | 4347 | 185,539 | 606 | 2203 | 2263 | 200,328 | 13,480 | 326 | 45,892 | 113,051 |
| Comparison | Na | Mg | Si | S | K | Mn | Fe | Sr | Ba | P | Ca |
|---|---|---|---|---|---|---|---|---|---|---|---|
| F/P | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS |
| F/M | S | S | NS | S | NS | S | S | NS | S | S | S |
| F/MW | S | NS | S | S | NS | S | S | NS | NS | S | S |
| P/M | NS | S | S | S | NS | S | S | NS | S | S | S |
| P/MW | NS | NS | S | S | NS | S | S | NS | S | S | S |
| M/MW | NS | NS | S | S | NS | S | S | S | NS | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denys, C.; Otero, O.; Kullmer, O.; Sandrock, O.; Bromage, T.G.; Schrenk, F.; Dauphin, Y. Biominerals Fossilisation: Fish Bone Diagenesis in Plio–Pleistocene African Hominid Sites of Malawi. Minerals 2020, 10, 1049. https://doi.org/10.3390/min10121049
Denys C, Otero O, Kullmer O, Sandrock O, Bromage TG, Schrenk F, Dauphin Y. Biominerals Fossilisation: Fish Bone Diagenesis in Plio–Pleistocene African Hominid Sites of Malawi. Minerals. 2020; 10(12):1049. https://doi.org/10.3390/min10121049
Chicago/Turabian StyleDenys, Christiane, Olga Otero, Ottmar Kullmer, Oliver Sandrock, Timothy G. Bromage, Friedemann Schrenk, and Yannicke Dauphin. 2020. "Biominerals Fossilisation: Fish Bone Diagenesis in Plio–Pleistocene African Hominid Sites of Malawi" Minerals 10, no. 12: 1049. https://doi.org/10.3390/min10121049