
The Effects of Vegetation Structure and Timber Harvesting on Ground Beetle (Col.: Carabidae) and Arachnid Communities (Arach.: Araneae, Opiliones) in Short-Rotation Coppices

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Taxonomy and Nomenclature
2.4. Ecological Groups
2.4.1. Ecological Types (ETs)
2.4.2. Habitat Preferences (HPs)
2.5. Species, Habitat Preference, and Vegetation Structure Diversity
2.6. Statistical Analysis
3. Results
3.1. Vegetation Structure
3.2. Species and Individual Numbers
3.3. Characterisation of Alpha Diversity
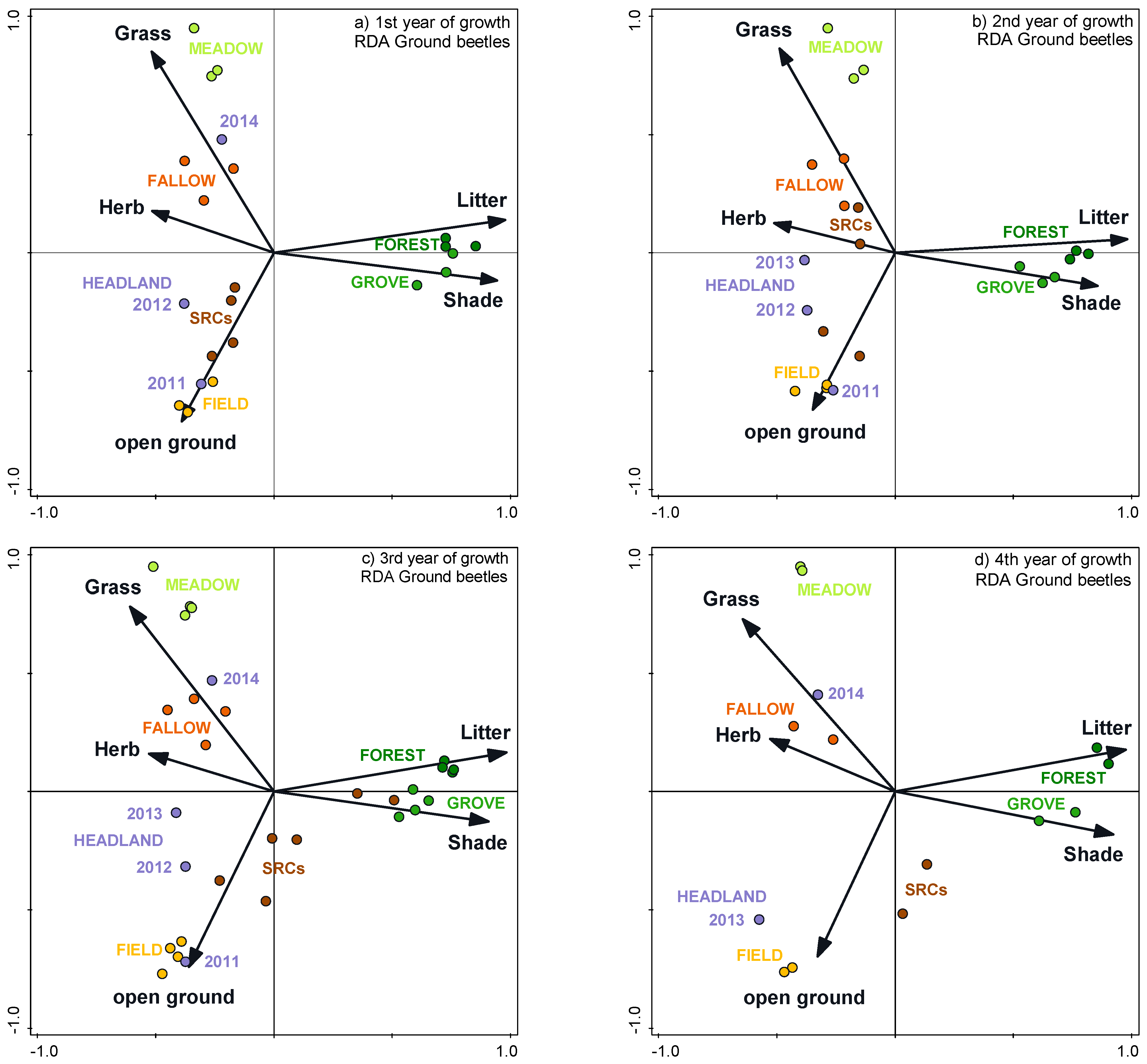
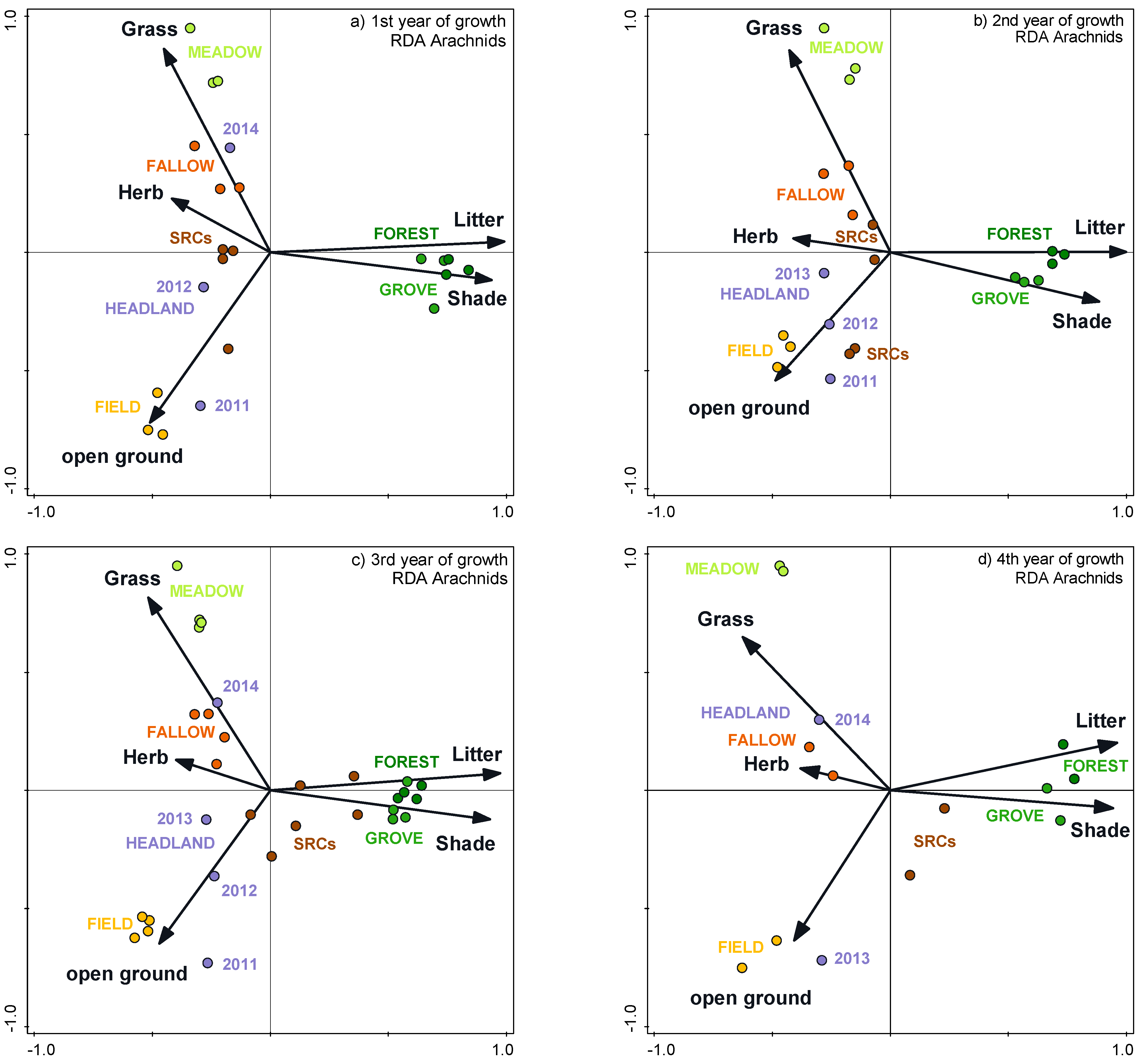
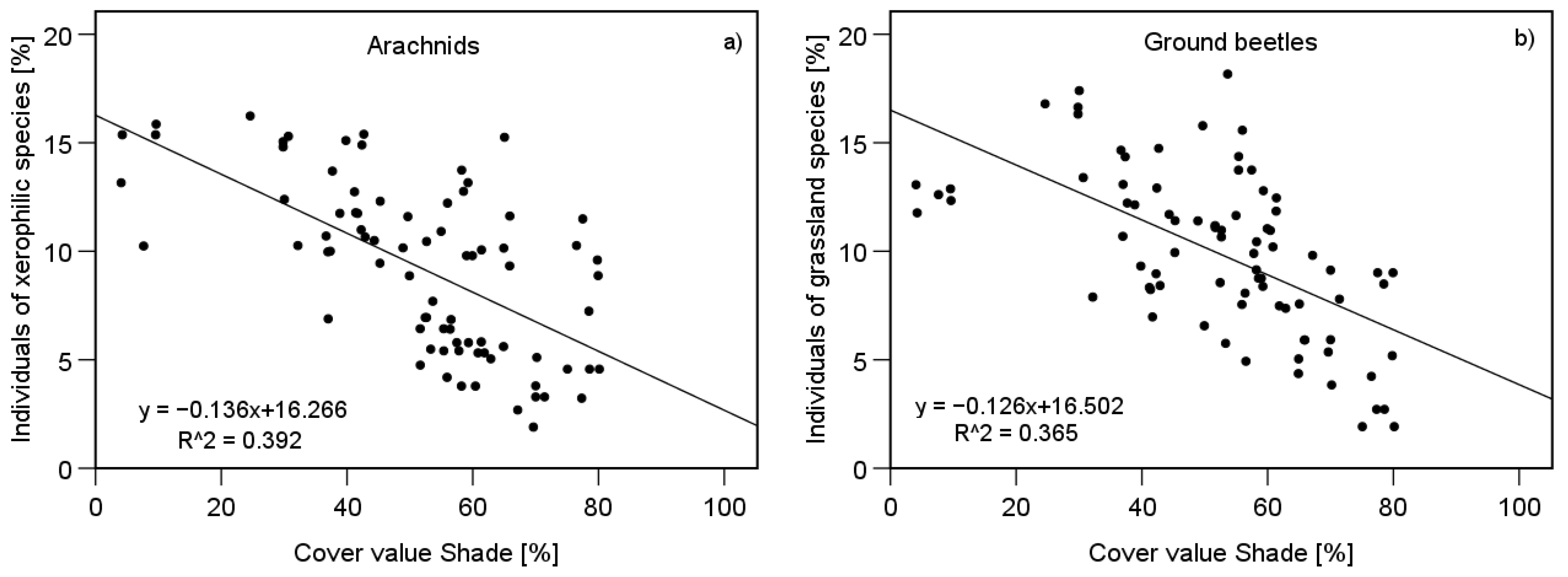
3.4. Impacts of Vegetation Structure on Communities
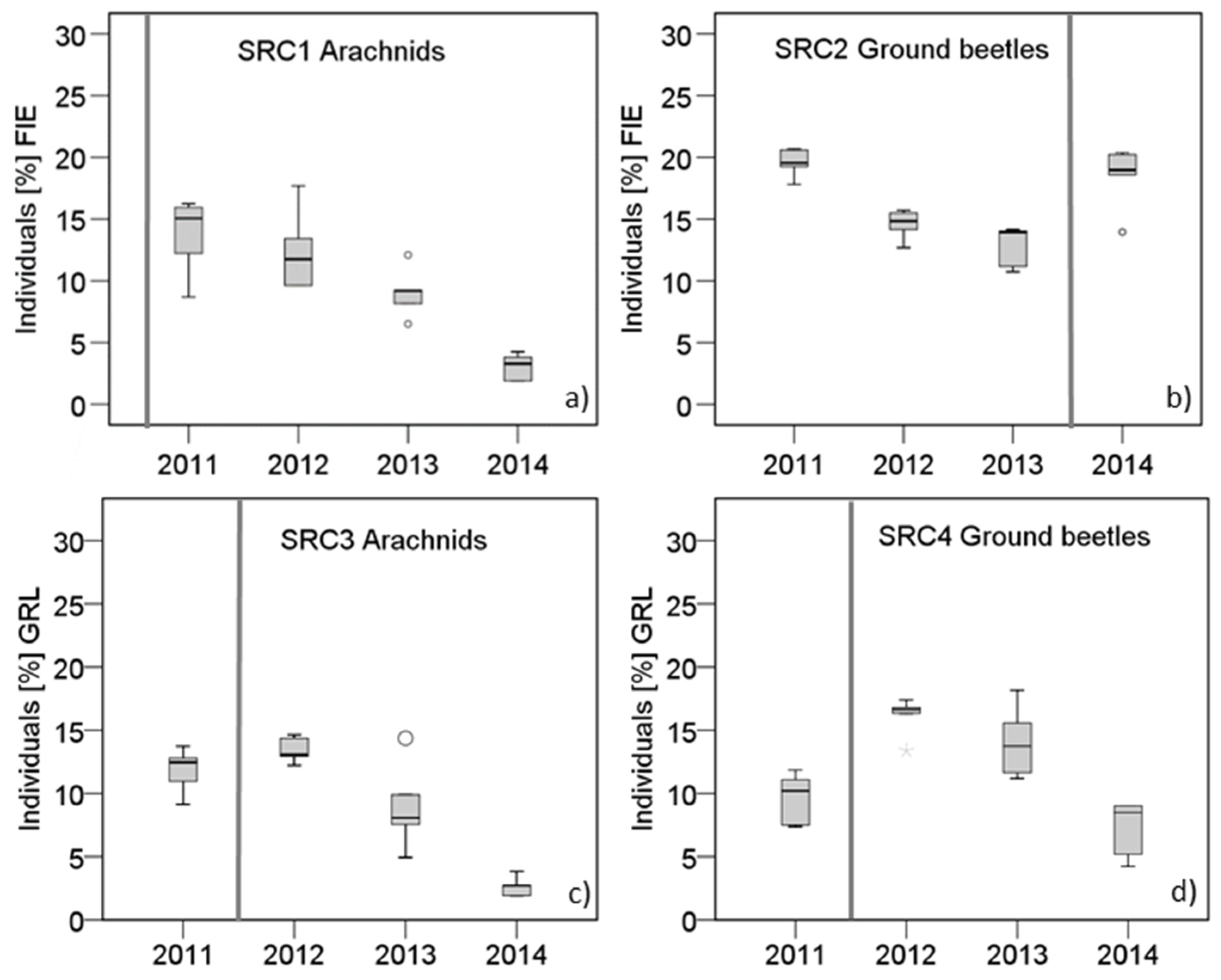
3.5. Cenoses of the SRCs in the Individual Study Years
3.6. Cenoses of the SRCs throughout the Study Period
3.7. Structural and Ecological Diversity
4. Discussion
4.1. Composition of Communities
4.2. Cenotic Changes during SRC Woody Growth
4.3. Vegetation Structure’s Impact on Ecological Species Traits within Communities
4.4. Effects of Timber Harvesting
4.5. Structural Diversity’s Impact on Ecological Variety
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ortiz, A.M.D.; Outhwaite, C.L.; Dalin, C.; Newbold, T. A review of the interactions between biodiversity, agriculture, climate change, and international trade: Research and policy priorities. One Earth 2021, 4, 88–101. [Google Scholar] [CrossRef]
- Dudley, N.; Alexander, S. Agriculture and biodiversity: A review. Biodiversity 2017, 18, 45–49. [Google Scholar] [CrossRef]
- Uhler, J.; Redlich, S.; Zhang, J.; Hothorn, T.; Tobisch, C.; Ewald, J.; Thorn, S.; Seibold, S.; Mitesser, O.; Morinière, J. Relationship of insect biomass and richness with land use along a climate gradient. Nat. Commun. 2021, 12, 5946. [Google Scholar] [CrossRef] [PubMed]
- Outhwaite, C.L.; McCann, P.; Newbold, T. Agriculture and climate change are reshaping insect biodiversity worldwide. Nature 2022, 605, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Barros-Rodríguez, A.; Rangseekaew, P.; Lasudee, K.; Pathom-Aree, W.; Manzanera, M. Impacts of agriculture on the environment and soil microbial biodiversity. Plants 2021, 10, 2325. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Wyckhuys, K.A. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Tilman, D.; Fargione, J.; Wolff, B.; D’antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting agriculturally driven global environmental change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, G.; Roslin, T.; Miller, K.; Tamburini, G.; Birkhofer, K.; Caballero-Lopez, B.; Lindström, S.A.M.; Öckinger, E.; Rundlöf, M.; Rusch, A. Crop diversity benefits carabid and pollinator communities in landscapes with semi-natural habitats. J. Appl. Ecol. 2020, 57, 2170–2179. [Google Scholar] [CrossRef]
- O’Rourke, M.E.; Liebman, M.; Rice, M.E. Ground beetle (Coleoptera: Carabidae) assemblages in conventional and diversified crop rotation systems. Environ. Entomol. 2008, 37, 121–130. [Google Scholar] [CrossRef]
- Estrada-Carmona, N.; Sánchez, A.C.; Remans, R.; Jones, S.K. Complex agricultural landscapes host more biodiversity than simple ones: A global meta-analysis. Proc. Natl. Acad. Sci. USA 2022, 119, e2203385119. [Google Scholar] [CrossRef]
- Duelli, P.; Obrist, M.K. Regional biodiversity in an agricultural landscape: The contribution of seminatural habitat islands. Basic Appl. Ecol. 2003, 4, 129–138. [Google Scholar] [CrossRef]
- Thies, C.; Tscharntke, T. Landscape structure and biological control in agroecosystems. Science 1999, 285, 893–895. [Google Scholar] [CrossRef] [PubMed]
- Statistisches Bundesamt Destatis. Data licence Germany—Bodenfläche (Tatsächliche Nutzung): Deutschland, Stichtag, Nutzungsarten–33111-0001—Version 2.0. Available online: https://www-genesis.destatis.de/genesis/downloads/00/tables/33111-0001_00.csv (accessed on 6 December 2023).
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed]
- Seibold, S.; Gossner, M.M.; Simons, N.K.; Blüthgen, N.; Müller, J.; Ambarlı, D.; Ammer, C.; Bauhus, J.; Fischer, M.; Habel, J.C. Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 2019, 574, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Russo, D.; Di Febbraro, M.; Cistrone, L.; Jones, G.; Smeraldo, S.; Garonna, A.P.; Bosso, L. Protecting one, protecting both? Scale-dependent ecological differences in two species using dead trees, the rosalia longicorn beetle and the barbastelle bat. J. Zool. 2015, 297, 165–175. [Google Scholar] [CrossRef]
- Wagner, D.L.; Fox, R.; Salcido, D.M.; Dyer, L.A. A window to the world of global insect declines: Moth biodiversity trends are complex and heterogeneous. Proc. Natl. Acad. Sci. USA 2021, 118, e2002549117. [Google Scholar] [CrossRef]
- Müller, J.; Hothorn, T.; Yuan, Y.; Seibold, S.; Mitesser, O.; Rothacher, J.; Freund, J.; Wild, C.; Wolz, M.; Menzel, A. Weather explains the decline and rise of insect biomass over 34 years. Nature 2023, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Staab, M.; Gossner, M.M.; Simons, N.K.; Achury, R.; Ambarlı, D.; Bae, S.; Schall, P.; Weisser, W.W.; Blüthgen, N. Insect decline in forests depends on species’ traits and may be mitigated by management. Commun. Biol. 2023, 6, 338. [Google Scholar] [CrossRef]
- Purvis, G.; Fadl, A. The influence of cropping rotations and soil cultivation practice on the population ecology of carabids (Coleoptera: Carabidae) in arable land. Pedobiologia 2002, 46, 452–474. [Google Scholar] [CrossRef]
- Requier, F.; Jowanowitsch, K.K.; Kallnik, K.; Steffan-Dewenter, I. Limitation of complementary resources affects colony growth, foraging behavior, and reproduction in bumble bees. Ecology 2020, 101, e02946. [Google Scholar] [CrossRef]
- Gómez, J.E.; Lohmiller, J.; Joern, A. Importance of vegetation structure to the assembly of an aerial web-building spider community in North American open grassland. J. Arachnol. 2016, 44, 28–35. [Google Scholar] [CrossRef]
- Tscharntke, T.; Grass, I.; Wanger, T.C.; Westphal, C.; Batáry, P. Beyond organic farming–harnessing biodiversity-friendly landscapes. Trends Ecol. Evol. 2021, 36, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Chetcuti, J.; Kunin, W.E.; Bullock, J.M. Habitat fragmentation increases overall richness, but not of habitat-dependent species. Front. Ecol. Evol. 2020, 8, 607619. [Google Scholar] [CrossRef]
- Kotze, D.J.; Niemelä, J.; O’Hara, R.B.; Turin, H. Testing abundance-range size relationships in European carabid beetles (Coleoptera, Carabidae). Ecography 2003, 26, 553–566. [Google Scholar] [CrossRef]
- Glück, E.; Kreisel, A. Die Hecke als Lebensraum, Refugium und Vernetzungsstruktur und ihre Bedeutung für die Dispersion von Waldcarabidenarten. Laufene Seminarbeiträge 10/86, Akademie für Landschaftspflege und Naturschutz (ANL) (1988). In Proceedings of the Biotopverbund in der Landschaft, Symposium, Laufen, Germany, 3–5 November 1986; Volume 3, pp. 64–83. [Google Scholar]
- Veste, M.; Böhm, C. Agrarholz-Schnellwachsende Bäume in der Landwirtschaft; Springer: Berlin/Heidelberg, Germany, 2018; p. 529. [Google Scholar]
- Dauber, J.; Jones, M.B.; Stout, J.C. The impact of biomass crop cultivation on temperate biodiversity. GCB Bioenergy 2010, 2, 289–309. [Google Scholar] [CrossRef]
- Schulz, U.; Brauner, O.; Gruß, H. Animal diversity on short-rotation coppices—A review. Landbauforsch. Volkenrode 2009, 59, 171–181. [Google Scholar]
- Baum, S.; Bolte, A.; Weih, M. Short rotation coppice (SRC) plantations provide additional habitats for vascular plant species in agricultural mosaic landscapes. BioEnergy Res. 2012, 5, 573–583. [Google Scholar] [CrossRef]
- Glemnitz, M.; Platen, R.; Krechel, R.; Konrad, J.; Wagener, F. Can short-rotation coppice strips compensate structural deficits in agrarian landscapes? Asp. Appl. Biol. 2013, 118, 153–162. [Google Scholar]
- Müller-Kroehling, S.; Hohmann, G.; Helbig, C.; Liesebach, M.; Lübke-Al Hussein, M.; Al Hussein, I.A.; Burmeister, J.; Jantsch, M.C.; Zehlius-Eckert, W.; Müller, M. Biodiversity functions of short rotation coppice stands-results of a meta study on ground beetles (Coleoptera: Carabidae). Biomass Bioenergy 2020, 132, 13. [Google Scholar] [CrossRef]
- Dimitriou, I.; Baum, C.; Baum, S.; Busch, G.; Schulz, U.; Köhn, J.; Lamersdorf, N.; Leinweber, P.; Aronsson, P.; Weih, M. Quantifying environmental effects of short rotation coppice (SRC) on biodiversity, soil and water. In IEA Bioenergy Task; IEA: Paris, France, 2011; pp. 1–34. [Google Scholar]
- Kotze, D.J.; Brandmayr, P.; Casale, A.; Dauffy-Richard, E.; Dekoninck, W.; Koivula, M.J.; Lövei, G.L.; Mossakowski, D.; Noordijk, J.; Paarmann, W. Forty years of carabid beetle research in Europe–from taxonomy, biology, ecology and population studies to bioindication, habitat assessment and conservation. ZooKeys 2011, 100, 55. [Google Scholar] [CrossRef]
- Sarma, S.; Pujari, D.; Rahman, Z. Role of spiders in regulating insect pests in the agricultural ecosystem-an overview. J. Int. Acad. Res. Multidiscip. 2013, 1, 100–117. [Google Scholar]
- Ysnel, F.; Canard, A. Spider biodiversity in connection with the vegetation structure and the foliage orientation of hedges. J. Arachnol. 2000, 28, 107–114. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef]
- Bianchi, F.J.; Booij, C.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Ribera, I.; Dolédec, S.; Downie, I.S.; Foster, G.N. Effect of land disturbance and stress on species traits of ground beetle assemblages. Ecology 2001, 82, 1112–1129. [Google Scholar] [CrossRef]
- Langeveld, H.; Quist-Wessel, F.; Dimitriou, I.; Aronsson, P.; Baum, C.; Schulz, U.; Bolte, A.; Baum, S.; Köhn, J.; Weih, M. Assessing environmental impacts of short rotation coppice (SRC) expansion: Model definition and preliminary results. Bioenergy Res. 2012, 5, 621–635. [Google Scholar] [CrossRef]
- Vanbeveren, S.P.; Ceulemans, R. Biodiversity in short-rotation coppice. Renew. Sustain. Energy Rev. 2019, 111, 34–43. [Google Scholar] [CrossRef]
- Hessisches Landesamt für Naturschutz–Umwelt und Geologie (HLNUG). BodenViewer Hessen. Available online: http://bodenviewer.hessen.de (accessed on 14 January 2019).
- GeoBasis-DE/BKG. Verwaltungsgebiete der Bundesländer Deutschlands. VG250 im Maßstab 1:250.000; Bundesamt für Kartographie und Geodäsie (BKG): Frankfurt (am Main), Germany, 2011.
- Hessisches Statistisches Landesamt (HSL). Agrarstrukturerhebung–Kennziffer: CIV9 1a-2–4j/16; Hessisches Statistisches Landesamt (Hrsg.): Wiesbaden, Germany, 2017.
- Destatis. Bodennutzung der Betriebe (Struktur der Bodennutzung), Fachserie 3, Reihe 2.1.2, Vj 2020; 2030212207004; Statistisches Bundesamt: Wiesbaden, Germany, 2021.
- Deutscher Wetterdienst (DWD). Open-Data-Server des Deutschen Wetterdienstes; Deutscher Wetterdienst, Ed.; Deutscher Wetterdienst: Offenbach, Germany, 2018. Available online: https://opendata.dwd.de (accessed on 21 October 2018).
- Dierschke, H. Pflanzensoziologie: Grundlagen und Methoden; Ulmer Stuttgart: Stuttgart, Germany, 1994; p. 683. [Google Scholar]
- Barber, H.S. Traps for cave-inhabiting insects. J. Elisha Mitchell Sci. Soc. 1931, 46, 259–266. [Google Scholar]
- Müller-Motzfeld, G. Vol. 2 Adephaga 1: Carabidae (Laufkäfer). In Die Käfer Mitteleuropas; Freude, H., Harde, K.W., Lohse, G.A., Klausnitzer, B., Eds.; Spektrum Akademischer Verlag (Elsevier): Heidelberg, Germany, 2004; p. 521. [Google Scholar]
- Almquist, S. Swedish Araneae, Part. 1 Families Atypidae to Hahniidae (Linyphiidae excluded); Scandinavian Entomology: Lund, Sweden, 2005; Volume 62, p. 284. [Google Scholar]
- Almquist, S. Swedish Araneae, Part. 2 Families Dictynidae to Salticidae; Brill: Leiden, The Netherlands, 2006; Volume 63, p. 318. [Google Scholar]
- Heimer, S.; Nentwig, W. Spinnen Mitteleuropas: Ein Bestimmungsbuch; Parey: Berlin, Germany, 1991; p. 543. [Google Scholar]
- Locket, G.H.; Millidge, A.F. British Spiders Vol. I; Ray Society: London, UK, 1951; p. 310. [Google Scholar]
- Locket, G.H.; Millidge, A.F. British Spiders Vol. II; Ray Society: London, UK, 1953; p. 449. [Google Scholar]
- Locket, G.A.; Millidge, A.F.; Merrett, P. British Spiders Vol. III; Ray Society: London, UK, 1974; p. 315. [Google Scholar]
- Roberts, M.J. The Spider Fauna of Great Britain and Ireland. Linyphiidae; Brill Archive: Colchester, UK, 1987; Volume 2, p. 204. [Google Scholar]
- Roberts, M.J. The Spider Fauna of Great Britain and Ireland. Atypidae-Theridiosomatidae; Brill Archive: Colchester, UK, 1985; Volume 1, p. 229. [Google Scholar]
- Roberts, M.J. Spiders of Britain & Northern Europe; HarperCollins Publishers: London, UK, 1995; p. 383. [Google Scholar]
- Wiehle, H. Linyphiidae-Baldachinspinnen 44. Teil; VEB Gustav Fischer Verlag: Jena, Germany, 1956; Volume 44, p. 337. [Google Scholar]
- Wiehle, H. Micryphantidae-Zwergspinnen 47. Teil; VEB Gustav Fischer Verlag: Jena, Germany, 1960; Volume 47, p. 620. [Google Scholar]
- Martens, J. Weberknechte, Opiliones. Die Tierwelt Deutschlands 64. Teil; VEB Gustav Fischer Verlag: Jena, Germany, 1978; p. 464. [Google Scholar]
- Schmidt, J.; Trautner, J.; Müller-Motzfeld, G. Rote Liste und Gesamtartenliste der Laufkäfer (Coleoptera: Carabidae) Deutschlands. Naturschutz Biol. Vielfalt 2016, 70, 139–204. [Google Scholar]
- Platnick, N. World Spider Catalog. Version 21.5; Natural History Museum Bern: Bern, Switzerland, 2020. [Google Scholar] [CrossRef]
- Muster, C.; Blick, T.; Schönhofer, A. Rote Liste und Gesamtartenliste der Weberknechte (Arachnida: Opiliones) Deutschlands. Naturschutz Biol. Vielfalt 2016, 70, 513–536. [Google Scholar]
- Barndt, D.; Brase, S.; Glauche, M.; Gruttke, H.; Kegel, B.; Platen, R.; Winkelmann, H. Die Laufkäferfauna von Berlin (West)-mit Kennzeichnung und Auswertung der verschollenen und gefährdeten Arten (Rote Liste, 3. Fassung). In Rote Liste der Gefährdeten Pflanzen und Tiere in Berlin. Landschaftsentwicklung und Umweltforschung. Sonderausgabe 6; Auhagen, A., Platen, R., Sukopp, H., Eds.; Technische Universität: Berlin, Germany, 1991; Volume 6, pp. 243–275. [Google Scholar]
- Platen, R.; Moritz, M.; Broen, B.V. Liste der Webspinnen-und Weberknechtarten (Arach.: Araneida, Opilionida) des Berliner Raumes und ihre Auswertung für Naturschutzzwecke (Rote Liste). In Rote Liste der Gefährdeten Pflanzen und Tiere in Berlin. Landschaftsentwicklung und Umweltforschung. Sonderband 6; Auhagen, A., Platen, R., Sukopp, H., Eds.; Technische Universität: Berlin, Germany, 1991; Volume 6, pp. 169–205. [Google Scholar]
- Konrad, J. Der Einfluss der Vegetationsstruktur und die Auswirkungen des Kurzumtriebs auf die Diversität und Zönosenstruktur Ausgewählter Arthropodengemeinschaften (Col.: Carabidae; Arach.: Araneae et Opiliones) in Agrarholzflächen Nordhessens. Ph.D. Thesis, Martin-Luther-Universität Halle-Wittenberg, Halle (Saale), Germany, 2022; 132p. + Appendix. [Google Scholar]
- Gesellschaft Für Angewandte Carabidologie (GAC). Lebensraumpräferenzen der Laufkäfer Deutschlands–Wissensbasierter Katalog. Angew. Carabidol. Suppl. V 2009, 1, 45. [Google Scholar]
- Magurran, A.E.; McGill, B.J. Biological Diversity. Frontiers in Measurement and Assessment; Oxford University Press: Oxford, NY, USA, 2013; p. 345. [Google Scholar]
- Platen, R.; Konrad, J.; Glemnitz, M. Novel energy crops: An opportunity to enhance the biodiversity of arthropod assemblages in biomass feedstock cultures? Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2017, 13, 162–171. [Google Scholar] [CrossRef]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5; Cambridge University Press: Cambridge, MA, USA, 2014; p. 376. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P.; Legendre, L.F. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 24, p. 990. [Google Scholar]
- Darlington, R.B.; Hayes, A.F. Regression Analysis and Linear Models: Concepts, Applications, and Implementation; Guilford Publications: New York, NY, USA, 2016; p. 661. [Google Scholar]
- Newey, W.K.; West, K.D. A Simple, Positive Semi-Definite, Heteroskedasticity and Autocorrelation Consistent Covariance Matrix; NBER Technical Paper; NBER: Cambridge, MA, USA, 1986; Volume 55, p. 703. [Google Scholar]
- Cohen, J.; Cohen, P.; West, S.; Aiken, L. Applied Multiple Regression/Correlation Analysis for the Behavioral Sciences (3. Aufl.); Lawrence Erlbaum Associates: Mahwah, NJ, USA, 2003; p. 736. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows; Version 26.0; IBM Corp.: Armonk, NY, USA, 2019. [Google Scholar]
- Hayes, A.F.; Cai, L. Using heteroscedasticity-consistent standard error estimators in OLS regression: An introduction and software implementation. Behav. Res. Methods 2007, 39, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Grüner, H. Regression Diagnostic Tests with R. with R Libraries Lmtest and Zoo; Freie Universität Berlin: Berlin, Germany, 2021; Available online: http://gruener.userpage.fu-berlin.de (accessed on 6 October 2021).
- Boháč, J.; Celjak, I.; Moudrý, J.; Kohout, P.; Wotavová, K. Communities of beetles in plantations of fast growing plant species for energetic purposes. Entomol. Rom. 2007, 12, 213–221. [Google Scholar]
- Havlíčková, K.; Rudišová, I. A short rotation coppice of fast-growing trees, their landscape aspects and biodiversity. Ekológia 2011, 30, 12–21. [Google Scholar] [CrossRef]
- Helbig, C.; Müller, M. Habitatqualität von Kurzumtriebsplantagen für die epigäische Fauna am Beispiel der Laufkäfer (Coleoptera, Carabidae). Agrowood Kurzumtriebsplantagen Dtschl. Eur. Perspekt. Hg. Bemman A 2010, 147–152. [Google Scholar]
- Verheyen, K.; Buggenhout, M.; Vangansbeke, P.; De Dobbelaere, A.; Verdonckt, P.; Bonte, D. Potential of Short Rotation Coppice plantations to reinforce functional biodiversity in agricultural landscapes. Biomass Bioenergy 2014, 67, 435–442. [Google Scholar] [CrossRef]
- Blick, T.; Weiss, I.; Burger, F. Spinnentiere einer neu angelegten Pappel-Kurzumtriebsfläche (Energiewald) und eines Ackers bei Schwarzenau (Lkr. Kitzingen, Unterfranken, Bayern). Arachnol. Mitteilungen 2003, 25, 1–16. [Google Scholar] [CrossRef]
- Allegro, G.; Sciaky, R. Assessing the potential role of ground beetles (Coleoptera, Carabidae) as bioindicators in poplar stands, with a newly proposed ecological index (FAI). For. Ecol. Manag. 2003, 175, 275–284. [Google Scholar] [CrossRef]
- Britt, C.; Fowbert, J.; McMillan, S. The ground flora and invertebrate fauna of hybrid poplar plantations: Results of ecological monitoring in the PAMUCEAF project. Asp. Appl. Biol. 2007, 82, 83–90. [Google Scholar]
- Sachs, D.; Brauner, O.; Schulz, U. Laufkäfer (Carabidae) auf Energieholzflächen–die Bedeutung von Begleitstrukturen für Diversität und Abundanz. Mitteilungen Dtsch. Ges. Allg. Angew. Entomol. 2012, 18, 157–161. [Google Scholar]
- Ulrich, W.; Buszko, J.; Czarnecki, A. The contribution of poplar plantations to regional diversity of ground beetles (Coleoptera: Carabidae) in agricultural landscapes. Ann. Zool. Fenn. 2004, 41, 501–512. [Google Scholar]
- Schulz, U. Tierartenvielfalt. In Energieholzproduktion in der Landwirtschaft–Chancen und Risiken aus Sicht des Natur- und Umweltschutzes. Naturschutzbund Deutschland; NABU, Ed.; Warlich Druck: Berlin, Germany, 2008; pp. 41–49. [Google Scholar]
- Šťastná, P. Diversity of ground beetles (Carabidae) in the plantations of fast growing trees. Acta Univ. Agric. Silvic. Mendel. Brun. 2013, 60, 309–316. [Google Scholar] [CrossRef]
- Weger, J.; Vávrová, K.; Kašparová, L.; Bubeník, J.; Komárek, A. The influence of rotation length on the biomass production and diversity of ground beetles (Carabidae) in poplar short rotation coppice. Biomass Bioenergy 2013, 54, 284–292. [Google Scholar] [CrossRef]
- Blick, T.; Burger, F. Wirbellose in Energiewäldern. Am Beispiel der Spinnentiere der Kurzumtriebsfläche Wöllershof (Oberpfalz, Bayern). Naturschutz Landschaftsplanung 2002, 34, 276–284. [Google Scholar]
- Brauner, O.; Schulz, U. Laufkäfer auf Energieholzplantagen und angrenzenden Vornutzungsflächen (Carabidae)–Untersuchungen in Sachsen und Brandenburg. Entomol. Blätter Biol. Syst. Käfer 2011, 107, 31–64. [Google Scholar]
- Kriegel, P.; Fritze, M.A.; Thorn, S. Surface temperature and shrub cover drive ground beetle (Coleoptera: Carabidae) assemblages in short-rotation coppices. Agric. For. Entomol. 2021, 23, 400–410. [Google Scholar] [CrossRef]
- Nerlich, K.; Seidl, F.; Mastel, K.; Graeff-Hönninger, S.; Claupein, W. Auswirkungen von Weiden (Salix spp.) und Pappeln (Populus spp.) im Kurzumtrieb auf die biologische Vielfalt am Beispiel von Laufkäfern (Carabidae). Gesunde Pflanz. 2012, 64, 129–139. [Google Scholar] [CrossRef]
- Schardt, M.; Burger, F.; Blick, T. Ökologischer Vergleich der Spinnenfauna (Arachnida: Araneae) von Energiewäldern und Ackerland. DGaaE Mitteilungen 2008, 16, 131–135. [Google Scholar]
- Al Hussein, I.A.; Röhricht, C.; Ruscher, K.; Lübke-Al Hussein, M. Ökologische Bewertung von Energieholzanlagen und einer Naturschutzhecke auf großen Ackerschlägen am Beispiel der Laufkäfer. Angew. Carabidol. 2014, 10, 87–95. [Google Scholar]
- Liesebach, M.; Mecke, R. Die Laufkäferfauna einer Kurzumtriebsplantage, eines Gerstenackers und eines Fichtenwaldes im Vergleich. Die Holzzucht 2003, 54, 11–15. [Google Scholar]
- Burger, F. Zur Ökologie von Energiewäldern. Schriftenreihe Dtsch. Rates Landespfl. 2006, 79, 74–80. [Google Scholar]
- Lamersdorf, N.; Bielefeldt, J.; Bolte, A.; Busch, G.; Dohrenbusch, A.; Knust, C.; Kroiher, F.; Schulz, U.; Stoll, B. Das Projekt NOVALIS–zur naturverträglichen Produktion von Energieholz in der Landwirtschaft. Arch. Forstwes. Landschaftsökologie 2008, 43, 138–141. [Google Scholar]
- Gruttke, H.; Willeke, S. Tierökologische Langzeitstudie zur Besiedlung neu angelegter Gehölzpflanzungen in der intensiv bewirtschafteten Agrarlandschaft-ein E+ E-Vorhaben. Nat. Landsch. 1993, 68, 367–376. [Google Scholar]
- Piotrowska, N.S.; Czachorowski, S.Z.; Stolarski, M.J. Ground Beetles (Carabidae) in the Short-Rotation Coppice Willow and Poplar Plants—Synergistic Benefits System. Agriculture 2020, 10, 648. [Google Scholar] [CrossRef]
- Bruggisser, O.T.; Sandau, N.; Blandenier, G.; Fabian, Y.; Kehrli, P.; Aebi, A.; Naisbit, R.E.; Bersier, L.-F. Direct and indirect bottom-up and top-down forces shape the abundance of the orb-web spider Argiope bruennichi. Basic Appl. Ecol. 2012, 13, 706–714. [Google Scholar] [CrossRef]
- Brunk, I. Diversität von Laufkäfern (Coleoptera, Carabidae), höheren Pflanzen und struktureller Diversität in frühen Sukzessionsstadien. In Biodiversität und Sukzession in der Niederlausitzer Bergbaufolgelandschaft: BIOLOG; SUBICON; Bröring, U., Wiegleb, G., Eds.; Books on Demand GmbH: Norderstedt, Germany, 2006; pp. 22–44. [Google Scholar]
- Ings, T.; Hartley, S. The effect of habitat structure on carabid communities during the regeneration of a native Scottish forest. For. Ecol. Manag. 1999, 119, 123–136. [Google Scholar] [CrossRef]
- Mcnett, B.J.; Rypstra, A.L. Habitat selection in a large orb-weaving spider: Vegetational complexity determines site selection and distribution. Ecol. Entomol. 2000, 25, 423–432. [Google Scholar] [CrossRef]
- Meißner, A. Die Bedeutung der Raumstruktur für die Habitatwahl von Lauf- und Kurzflügelkäfern–Freilandökologische und Experimentelle Untersuchungen Einer Niedermoorzönose. Ph.D. Thesis, Technische Universität Berlin, Berlin, Germany, 1998; p. 184. [Google Scholar]
- Robinson, J.V. The effect of architectural variation in habitat on a spider community: An experimental field study. Ecology 1981, 62, 73–80. [Google Scholar] [CrossRef]
- Uetz, G. Habitat structure and spider foraging. In Habitat Structure; Mc Coy, E.D., Mushinsky, H.R., Eds.; Chapman and Hall: London, UK, 1991; pp. 325–348. [Google Scholar]
- Brose, U. Bottom-up control of carabid beetle communities in early successional wetlands: Mediated by vegetation structure or plant diversity? Oecologia 2003, 135, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Brunk, I. Diversität und Sukzession von Laufkäferzönosen in Gestörten Landschaften Südbrandenburgs. Ph.D. Thesis, Universität Cottbus, Cottbus, Germany, 2007. [Google Scholar]
- Gibb, H.; Cunningham, S. Revegetation of farmland restores function and composition of epigaeic beetle assemblages. Biol. Conserv. 2010, 143, 677–687. [Google Scholar] [CrossRef]
- Jukes, M.R.; Peace, A.J.; Ferris, R. Carabid beetle communities associated with coniferous plantations in Britain: The influence of site, ground vegetation and stand structure. For. Ecol. Manag. 2001, 148, 271–286. [Google Scholar] [CrossRef]
- Liu, J.-L.; Li, F.-R.; Sun, T.-S.; Ma, L.-F.; Liu, L.-L.; Yang, K. Interactive effects of vegetation and soil determine the composition and diversity of carabid and tenebrionid functional groups in an arid ecosystem. J. Arid Environ. 2016, 128, 80–90. [Google Scholar] [CrossRef]
- Schwab, A.; Dubois, D.; Fried, P.M.; Edwards, P.J. Estimating the biodiversity of hay meadows in north-eastern Switzerland on the basis of vegetation structure. Agric. Ecosyst. Environ. 2002, 93, 197–209. [Google Scholar] [CrossRef]
- Spake, R.; Barsoum, N.; Newton, A.C.; Doncaster, C.P. Drivers of the composition and diversity of carabid functional traits in UK coniferous plantations. For. Ecol. Manag. 2016, 359, 300–308. [Google Scholar] [CrossRef]
- Taboada, A.; Kotze, D.J.; Tárrega, R.; Salgado, J.M. Traditional forest management: Do carabid beetles respond to human-created vegetation structures in an oak mosaic landscape? For. Ecol. Manag. 2006, 237, 436–449. [Google Scholar] [CrossRef]
- Guillemain, M.; Loreau, M.; Daufresne, T. Relationships beetween the regional distribution of carabid beetles (Coleoptera, Carabidae) and the abundance of their potential prey. Acta Oecologica 1997, 18, 465–483. [Google Scholar] [CrossRef]
- Koivula, M.; Punttila, P.; Haila, Y.; Niemelä, J. Leaf litter and the small-scale distribution of carabid beetles (Coleoptera, Carabidae) in the boreal forest. Ecography 1999, 22, 424–435. [Google Scholar] [CrossRef]
- Lassau, S.A.; Hochuli, D.F.; Cassis, G.; Reid, C.A. Effects of habitat complexity on forest beetle diversity: Do functional groups respond consistently? Divers. Distrib. 2005, 11, 73–82. [Google Scholar] [CrossRef]
- Loreau, M. Vertical distribution of the activity of carabid beetles in a beech forest floor. Pedobiologia 1987, 30, 173–178. [Google Scholar] [CrossRef]
- Magura, T.; Tóthmérész, B.; Elek, Z. Impacts of leaf-litter addition on carabids in a conifer plantation. Biodivers. Conserv. 2005, 14, 475–491. [Google Scholar] [CrossRef]
- Stevenson, B.G.; Dindal, D.L. Effect of leaf shape on forest litter spiders: Community organization and microhabitat selection of immature Enoplognatha ovata (Clerck)(Theridiidae). J. Arachnol. 1982, 10, 165–178. [Google Scholar]
- Bonn, A.; Kleinwächter, M. Microhabitat distribution of spider and ground beetle assemblages (Araneae, Carabidae) on frequently inundated river banks of the River Elbe. Z. Okol. Naturschutz 1999, 8, 109–123. [Google Scholar]
- Diehl, E.; Mader, V.L.; Wolters, V.; Birkhofer, K. Management intensity and vegetation complexity affect web-building spiders and their prey. Oecologia 2013, 173, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Hawthorne, A.; Hassall, M. The effect of cereal headland treatments on carabid communities. Acta Jutl. 1995, 70, 185–198. [Google Scholar]
- Kalinkat, G.; Brose, U.; Rall, B.C. Habitat structure alters top-down control in litter communities. Oecologia 2013, 172, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, J.; Walter, R. Auswirkungen von Energieholzstreifen auf Laufkäfer und Nacktschnecken Ökologisch Bewirtschafteter Ackerflächen. In Proceedings of the WTA 2017 Vs. 2.2, Delft, The Netherlands, 13–16 September 2017; pp. 240–243. [Google Scholar]
- Liesebach, M.; Mecke, R.; Rose, A. Die epigäische Wirbellosenfauna einer Kurzumtriebsplantage im Vergleich zu der eines angrenzenden Gerstenackers und der eines Fichtenwaldes. Holzzucht 1999, 53, 21–25. [Google Scholar]
- Rowe, R.L.; Goulson, D.; Doncaster, C.P.; Clarke, D.J.; Taylor, G.; Hanley, M.E. Evaluating ecosystem processes in willow short rotation coppice bioenergy plantations. GCB Bioenergy 2013, 5, 257–266. [Google Scholar] [CrossRef]
- Platen, R.; Konrad, J.; Glemnitz, M. Sägezahnhypothese, Arten/Strukturvielfalt. In Nachwachsende Rohstoffe als Option für den Naturschutz. Naturschutz durch Landbau? Schlussbericht zu ELKE III.; Heck, P., Böhmer, J., Eds.; Umwelt-Campus Birkenfeld: Hoppstädten-Weiersbach, Germany, 2013; p. 802. [Google Scholar]
- Loreau, M.; Barbier, M.; Filotas, E.; Gravel, D.; Isbell, F.; Miller, S.J.; Montoya, J.M.; Wang, S.; Aussenac, R.; Germain, R. Biodiversity as insurance: From concept to measurement and application. Biol. Rev. 2021, 96, 2333–2354. [Google Scholar] [CrossRef]
- Ives, A.R.; Klug, J.L.; Gross, K. Stability and species richness in complex communities. Ecol. Lett. 2000, 3, 399–411. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Cabido, M. Vive la difference: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Walker, B. Conserving biological diversity through ecosystem resilience. Conserv. Biol. 1995, 9, 747–752. [Google Scholar] [CrossRef]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.; Hector, A.; Hooper, D.; Huston, M.; Raffaelli, D.; Schmid, B. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M.; Mouquet, N.; Gonzalez, A. Biodiversity as spatial insurance in heterogeneous landscapes. Proc. Natl. Acad. Sci. USA 2003, 100, 12765–12770. [Google Scholar] [CrossRef] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
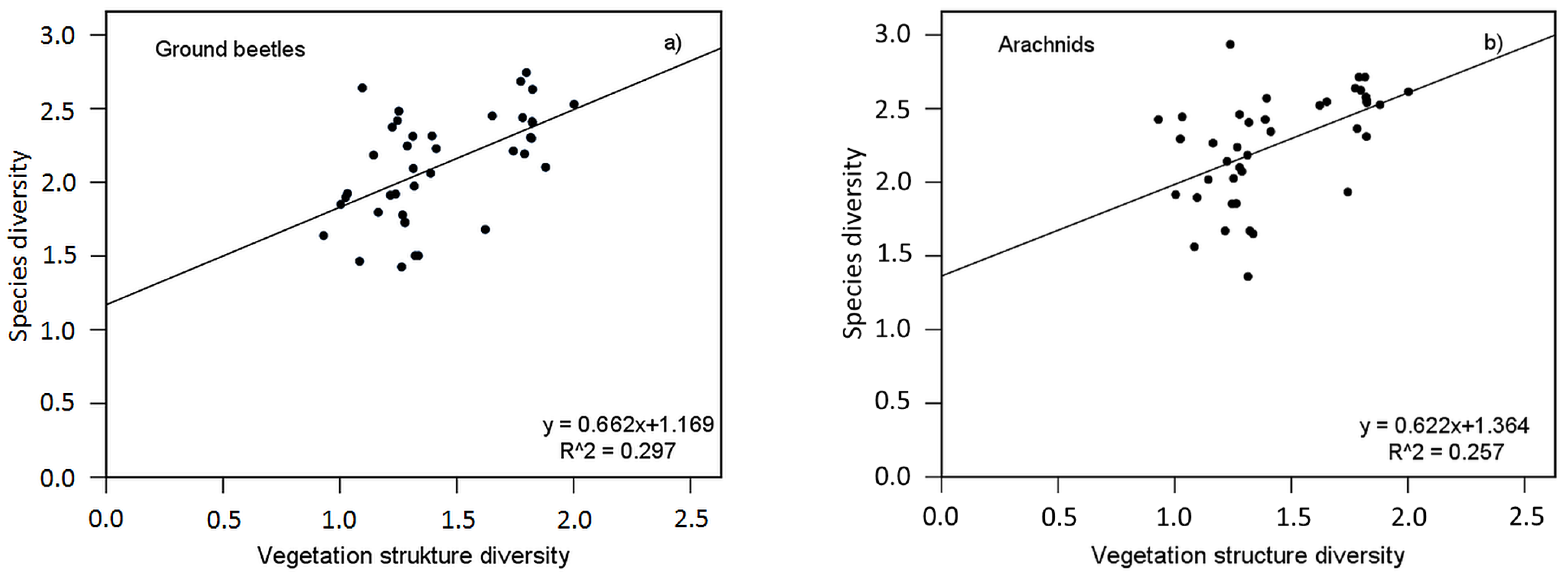
| Ground Beetles | Arachnids | |||
|---|---|---|---|---|
| Shan SpecDiv | Shan HabDiv | Shan SpecDiv | Shan HabDiv | |
| Model overview | ||||
| R2 | 0.297 | 0.380 | 0.257 | 0.445 |
| B | 0.662 | 0.834 | 0.622 | 0.814 |
| F | 16.081 | 23.311 | 13.123 | 30.528 |
| FHAC | 22.676 | 47.613 | 109.518 | 67.629 |
| df | 1. 38 | 1. 38 | 1. 38 | 1. 38 |
| p [Model} | <0.001 | <0.001 | 0.001 | <0.001 |
| Coefficients | ||||
| SE (B) | 0.165 | 0.173 | 0.172 | 0.147 |
| t | 4.010 | 4.828 | 3.623 | 5.525 |
| tHAC | 4.762 | 6.900 | 10.465 | 8.224 |
| pHAC [Coeff.] | <0.001 | <0.001 | 0.001 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konrad, J.; Platen, R.; Glemnitz, M. The Effects of Vegetation Structure and Timber Harvesting on Ground Beetle (Col.: Carabidae) and Arachnid Communities (Arach.: Araneae, Opiliones) in Short-Rotation Coppices. Land 2024, 13, 145. https://doi.org/10.3390/land13020145
Konrad J, Platen R, Glemnitz M. The Effects of Vegetation Structure and Timber Harvesting on Ground Beetle (Col.: Carabidae) and Arachnid Communities (Arach.: Araneae, Opiliones) in Short-Rotation Coppices. Land. 2024; 13(2):145. https://doi.org/10.3390/land13020145
Chicago/Turabian StyleKonrad, Jessika, Ralph Platen, and Michael Glemnitz. 2024. "The Effects of Vegetation Structure and Timber Harvesting on Ground Beetle (Col.: Carabidae) and Arachnid Communities (Arach.: Araneae, Opiliones) in Short-Rotation Coppices" Land 13, no. 2: 145. https://doi.org/10.3390/land13020145