

The Longevity of Fruit Trees in Basilicata (Southern Italy): Implications for Agricultural Biodiversity Conservation
 ,
,  ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
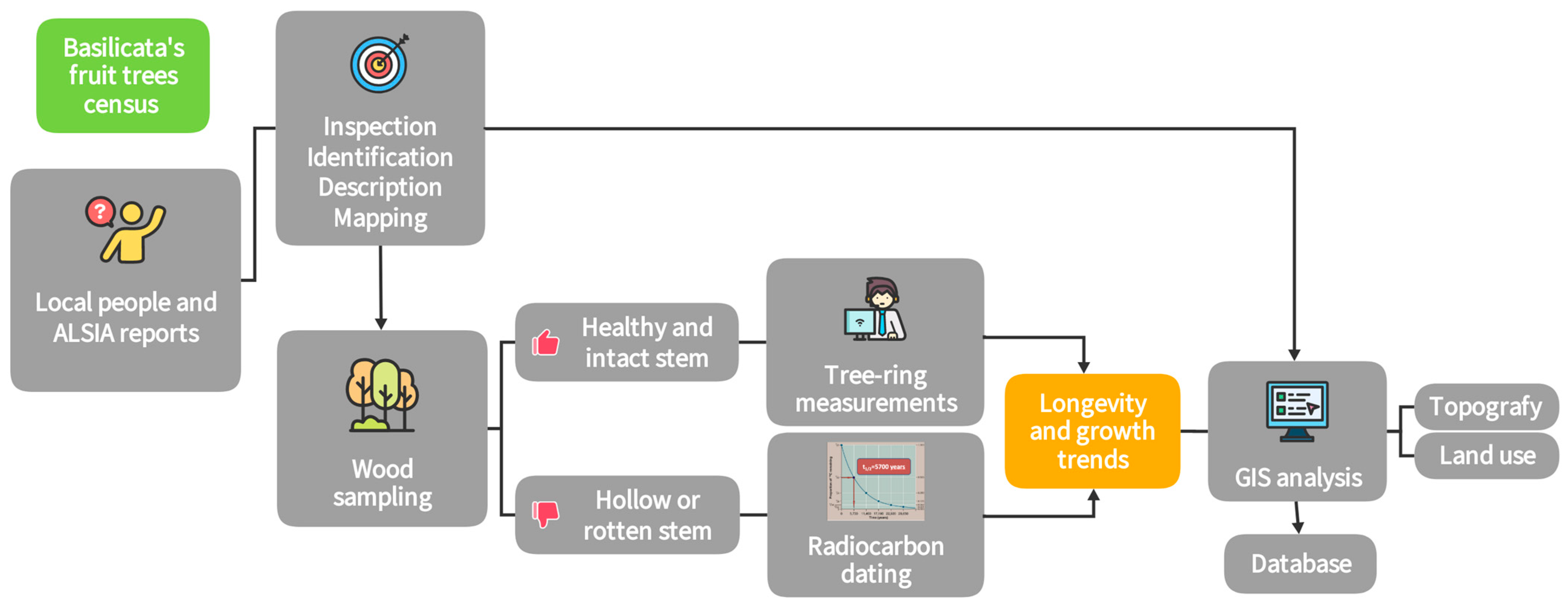
2. Materials and Methods
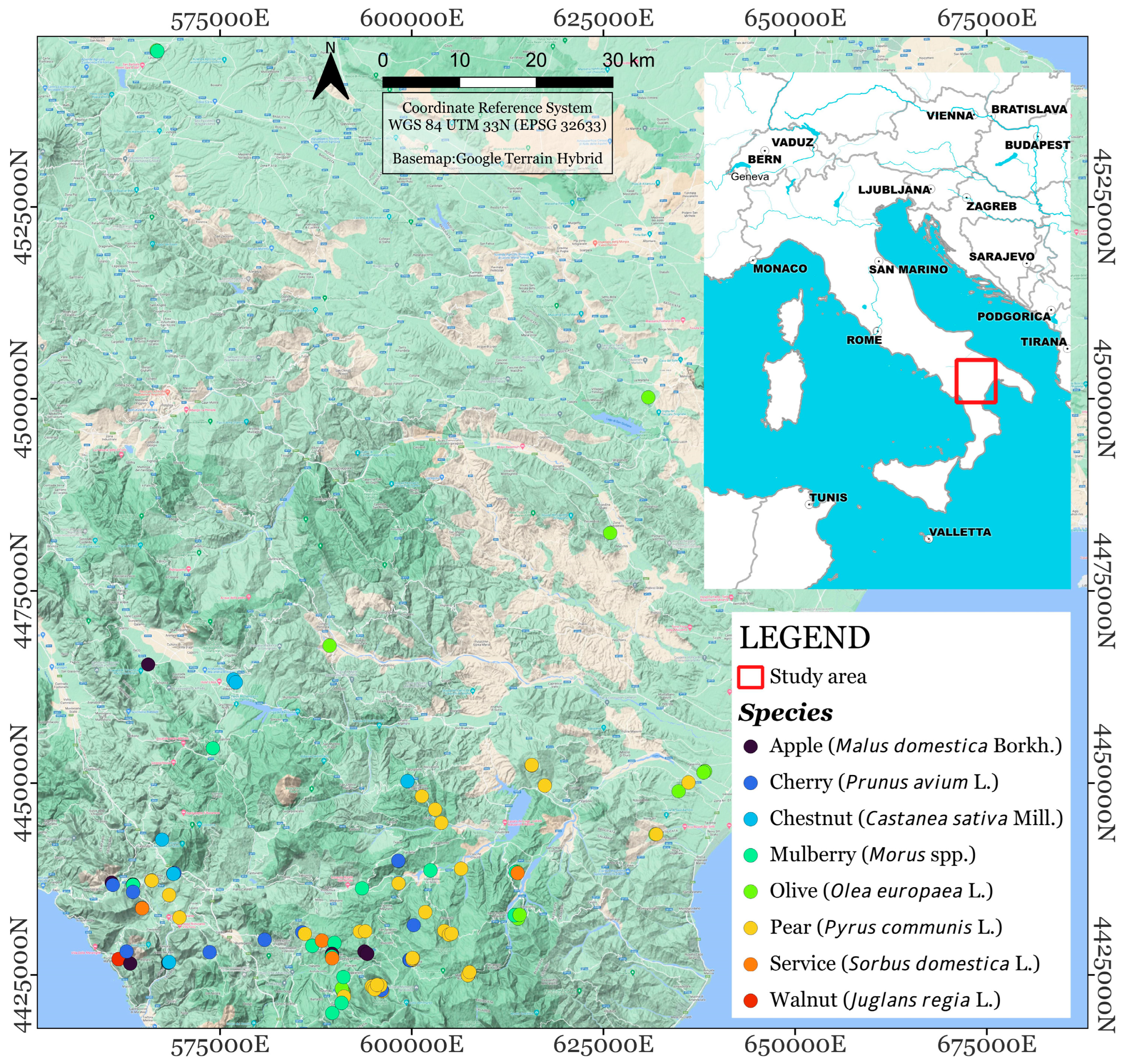
2.1. The Survey
2.2. Longevity, Growth and GIS Analysis
3. Results
3.1. The Census of Fruit Patriarchs in Basilicata
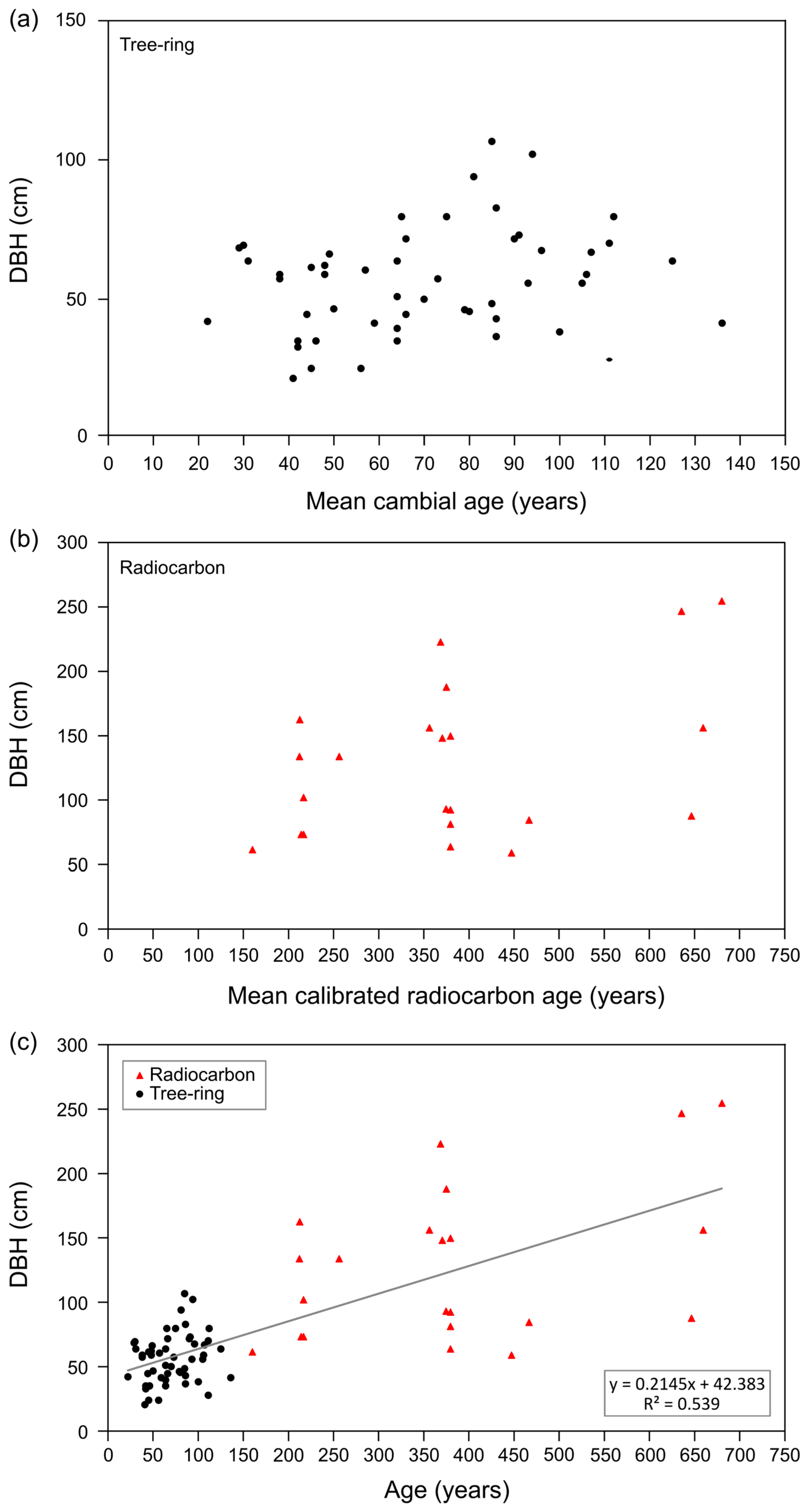
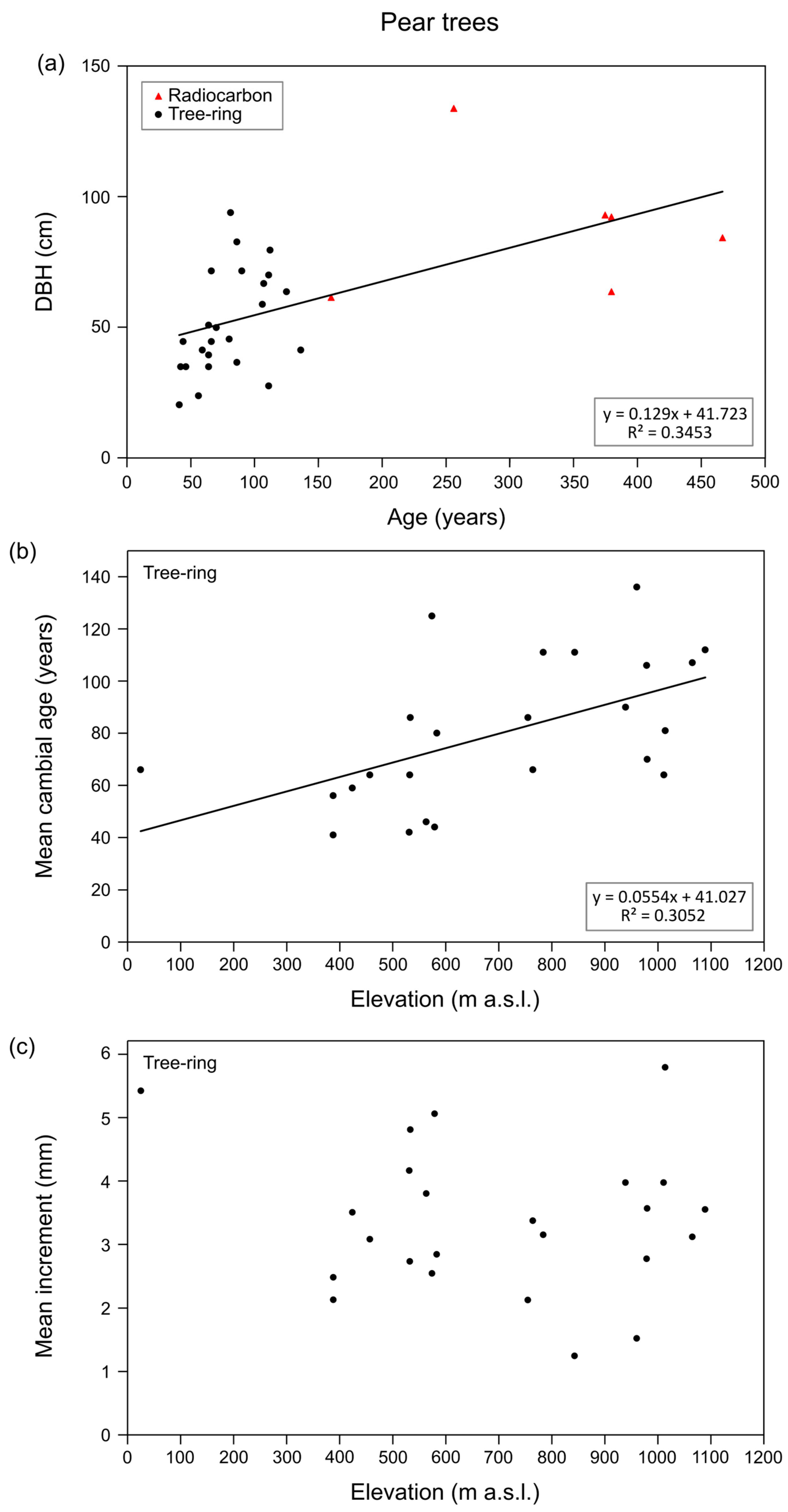
3.2. Growth and Longevity of Fruit Trees
4. Discussion
4.1. The Age of Fruit Patriarchs in Basilicata
4.2. Factors That Favor Old Ages in Fruit Trees
4.3. The Importance of Fruit Tree Longevity for Conservation Biology and Genetic Resources
4.4. Future Research
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diamond, J. Evolution, Consequences and Future of Plant and Animal Domestication. Nature 2002, 418, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Dudley, N.; Alexander, S. Agriculture and Biodiversity: A Review. Biodiversity 2017, 18, 45–49. [Google Scholar] [CrossRef]
- Tilman, D.; Fargione, J.; Wolff, B.; D’Antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting Agriculturally Driven Global Environmental Change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiarucci, A.; Piovesan, G. Need for a Global Map of Forest Naturalness for a Sustainable Future. Conserv. Biol. 2020, 34, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Palli, J.; Mensing, S.A.; Schoolman, E.M.; Solano, F.; Piovesan, G. Historical Ecology Identifies Long-Term Rewilding Strategy for Conserving Mediterranean Mountain Forests in South Italy. Ecol. Appl. 2022, e2758. [Google Scholar] [CrossRef]
- Piovesan, G.; Biondi, F.; Baliva, M.; Dinella, A.; Di Fiore, L.; Marchiano, V.; Presutti Saba, E.; De Vivo, G.; Schettino, A.; Di Filippo, A. Tree Growth Patterns Associated with Extreme Longevity: Implications for the Ecology and Conservation of Primeval Trees in Mediterranean Mountains. Anthropocene 2019, 26, 100199. [Google Scholar] [CrossRef]
- Cortonesi, A. Il Medioevo Degli Alberi: Piante e Paesaggi d’Italia (Secoli XI-XV); Carocci Editore: Rome, Italy, 2022; ISBN 9788829010974. [Google Scholar]
- Schicchi, R.; Speciale, C.; Amato, F.; Bazan, G.; Di Noto, G.; Marino, P.; Ricciardo, P.; Geraci, A. The Monumental Olive Trees as Biocultural Heritage of Mediterranean Landscapes: The Case Study of Sicily. Sustainability 2021, 13, 6767. [Google Scholar] [CrossRef]
- Krebs, P.; Poli, A.; Conedera, M. I Castagni Monumentali Dell’Alpe Di Brusino (Cantone Ticino, Svizzera): Indicazioni per Chi Volesse Azzardarne l’età. Boll. Della Soc. Ticin. di Sci. Nat. 2019, 107, 41–53. [Google Scholar]
- Petruccelli, R.; Giordano, C.; Salvatici, M.C.; Beghè, D.; Rodolfi, M.; Fabbri, A.; Benelli, C. Characterization and Conservation of “Olivo Della Strega”: An Ancient Olive Tree, Precious Resource for Natural and Cultural Heritage. Rend. Lincei 2021, 32, 311–324. [Google Scholar] [CrossRef]
- Ninot, A.; Howad, W.; Aranzana, M.J.; Senar, R.; Romero, A.; Mariotti, R.; Baldoni, L.; Belaj, A. Survey of over 4, 500 Monumental Olive Trees Preserved on-Farm in the Northeast Iberian Peninsula, Their Genotyping and Characterization. Sci. Hortic. 2018, 231, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Conedera, M.; Krebs, P.; Tinner, W.; Pradella, M.; Torriani, D. The Cultivation of Castanea Sativa (Mill.) in Europe, from Its Origin to Its Diffusion on a Continental Scale. Veg. Hist. Archaeobot. 2004, 13, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Besnard, G.; Terral, J.F.; Cornille, A. On the Origins and Domestication of the Olive: A Review and Perspectives. Ann. Bot. 2018, 121, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fideghelli, C. (Coordinator) Atlante Dei Fruttiferi Autoctoni Italiani. 2016. Available online: https://iris.unito.it/retrieve/handle/2318/1681120/450898/Capitolo_Piemonte.pdf (accessed on 15 January 2023).
- Routson, K.J.; Routson, C.C.; Sheppard, P.R. Dendrochronology Reveals Planting Dates of Historic Apple Trees in the Southwestern United States. J. Am. Pomol. Soc. 2012, 66, 9–15. [Google Scholar]
- Lageard, J.G.A. Aging Mulberry Trees (Morus Nigra L): The Charterhouse, London, UK. Arboric. J. 2022, 44, 127–139. [Google Scholar] [CrossRef]
- Piovesan, G.; Cannon, C.H.; Liu, J.; Munné-Bosch, S. Ancient Trees: Irreplaceable Conservation Resource for Ecosystem Restoration. Trends Ecol. Evol. 2022, 37, 1025–1028. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Lorenzo, S.; Ballester, A.; Corredoira, E.; Vieitez, A.M.; Agnanostakis, S.; Costa, R.; Bounous, G.; Botta, R.; Beccaro, G.L.; Kubisiak, T.L.; et al. Chestnut. In Fruit Breeding. Handbook of Plant Breeding; Badenes, M., Byrne, D., Eds.; Springer: Boston, MA, USA, 2012; Volume 8, pp. 729–769. [Google Scholar] [CrossRef]
- Salimonti, A.; Simeone, V.; Cesari, G.; Lamaj, F.; Cattivelli, L.; Perri, E.; Desiderio, F.; Fanizzi, F.P.; Del Coco, L.; Zelasco, S. A First Molecular Investigation of Monumental Olive Trees in Apulia Region. Sci. Hortic. 2013, 162, 204–212. [Google Scholar] [CrossRef]
- Hajdas, I.; Ascough, P.; Garnett, M.H.; Fallon, S.J.; Pearson, C.L.; Quarta, G.; Spalding, K.L.; Yamaguchi, H.; Yoneda, M. Radiocarbon Dating. Nat. Rev. Methods Prim. 2021, 1, 62. [Google Scholar] [CrossRef]
- Piovesan, G.; Biondi, F.; Baliva, M.; Calcagnile, L.; Quarta, G.; Di Filippo, A. Dating Old Hollow Trees by Applying a Multistep Tree-Ring and Radiocarbon Procedure to Trunk and Exposed Roots. MethodsX 2018, 5, 495–502. [Google Scholar] [CrossRef]
- Piovesan, G.; Baliva, M.; Calcagnile, L.; D’Elia, M.; Dorado-Liñán, I.; Palli, J.; Siclari, A.; Quarta, G. Radiocarbon Dating of Aspromonte Sessile Oaks Reveals the Oldest Dated Temperate Flowering Tree in the World. Ecology 2020, 101, 1–4. [Google Scholar] [CrossRef]
- Aniol, R.W. A New Device for Computer Assisted Measurement of Tree-Ring Widths. Dendrochronologia 1987, 5, 135–141. [Google Scholar]
- Aniol, R.W. Tree-Ring Analysis Using CATRAS. Dendrochronologia 1983, 1, 45–53. [Google Scholar]
- Calcagnile, L.; Maruccio, L.; Scrimieri, L.; delle Side, D.; Braione, E.; D’Elia, M.; Quarta, G. Development and Application of Facilities at the Centre for Applied Physics, Dating and Diagnostics (CEDAD) at the University of Salento during the Last 15 years. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 2019, 456, 252–256. [Google Scholar] [CrossRef]
- Reimer, P.J.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 Cal KBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Cherubini, P.; Humbel, T.; Beeckman, H.; Gärtner, H.; Mannes, D.; Pearson, C.; Schoch, W.; Tognetti, R.; Lev-Yadun, S. Olive Tree-Ring Problematic Dating: A Comparative Analysis on Santorini (Greece). PLoS ONE 2013, 8, e54730. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.J.; Colangelo, M.; Gracia-Balaga, A.; Ortega-Martínez, M.A.; Büntgen, U. Demystifying the Age of Old Olive Trees. Dendrochronologia 2021, 65, 125802. [Google Scholar] [CrossRef]
- Jarman, R.; Moir, A.K.; Webb, J.; Chambers, F.M.; Russell, K. Dendrochronological Assessment of British Veteran Sweet Chestnut (Castanea sativa) Trees: Successful Cross-Matching, and Cross-Dating with British and French Oak (Quercus) Chronologies. Dendrochronologia 2018, 51, 10–21. [Google Scholar] [CrossRef]
- Bernabei, M. The Age of the Olive Trees in the Garden of Gethsemane. J. Archaeol. Sci. 2015, 53, 43–48. [Google Scholar] [CrossRef]
- Ehrlich, Y.; Regev, L.; Kerem, Z.; Boaretto, E. Radiocarbon Dating of an Olive Tree Cross-Section: New Insights on Growth Patterns and Implications for Age Estimation of Olive Trees. Front. Plant Sci. 2017, 8, 1918. [Google Scholar] [CrossRef]
- Biondi, F.; Meko, D.M.; Piovesan, G. Maximum Tree Lifespans Derived from Public-Domain Dendrochronological Data. iScience 2023. [Google Scholar] [CrossRef]
- Liu, J.; Xia, S.; Zeng, D.; Liu, C.; Li, Y.; Yang, W.; Yang, B.; Zhang, J.; Slik, F.; Lindenmayer, D.B. Age and Spatial Distribution of the World’s Oldest Trees. Conserv. Biol. 2022, 36, e13907. [Google Scholar] [CrossRef]
- Piovesan, G.; Biondi, F. On Tree Longevity. New Phytol. 2021, 231, 1318–1337. [Google Scholar] [CrossRef] [PubMed]
- Lageard, J.G.A.; Sultana, D.; Brearley, F.Q. Veteran Trees in an Historic Landscape: The Bidnija Olive Grove, Malta. J. Archaeol. Sci. Rep. 2021, 38, 103094. [Google Scholar] [CrossRef]
- Christian, T.; Coles, P. “Morus” from the Website Trees and Shrubs Online. Available online: treesandshrubsonline.org/articles/morus/ (accessed on 15 January 2023).
- Vavilov, N.I.; Dorofeev, V.F. Origin and Geography of Cultivated Plants; Cambridge University Press: Cambridge, UK, 1992; p. 498. [Google Scholar]
- Hancock, J.F.; Lobos, G.A. Pears. Temp. Fruit Crop Breed. Germplasm Genom. 2008, 9781402069079, 299–335. [Google Scholar] [CrossRef]
- Janick, J. The Pear in History, Literature, Popular Culture and Art. Acta Hortic. 2002, 596, 41–52. [Google Scholar] [CrossRef]
- Eccher, T.; Pontiroli, R. Old Pear Varieties in Northern Italy. Acta Hortic. 2005, 671, 243–246. [Google Scholar] [CrossRef]
- Silva, G.J.; Souza, T.M.; Barbieri, R.L.; Costa De Oliveira, A. Origin, Domestication, and Dispersing of Pear (Pyrus spp.). Adv. Agric. 2014, 2014, 541097. [Google Scholar] [CrossRef] [Green Version]
- McKenna, D. A Study of Artifacts from the Collection of Leonard Russell, Danversport, Massachusetts. Bull. Mass. Archaeol. Soc. 2018, 79, 23. [Google Scholar]
- Winter, M.B.; Wolff, B.; Gottschling, H.; Cherubini, P. The Impact of Climate on Radial Growth and Nut Production of Persian Walnut (Juglans Regia L.) in Southern Kyrgyzstan. Eur. J. For. Res. 2009, 128, 531–542. [Google Scholar] [CrossRef]
- Berni, R.; Cantini, C.; Romi, M.; Hausman, J.F.; Guerriero, G.; Cai, G. Agrobiotechnology Goes Wild: Ancient Local Varieties as Sources of Bioactives. Int. J. Mol. Sci. 2018, 19, 2248. [Google Scholar] [CrossRef] [Green Version]
- Hofmeister, B.T.; Denkena, J.; Colomé-Tatché, M.; Shahryary, Y.; Hazarika, R.; Grimwood, J.; Mamidi, S.; Jenkins, J.; Grabowski, P.P.; Sreedasyam, A.; et al. A Genome Assembly and the Somatic Genetic and Epigenetic Mutation Rate in a Wild Long-Lived Perennial Populus Trichocarpa. Genome Biol. 2020, 21, 259. [Google Scholar] [CrossRef]
- Gallusci, P.; Agius, D.R.; Moschou, P.N.; Dobránszki, J.; Kaiserli, E.; Martinelli, F. Deep inside the Epigenetic Memories of Stressed Plants. Trends Plant Sci. 2023, 28, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Padovan, A.; Keszei, A.; Foley, W.J.; Külheim, C. Differences in Gene Expression within a Striking Phenotypic Mosaic Eucalyptus Tree That Varies in Susceptibility to Herbivory. BMC Plant Biol. 2013, 13, 29. [Google Scholar] [CrossRef] [Green Version]
- Coomes, D.A.; Allen, R.B. Effects of Size, Competition and Altitude on Tree Growth. J. Ecol. 2007, 95, 1084–1097. [Google Scholar] [CrossRef]
- Bigler, C. Trade-Offs between Growth Rate, Tree Size and Lifespan of Mountain Pine (Pinus montana) in the Swiss National Park. PLoS ONE 2016, 11, e0150402. [Google Scholar] [CrossRef] [Green Version]
- Cannon, C.H.; Piovesan, G.; Munné-Bosch, S. Old and Ancient Trees Are Life History Lottery Winners and Vital Evolutionary Resources for Long-Term Adaptive Capacity. Nat. Plants 2022, 8, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Horák, J. Insect Ecology and Veteran Trees. J. Insect Conserv. 2017, 21, 1–5. [Google Scholar] [CrossRef]
- Matwiejuk, A. Lichens of Fruit Trees in the Selected Locations in Podlaskie Voivodeship [North-Eastern Poland]. Ochr. Sr. I Zasobow Nat. 2017, 28, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Michalczuk, J. The Importance of Non-Forest Tree Stand Features for Protection of the Syrian Woodpecker Dendrocopos Syriacus in Agricultural Landscape: A Case Study from South-Eastern Poland. Agrofor. Syst. 2020, 94, 1825–1835. [Google Scholar] [CrossRef]
- Horak, J. Fragmented Habitats of Traditional Fruit Orchards Are Important for Dead Wood-Dependent Beetles Associated with Open Canopy Deciduous Woodlands. Naturwissenschaften 2014, 101, 499–504. [Google Scholar] [CrossRef]
- Žarnovičan, H.; Kollár, J.; Falt’an, V.; Petrovič, F.; Gábor, M. Management and Land Cover Changes in the Western Carpathian Traditional Orchard Landscape in the Period after 1948. Agronomy 2021, 11, 366. [Google Scholar] [CrossRef]
- Anderson, J.T.; Song, B.H. Plant Adaptation to Climate Change—Where Are We? J. Syst. Evol. 2020, 58, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Pasques, O.; Munné-Bosch, S. Physiological Mechanisms Underlying Extreme Longevity in Mountain Pine Trees. Plant Physiol. 2022, 191, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Fang, K.; Li, Y.; Chen, D.; Liu, X.; Dong, Z.; Zhou, F.; Guo, G.; Shi, F.; Xu, C.; et al. Climate, Intrinsic Water-Use Efficiency and Tree Growth over the Past 150 Years in Humid Subtropical China. PLoS ONE 2017, 12, e0172045. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Elevation (m a.s.l.) | Slope (%) | Aspect (%) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Min | Max | Mean | SD | Min | Max | N | NE | E | SE | S | SW | W | NW | |
| Apple | 628 | 163 | 433 | 842 | 18 | 10 | 2 | 29 | - | 14 | - | 14 | 29 | 43 | - | - |
| Cherry | 753 | 157 | 488 | 932 | 14 | 10 | 1 | 36 | - | - | - | 8 | 25 | 8 | 42 | 17 |
| Chestnut | 485 | 281 | 203 | 874 | 12 | 8 | 2 | 25 | - | 9 | 9 | 9 | 36 | 36 | - | - |
| Mulberry | 579 | 180 | 260 | 857 | 23 | 8 | 1 | 34 | - | - | 40 | 20 | 20 | 13 | 7 | - |
| Olive | 354 | 256 | 73 | 807 | 15 | 10 | 3 | 32 | - | - | 36 | 21 | 7 | 7 | 14 | 14 |
| Pear | 698 | 261 | 43 | 1097 | 22 | 9 | 9 | 63 | 5 | 8 | 13 | 15 | 5 | 20 | 25 | 10 |
| Service | 590 | 124 | 479 | 762 | 25 | 4 | 20 | 30 | - | - | 25 | 25 | 25 | 25 | - | - |
| Walnut | 704 | 347 | 325 | 1006 | 26 | 21 | 9 | 49 | - | - | 33 | - | - | - | 33 | 33 |
| Species | Land Use/Land Cover (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 112 | 211 | 222 | 223 | 242/243 | 3111 | 3112 | 3114 | 3115 | 321 | 324 | |
| Apple | - | - | - | - | 86 | - | - | - | - | 14 | - |
| Cherry | - | - | - | - | 50 | 8 | 33 | - | - | - | 8 |
| Chestnut | - | - | - | - | 27 | - | 9 | 9 | 9 | 45 | - |
| Mulberry | 20 | 7 | - | - | 67 | - | 7 | - | - | - | - |
| Olive | 14 | 7 | - | 7 | 71 | - | - | - | - | - | - |
| Pear | 3 | 20 | 3 | - | 63 | - | 8 | - | - | 3 | 3 |
| Service | - | - | - | - | 75 | - | - | - | - | - | 25 |
| Walnut | - | - | - | - | 67 | 33 | - | - | - | - | - |
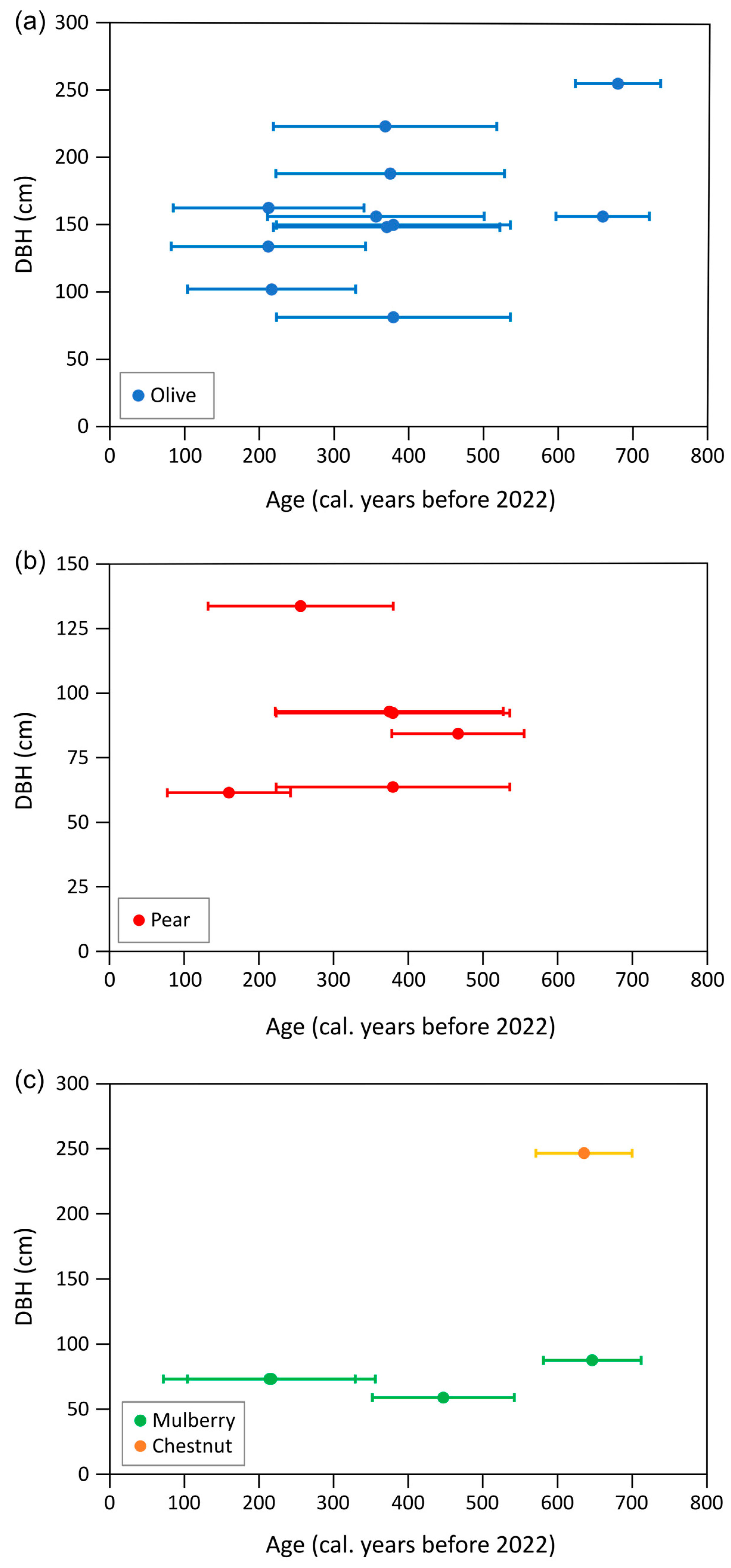
| Species | No. Trees | No. Trees | DBH (cm) | Radiocarbon Age (Years) | Elevation (m a.s.l.) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (Total) | (After 1950) | Mean | Max | Min | Mean | Max | Min | Mean | Max | Min | |
| Chestnut | 2 | 1 * | - | 247 | - | - | 636 | - | - | 865 | - |
| Mulberry | 8 | 4 * | 73 | 88 | 59 | 381 | 646 | 214 | 586 | 824 | 351 |
| Olive | 14 | 3 * | 159 | 255 | 81 | 382 | 680 | 212 | 288 | 776 | 73 |
| Pear | 13 | 6 * | 88 | 134 | 61 | 336 | 467 | 160 | 738 | 1078 | 270 |
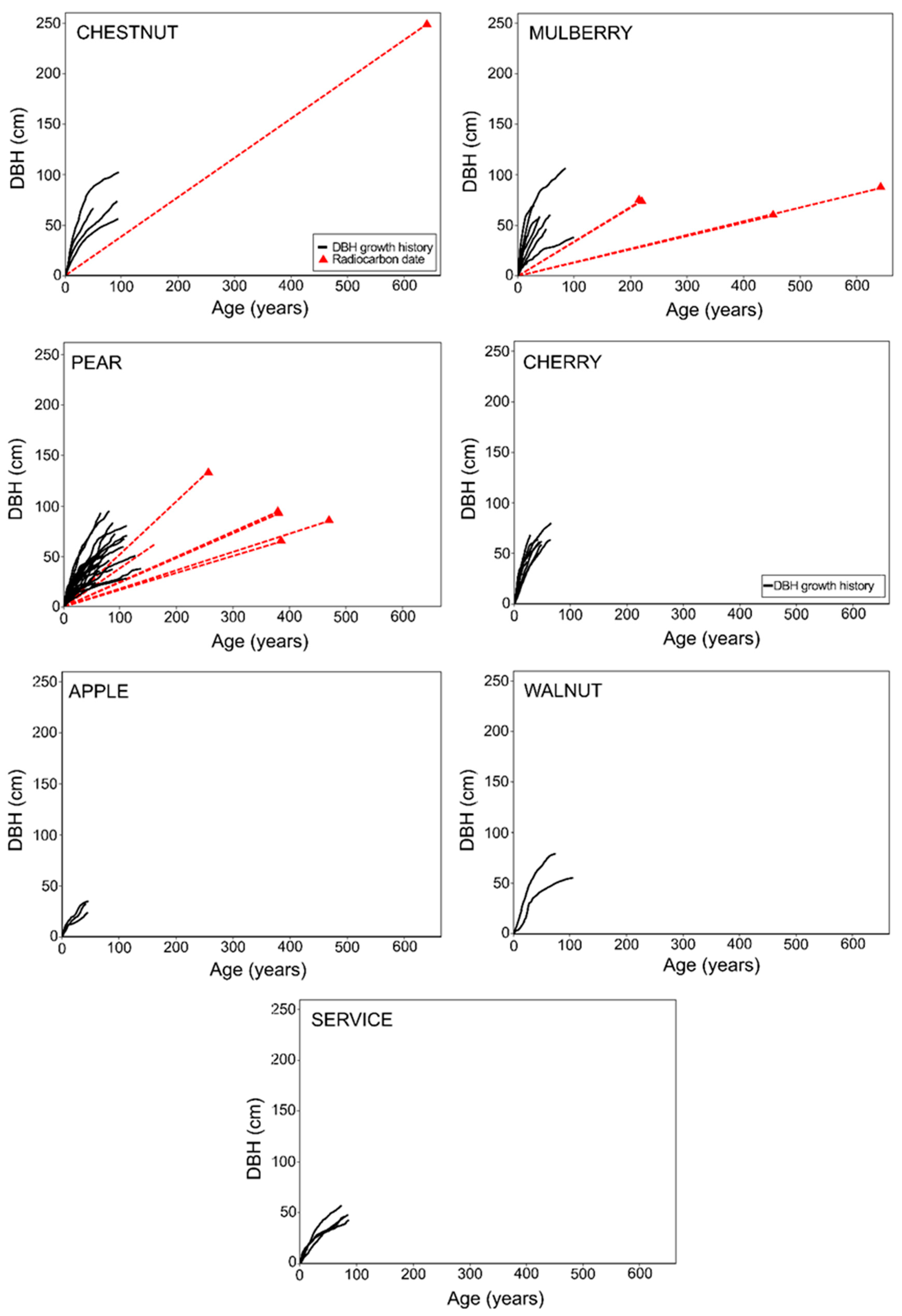
| Species | No. Trees | No. Trees | DBH (cm) | Age (Years) | Elevation (m a.s.l.) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (Total) | (No Pith) | Mean | Max | Min | Mean | Max | Min | Mean | Max | Min | |
| Chestnut | 9 | 5 * | 74 | 102 | 56 | 82 | 94 | 49 | 404 | 618 | 210 |
| Cherry | 12 | 3 * | 63 | 80 | 42 | 50 | 96 | 22 | 803 | 945 | 586 |
| Mulberry | 7 | - | 62 | 107 | 38 | 57 | 100 | 30 | 565 | 853 | 329 |
| Apple | 7 | 4 * | 31 | 35 | 24 | 44 | 42 | 46 | 639 | 820 | 454 |
| Walnut | 3 | 1 * | 68 | 80 | 56 | 90 | 105 | 75 | 563 | 819 | 308 |
| Pear | 27 | 4 * | 51 | 94 | 20 | 80 | 136 | 41 | 707 | 1089 | 25 |
| Service | 4 | - | 49 | 57 | 43 | 81 | 86 | 73 | 482 | 776 | 52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palli, J.; Baliva, M.; Biondi, F.; Calcagnile, L.; Cerbino, D.; D’Elia, M.; Muleo, R.; Schettino, A.; Quarta, G.; Sassone, N.; et al. The Longevity of Fruit Trees in Basilicata (Southern Italy): Implications for Agricultural Biodiversity Conservation. Land 2023, 12, 550. https://doi.org/10.3390/land12030550
Palli J, Baliva M, Biondi F, Calcagnile L, Cerbino D, D’Elia M, Muleo R, Schettino A, Quarta G, Sassone N, et al. The Longevity of Fruit Trees in Basilicata (Southern Italy): Implications for Agricultural Biodiversity Conservation. Land. 2023; 12(3):550. https://doi.org/10.3390/land12030550
Chicago/Turabian StylePalli, Jordan, Michele Baliva, Franco Biondi, Lucio Calcagnile, Domenico Cerbino, Marisa D’Elia, Rosario Muleo, Aldo Schettino, Gianluca Quarta, Nicola Sassone, and et al. 2023. "The Longevity of Fruit Trees in Basilicata (Southern Italy): Implications for Agricultural Biodiversity Conservation" Land 12, no. 3: 550. https://doi.org/10.3390/land12030550